
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
الاستفادة من مجمع بين الافراج عن الإثيلين، حمض 2-Chloroethylphosphonic، كأداة لدراسة استجابة الإثيلين في البكتيريا
In This Article
Summary
The protocols outlined herein facilitate the convenient investigation of bacterial ethylene responses by utilizing 2-chloroethylphosphonic acid (CEPA). Ethylene is produced in situ through the decomposition of CEPA in an aqueous bacterial growth medium, circumventing the requirement for pure ethylene gas.
Abstract
Ethylene (C2H4) is a gaseous phytohormone that is involved in numerous aspects of plant development, playing a dominant role in senescence and fruit ripening. Exogenous ethylene applied during early plant development triggers the triple response phenotype; a shorter and thicker hypocotyl with an exaggerated apical hook. Despite the intimate relationship between plants and bacteria, the effect of exogenous ethylene on bacteria has been greatly overlooked. This is partly due to the difficulty of controlling gaseous ethylene within the laboratory without specialized equipment. 2-Chloroethylphosphonic acid (CEPA) is a compound that decomposes into ethylene, chlorine, and phosphate in a 1:1:1:1 molar ratio when dissolved in an aqueous medium of pH 3.5 or greater. Here we describe the use of CEPA to produce in situ ethylene for the investigation of ethylene response in bacteria using the fruit-associated, cellulose-producing bacterium Komagataeibacter xylinus as a model organism. The protocols described herein include both the verification of ethylene production from CEPA via the Arabidopsis thaliana triple response assay and the effects of exogenous ethylene on K. xylinus cellulose production, pellicle properties and colonial morphology. These protocols can be adapted to examine the effect of ethylene on other microbes using appropriate growth media and phenotype analyses. The use of CEPA provides researchers with a simple and efficient alternative to pure ethylene gas for the routine determination of bacterial ethylene response.
Introduction
الاثيلين الأوليفين (C 2 H 4) اكتشفت لأول مرة كما هرمون مصنع في عام 1901 عندما لوحظ أن الشتلات البازلاء، ونمت في المختبر التي تستخدم مصابيح الغاز والفحم، وعرضت على التشكل الشاذ الذي ينبع كانت (hypocotyls) أقصر، وأكثر سمكا وعازمة جانبية مقارنة الشتلات البازلاء العادية. وصف النمط الظاهري في وقت لاحق 1،2 استجابة الثلاثي. أثبتت الدراسات اللاحقة أن الاثيلين هو هرمون نباتي الحيوي الذي ينظم العمليات التنموية العديدة مثل النمو والاستجابة للضغط النفسي، نضج الثمار والشيخوخة 3. thaliana نبات الأرابيدوبسيس، كائن نموذج للأبحاث بيولوجيا النبات، قد درست جيدا في ما يخص ردها على الإيثيلين. وقد تم عزل المسوخ استجابة العديد من الاثيلين من خلال استغلال النمط الظاهري استجابة الثلاثي لوحظ في الظلام نمت أ. الشتلات thaliana في وجود من الاثيلين 1،4،5. مقدمة السكروز لإنتاج الإثيلين في النباتات هو 1-لحمض minocyclopropane الكربوكسيلية (ACC) 6، ويستخدم عادة خلال فحص استجابة الثلاثي لزيادة انتاج الاثيلين الذاتية التي تؤدي إلى الثلاثي 1،4،5 استجابة النمط الظاهري.
على الرغم من أن استجابة الاثيلين تدرس على نطاق واسع في النباتات، ودراسة سلوكه تأثير الاثيلين دخيلة على البكتيريا إلى حد كبير على الرغم من ارتباط وثيق من البكتيريا مع النباتات. ذكرت إحدى الدراسات أن بعض سلالات الزائفة يمكن البقاء على قيد الحياة باستخدام الاثيلين كمصدر وحيد للكربون والطاقة 7. ومع ذلك، فقد أظهرت اثنين فقط من الدراسات أن البكتيريا تستجيب للالاثيلين. وأظهرت الدراسة الأولى أن سلالات الزائفة الزنجارية، P. المتألقة، P. الكريهة، وP. كانت syringae الكيميائي نحو الاثيلين باستخدام فحص المكونات الاغاروز التي الاغاروز المنصهر كانت مختلطة مع العازلة الكيميائي معايرتها مع غاز الاثيلين النقي 8. ومع ذلك، على حد علمنا، لم تكن هناك أي فورثتقارير إيه باستخدام غاز الاثيلين النقي لتوصيف استجابة الاثيلين البكتيرية، ويرجح ذلك بسبب صعوبة التعامل مع الغازات في المختبر بدون المعدات المتخصصة. أظهر التقرير الثاني للاستجابة الاثيلين البكتيرية التي الاثيلين زاد إنتاج السليلوز البكتيريا والتعبير الجيني أثرت في البكتيريا المصاحبة الفاكهة، Komagataeibacter (سابقا Gluconacetobacter) xylinus 9. في هذه الحالة، ومجمع الإفراج الإثيلين، واستخدم حمض 2-chloroethylphosphonic (CEPA) لإنتاج الإثيلين في الموقع في المتوسط نمو البكتيريا، تجاوز الحاجة للغاز الاثيلين النقي أو المعدات المتخصصة.
CEPA تنتج الاثيلين في نسبة 1: 1 الرحى فوق درجة الحموضة 3.5 10،11 من خلال المحفز الأساس، من الدرجة الأولى رد فعل 12-14. ويرتبط تدهور CEPA بإيجابية مع الرقم الهيدروجيني ودرجة الحرارة 13،14 والنتائج في إنتاج إيثيلإيني، كلوريد والفوسفات. يوفر CEPA الباحثين المهتمين بدراسة ردود البكتيرية إلى الإثيلين مع بديل مناسب لالاثيلين الغازي.
ويتمثل الهدف العام من البروتوكولات التالية هو توفير وسيلة بسيطة وفعالة لدراسة استجابة الاثيلين البكتيرية ويتضمن المصادقة على المستويات ذات الصلة من الناحية الفسيولوجية لانتاج الاثيلين من CEPA التحلل في المتوسط نمو البكتيريا، وتحليل درجة الحموضة الثقافة لضمان CEPA لم تضعف التحلل خلال نمو البكتيريا، وتقييم تأثير الاثيلين على مورفولوجيا البكتيرية والنمط الظاهري. علينا أن نظهر هذه البروتوكولات تستخدم ك. xylinus، ومع ذلك، يمكن تكييف هذه البروتوكولات لدراسة استجابة الاثيلين في غيرها من البكتيريا باستخدام متوسط النمو المناسب وتحليل النمط الظاهري.
Protocol
1. المواد الكيميائية
- إعداد محلول من 500 ملي CEPA (144.49 جم / مول)، والتوصل إلى حل تتألف من 500 ملي كلوريد الصوديوم (58.44 جم / مول) و 500 ملي ناه 2 ص 4 · H 2 O (137.99 جم / مول) في المحمضة (الرقم الهيدروجيني 2.5) ماء نقي للغاية أو 0.1 N حمض الهيدروكلوريك. مزيج باستخدام الدوامة حتى حلول واضحة.
- تمييع متسلسل (10X) الحلول ملي 500 في نفس مذيب للحصول على 5 ملي و 50 ملي الأسهم.
- إعداد محلول 10 ملم من 1 aminocyclopropane حمض الكربوكسيلية (ACC، 101.1 غرام / مول) في مياه نقي للغاية.
- تصفية تعقيم حلول الأسهم وقسامات مخزن في -20 درجة مئوية.
- تصفية تعقيم السيلولوز. مخزن aliquots في 4 درجات مئوية.
2. التحقق من إنتاج الإثيلين من 2 Chloroethylphosphonic حمض التحلل: الثلاثي الفحص الاستجابة
- سطح تعقيم نبات الأرابيدوبسيس thaliana بذور نوع إيكولوجي كولومبيا باستخدام بخار المرحلة الطريقة:
تنبيه: الخطوة التالية تنتج رغاز الكلور ناقصة الأكسدة. إجراء تعقيم البذور في غطاء الدخان.- الحصول على وعاء قابل للغلق عميقة بما فيه الكفاية لاستيعاب كوب زجاجي 250 مل لتعقيم البذور ووضعه في غطاء الدخان.
- إضافة A. thaliana البذور لأنبوب microcentrifuge ووضع أنبوب في رفوف. وضع رف يحتوي على أنبوب مفتوح في وعاء قابل للغلق.
ملاحظة: لا تملأ أنابيب الفردية أكثر من نصف كامل للسماح للغاز الكلور لاختراق البذور أقل. - وضع كوب 250 مل تحتوي على 100 مل من التبييض التجاري في الحاويات اغلاقها باحكام. بعناية إضافة 3 مل من حمض الهيدروكلوريك المركز إلى التبييض وختم على الفور حاوية مع غطاء لها.
تنبيه: بليتش وحمض الهيدروكلوريك رد فعل لإنتاج غاز الكلور السام الذي يعمل على السطح تعقيم البذور. - احتضان بذور في وجود غاز الكلور لمدة 4 ساعة في fumehood.
ملاحظة: مرة التعقيم أطول وتقلل من كفاءة الإنبات. - بعد التعقيم، والسماح للجالغاز hlorine للتنفيس في غطاء الدخان لا يقل عن 1 ساعة ثم ختم أنبوب microcentrifuge. ترك التبييض وحمض الهيدروكلوريك الخليط في fumehood لا يقل عن 24 ساعة قبل رميه.
ملاحظة: قد تكون مخزنة تعقيم البذور في درجة حرارة الغرفة للاستخدام الفوري. - إبقاء البذور في 4 درجة مئوية لمدة التخزين على المدى الطويل. جلب البذور إلى درجة حرارة الغرفة قبل فتح أنبوب microcentrifuge لمنع التكثيف. تخزين البذور في الظلام.
- إعداد لوحات أجار في القطاع 4 أطباق بتري (90 × 15 ملم):
- إعداد 110 مل من متوسط النمو للأ. thaliana الشتلات ما يلي: 1X Murashige وسكوغ (MS) متوسطة القاعدية 15 (4.33 جم / لتر) يحتوي على 1٪ (ث / ت) السكروز و 0.8٪ (ث / ت) أجار. ضبط متوسطة MS لدرجة الحموضة 6 مع هيدروكسيد الصوديوم.
- إعداد 100 مل من متوسط النمو البكتيري للCEPA التحلل: شرام وHestrin (SH) المتوسطة 16 تحتوي على 1.5٪ (ث / ت) أجار. ضبط المتوسطة SH لدرجة الحموضة 7 مع هيدروكسيد الصوديوم.
- تعقيم وسائل الإعلام قبل التعقيم وتمب(ص) في 55 ° C حمام الماء.
- مرة واحدة وقد خفف آغار MS، إضافة إلى 40 مل قارورة معقمة. لإعداد متوسط النمو تحكم إيجابية لأ. thaliana الشتلات، تكملة قسامة 40 مل من مرض التصلب العصبي المتعدد أجار مع 40 ميكرولتر من 10 ملي ACC للحصول على تركيز النهائي من 10 ميكرومتر ACC.
- إضافة 5 مل من المتوسط إلى الأرباع المناسبة للأطباق بتري قطاعية (الشكل 1)، والسماح للأجار ليصلب. إعداد جميع لوحات في ثلاث نسخ.
ملاحظة: لم يتم إضافة CEPA إلى وسط النمو حتى في وقت لاحق في البروتوكول.

الشكل 1: إعداد لوحات أجار يستخدم لفحص استجابة الثلاثي مع CEPA يوضح التخطيطي الأرباع محددة للسيطرة سلبية (A)، مراقبة إيجابية (ب)، واللوحات التجريبية (C). تم تعديل هذا الرقم من Augimeri والشريط 9. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
- تطبق أ. thaliana بذور لضمان الإنبات متزامن:
- إضافة ما يقرب من خمسين ألف بذور thaliana على كل رباعي يحتوي MS أو MS + ACC أجار. وتوزع ضمان بذور بالتساوي لتسهيل إزالة الشتلات والتحليل.
- احتضان لوحات تحتوي على البذور في الظلام في 4 ° مئوية لمدة 3-4 أيام.
- تتحلل CEPA على البكتيريا النمو المتوسطة (SH) ونفذ الثلاثي الاستجابة الفحص:
- بعد التقسيم الطبقي، وتعريض البذور لضوء الفلورسنت لمدة 2 ساعة.
- نشر 10 ميكرولتر من ملي CEPA حل سهم 500 على الأرباع لوحات تجريبية تحتوي على أجار SH (الرقم الهيدروجيني 7؛ الشكل 1) للحصول على تركيز CEPA النهائيمن 1 ملم.
- ختم لوحات مع فيلم المختبرات ويغطى بورق لخلق بيئة مظلمة للبذور.
- تنبت البذور التي يحتضنها لوحات في الظلام في 23 درجة مئوية لمدة 3 أيام مع الجانب آغار أسفل.
ملاحظة: يمكن أن تكون مكدسة لوحات، ولكن يجب وضع ضوابط السلبية على الجزء السفلي منذ الاثيلين هو أخف من الهواء.
- تحليل الثلاثي الاستجابة بيانات الفحص:
- مع ملقط معقم لهب، وإزالة الشتلات واحدة من لوحة المقابلة لكل معاملة وعرض تحت المجهر تشريح أو USB الرقمية. ضمان الجزء السفلي من الشتلات تتماشى وصورة.
- إزالة 30 شتلة من كل رباعي (60 شتلة في تكرار البيولوجي و 180 شتلة لكل معاملة). محاذاة على السطح مع خلفية سوداء وصورة مع الحاكم.
- قياس طول التحتفلقي (ملم) من الشتلات تكرار استخدام يماغيج البرمجيات 17. تعيين نطاق عن طريق النقر والسحب * مستقيم *أداة لتحديد طول 10 ملم. تحديد "جدول تعيين" تحت علامة التبويب "تحليل" وتعيين مسافة معروفة إلى 10. باستخدام أداة * مقسمة *، انقر واسحب لتحديد طول التحتفلقي ثم اضغط M لقياس المسافة. مقارنة وسائل مكررات البيولوجية باستخدام في اتجاه واحد أنوفا مع التجارب المقارنة متعددة لتوكي و. تعتبر الاختلافات كبير إذا ف <0.05.
3. تحليل درجة الحموضة في جميع أنحاء النمو الجرثومي
- تنمو وتحديد Komagataeibacter كاتب xylinus الثقافات في ثلاث نسخ:
- تلقيح مستعمرة واحدة من ك. xylinus إلى 5 مل من المتوسط SH (الرقم الهيدروجيني 5) تستكمل مع 0.2٪ (ت / ت) السيلولوز فلتر تعقيم. احتضان الثقافات في 30 درجة مئوية مع الإثارة في 150 دورة في الدقيقة حتى لOD يتم التوصل إلى 600 من 0،3-0،4 (حوالي 72 ساعة).
- الثقافات الحصاد بداية بواسطة الطرد المركزي (2000 x ج، 4 درجة مئوية و 10 دقيقة). مع 5 مل العقيمة 0.85٪ (ث / ت) NACل حل، وغسل الخلايا مرتين و resuspend بيليه الخلية. تبقي الخلايا على الجليد.
- تحديد الخلايا باستخدام غرفة عد بيتروف-Hausser.
- تطعيم الثقافات لتحليل درجة الحموضة:
- إضافة 150 مل من مرق SH (الرقم الهيدروجيني 7) تستكمل مع 0.2٪ (ت / ت) السيلولوز فلتر تعقيم إلى 500 اثني عشر مل القوارير ذات الأغطية احباط. تطعيم قوارير مع ك. بداية الثقافات xylinus بتركيز 10 5 خلية / مل. باستخدام بداية الثقافات الثلاث، إعداد ثلاث مكررات البيولوجية لكل معاملة.
- الثقافات الملحق مع 300 ميكرولتر من 5، 50، أو 500 الحلول الأسهم ملم CEPA للحصول على تركيزات CEPA النهائية من 0.01، 0.1، و 1.0 ملم على التوالي. تكملة الثقافات ضابطة مع 300 ميكرولتر من المذيبات المستخدمة لإذابة CEPA.
- ختم القوارير عن طريق تسجيل بإحكام الأغطية احباط لقوارير واحتضان الثقافات لمدة 14 يوما حتى 30 درجة مئوية مع الإثارة في 150 دورة في الدقيقة.
- كل يوم، asepticaLLY إزالة 5 مل عينات من كل قارورة وبيليه الخلايا بواسطة الطرد المركزي (2000 x ج، 4 درجة مئوية و 10 دقيقة). نقل supernatants إلى أنبوب نظيفة وقياس درجة الحموضة من كل تكرار البيولوجي باستخدام مقياس درجة الحموضة.
- تحليل البيانات في الوقت بالطبع عن طريق الرسوم البيانية يعني الرقم الهيدروجيني للمكررات البيولوجية.
ملاحظة: من المهم أن الرقم الهيدروجيني الثقافة لا تنخفض الى اقل من 3.5. الرقم الهيدروجيني الثقافة من 5 يقلل بشكل ملحوظ الافراج الاثيلين من CEPA، ودرجة حموضة أقل من 3.5 يمنع تماما الإفراج الاثيلين. - للسيطرة على مستويات الكلور والفوسفات، إجراء تجربة مماثلة باستخدام 0.01، 0.1، و 1.0 ملم من 4 حل كلوريد الصوديوم-ناه 2 ص باستخدام 5، 50، و 500 أسهم ملم على التوالي.
4. مستعمرة الصرف
- تنمو ك. الثقافات xylinus كاتب في ثلاث نسخ:
- تلقيح مستعمرة واحدة من ك. xylinus إلى 5 مل من المتوسط SH (الرقم الهيدروجيني 5) تستكمل مع 0.2٪ (ت / ت) السيلولوز فلتر تعقيم. Incubaالثقافات الشركة المصرية للاتصالات عند 30 درجة مئوية مع الإثارة في 150 دورة في الدقيقة حتى على OD 600 من 0،3-0،4 يتم التوصل إلى (حوالي 72 ساعة).
- الثقافات الحصاد بداية بواسطة الطرد المركزي (2000 x ج، 4 درجة مئوية و 10 دقيقة). مع 5 مل من محلول ملحي، وغسل الخلايا ثم resuspend الكرية الخلية. تبقي الخلايا على الجليد.
- إعداد 24 أجار لوحات تحتوي على 25 مل من SH (الرقم الهيدروجيني 7) المتوسطة مع 1.5٪ (ث / ت) أجار.
- مرة واحدة وقد عززت أجار، نشر 50 ميكرولتر من 5، 50، و 500 الحلول الأسهم ملم CEPA على أجار للحصول على تركيزات CEPA النهائية من 0.01، 0.1، و 1.0 ملم على التوالي. إعداد لوحات تحكم غير المعالجة والمذيبات، والتي تتكون من أي تعديل ونشر 50 ميكرولتر من المذيبات المستخدمة لإذابة مركبات الاختبار.
- لوحات متتالية للمستعمرات معزولة مع حلقة كاملة (~ 5 ميكرولتر) من ك. xylinus ثقافة المبتدئين وختم لهم مع فيلم البارافين. تطعيم جميع لوحات في ثلاث نسخ. احتضان لوحات لمدة خمسة أيام في 30 درجة مئوية.
- الفت وحدة التدفق الضوئيالمستعمرات ograph في 20X التكبير باستخدام مجهر USB الرقمية ونوعيا تقييم الاستعماري التشكل والسليلوز الإنتاج في وسط صلب.
يبدو السليلوز باعتباره مادة ضبابية على هامش مستعمرة: مذكرة. - للسيطرة على مستويات الكلور والفوسفات، إجراء تجربة مماثلة باستخدام 0.01، 0.1، و 1.0 ملم من 4 حل كلوريد الصوديوم-ناه 2 ص باستخدام 5، 50، و 500 أسهم ملم على التوالي.
5. جليدة فحوصات
- تنمو وتحديد ك. الثقافات xylinus كاتب في ثلاث نسخ:
- في ثلاث نسخ، تلقيح مستعمرة واحدة من ك. xylinus إلى 5 مل من المتوسط SH (الرقم الهيدروجيني 5) تستكمل مع 0.2٪ (ت / ت) السيلولوز فلتر تعقيم. احتضان الثقافات في 30 درجة مئوية مع الإثارة في 150 دورة في الدقيقة حتى لOD يتم التوصل إلى 600 من 0،3-0،4 (حوالي 72 ساعة).
- الثقافات الحصاد بداية بواسطة الطرد المركزي (2000 x ج، 4 درجة مئوية و 10 دقيقة). مع 5 مل من محلول ملحي، وغسل رانه خلايا مرتين و resuspend بيليه الخلية. تبقي الخلايا على الجليد.
- تحديد الخلايا باستخدام غرفة عد بيتروف-Hausser.
- إعداد ماستر خلطات لتلقيح لوحات 24-جيدا:
- تكملة 60 مل من المتوسط SH (الرقم الهيدروجيني 7) مع 120 ميكرولتر من 5، 50، و 500 أسهم ملي CEPA الحصول على تركيزات CEPA النهائية من 0.01، 0.1، و 1.0 ملم على التوالي. تكملة 60 مل أخرى متوسطة SH (الرقم الهيدروجيني 7) مع 120 ميكرولتر من المذيبات المستخدمة لإذابة CEPA. دوامة لخلط.
- تقسيم كل 60 مل من CEPA-تحتوي على المتوسط إلى أربعة 14 مل قسامات. تطعيم ثلاثة من aliquots مل 14 مع مكررات البيولوجية للثقافة المبتدئين بتركيز 10 5 خلية / مل. إبقاء الأنابيب مع الخلايا على الجليد لمنع إنتاج السليلوز.
ملاحظة: سيتم استخدام قسامة 14 مل المتبقية لآبار المراقبة معقمة.
- تطعيم لوحات 24-جيدا:
- اختبار كل معاملة في لوحة خاصة بها مع ثلاثة صفوف من مكررات البيولوجية وصف من الضوابط معقمة (الشكل 2).

وaliquoted تدفق البياني يوضح البروتوكول المستخدم لفحص جليدة وتحليل تستكمل CEPA المخزن الرقم الهيدروجيني 7 المتوسطة SH (60 مل) لمدة ثلاث التطعيمات تكرار البيولوجي منفصلة والسيطرة العقيمة (14 مل لكل منهما): الشكل 2. ثم يتم aliquoted هذه الثقافات إلى ستة مكررات التقنية (2 مل) في 24 لوحة جيدا وبعد ذلك اغلقت مع فيلم البارافين. بعد مرور فترة الحضانة لمدة 7 أيام في 30 درجة مئوية، ويتم حصاد pellicles وتتميز تحديد الوزن الرطب، سمك، الوزن الجاف، والتبلور التي كتبها FT-IR. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم
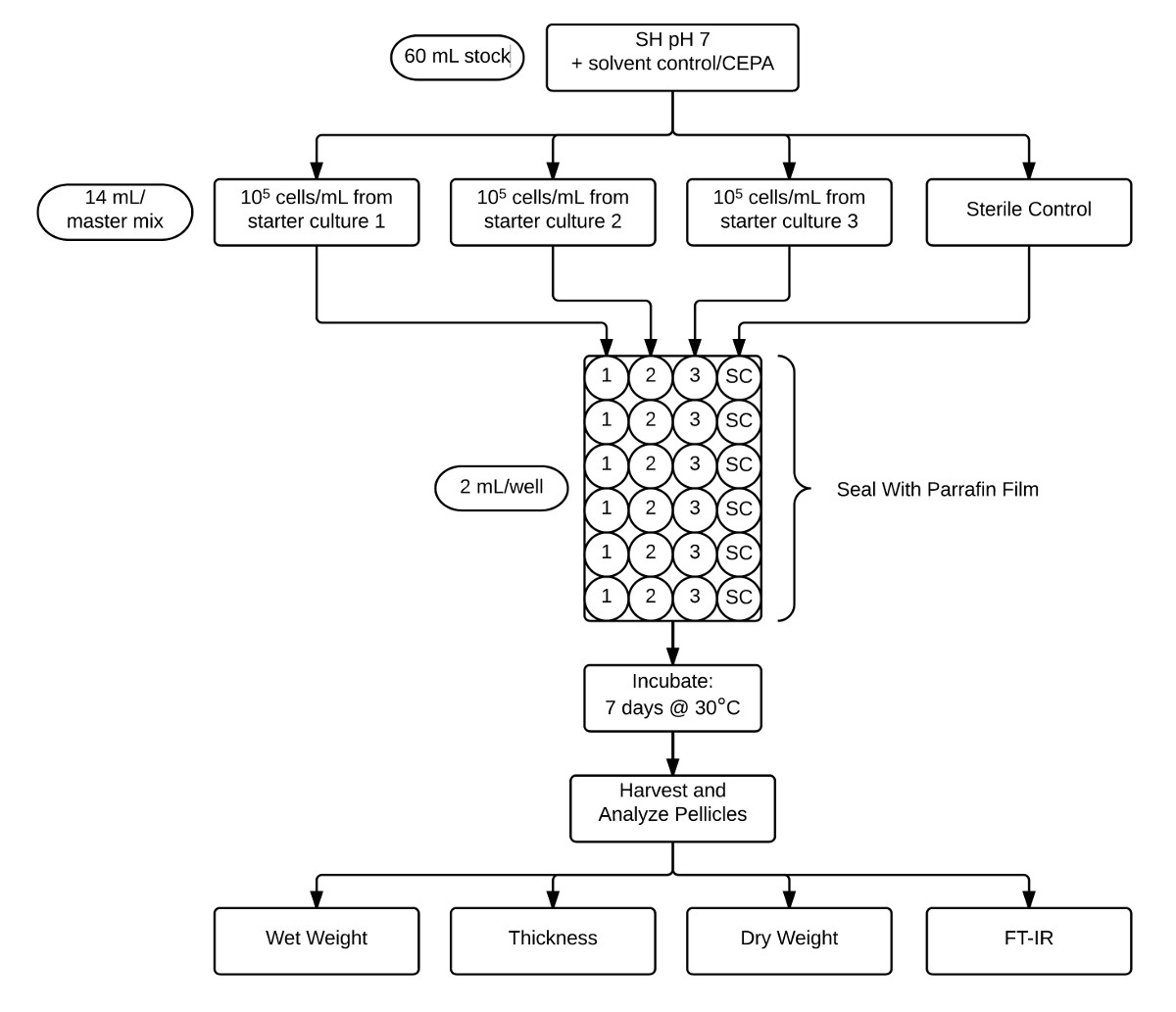{kind=link}
- باستخدام مزيج الرئيسي 14 مل، إضافة 2 مل في مجموعة شرق افريقياح ستة آبار العقيمة 24 لوحة جيدا. استكمال لمكررات البيولوجية ثلاثة والسيطرة العقيمة (الشكل 2). كرر لكل معاملة.
- لوحات ختم مع فيلم البارافين واحتضان ثابت لمدة 7 أيام عند 30 درجة مئوية.
- الحصاد وقياس جليدة ويت الوزن، سمك، الوزن الجاف (السليلوز الغلة) والتبلور (الشكل 2):
- خفض جانب واحد من جليدة لرفع حافة جليدة المعارضة وإزالة pellicles الفردية مع ملقط. مع الحفاظ على قبضة، ووضعها على منشفة ورقية جديدة لمدة 3 ثانية لإزالة المتوسطة الزائدة قبل وزنها لتحديد أوزانها الرطب.
- محاذاة pellicles المتاخمة للحاكم وصورة من الجانب باستخدام كاميرا رقمية عالية الدقة.
- باستخدام يماغيج البرمجيات 17، وقياس سماكة غشاء رقيق على الكتف الأيسر والكتف الأيمن ووسط كل جليدة. متوسط جميع مكررات التقنية لكل replicat البيولوجيه.
- نقل فردي pellicles في الآبار من لوحة 6 جيدا. علاج pellicles مع 12 مل من 0.1 هيدروكسيد الصوديوم N 80 درجة مئوية لمدة 20 دقيقة لليز الخلايا.
- إزالة هيدروكسيد الصوديوم وتحييد pellicles عن طريق الغسيل بالماء فائقة نقية لمدة 24 ساعة مع الإثارة. تغيير الماء كل 6 ساعات.
ملاحظة: يجب أن تكون بيضاء Pellicles عند الانتهاء من خطوة الغسيل. - مكان pellicles على الحصير السيليكون وجافة عند 50 درجة مئوية لمدة 48 ساعة على وزن ثابت. الجاف مرة واحدة، وترفع من الحصير وقياس الأوزان جليدة على نطاق تحليلي لتحديد العائد السليلوز البكتيريا.
- تحليل جليدة التبلور باستخدام فورييه تحويل مطياف الأشعة تحت الحمراء (FT-IR) باستخدام 32 المسح الضوئي وقرار من 4 سم -1 في حدود 4000 إلى 650 سم -1. حساب مؤشر التبلور، CI (الأشعة تحت الحمراء)، وذلك باستخدام 1437 / أ 895. نسبة الامتصاص من "الفرقة البلورية" و "الفرقة غير متبلور" كما وصفها سابقا 18.
- للسيطرة على مستويات الكلور والفوسفات، إجراء تجربة مماثلة باستخدام 0.01، 0.1، و 1.0 ملم من 4 حل كلوريد الصوديوم-ناه 2 ص باستخدام 5، 50، و 500 أسهم ملم على التوالي.
- تحليل البيانات جليدة:
- حساب جليدة الماء عن طريق تحديد الفرق بين جليدة الوزن الرطب والوزن الجاف.
- متوسط قيم كل مكررات التقنية للحصول على قيمة واحدة لكل تكرار البيولوجي للتحليل الإحصائي. مقارنة العلاجات باستخدام في اتجاه واحد أنوفا مع التجارب المقارنة متعددة توكي و. فروق ذات دلالة إذا ف <0.05.
- تطبيع البيانات مثل في المئة من الضوابط غير المعالجة ورسم وسائل مكررات البيولوجية.
النتائج
ويرد الإعداد لوحة التخطيطي للتحقق من تحرير الاثيلين من CEPA في المتوسط SH (الرقم الهيدروجيني 7) من خلال فحص استجابة الثلاثي في الشكل 1A - C. ويرد-الرسم البياني يوضح بروتوكول جليدة في الشكل 2. الظلام نمت أ. thaliana الشتلات يحمل ا...
Discussion
الأساليب المذكورة هنا تلخص في الإنتاج الموقعي من الاثيلين من CEPA لدراسة استجابة الاثيلين البكتيرية باستخدام كائن نموذج، K. xylinus. هذه الطريقة مفيدة جدا كما الاثيلين يمكن أن تنتج من خلال استكمال أي وسط مائي يحتوي على الأس الهيدروجيني أكبر من 3.5 10،11 م?...
Disclosures
The authors have nothing to disclose.
Acknowledgements
The authors thank Dr. Dario Bonetta for providing Arabidopsis thaliana seeds and for technical assistance in regards to the triple response assay, as well as Simone Quaranta for help with FT-IR. This work was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC-DG) to JLS, an Ontario Graduate Scholarship (OGS) to RVA, and a Queen Elizabeth II Graduate Scholarship in Science and Technology (QEII-GSST) to AJV.
Materials
| Name | Company | Catalog Number | Comments |
| 1-aminocyclopropane carboxylic acid (ACC) | Sigma | A3903 | Biosynthetic precursor of ethylene in plants |
| 4-sector Petri dish | Phoenix Biomedical | CA73370-022 | For testing triple response |
| Agar | BioShop | AGR001.1 | To solidify medium |
| Canon Rebel T1i DLSR camera | Canon | 3818B004 | For pictures of pellicles |
| Cellulase from Trichoderma reesei ATCC 26921 | Sigma | C2730 | Aqueous solution |
| Citric acid | BioShop | CIT002.500 | For SH medium |
| Commercial bleach | Life Brand | 57800861874 | Bleach for seed sterilization |
| Concentrated HCl | BioShop | HCL666.500 | Hydrochloric acid for pH adjustment |
| Digital USB microscope | Plugable | N/A | For pictures of colonies |
| Ethephon (≥96%; 2-chloroethylphosphonic acid) | Sigma | C0143 | Ethylene-releasing compound |
| Glucose | BioBasic | GB0219 | For SH medium |
| Komagataeibacter xylinus ATCC 53582 | ATCC | 53582 | Bacterial cellulose-producing alphaproteobacterium |
| Microcentrifuge tube | LifeGene | LMCT1.7B | 1.7 ml microcentrifuge tube |
| Murashige and Skoog (MS) basal medium | Sigma | M5519 | Arabidopsis thaliana growth medium |
| Na2HPO4·7H2O | BioShop | SPD579.500 | Sodium phosphate, dibasic heptahydrate for SH medium |
| NaCl | BioBasic | SOD001.1 | Sodium chloride for saline and control solution |
| NaH2PO4·H2O | BioShop | SPM306.500 | Sodium phosphate, monobasic monohydrate for control solution |
| NaOH | BioShop | SHY700.500 | Sodium hydroxide for pH adjustment |
| Paraffin film | Parafilm | PM996 | For sealing plates and flasks |
| Peptone (bacteriological) | BioShop | PEP403.1 | For SH medium |
| Petroff-Hausser counting chamber | Hausser scientific | 3900 | Bacterial cell counting chamber |
| Polyethersulfone sterilization filter 0.2 µm | VWR | 28145-501 | For sterilizing cellulase |
| Sucrose | BioShop | SUC600.1 | Sucrose for MS medium |
| Yeast extract | BioBasic | G0961 | For SH medium |
References
- Guzmán, P., Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell. 2 (6), 513-523 (1990).
- Bakshi, A., Shemansky, J. M., Chang, C., Binder, B. M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 34 (4), 809-827 (2015).
- Schaller, G. E. Ethylene and the regulation of plant development. BMC Biol. 10 (1), (2012).
- Hua, J., Sakai, H., et al. EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis. Plant Cell. 10 (8), 1321-1332 (1998).
- Bleecker, A. B., Estelle, M. A., Somerville, C., Kende, H. Insensitivity to ethylene conferred by a dominant Mutation in Arabidopsis thaliana. Science. 241 (4869), 1086-1089 (1988).
- Hamilton, A. J., Bouzayen, M., Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. 88 (16), 7434-7437 (1991).
- Kim, J. Assessment of ethylene removal with Pseudomonas strains. J. Hazard. Mater. 131 (3), 131-136 (2006).
- Kim, H. E., Shitashiro, M., Kuroda, A., Takiguchi, N., Kato, J. Ethylene chemotaxis in Pseudomonas aeruginosa and other Pseudomonas species. Microbes Environ. 22 (2), 186-189 (2007).
- Augimeri, R. V., Strap, J. L. The phytohormone ethylene enhances bacterial cellulose production, regulates CRP/FNRKx transcription and causes differential gene expression within the cellulose synthesis operon of Komagataeibacter (Gluconacetobacter) xylinus ATCC 53582. Front. Microbiol. 6, 1459 (2015).
- Zhang, W., Wen, C. K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 48 (1), 45-53 (2010).
- Zhang, W., Hu, W., Wen, C. K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 5 (4), 453-457 (2010).
- Warner, H. L., Leopold, A. C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44 (1), 156-158 (1969).
- Biddle, E., Kerfoot, D. G. S., Kho, Y. H., Russell, K. E. Kinetic studies of the thermal decomposition of 2-chloroethylphosphonic acid in aqueous solution. Plant Physiol. 58 (5), 700-702 (1976).
- Klein, I., Lavee, S., Ben-Tal, Y. Effect of water vapor pressure on the thermal decomposition of 2-chloroethylphosphonic acid. Plant Physiol. 63 (3), 474-477 (1979).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15 (3), 473-497 (1962).
- Schramm, M., Hestrin, S. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum. J. Gen. Microbiol. 11 (1), 123-129 (1954).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671-675 (2012).
- Ciolacu, D., Ciolacu, F., Popa, V. I. Amorphous cellulose-structure and characterization. Cellul. Chem. Technol. 45 (1), 13-21 (2011).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved