
Method Article
قياس ارتباط النوكليوتيدات ببروتينات الغشاء الوظيفية السليمة في الوقت الفعلي
In This Article
Summary
يقدم هذا البروتوكول طريقة لقياس ارتباط نيوكليوتيدات الأدينين بالمستقبلات في الوقت الفعلي في بيئة خلوية. يتم قياس الارتباط على أنه نقل طاقة الرنين Förster (FRET) بين مشتقات نيوكليوتيدات ثلاثي نيتروفينيل والبروتين المسمى بحمض أميني فلوري غير قانوني.
Abstract
لقد طورنا طريقة لقياس ارتباط نيوكليوتيدات الأدينين بمستقبلات غشائية سليمة ووظيفية في بيئة خلوية أو غشائية. تجمع هذه الطريقة بين التعبير عن البروتينات الموسومة بالحمض الأميني الفلوري غير المتعارف عليه ANAP ، والحنق بين مشتقات النوكليوتيدات ANAP والفلورية (ثلاثي النيتروفينيل). نقدم أمثلة على ارتباط النوكليوتيدات بقنوات أيون KATP الموسومة ب ANAP المقاسة في أغشية البلازما غير المسقوفة وبقع الغشاء المستأصلة من الداخل إلى الخارج تحت مشبك الجهد. هذا الأخير يسمح بالقياسات المتزامنة لربط الليجند وتيار القناة ، وهي قراءة مباشرة لوظيفة البروتين. تتم مناقشة معالجة البيانات وتحليلها على نطاق واسع ، إلى جانب المزالق والمصنوعات اليدوية المحتملة. توفر هذه الطريقة رؤى ميكانيكية غنية حول البوابة المعتمدة على الرباط لقنوات KATP ويمكن تكييفها بسهولة لدراسة البروتينات الأخرى التي ينظمها النيوكليوتيدات أو أي مستقبل يمكن تحديد ليجند فلوري مناسب له.
Introduction
يتم تنظيم عدة فئات مهمة من البروتين مباشرة عن طريق ربط الرباط. تتراوح هذه من الإنزيمات القابلة للذوبان إلى البروتينات المضمنة في الغشاء بما في ذلك مستقبلات التيروزين كينازات والمستقبلات المقترنة بالبروتين G (GPCRs) والقنوات الأيونية. تمثل GPCRs والقنوات ~ 34٪ و ~ 15٪ من جميع أهداف الأدوية الحالية ،على التوالي 1,2. لذلك ، هناك اهتمام كيميائي حيوي كبير ، بالإضافة إلى اهتمام طبي بتطوير الأساليب التي توفر رؤى ميكانيكية في تفاعلات مستقبلات الرباط. تتطلب الطرق التقليدية لقياس ارتباط الربيطة ، بما في ذلك وضع العلامات على التقارب الضوئي ودراسات ربط الليجند الإشعاعي ، كميات كبيرة من البروتين المنقى جزئيا ويتم إجراؤها عادة في ظل ظروف غير فسيولوجية ومقاييس زمنية. تتطلب الطريقة المثالية كميات صغيرة فقط من البروتين ، ويمكن إجراؤها على بروتينات سليمة يتم التعبير عنها في بيئة خلوية أو غشائية ، ويمكن مراقبتها في الوقت الفعلي ، وستكون متوافقة مع القراءات المباشرة لوظيفة البروتين.
نقل طاقة الرنين Förster (FRET) هي طريقة تكتشف القرب بين جزيئين موسومين بالفلورسنت3. يحدث الحنق عندما ينقل فلوروفور متبرع متحمس الطاقة بطريقة غير إشعاعية إلى جزيء متقبل (عادة فلوروفور آخر). يؤدي نقل الطاقة إلى إخماد انبعاث مضان المتبرع وتوعية انبعاث المستقبل (إذا كان المستقبل عبارة عن فلوروفور). تعتمد كفاءة النقلعلى قوة 6 للمسافة بين المتبرع والمستقبل. علاوة على ذلك ، يجب أن يكون المتبرع والمتقبل على مقربة (عادة أقل من 10 نانومتر) حتى يحدث FRET. على هذا النحو ، يمكن استغلال FRET لقياس الارتباط المباشر بين مستقبلات البروتين الموسومة بالفلورسنت والرباط الفلوري.
يتم تنظيم أو تنشيط العديد من البروتينات المختلفة عن طريق ربط نيوكليوتيدات الأدينين داخل الخلايا أو خارج الخلية (ATP ، ADP ، AMP ، cAMP). تتطلب العديد من بروتينات النقل التحلل المائي ATP لدورة تفاعلها ، بما في ذلك ناقلات الكاسيت المرتبطة ب ATP و ATPases من النوع P مثل مضخة Na + / K + 4,5 . قنوات K + (K ATP) الحساسة لATP ، ومنظم التوصيل عبر الغشاء للتليف الكيسي (CFTR) ، والقنوات المنظمة للنيوكليوتيدات الحلقية كلها قنوات أيونية مسورة بربط نيوكليوتيدات الأدينين داخل الخلايا ، مما يجعلها حساسة بشكل رائع للتغيرات في التمثيل الغذائي الخلوي ونقل الإشارة6،7،8. تستجيب مستقبلات P2X و P2Y Purinergic للتغيرات في ATP خارج الخلية ، والتي يمكن إطلاقها كناقل عصبي أو نتيجة لتلف الأنسجة9. لقد طورنا مقايسة قائمة على FRET لقياس ارتباط نيوكليوتيدات الأدينين ببروتينات الغشاء في الوقت الفعلي. لقد طبقنا هذه الطريقة سابقا لدراسة ارتباط النوكليوتيدات بقنوات KATP 10,11.
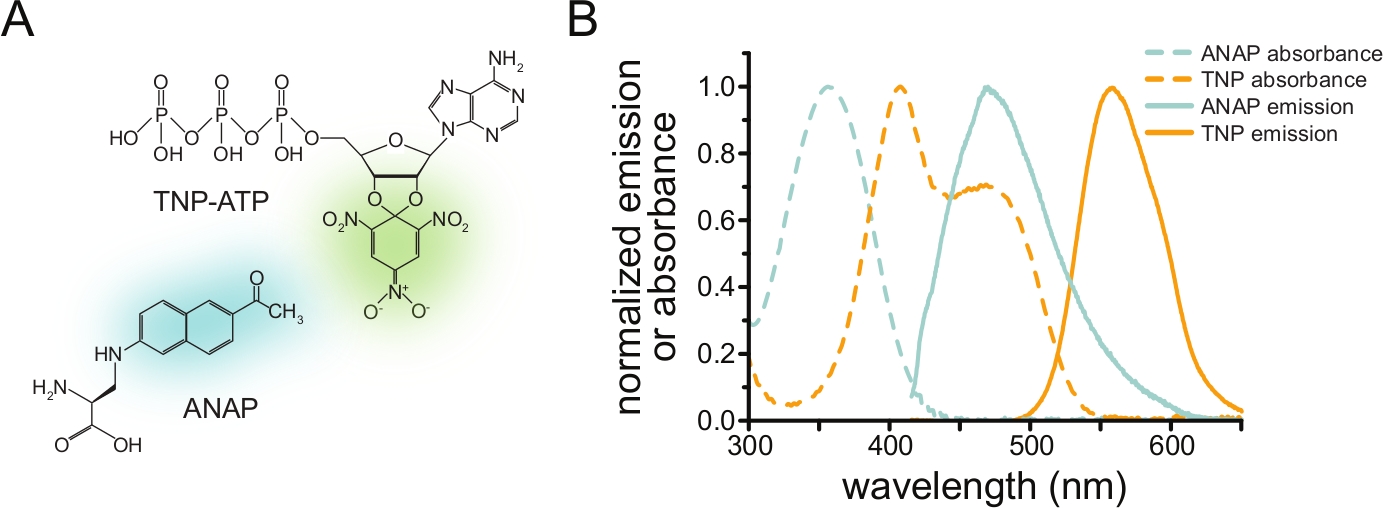
لقياس ارتباط النوكليوتيدات عبر FRET ، يجب أولا تمييز البروتين محل الاهتمام بالفلوروفور. يجب إدخال علامة الفلورسنت في الموقع على وجه التحديد في البروتين محل الاهتمام بحيث يكون قريبا بدرجة كافية من موقع ربط الليجند لحدوث FRET ، مع توخي الحذر بشكل خاص لضمان عدم تأثير العلامة على الهيكل العام للبروتين ووظيفته. لتحقيق ذلك ، نستخدم تقنية طورها Chatterjee et al. ، باستخدام قمع كودون إيقاف الكهرمان لإدخال حمض أميني فلوري غير قانوني (l-3- (6-acetylnaphthalen-2-ylamino) -2-aminopropionic; ANAP) في الموقع المطلوب12. نقيس ارتباط النوكليوتيدات على أنه FRET بين البروتين المسمى ANAP ومشتقات النوكليوتيدات الفلورية ، ثلاثي نيتروفينيل (TNP) (الشكل 1 أ). يتداخل طيف الانبعاث ل ANAP مع طيف امتصاص نيوكليوتيدات TNP ، وهي حالة ضرورية لحدوث FRET (الشكل 1B). نوضح هنا نوعين مختلفين من تجارب الربط. في الأول ، يتم قياس ارتباط النوكليوتيدات بالجانب داخل الخلايا من قنوات KATP التي تحمل علامة ANAP في الخلايا التي تم تسقيفها عن طريق صوتنة تاركة شظايا ملتصقة من غشاء البلازما على انزلاق غطاء زجاجي10،11،13،14.
في الطريقة الثانية ، يتم قياس ارتباط النوكليوتيدات بقنوات KATP التي تحمل علامة ANAP في رقعة غشائية تحت مشبك الجهد ، مما يسمح بالقياس المتزامن للتيارات الأيونية والتألق. من خلال الجمع بين هذين النهجين التجريبيين ، يمكن ربط التغييرات في الربط ارتباطا مباشرا بالتغيرات في وظيفة القناة11. تتم مناقشة النتائج النموذجية والمزالق المحتملة وتحليل البيانات.
Protocol
1. إعداد زلات الغطاء
ملاحظة: يجب أن تتم هذه الخطوات في غطاء زراعة الأنسجة المعقمة. يتم إعطاء كميات لإعداد 10 أطباق.
- ضع عشرة عبوات زجاجية من البورسليكات المعقم مقاس 30 مم بشكل فردي في عشرة أطباق معقمة غير معالجة مقاس 35 مم واشطفها مرة واحدة ب 2 مل من الماء المقطر المعقم.
- تمييع 1 مل من 0.1٪ وزن / في محلول بولي-L-ليسين في ماء مقطر معقم إلى حجم إجمالي قدره 10 مل (التركيز النهائي 0.01٪ وزن / حجم). تخلط جيدا ، ثم ماصة 1 مل على كل غطاء زلة واحتضانها في درجة حرارة الغرفة لمدة 20 دقيقة.
- نضح بولي-L-ليسين واغسل كل غطاء مرتين مع ما لا يقل عن 2 مل من الماء المقطر المعقم. اتركيه حتى يجف تماما ، أي 3 ساعات على الأقل.
2. بذر خلايا HEK-293T
ملاحظة: يجب أن تتم هذه الخطوات في غطاء زراعة الأنسجة. تم اختيار خلايا HEK-293T لخلفيتها الحالية المنخفضة وسهولة النمو في الثقافة. يمكن تكييف هذا البروتوكول مع أنواع الخلايا الأخرى.
- اشطف قارورة T75 متقاربة بنسبة 80-90٪ من خلايا HEK-293T مرة واحدة باستخدام 12 مل من محلول ملحي مخزن للفوسفات (PBS) قبل احتضانها ب 2 مل من التربسين لمدة 2-5 دقائق ، أو حتى تنفصل الخلايا تماما وتنفصل تماما تقريبا.
- أعد تعليق الخلايا بإضافة 10 مل من وسط النسر المعدل من Dulbecco (DMEM) المكمل بمصل بقري جنيني بنسبة 10٪ و 100 وحدة / مل من البنسلين و 100 ميكروغرام / مل من الستربتومايسين. ماصة بلطف على الجزء السفلي من القارورة لتفتيت كتل الخلايا المتبقية.
- أضف 2 مل من DMEM المكمل إلى العدد المطلوب من الأطباق مقاس 35 مم التي تحتوي على زلات غطاء مطلية. أضف 100 ميكرولتر من الخلايا المعلقة إلى كل طبق. احتضان بين عشية وضحاها في 37 درجة مئوية.
3. النقل
ملاحظة: يجب أن تتم هذه الخطوات في غطاء زراعة الأنسجة. يتم إعطاء كميات لنقل 10 أطباق. لدمج ANAP الخاص بالموقع ، يجب استبدال كودون الحمض النووي في الموضع المخصص لوضع العلامات بكودون توقف الكهرماني (TAG). يتم نقل هذا البناء مع اثنين من البلازميدات: pANAP و peRF1-E55D12,15. يشفر pANAP عدة نسخ من زوج سينثيتاز tRNA / tRNA الخاص ب ANAP. في وجود ANAP ، ينتج عن نقل هذا البلازميد tRNA مشحون ب ANAP الذي يتعرف على كودون توقف الكهرماني. يشفر peRF1-E55D عامل إطلاق الريبوسوم السلبي السائد الذي يزيد من إنتاجية البروتين الكامل الطول الموسوم ب ANAP.
- قم بإعداد أنبوب سعة 1.5 مل مع 10 ميكروغرام pANAP ، 10 ميكروغرام peRF1-E55D ، والحمض النووي للبناء المخصص لوضع العلامات باستخدام ANAP. يصل الحجم النهائي إلى 500 ميكرولتر مع DMEM غير المكمل.
- في أنبوب منفصل ، قم بإعداد 3 ميكرولتر من كاشف النقل القائم على الدهون (انظر جدول المواد) لكل 1 ميكروغرام من الحمض النووي والوصول إلى الحجم النهائي البالغ 500 ميكرولتر مع DMEM غير المكمل.
- الجمع بين الحمض النووي ومخاليط كاشف نقل في أنبوب واحد واحتضان لمدة 20 دقيقة في درجة حرارة الغرفة.
- أضف 400 ميكرولتر من مخزون 1 mM ANAP (ملح ثلاثي فلورو أسيتات في 30 mM NaOH) إلى 20 مل مكمل DMEM لتركيز نهائي قدره 20 ميكرومتر ANAP. استبدل الوسائط القديمة من الخلايا المطلية ب 2 مل من الوسائط المحتوية على ANAP لكل طبق.
- ماصة 10٪ من مزيج نقل الحمض النووي على كل طبق. احتضان في 33 درجة مئوية لمدة 2-4 أيام قبل التجارب. الحضانة عند 33 درجة مئوية تبطئ انقسام الخلايا وتزيد من إنتاج البروتين لكل خلية16.
4. تجارب الأغشية غير المسقوفة
- استخدم زوجا من الملقط لكسر زلة الغطاء مع الخلايا المنقولة إلى أجزاء أصغر.
- اتبع أحد الإجراءات أدناه لفتح الخلايا.
- في حالة استخدام زلات غطاء مطلية مسبقا ، اشطف جزءا باستخدام PBS ، ثم ضعه في قاع طبق 35 مم يحتوي على 2 مل PBS. صوتنة لفترة وجيزة باستخدام صوتنة مسبار (50 واط ، سعة 20٪ -40٪ ، مسبار 3 مم) موضوعة 3-5 مم فوق العينة لكشف الخلايا وترك شظايا غشاء البلازما الملتصقة (الشكل 2أ ، ج).
ملاحظة: يمكن أن تختلف قوة Sonicator ومدتها وارتفاع المسبار فوق العينة للحصول على إنتاجية عالية من الأغشية غير المسقوفة دون تعرية انزلاق الغطاء تماما. - إذا لم تستخدم زلات غطاء مطلية مسبقا ، اشطف جزءا من انزلاق الغطاء باستخدام PBS ، ثم اغمس في أنبوب يحتوي على 0.1٪ w / v poly-L-lysine لمدة ~ 30 ثانية قبل صوتنة لفترة وجيزة (كما في الخطوة 4.2.1) لفك الخلايا وترك شظايا غشاء البلازما غير المسقوفة / غير المسقوفة جزئيا (الشكل 2أ ، ج ، د). وقد ثبت أن التعرض القصير لبولي-L-lysine يحسن الالتصاق بغطاءالغطاء 13.
- في حالة استخدام زلات غطاء مطلية مسبقا ، اشطف جزءا باستخدام PBS ، ثم ضعه في قاع طبق 35 مم يحتوي على 2 مل PBS. صوتنة لفترة وجيزة باستخدام صوتنة مسبار (50 واط ، سعة 20٪ -40٪ ، مسبار 3 مم) موضوعة 3-5 مم فوق العينة لكشف الخلايا وترك شظايا غشاء البلازما الملتصقة (الشكل 2أ ، ج).
- ضع الجزء الصوتي في طبق زجاجي بقاع 35 مم يحتوي على محلول حمام سعة 2 مل وقم بتركيبه على مجهر مقلوب مزود بهدف غمر مائي مرتفع NA و 60x. يتم توصيل منفذ الكاميرا الخاص بالمجهر بمطياف في سلسلة بكاميرا CCD عالية الحساسية. قم بتهوية حجرة الحمام (0.5 - 1 مل / دقيقة) مع المخزن المؤقت باستخدام مضخة تمعجية. يختلف تكوين المخزن المؤقت اعتمادا على البروتين قيد الدراسة.
ملاحظة: إذا لم يكن لدى المستخدم إمكانية الوصول إلى هدف بمسافة عمل طويلة ، فقد يكون من المستحيل التركيز على شظايا الغشاء غير المسقوفة بسبب الارتفاع الإضافي لانزلاق الغطاء. البديل هو زرع الخلايا مباشرة على الأطباق ذات القيعان الزجاجية poly-L-lysine (انظر جدول المواد على سبيل المثال). سيؤدي ذلك أيضا إلى تقليل الانحرافات المحتملة في الصورة المرتبطة بالتركيز من خلال قطعتين من الزجاج. هذه الانحرافات لا تؤثر على شكل الأطياف المكتسبة. - حدد شظايا الغشاء غير المسقوفة التي تعبر عن القناة التي تحمل علامة ANAP من خلال البحث عن مضان القناة (الشكل 2ج ، د).
ملاحظة: يوصى باستخدام ملصق فلورسنت إضافي (حيث يمكن تمييز طيف الانبعاث عن طيف انبعاث ANAP) للمساعدة في تحديد الأغشية غير المسقوفة التي تحتوي على البروتين محل الاهتمام. أجريت التجارب في الشكل 2 C ، D على القنوات التي تحمل علامة ANAP مع علامات البروتين الفلوري C-terminal. - قم بإشراك قناع مقياس الطيف جزئيا (رفع ~ 10٪) بين منفذ الكاميرا على المجهر والمطياف. سيظهر ظل القناع على صورة الكاميرا. قم بمحاذاة الغشاء غير المسقوف مع قناع مقياس الطيف ، عن طريق ضبط مرحلة المجهر. الحصول على حقل مشرق وصورة مضان للغشاء غير المسقوف. سيتم استخدام هذه لتحديد منطقة الاهتمام للتحليل.
- اجعل طرف نظام التروية microvolume قريبا من الغشاء غير المسقوف.
ملاحظة: لتقليل مضان الخلفية ، تم استبدال تدفق نظام التروية بطرف مخصص مصنوع من زجاج البورسليكات. - لتصوير أطياف التألق ، قم بإثارة الغشاء باستخدام مصباح LED 385 نانومتر من خلال مرشح إثارة تمرير النطاق 390/18 نانومتر وثنائي اللون بحافة 416 نانومتر. اجمع الضوء المنبعث من خلال مرشح انبعاث طويل 400 نانومتر (الشكل 2 ب).
- قم بإشراك قناع مقياس الطيف وتأكد من مرور الضوء المنبعث. قم بإشراك حواجز شبكية مطياف (300 أخاديد / مم). مع وجود حواجز شبكية في مكانها ، سيتم إسقاط الضوء المنحرف بواسطة مقياس الطيف على شريحة كاميرا CCD لإنتاج صور طيفية (الشكل 3 أ). تحتفظ هذه الصور بالمعلومات المكانية في البعد y . يتم استبدال البعد x بالطول الموجي.
- اختياريا ، إذا تم تمييز البروتين محل الاهتمام ببروتين فلوري ، فاحصل على صورة طيفية للبروتين الفلوري باستخدام مجموعة المرشح المناسبة.
- خذ واحدا أو أكثر من التعرض 0.1-10 ثانية في بداية التجربة أثناء تعطير محلول عازل خال من النيوكليوتيدات. سيتم استخدامها لتصحيح البيانات وتطبيعها طوال بقية التجربة (انظر القسم 5 أدناه).
ملاحظة: يعتمد اختيار وقت التعرض على مستوى التعبير الذي تم تحقيقه وسطوع الفلوروفور والبصريات. يجب اختيار وقت التعرض لزيادة الإشارة وتقليل معدل التبييض المرصود. النطاق الزمني للتعرض الوارد في 4.10 مناسب لقياسات ربط التوازن ولكنه قد يكون مفيدا لقياس التغيرات الحركية البطيئة10. ستكون القدرة على استخدام أوقات التعرض القصيرة لتتبع الحركية الأسرع محدودة بمستويات تعبير البروتين والتبييض الضوئي ، بدلا من الأجهزة. - تطبيق مجموعة من تركيزات TNP-ATP (عادة ما يتم تحضيرها في محلول الحمام) لإنشاء منحنى استجابة التركيز. استخدم كل محلول لمدة دقيقة واحدة على الأقل لضمان الوصول إلى حالة مستقرة واغسل كل تركيز بمحلول الاستحمام لمدة دقيقة واحدة على الأقل.
ملاحظة: من المهم التأكد من أن نظام التروية يمكن أن يصل بسرعة إلى التوازن (الشكل 2E) وتحقيق التركيز المحلي الصحيح ل TNP-ATP (الشكل 2F). - خذ التعرض (بنفس المدة المستخدمة في الخطوة 4.10) عند كل تركيز وفي نهاية كل غسلة.
5. التحليل الطيفي
ملاحظة: تمت كتابة هذه التعليمات للاستخدام مع رمز التحليل "pcf.m" ، والذي يمكن العثور عليه في GitHub. https://github.com/mpuljung/spectra-analysis10. يمكن العثور على رمز إضافي وبديل في https://github.com/smusher/KATP_paper_201911. لقد وصفنا العمليات التي يقوم بها البرنامج هنا حتى يتمكن المستخدم من إنشاء التعليمات البرمجية الخاصة به أو اختيار تحليل البيانات يدويا.
- بدء تشغيل برنامج التحليل عن طريق كتابة اسم البرنامج ("pcf") في سطر الأوامر.
- عند فتح مربع حوار ملف / مجلد مفتوح مع المطالبة: "تحديد ملفات لعائد الاستثمار" ، حدد أسماء الملفات المرتبطة بصور الحقل الساطع والتألق للغشاء غير المسقوف. ستظهر مطالبة في سطر الأوامر لكتابة اسم ملف الإخراج.
- اكتب اسم الملف واضغط على Enter.
- عندما يعرض البرنامج صور الحقل الساطع والتألق ، حدد منطقة الاهتمام (ROI) في الصورة الطيفية المقابلة لموقع جزء الغشاء غير المسقوف أو الرقعة المستأصلة (انظر القسم 6) باتباع مطالبات البرنامج. حدد منطقة خلفية في نفس الصورة الطيفية (تمثل نفس نطاق الطول الموجي كما في عائد الاستثمار) المقابلة لقسم من انزلاق الغطاء أو الطبق بدون غشاء متصل (الشكل 3 أ). سيطالبك البرنامج بالنقر فوق الجزء العلوي من عائد الاستثمار والضغط على Enter ، والنقر فوق الجزء السفلي من عائد الاستثمار والضغط على Enter ثم كرر هذه العملية لمنطقة الخلفية.
- عند فتح مربع حوار ملف / مجلد مفتوح مع المطالبة: "تحديد ملف ل FP Spectrum" ، حدد اسم الملف المرتبط بطيف البروتين الفلوري (FP) (الخطوة 4.9 الاختيارية). إذا لم يتم الحصول على طيف FP ، فحدد ملف طيف مختلف. يعمل طيف FP كمراقبة جودة للتمييز بين البروتين الموسوم ومضان الخلفية.
- عند فتح مربع حوار ملف / مجلد مفتوح مع المطالبة: "تحديد ملفات للتحليل" ، حدد جميع الملفات المقابلة لأطياف ANAP (من الخطوات 4.10 إلى 4.12) ، بما في ذلك الملفات اللازمة لتصحيح التبييض.
- عند فتح مربع حوار ملف / مجلد مفتوح مع المطالبة: "تحديد ملفات لجمع التبييض" ، حدد مجموعة فرعية من الملفات من الخطوة 5.6 المقابلة للأطياف الأولية المكتسبة في محلول خال من النيوكليوتيدات في بداية التجربة أو الأطياف المكتسبة أثناء الغسيل في محلول خال من النيوكليوتيدات لاستخدامها في التصحيح (من الخطوات 4.10 إلى 4.12).
- متوسط الخط لكل صورة لإنتاج أطياف ، أي متوسط كثافة جميع وحدات البكسل في البعد y لعائد الاستثمار أو منطقة الخلفية عند كل طول موجي. (الشكل 3 ب). اطرح متوسط طيف الخلفية الناتج من متوسط الطيف المكتسب من عائد الاستثمار لإزالة مضان الخلفية والتألق من TNP-ATP غير المنضم (الشكل 3C). يتم تنفيذ هذه الخطوات تلقائيا بواسطة البرنامج.
- حدد شدة ANAP لكل تعرض عن طريق حساب متوسط شدة نافذة 5 نانومتر تتمحور حول ذروة ANAP للأطياف المطروحة (عادة ~ 470 نانومتر ولكنها قد تختلف اعتمادا على البيئة المكروية المحلية لبقايا ANAP).
ملاحظة: يوضح الشكل 3D 6 أطياف تم الحصول عليها من التعرض المتتالي لمدة 10 ثوان لجزء غشاء غير مسقوف يعبر عن القنوات الموسومة ب ANAP. يظهر الجزء الداخلي متوسط شدة ذروة كل طيف. سيجد البرنامج تلقائيا الطول الموجي الذروة في الطيف الأول المكتسب ويستخدم هذه القيمة طوال الوقت. سيتم حساب الكثافة تلقائيا بواسطة البرنامج. - قم بتطبيع شدة ANAP لكل تجربة بقسمة شدة ANAP لتعرض معين (F) على شدة ANAP للتعرض الأول في السلسلة الزمنية ، والتي تم التقاطها في الخطوة 4.10 (Fmax). مرة أخرى ، يقوم البرنامج بإجراء هذه الحسابات تلقائيا.
- نفذ الخطوات أدناه للحصول على البيانات.
- لتصحيح التبييض الضوئي ANAP ، قم أولا بتركيب اضمحلال أسي واحد ، (F / F كحد أقصى) = A * exp (-t / τ) + (1-A) ، حيث t هو وقت التعرض التراكمي ، τ هو ثابت الوقت و A هي السعة) إما لخطوات الغسيل الوسيطة بين تطبيقات TNP-ATP أو إلى التعرضات الأولية المتعددة المأخوذة قبل الغسيل على TNP-ATP (الشكل 3D ، أقحم).
ملاحظة: سيعرض البرنامج هذا الملاءمة ويطالب بقبوله أو رفضه. إذا تم رفض الملاءمة ، توفير فرصة أخرى لتحديد الملفات لتصحيح التبييض. - قسم أطياف ANAP الطبيعية (في الخطوة 5.10) على القيمة المتوقعة للملاءمة الأسية من الخطوة 5.11.1 في كل نقطة زمنية (الشكل 3E).
ملاحظة: بالنسبة للمثال الموضح ، فإن ذروة التألق الطبيعية المرصودة عند 50 ثانية هي 0.65 والتألق المتوقع من الملاءمة الأسية هو 0.64. لتصحيح التبييض ، قسم القيمة المرصودة (0.65 ، أقحم الشكل 3E ، دائرة فارغة) على القيمة المتوقعة (0.64 ، أقحم الشكل 3E ، خط متقطع) لإنتاج القيمة المصححة (~ 1 ، أقحم الشكل 3E ، دائرة ملونة). إذا كان تصحيح التبييض كافيا ، فيجب أن تكون شدة ANAP من جميع التعرضات المكتسبة في غياب النيوكليوتيدات متساوية تقريبا (الشكل 3E). يتم إجراء هذه الحسابات تلقائيا بواسطة البرنامج. - احصل على الإخراج كصورة ترسم البيانات وجدول بيانات مبوب يحتوي على الأطياف الأولية والأطياف المطروحة والأطياف المصححة للتبييض الضوئي وبيانات الذروة لكل ملف بحيث يمكن إجراء مزيد من التحليل.
- لتصحيح التبييض الضوئي ANAP ، قم أولا بتركيب اضمحلال أسي واحد ، (F / F كحد أقصى) = A * exp (-t / τ) + (1-A) ، حيث t هو وقت التعرض التراكمي ، τ هو ثابت الوقت و A هي السعة) إما لخطوات الغسيل الوسيطة بين تطبيقات TNP-ATP أو إلى التعرضات الأولية المتعددة المأخوذة قبل الغسيل على TNP-ATP (الشكل 3D ، أقحم).
6. تجارب قياس الفلورة بمشبك التصحيح
- اسحب ماصات التصحيح من الشعيرات الدموية الزجاجية البورسليكات ذات الجدران السميكة إلى مقاومة من 1.5 MΩ إلى 2.5 MΩ عند ملئها بمحلول ماصة. يختلف تكوين محلول الماصة اعتمادا على البروتين قيد الدراسة.
- انقل زلة غطاء مع خلايا منقولة إلى طبق زجاجي القاع 35 مم يحتوي على محلول حمام سعة 2 مل وقم بتركيبه على مجهر مقلوب مزود بهدف غمر مائي عالي NA و 60x. قم بتهوية حجرة الاستحمام (0.5 - 1 مل / دقيقة) بمحلول الاستحمام باستخدام مضخة تمعجية. أما بالنسبة لمحلول الماصة ، فسيختلف محلول الاستحمام اعتمادا على البروتين قيد الدراسة.
- حدد الخلية التي تعبر عن القنوات التي تحمل علامة ANAP من خلال البحث عن التألق في غشاء الخلية.
- املأ ماصة التصحيح بمحلول ماصة. ضع ضغطا إيجابيا لطيفا على الماصة وضعه في حجرة الحمام. اضغط على الماصة على غشاء الخلية وقم بتطبيق شفط لطيف لتحقيق ختم GΩ (الشكل 4 أ).
- قم باستئصال الرقعة عن طريق تحريك حامل الماصة بسرعة بعيدا عن الخلية (الشكل 4 أ).
ملاحظة: يجب أن يشكل استئصال الرقعة بهذه الطريقة رقعة من الداخل إلى الخارج ، مع تعرض المجالات الخلوية للبروتين لنظام التروية. إذا لم يكن موقع ارتباط النوكليوتيدات قيد الدراسة خلويا ، فسيكون من الضروري استخدام بقع خارجية أو تسجيلات خلية كاملة لإجراء تجارب PCF. - اجعل طرف ماصة الرقعة بالقرب من طرف نظام التروية ، وتحقق من أن الرقعة داخل شق قناع مقياس الطيف (الشكل 4 أ).
- قم بتطبيق TNP-ATP وأطياف الصور كما في الخطوات 4.10-4.12 ، مع تسجيل استجابة التيار الأيوني لتطبيق النوكليوتيدات في نفس الوقت.
ملاحظة: قد يحدث زجاج الماصة انحرافات مكانية وانعكاسات في الصور التي تم الحصول عليها. ومع ذلك ، فإن هذه الانحرافات لن تؤثر على شكل الأطياف المكتسبة ويمكن فصل ضوء الإثارة المنعكس بسهولة عن التألق باستخدام إما المطياف أو مرشح الانبعاث طويل التمرير. - تحليل الأطياف. يمكن أن تظهر الأطياف المصورة من البقع المستأصلة الإفراط في طرح مضان TNP-ATP غير المرتبط بسبب استبعاد TNP-ATP من زجاج ماصة الرقعة (الشكل 4C-E). لا يؤثر هذا الطرح الزائد على طيف انبعاث ANAP وبالتالي يمكن تجاهله.
ملاحظة: نظرا لأن إشارة التألق في البقع المستأصلة ستكون أقل منها في الأغشية غير المسقوفة ، فمن المهم استخدام وقت التعرض الذي يعطي إشارة عالية بما يكفي للضوضاء دون تبييض ANAP بسرعة كبيرة.
النتائج
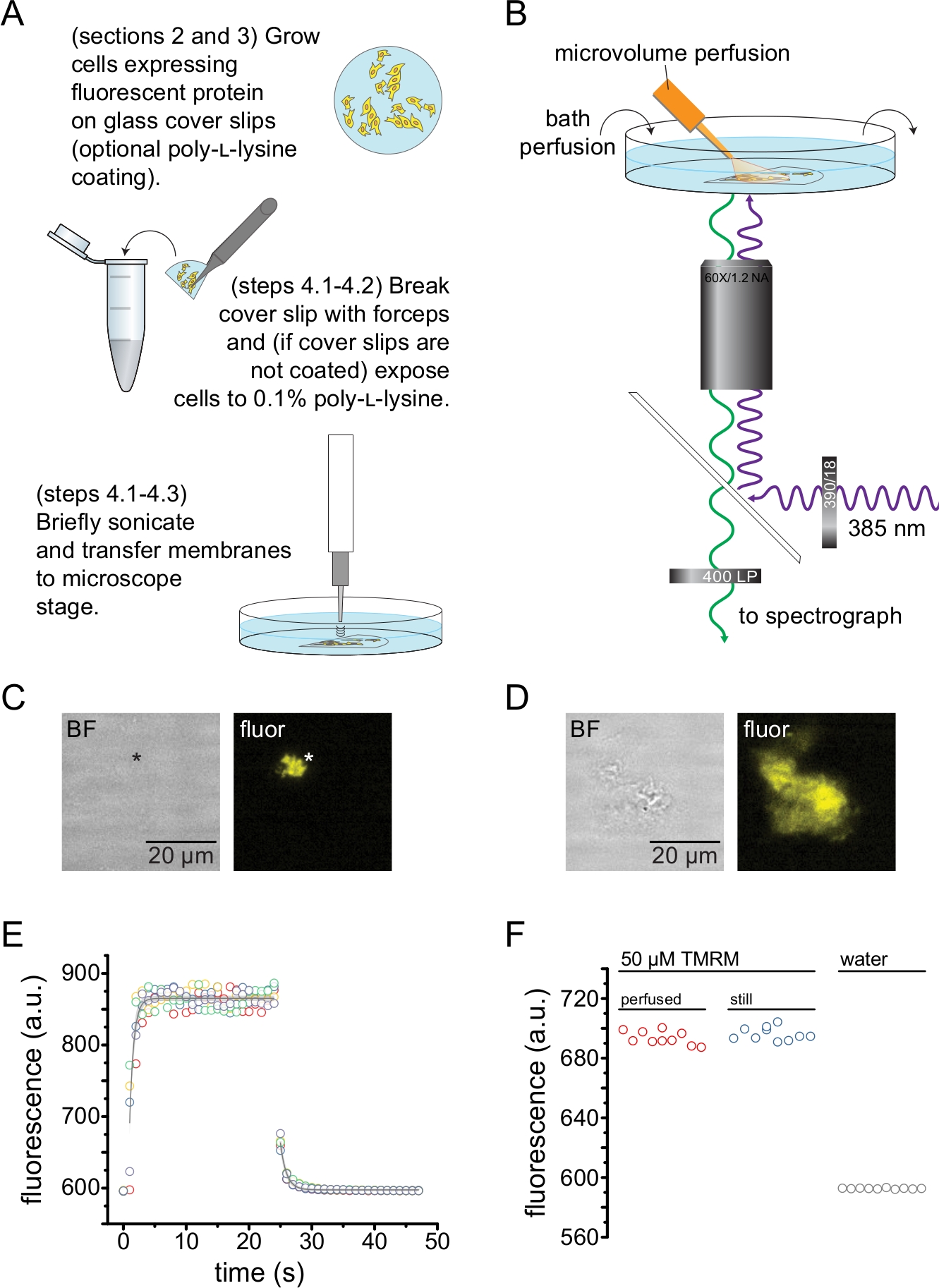
يصور الشكل 2 الإعداد التجريبي الأساسي لقياس ارتباط النوكليوتيدات ببروتينات الفلورسنت في شظايا الغشاء غير المسقوفة التي تم الحصول عليها عن طريق الصوتنة (الشكل 2أ ، ب). تم استخدام طريقتين مختلفتين للحصول على أغشية غير مسقوفة ، زراعة الخلايا مباشرة على زلات الغطاء المطلي بالبولي L-lysine أو زراعة الخلايا على الزجاج غير المعالج وتعريضها لفترة وجيزة ل poly-L-lysine (0.1٪ في الماء) قبل فك التسقيف. يصور الشكل 2C جزءا نموذجيا من غشاء غير مسقوف من خلية HEK-293T تعبر عن قنوات K ATP الموسومة ببروتين الفلورسنت البرتقالي (OFP). كانت الأغشية غير المسقوفة غير مرئية تقريبا في صور المجال الساطع وتم تحديدها من خلال مضان بروتينات الغشاء الموسومة أو عن طريق تلطيخ مضاد بصبغة غشائية مثل أوكتاديسيل رودامين ب13. بالإضافة إلى الأغشية غير المسقوفة ، أنتجت صوتنة خلايا HEK-293T أيضا شظايا خلايا غير مسقوفة جزئيا (الشكل 2D)10,17. كانت هذه الشظايا مرئية في حقل مشرق. قد يكون هذا نتيجة لأغشية البلازما المكشكشة التي لا تلتصق إلا بشكل سيئ بزجاج الغطاء. بدلا من ذلك ، قد تحتوي هذه الشظايا على حويصلات وأغشية من عضيات داخل الخلايا. على هذا النحو ، من الأفضل الحصول على صور فقط من الأغشية غير المسقوفة "الحقيقية" ، حيث أن البروتين المستهدف المسمى المرتبط بالأغشية داخل الخلايا قد يعكس المراحل الوسيطة للمعالجة والتجميع بعد الترجمة. يوصى بزراعة الخلايا على الزجاج المطلي بولي-L-lysine لأن هذا أدى إلى إنتاجية أعلى من الأغشية غير المسقوفة "الحقيقية" عند الصوتنة.
تم تطبيق نظام نضح ميكروحجمي على النيوكليوتيدات الفلورية لتقليل الكميات المطلوبة في تجربة نموذجية (الشكل 2 ب). تم استبدال الطرف الزجاجي المطلي بالبوليميد المقدم بطرف زجاجي من البورسليكات المسحوب يدويا في إعداد التروية لدينا ، مما قلل من الخلفية الفلورية. لتقليل تراكم النوكليوتيدات حول الأغشية غير المسقوفة التي يتم تصويرها ، تم ترشيح غرفة الاستحمام بأكملها ببطء مع العازلة. على هذا النحو ، أردنا قياس معدل تغير المحلول من نظام التروية ذات الحجم الصغير والتحقق من أننا تمكنا من تحقيق تركيز الربيطة المقصود في منطقة اهتمامنا ، أي أن الرباط من نظام التروية لدينا لم يتم تخفيفه مباشرة في وسط الاستحمام قبل الوصول إلى الغشاء غير المسقوف. للتحكم في هذه الاحتمالات ، تم قياس غسل وغسل محلول 50 ميكرومتر من رباعي ميثيل رودامين -5-ماليميد (TMRM) من نظام التروية المجهرية الخاص بنا الموجه إلى سطح طبق زجاجي القاع مغطى بالماء (الشكل 2E). كانت حركية تبادل المحلول قابلة للتكرار وموصوفة جيدا من خلال اضمحلال أسي واحد مع ثوابت زمنية أقل من 1 s لكل من الغسيل والغسيل. تحد أوقات تبادل الحلول هذه من قدرتنا على قياس حركية ربط الليجند وفك الارتباط في إعدادنا الحالي. للتحقق من أننا تمكنا من تحقيق تركيز الرباط المطلوب على سطح انزلاق الغطاء ، قارنا شدة التألق البالغة 50 ميكرومتر TMRM التي يتم تسليمها إلى انزلاق الغطاء بواسطة نظام التروية microvolume الخاص بنا ب 50 μM TMRM في حمام ثابت (الشكل 2F). لم يلاحظ أي اختلاف في الشدة ، مما تحقق من أنه يمكن تحقيق تركيزات الربيطة المناسبة على سطح انزلاق الغطاء باستخدام نظام التروية المجهرية الخاص بنا ، حتى عندما يكون الحمام معطرا.
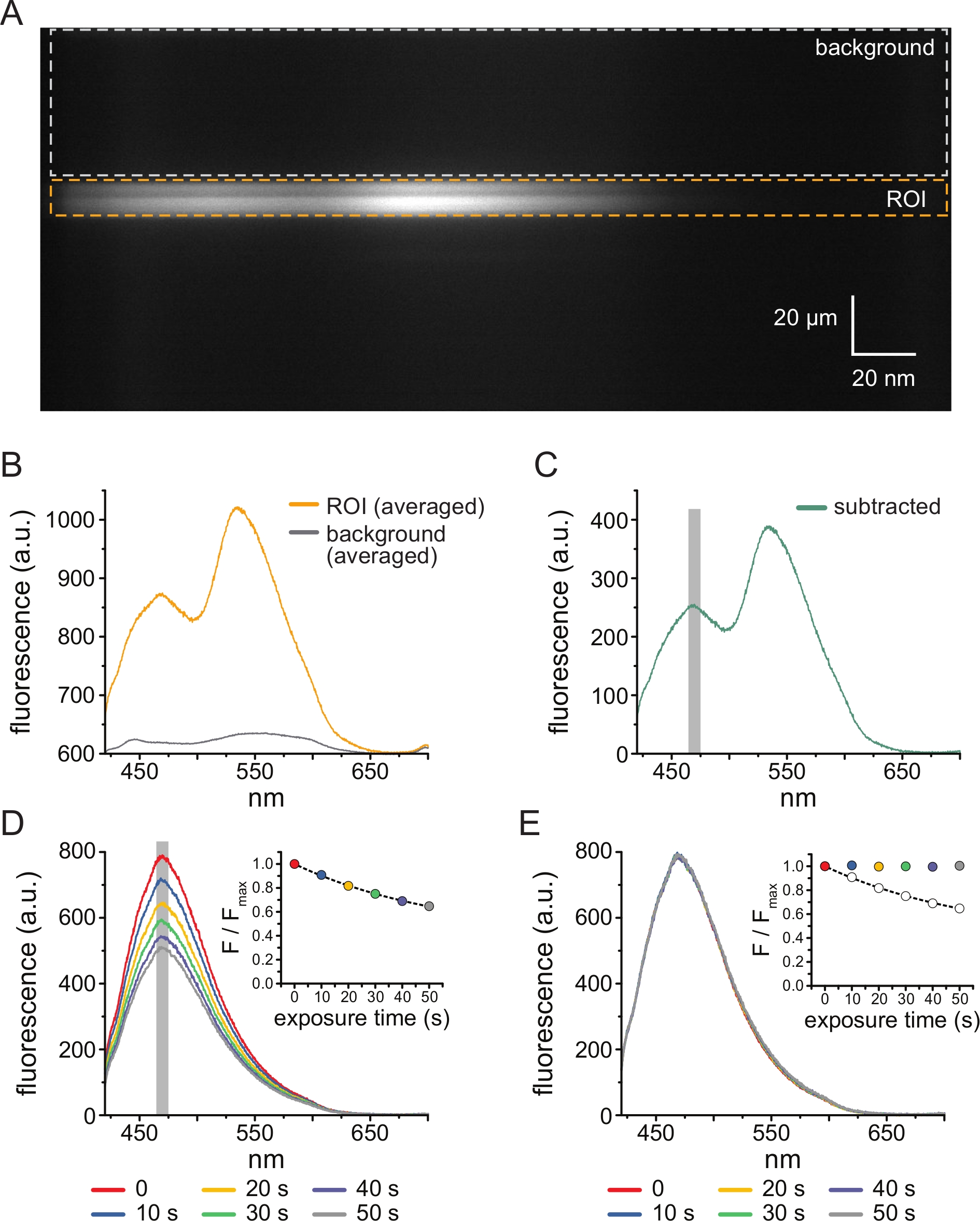
يوضح الشكل 3A صورة طيفية تم الحصول عليها من قنوات KATP الموسومة ب ANAP في غشاء غير مسقوف من خلية HEK-239T معرضة ل 5 μM TNP-ATP. للحصول على مثل هذه الصور ، تم توجيه الضوء المنبعث من الغشاء غير المسقوف من خلال مطياف في سلسلة مع كاميرا CCD. تم انحراف التألق المنبعث من حواجز شبكية وإسقاطه على شريحة الكاميرا ، مما أدى إلى إنتاج أطياف. تحتفظ الصور الناتجة بالمعلومات المكانية في البعد y ، ولكن تم استبدال البعد x بالطول الموجي. تم تحديد منطقة الاهتمام (ROI) ، المقابلة للغشاء غير المسقوف باللون البرتقالي. تتجلى منطقتان من الكثافة العالية في الصورة ، تقابلان ذروة انبعاث ANAP و TNP-ATP. تم تقدير ذلك بشكل أفضل في متوسط الطول الموجي (على مدار عائد الاستثمار بأكمله) الطيف الموضح في الشكل 3B. الذروة ~ 470 نانومتر يتوافق مع ANAP المدمج في KATP ؛ الذروة ~ 535 نانومتر يتوافق مع TNP-ATP. لتصحيح مضان الخلفية والإثارة المباشرة ل TNP-ATP في المحلول ، تم اختيار منطقة خلفية (الشكل 3A ، رمادي) من كل صورة. يظهر متوسط طيف الخلفية في الشكل 3B. تم الحصول على الطيف النهائي عن طريق طرح متوسط طيف الخلفية من متوسط طيف عائد الاستثمار (الشكل 3C).
ANAP عرضة للقطع الأثرية الضوئية. يوضح الشكل 3D الانخفاض في ذروة مضان ANAP بعد التعرض المتعدد. تم تركيب ذروة التألق من عدة تعرضات في غياب TNP-ATP (أو من الغسلات بين تركيزات TNP-ATP) على اضمحلال أسي واحد وتم استخدام هذا لتصحيح القطع الأثرية المبيضة ضوئيا (الشكل 3E). يوصى بإجراء تجارب الاستجابة للتركيز من تركيزات النوكليوتيدات المنخفضة إلى العالية والعالية إلى المنخفضة. إذا لم يقدم تصحيح التبييض أي قطع أثرية إضافية ، فيجب أن تكون النتائج قابلة للمقارنة11.
يوضح الشكل 5A صورا طيفية تمثيلية من غشاء غير مسقوف تم الحصول عليه من خلية تعبر عن قنوات KATP الموسومة ب ANAP في غياب ووجود TNP-ATP. الأطياف المصححة موضحة في الشكل 5B. عند مراقبة أطياف الانبعاثات ، كان هناك فصل واضح بين انبعاث مضان المانح والمستقبل. كما لوحظ بعض الارتباط غير المحدد ل TNP-ATP بأغشية البلازما الساذجة من خلايا HEK-293T غير المنقولة ، فمن المستحسن تحديد FRET كانخفاض في مضان المانحين (ANAP)10,11. كانت هذه الذروة خاصة بالمستقبل المسمى.
بالنسبة للروابط التي تحدث تغييرا مطابقا في مستقبلاتها ، لا توفر دراسات الربط بمعزل عن بعضها البعض معلومات مباشرة وذات مغزى ميكانيكي حول عملية ربط الرباط18. لا تعتمد علاقة التركيز والاستجابة لربط الليجند على تقارب الارتباط الجوهري فحسب ، بل تعتمد أيضا على التغيير التوافقي الناجم عن ربط الربيطة ، والميل المتأصل للمستقبل لتغيير التشكل في غياب الرباط. لفهم العمليات التي تؤكد بشكل أفضل على تفاعلات مستقبلات الرباط ، يمكن إقران قياسات الربط بالتجارب التي توفر قراءة لوظيفة البروتين. تحقيقا لهذه الغاية ، تعد القنوات الأيونية نظاما نموذجيا مثاليا ، حيث يمكن قياس تياراتها بدقة زمنية دون مللي ثانية وصولا إلى مستوى الجزيء الفردي باستخدام مشبك الجهد. تاريخيا ، قدمت قياسات التيار المزدوج والتألق رؤى مهمة حول فتح وإغلاق (بوابة) القنوات الأيونية ذات الجهد والرباط19،20،21. أجريت تجارب لقياس التيارات الأيونية وارتباط النوكليوتيدات الحلقية الفلورية في وقت واحد بمختلف القنوات المنظمة للنيوكليوتيدات الحلقية22،23،24. استخدمت هذه الدراسات ليجند زاد من عائده الكمي عند الربط. يمكن طرح التألق من الرباط غير المنضم في حجم المحلول بالقرب من الرقعة عن طريق تصوير البقع باستخدام المجهر متحد البؤر22,23. في دراساتنا ، تم قياس الارتباط باستخدام الانخفاض في مضان ANAP. نظرا لأن هذه الإشارة خاصة بالقناة و FRET بين ANAP و TNP-ATP تعتمد بشدة على المسافة (نصف الحد الأقصى عند ~ 43 Å) ، فقد تم تجنب تلوث إشارتنا بواسطة النيوكليوتيدات غير المرتبطة وغير المرتبطة على وجه التحديد.
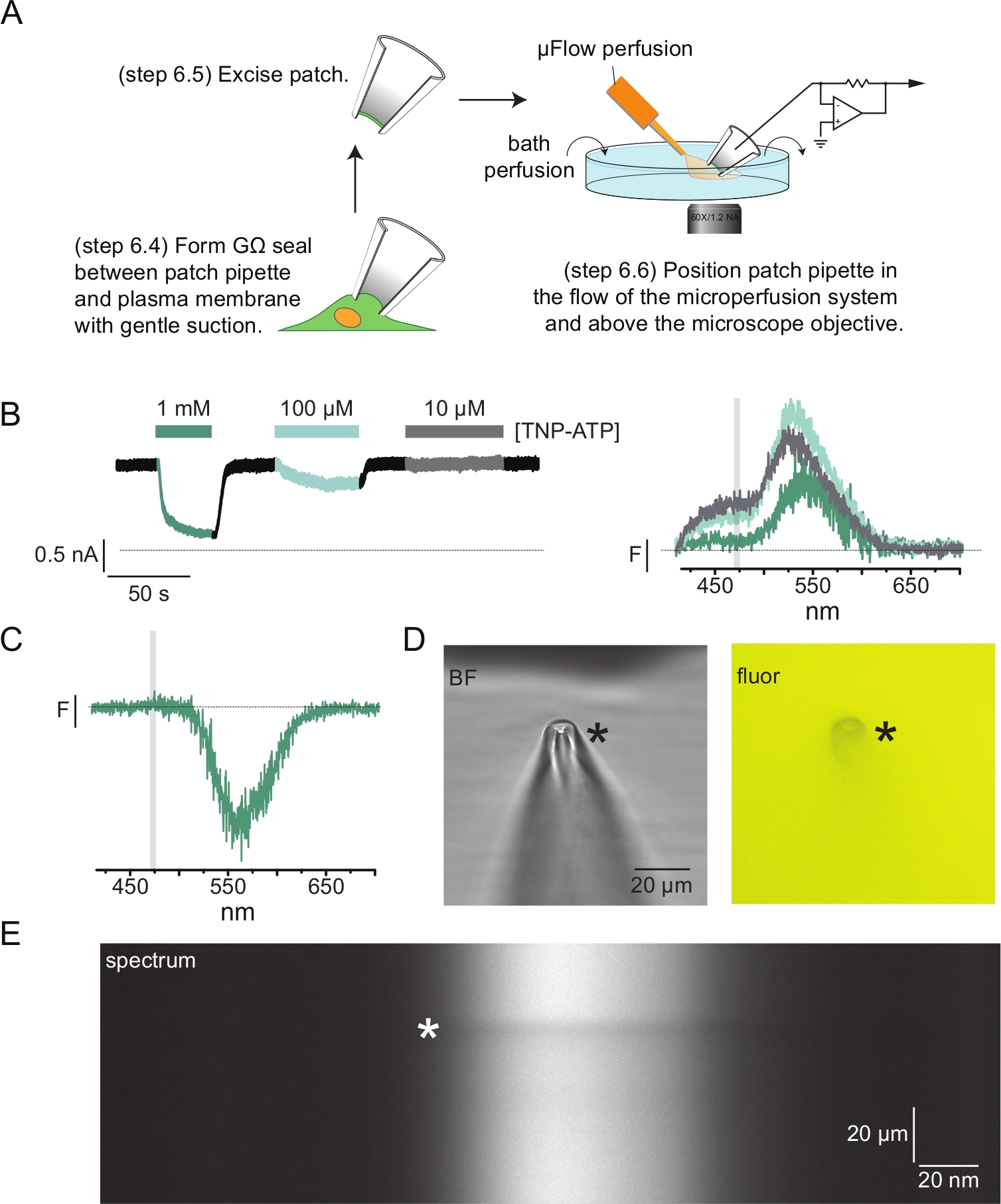
يوضح الشكل 4 أ تجربة نموذجية لقياس الفلور بمشبك التصحيح (PCF). تم تشكيل ختم عالي المقاومة (GΩ) بين ماصة زجاجية من البورسليكات مملوءة بالمحلول الملحي (متصلة بمضخم مشبك الجهد) وخلية تعبر عن KATP الموسوم ANAP. بعد تشكيل الختم ، تم سحب الماصة بعيدا عن الخلية ، مما يسمح بالوصول إلى مواقع ربط النوكليوتيدات داخل الخلايا. ثم تم وضع الماصة فوق هدف المجهر ، متمركزا على شق قناع مقياس الطيف وتم تقريب تدفق نظام التروية المجهري (المعدل بطرف زجاجي من البورسليكات) من الماصة (الشكل 4 د). تم التحكم في الجهد وتم قياس التيارات من القنوات الموجودة في الرقعة. يتم عرض التيارات والأطياف التمثيلية من قنوات KATP الموسومة ANAP في الشكل 4B ، مشفرة بالألوان لمطابقة الأطياف مع التيارات. تم تصحيح أطياف الانبعاث للخلفية والتبييض كما هو الحال بالنسبة للأغشية غير المسقوفة.

الشكل 1: ANAP و TNP-ATP يصنعان زوجا مناسبا من الحنق. (أ) هياكل ANAP و TNP-ATP. يتم تسليط الضوء على أجزاء الفلورسنت. (ب) أطياف انبعاث الامتصاص والتألق ل ANAP و TNP-ATP. التداخل بين انبعاث ANAP وامتصاص TNP-ATP مطلوب ل FRET. مقتبس من Puljung et al. (نشر بموجب رخصة المشاع الإبداعي ، https://creativecommons.org/licenses/by/4.0/)10. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: قياس ارتباط النيوكليوتيدات في أغشية البلازما غير المسقوفة. أ: رسم تخطيطي لتحضير أغشية البلازما غير المسقوفة من الخلايا الملتصقة التي تعبر عن بروتين غشاء فلوري. يتم توفير التعليمات للخلايا المزروعة على زلات غطاء مغلفة بالبولي L-lysine أو غير معالجة. ب: الإعداد التجريبي لقياس ارتباط النيوكليوتيدات في الأغشية غير المسقوفة. (ج) صور المجال الساطع والفلورسنت لغشاء بلازمي غير مسقوف تماما مشتق من خلية تعبر عن بروتين فلوري برتقالي (OFP) موسوم بقنوات KATP. تشير العلامة النجمية إلى موضع الغشاء ، والذي يكاد يكون غير مرئي في صورة المجال الساطع. كان OFP متحمسا بمصباح LED عريض 565 نانومتر من خلال مرشح تمرير نطاق 531/40 نانومتر وتم جمع ضوء ثنائي اللون ومنبعث بحافة 562 نانومتر من خلال مرشح تمرير النطاق 593/40 نانومتر. (د) صور المجال الساطع والفلورسنت لجزء غشاء غير مسقوف جزئيا مشتق من خلية تعبر عن بروتين الفلورسنت البرتقالي (OFP) الموسوم بقنوات KATP. (ه) دورة وقت تبادل الحل المكتسبة باستخدام الإعداد الموضح في B. يتم عرض خمس مكررات تقنية. تم تحميل نظام التروية المجهرية ب 50 ميكرومتر رباعي ميثيل رودامين -5-ماليميد (TMRM). تم رش الحمام بالماء بمعدل ~ 0.5 مل / دقيقة. كانت البيانات من دورات وقت الغسيل (زيادة التألق) والغسيل (تناقص التألق) مناسبة مع اضمحلال أسي واحد للشكل F = A * exp (-x / τ) + y0. كان ثابت الوقت (τ) للغسيل ~ 0.6 ثانية. كان الثابت الزمني للغسيل ~ 1.0 ثانية. كان TMRM متحمسا بمصباح LED عريض 565 نانومتر من خلال مرشح تمرير النطاق 540/25 نانومتر وتم جمع الضوء ثنائي اللون والمنبعث بحافة 565 نانومتر من خلال مرشح تمرير النطاق 605/55 نانومتر. (و) مقارنة شدة التألق لمحلول 50 ميكرومتر من TMRM المطبق باستخدام نظام التروية المجهرية كما في B وحمام ثابت يحتوي على 50 ميكرومتر TMRM. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 3: طرح الخلفية وتصحيح التبييض. (أ) الصورة الطيفية (المعلومات المكانية في البعد y ، الطول الموجي في البعد x) لغشاء بلازمي غير مسقوف من خلية تعبر عن قنوات KATP المسماة ANAP. تم تطبيق 5 μM TNP-ATP باستخدام الإعداد الموضح في الشكل 2B. يشير المربع البرتقالي إلى منطقة الاهتمام (ROI) ، المقابلة للغشاء غير المسقوف. يشير المربع الرمادي إلى منطقة الخلفية المستخدمة لتصحيح الطيف. (ب) أطياف الانبعاث المشتقة من متوسطات الطول الموجي لكل طول موجي لعائد الاستثمار ومناطق الخلفية في A. (C) الطيف المشتق عن طريق طرح متوسط طيف الخلفية من متوسط طيف عائد الاستثمار في B. تظهر نافذة 5 نانومتر حول قمة ANAP المستخدمة لتحديد متوسط الكثافة كمنطقة مظللة باللون الرمادي. (د) الأطياف المكتسبة من ستة تعريضات متتالية لغشاء بلازمي غير مسقوف من خلية تعبر عن قنوات KATP التي تحمل علامة ANAP. لاحظ التناقص في التألق الناتج عن التبييض الضوئي. يظهر الجزء الداخلي أن مضان الذروة الطبيعي يتناسب مع اضمحلال أسي واحد للشكل F / Fmax = A * exp (-t / τ) + (1-A). الرموز الموجودة في الجزء الداخلي مرمزة بالألوان لمطابقة الأطياف. (ه) نفس الأطياف الموجودة في D مصححة للتبييض الضوئي. يظهر الجزء الداخلي مضان الذروة الطبيعي من D كدوائر مفتوحة ، مع عرض مضان الذروة المصحح باستخدام الدوائر المملوءة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 4: القياسات المتزامنة لارتباط النيوكليوتيدات وتيارات القناة باستخدام القياس الفلوري بمشبك التصحيح (PCF). (أ) رسم تخطيطي يوضح الإعداد التجريبي لقياس ارتباط النيوكليوتيدات والتيارات الأيونية. (ب) مثال التيارات (يسار) والأطياف (يمين) المكتسبة من رقعة غشائية مستأصلة من خلية تعبر عن قنوات KATP التي تحمل علامة ANAP. تم تسجيل التيارات عند إمكانية الاحتفاظ ب -60 مللي فولت ، ورقمنتها عند 20 كيلو هرتز ، وتصفيتها عند 5 كيلو هرتز. تتوافق المنطقة المظللة باللون الرمادي مع نطاق الطول الموجي الذي تم من خلاله تحديد شدة ANAP. مقتبس من Usher et al. (نشر بموجب رخصة المشاع الإبداعي ، https://creativecommons.org/licenses/by/4.0/)11. (ج) الطيف المكتسب من رقعة غشائية مستأصلة من خلية تعبر عن قنوات KATP الموسومة ب ANAP المعرضة ل 1 mMM TNP-ATP. لاحظ الذروة السالبة المقابلة لنطاق الطول الموجي الذي لوحظ خلاله مضان TNP-ATP. تشير المنطقة المظللة باللون الرمادي إلى نطاق الطول الموجي المستخدم لتحديد مضان ANAP كما في B. مقتبس من Usher et al. (نشر بموجب رخصة المشاع الإبداعي ، https://creativecommons.org/licenses/by/4.0/)11. (د) صور المجال الساطع والفلورسنت لماصة رقعة معرضة ل 1 mM TNP-ATP. تشير العلامة النجمية إلى طرف الماصة. (ه) صورة طيفية لنفس ماصة الرقعة في 1 mMM TNP-ATP. تشير العلامة النجمية إلى موضع الماصة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

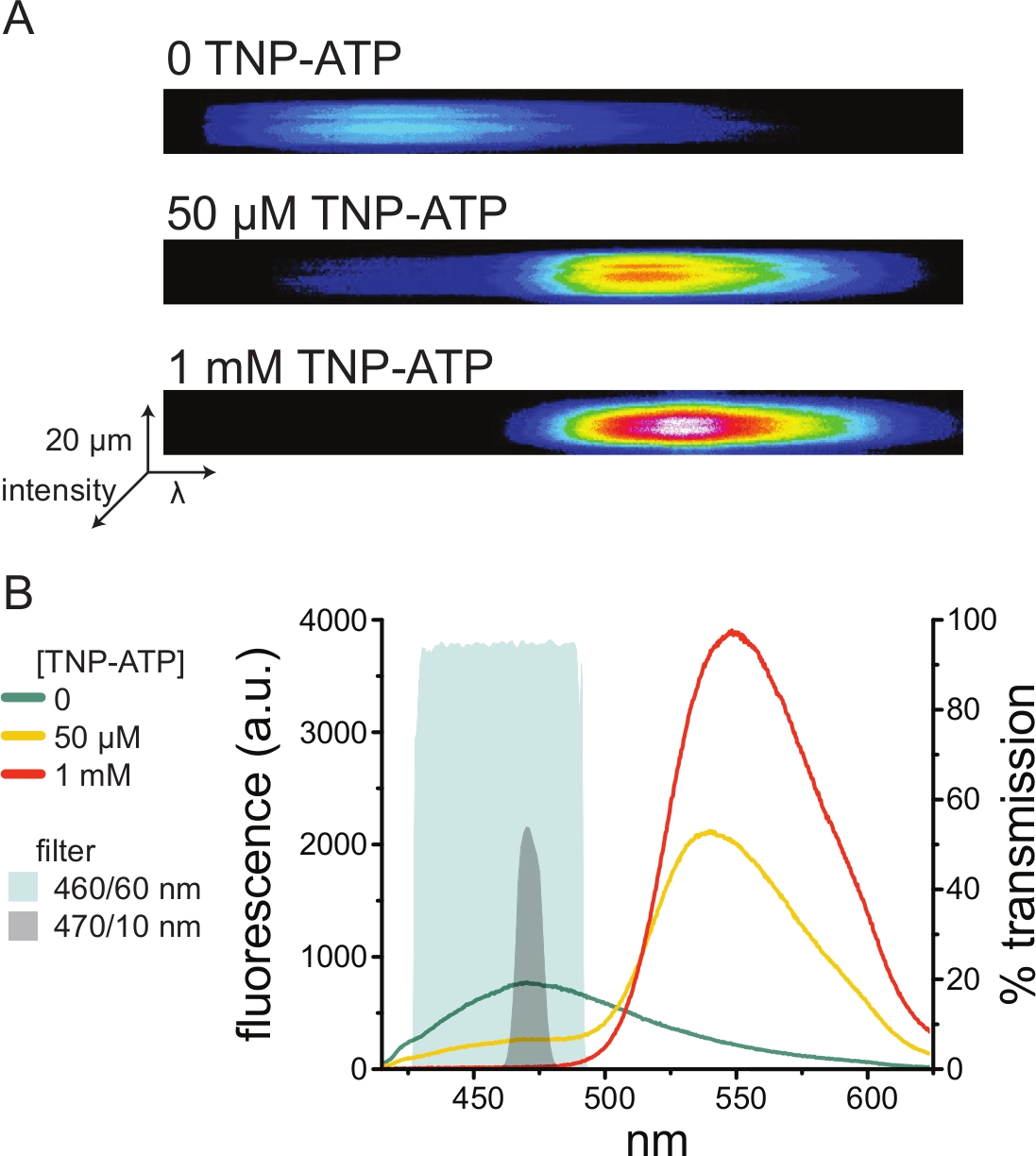
الشكل 5: ارتباط TNP-ATP بقنوات KATP التي تحمل علامة ANAP. (أ) صور طيفية لغشاء بلازمي غير مسقوف من خلية تعبر عن قنوات K ATP التي تحمل علامة ANAP في غياب TNP-ATP أو في وجود 50 ميكرومتر أو 1 مللي متر TNP-ATP. يتم عرض الشدة كخريطة حرارية. (ب) أطياف متوسط الطول الموجي بالطول الموجي من الصور في A تظهر تبريد مضان ANAP بواسطة TNP-ATP. تمثل المناطق المظللة مرشحين مختلفين لتمرير النطاق يمكن استخدامهما لقياس تبريد ANAP في حالة عدم توفر مقياس الطيف. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

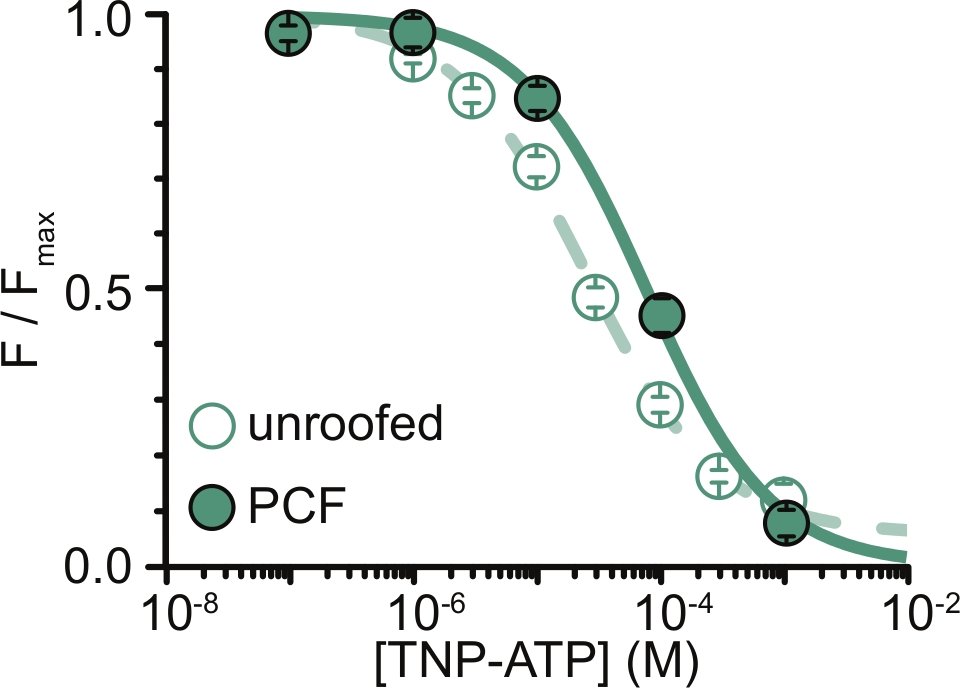
الشكل 6: تبريد قنوات K ATP التي تحمل علامة ANAP بواسطةTNP-ATP في الأغشية غير المسقوفة و PCF. تراكب البيانات من Usher et al. (نشرت بموجب رخصة المشاع الإبداعي ، https://creativecommons.org/licenses/by/4.0/)11. كانت البيانات مناسبة لمعادلة هيل: F / F max = E max + (1 - E max) / (1 + 10 (EC50 - [TNP-ATP]) * h). F هو التألق المقاس ، F max هو الحد الأقصى للتألق في غياب النيوكليوتيد ، Emax هو التبريد الأقصى عند تركيزات النوكليوتيدات المشبعة ، و h هو منحدر هيل. EC50 ، (تركيز النوكليوتيدات الذي يكون فيه التبريد نصف الحد الأقصى) و [TNP-ATP] هي قيم لوغاريتم. الأغشية غير المسقوفة: EC50 = -4.59 (25.7 ميكرومتر) ، ح = 0.82 ، Eكحد أقصى = 0.93. PCF: EC50 = -4.11 (77.6 ميكرومتر) ، ح = 0.87 ، Eكحد أقصى = 1.00. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
Discussion
لقد طورنا طريقة لقياس ارتباط نيوكليوتيدات الأدينين في الوقت الفعلي ببروتينات الغشاء السليمة. تعتمد طريقتنا على العديد من التقنيات الراسخة الأخرى بما في ذلك وضع العلامات على البروتينات باستخدام ANAP باستخدام كودون توقفالكهرماني 12 ، وفك سقف الخلية 14 ، وقياس فلورة مشبك الجهد / PCF19،20،21،22،23،24،25. يسمح توليف هذه الأساليب بقياس ارتباط النوكليوتيدات بدقة مكانية وزمانية عالية. في الواقع ، في عملنا السابق ، تمكنا من التمييز بين مواقع الربط المختلفة على نفس مركب البروتين باستخدام هذا النهج10,11. الأهم من ذلك ، يمكن تطبيق هذه التقنية مباشرة على كميات صغيرة من البروتين في بيئة خلوية في ظل ظروف تحافظ على وظيفة البروتين. يتيح لنا استخدام طريقة الربط الخاصة بنا جنبا إلى جنب مع القراءة الكهربية المباشرة لتيارات القناة الأيونية الحصول على رؤى غنية حول الأسس الجزيئية لبوابةالقناة 11.
نظرا لأن مقاييس الطيف هي قطعة غير قياسية من معدات المختبر ، يمكن أيضا مراقبة شدة ANAP في عزلة نسبية باستخدام مرشحات تمرير النطاق. يصور الشكل 5B الخصائص الطيفية لاثنين من هذه المرشحات. يقوم مرشح تمرير النطاق 470/10 نانومتر بحجب إشارة التألق بشكل فعال من TNP-ATP ويتداخل جيدا مع ذروة مضان ANAP. ومع ذلك ، فإن ذروة نفاذية هذا المرشح تبلغ حوالي 50٪ فقط ، مما قد يجعل من الصعب الحصول على إشارات جيدة من الأغشية المعتمة (أو في بقع الغشاء المستأصلة تحت مشبك الجهد). خيار آخر هو مرشح تمرير النطاق 460/60 نانومتر. هناك تداخل أكثر قليلا بين مرشح 460/60 نانومتر وقدم ذروة انبعاث TNP-ATP مقارنة بمرشح 470/10 نانومتر. ومع ذلك ، فإن ممر النطاق 460/60 نانومتر لديه نفاذية بنسبة 90-95٪ على نطاق واسع من ذروة ANAP ، والتي من المتوقع أن تعزز إشارة انبعاث التألق.
ANAP هو فلوروفور حساس بيئيا12،26،27. تختلف ذروة الانبعاث والعائد الكمي اعتمادا على موقع الدمج على البروتين محل الاهتمام وقد يتغير مع تغير شكل البروتين. ستكون هذه التغييرات واضحة على الفور من أطياف الانبعاثات ولكنها لن تكون واضحة عند قياس شدة ANAP باستخدام المرشحات. على أي حال ، هناك حاجة إلى ضوابط مناسبة لإثبات أن إشارة التألق لا تختلف بسبب التغيرات في البيئة المحلية حول ANAP بعد ارتباط النوكليوتيدات. يمكن أن تساعد تجارب التحكم مع النيوكليوتيدات غير المسماة في التحقق من أن أي تغييرات في شدة ANAP هي نتيجة FRET بين ANAP و TNP-nucleotides. يمكن أن ترتبط نيوكليوتيدات TNP بشكل غير محدد بالأغشية المشتقة من الخلايا غير المنقولة (إما بغشاء البلازما أو ببروتينات الغشاء الأصلية)10. نحن نحدد كمية الارتباط كتناقص في مضان المتبرع ، لأن هذه الإشارة خاصة بالقناة الموصوفة. ومع ذلك ، نوصي بإجراء تجارب تحكم إضافية لكل زوج من الناهضات / المستقبلات ، على سبيل المثال تحور موقع ربط النوكليوتيدات إذا كان معروفا ، للتحقق من أن التغيير في مضان المتبرع هو في الواقع نتيجة الارتباط المباشر بالمستقبل المسمى11. أخيرا ، يوصى بالعمل مع التركيبات التي تحتوي على علامة بروتين الفلورسنت بالإضافة إلى ملصق ANAP. هذا يساعد على التمييز بين مضان المستقبلات المسمى من الخلفية / التألق الذاتي. يمكن تمييز مضان الخلفية عن ANAP من خلال ذروة وشكل أطياف الانبعاث10 ، ولكن مثل هذه التحديدات قد تكون صعبة للغاية عند استخدام مجموعات المرشح فقط. بالإضافة إلى ذلك ، يمكن تحديد الخلايا والأغشية غير المسقوفة التي تعبر عن مستقبلات الفلورسنت باستخدام علامة البروتين الفلوري دون الحاجة إلى إثارة ANAP والمخاطرة بالتبييض الضوئي المفرط.
في العديد من سجلات PCF الخاصة بنا ، لاحظنا ذروة سلبية قوية في أطيافنا عند تركيزات عالية من TNP-ATP (الشكل 4C). هذه القمة السالبة هي قطعة أثرية من بروتوكول الطرح في الخلفية. يوضح الشكل 4D صورا ساطعة المجال وفلورية لماصة رقعة معرضة ل 1 mMM TNP-ATP. الظل عند طرف الماصة واضح ، ناتج عن استبعاد TNP-ATP من حجم جدران الماصة ، والذي يكون أكثر وضوحا داخل مستوى التركيز. تظهر الصورة الطيفية في الشكل 4E شريطا داكنا يتوافق مع هذا الظل. عند استخدام منطقة أعلى أو أسفل هذا الشريط الداكن لطرح الخلفية ، فإنها تنتج قمة سالبة. الأهم من ذلك ، أن هذه الذروة حدثت على نطاق الطول الموجي المقابل لانبعاث TNP-ATP ولم تؤثر على قياساتنا لتبريد ANAP.
كان القيد الرئيسي لتجاربنا هو الحصول على تعبير غشاء البلازما الكافي للتركيبات الموسومة ANAP لقياس التألق. كان من الأسهل عموما الحصول على أطياف عالية الجودة من الأغشية غير المسقوفة مقارنة ب PCF ، نظرا لحجمها الأكبر وقدرتنا على مسح طبق كامل من الأغشية غير المسقوفة بسرعة ، على عكس PCF حيث لا يمكن الحصول على بقع إلا واحدة في كل مرة. في تجاربنا ، كانت البيانات من الأغشية غير المسقوفة وتجارب PCF متشابهة ولكنها ليست متكافئة (الشكل 6)11. ومع ذلك ، لا يوجد سبب مسبق يجعل هذا يجب أن يكون ملاحظة عالمية لأن البروتينات في ماصة التصحيح قد تكون في حالة وظيفية مختلفة عن تلك الموجودة في الأغشية غير المسقوفة.
هنا ، بذلت محاولات لزيادة التعبير عن بنياتنا الموسومة ANAP ، ولا سيما خفض درجة حرارة ثقافة الخلية إلى 33 درجة مئوية10،11،16. في تجربتنا ، فإن محاولة تحديد المواقع في البروتين التي سيكون فيها ANAP بديلا محافظا لم تسفر باستمرار عن تركيبات يتم التعبير عنها بشكل جيد. لقد حققنا نجاحا أكبر في المسح المنهجي لمناطق البروتين بأكملها لمواقع دمج ANAP وفحص المرشحين للتعبير السطحي10. يعمل نظام وضع العلامات ANAP أيضا في بويضات Xenopus laevis ، مما يسمح باستئصال بقع غشائية أكبر بكثير ، وبالتالي زيادة الإشارة إلى الضوضاء26،27،28.
في حين أنه من المتوقع أن تؤدي مستويات التعبير الأكبر إلى إشارات أكثر إشراقا ، فإن الحد الأدنى لعدد القنوات المطلوبة لقياس التألق يعتمد على عدة عوامل ، بما في ذلك سطوع الفلوروفور ، ودرجة التبييض الضوئي ، وشدة ضوء الإثارة ، ومستوى التركيز. من الناحية النظرية ، يمكن إجراء تقديرات من خلال ربط شدة التألق وتيار القناة كما هو موضح سابقا28,29. ومع ذلك ، فإن موثوقية هذه التقديرات تتطلب بعض المعرفة بالتوصيل أحادي القناة والاحتمال المفتوح للقناة. بالإضافة إلى العوامل المذكورة أعلاه ، ستتأثر إشارة التألق أيضا بالقنوات المرتبطة بالحويصلات أو أقسام غشاء البلازما الملتصقة بزجاج الماصة التي لا تخضع لمشبك الجهد.
يتم تكييف هذه الطريقة بسهولة لدراسة القنوات الأيونية الأخرى الحساسة للنيوكليوتيدات. يشبه CFTR هيكليا الوحدة الفرعية لمستقبلات السلفونيل يوريا الملحقة ل KATP30,31. مثلK ATP CFTR يتم التحكم في البوابة عن طريق ربط النوكليوتيدات ، مما يجعلها هدفا مستقبليا واضحا لطريقتنا7. مستقبلات P2X Purinergic هي قنوات أيونية مسورة بواسطة ATP9 خارج الخلية. يعمل TNP-ATP كمضاد لمستقبلات P2X32,33. لذلك ، لن يكون مفيدا لدراسة تنشيط P2X ، على الرغم من أنه يمكن استخدامه في مقايسات المنافسة مع منبهات P2X. بدلا من ذلك ، يمكن استخدام مشتقات ATP الفلورية الأخرى ذات التداخل الطيفي الكافي مع انبعاث ANAP لدراسة التنشيط. Alexa-647-ATP هو ناهض فلوري P2X34. R0 المحسوب بين Alexa-647 و ANAP هو ~ 85 Å ، مما يعني أن الارتباط المباشر ب P2X يجب أن يؤدي إلى تبريد كبير ل ANAP المدمج في القناة. ومع ذلك ، فإن مثل هذا R0 الطويل سيؤدي أيضا إلى التبريد من Alexa-647-ATP المرتبط بالوحدات الفرعية المجاورة ويزيد من احتمال أن يؤدي ارتباط النوكليوتيدات غير المحدد إلى FRET. نظرا لأن موقع ربط الرباط في مستقبلات P2X خارج الخلية ، إجراء قياسات الربط على الخلايا السليمة ، أو في مشبك جهد الخلية بالكامل ، أو في بقع الغشاء الخارجي. يمكن أيضا توسيع طريقتنا لدراسة ربط وتنشيط الناقلات والمضخات الكهروجينية وغير الكهروجينية التي تعتمد على ATP لدورة تفاعلها بالإضافة إلى مستقبلات P2Y المقترنة بالبروتين G. أخيرا ، على الرغم من أننا طورنا هذه الطريقة لقياس ارتباط نيوكليوتيدات الأدينين (TNP-ATP ، TNP-ADP ، TNP-AMP) ، يمكن استخدام نفس النهج لدراسة الارتباط بأي مستقبل تقريبا تم تحديد ليجند فلوري مناسب له.
Disclosures
يعلن أصحاب البلاغ عدم وجود تضارب في المصالح.
Acknowledgements
نود أن نشكر راؤول تيرون إكسبوزيتو على المساعدة التقنية الممتازة. وقد مول هذا العمل مجلس بحوث التكنولوجيا الأحيائية والعلوم البيولوجية (BB/R002517/1; MCP و FMA) وصندوق ويلكوم (203731/Z/16/A; SGU)
Materials
| Name | Company | Catalog Number | Comments |
| T75 tissue-culture treated flask | StarLab | CC7682-4875 | |
| 0.1% w/v poly-L-lysine | Sigma-Aldrich | P8920 | |
| 30 mm borosilicate cover glass slips | VWR | 631-0174 | |
| 35 mm non-treated sterile dishes | CytoOne | CC7672-3340 | |
| 35 mm cover glass bottom dish | WPI | FD35-PDL-100 | |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 31966021 | |
| Foetal bovine serum (FBS) | Gibco | 10500-064 | |
| Penicillin/Streptomycin | Gibco | 15140-122 | |
| TrypLE select (tryosin) | Gibco | 12563-011 | Trypsin/EDTA reagent |
| Phosphate buffered saline (PBS) | Gibco | 14040-091 | |
| UltraPure distilled water | Invitrogen | 10977-035 | |
| HEK293T cells | ATTC | CRL-3216 | Used between passages 5-30 |
| ANAP-TFA | AsisChem | ASIS-0014 | Reconstituted in 30 mM NaOH to a final concentration of 1 mM |
| pANAP expression plasmid | Addgene | Plasmid #48696 | Encodes tRNA/tRNA synthetase pair for expression of ANAP-tagged protein |
| peRF1-E55D | Chin Lab (MRC Laboratory of Molecular Biology, Cambridge, UK) | Jason Chin: DOI: 10.1021/ja5069728 | Encodes dominant-negative eukaryotic ribosomal release factor |
| TransIT-LT1 | Mirus Bio | MIR 2300 | Lipopolyplex transfection reagent |
| Thick-walled borosilicate glass capillaries | Harvard Apparatus | GC150F-15 | |
| Tetramethylrhodamine-5-maleimide | Sigma-Aldrich | 94506 | |
| TNP-ATP | Jena Bioscience | NU-221L | Delivered at 10 mM in water |
| Nikon Eclipse TE2000-U inverted microscope microscope | Nikon | ||
| 60x water immersion objective (1.4 NA) | Nikon | MRD07602 | |
| 4-Wavelength High-Power LED Head | ThorLabs | LED4D245 | 385/490/565/625 nm LEDs |
| Four-Channel LED Driver | ThorLabs | DC4100 | |
| 390/18 nm band-pass excitation filter | ThorLabs | MF390-18 | For ANAP excitation |
| 400 nm long-pass emission filter | ThorLabs | FEL0400 | For imaging ANAP spectra |
| 416 nm edge dichroic | ThorLabs | MD416 | For imaging ANAP spectra |
| 460/60 nm band-pass emission filter | ThorLabs | MF460-60 | Suggested wide band-pass filter for imaging ANAP fluorescence (Figure 4B) |
| 470/10 nm band-pass emission filter | ThorLabs | FB470-10 | Suggested narrow band-pass filter for imaging ANAP fluorescence (Figure 4B) |
| 531/40 band-pass excitation filter | Brightline | FF01-531/40-25 | For orange fluorescent protein (OFP) excitation |
| 540/25 nm band-pass excitation filter | Chroma | D540/25X | For tetramethylrhodamine-5-maleimide (TMRM) excitation |
| 562 nm edge dichroic | Semrock | FF562-Di03 | For imaging OFP fluorescence |
| 565 nm edge dichroic | Chroma | 565DC | For imaging TMRM fluorescence |
| 593/40 nm band-pass excitation filter | Brightline | FF01-387/11-25 | For imaging OFP fluorescence |
| 605/55 nm band-pass emission filter | Chroma | D605/55M | For imaging TMRM fluorescence |
| IsoPlane-160 Imaging Spectrometer | Princeton Instruments | IsoPlane-160 | |
| PIXIS 400BR_eXcelon Camera | Princeton Instruments | PIXIS: 400BR_eXcelon | |
| Axopatch 200B amplifier | Molecular Devices | Axopatch 200B-2 | |
| Digidata 1440A digitizer | Molecular Devices | Digidata 1440A | |
| Probe sonicator | Sonics & Materials | VC-50 | For unroofing |
| REGLO digital peristaltic pump | Ismatec | ISM 832 | For bath perfusion |
| Microvolume perfusion system | ALA Scientific Instruments | ALA μFlow-8 | For TNP-ATP perfusion |
| pClamp 10.6.2 | Molecular Devices | Recording and analysing currents | |
| Lightfield 5.20.1507 | Princeton Instruments | Acquisition software for images and spectra | |
| Matlab | Mathworks | For data analysis | |
| Python 3.8.1 | Python Software Foundation | For data analysis |
References
- Garcia, M. L., Kaczorowski, G. J. Ion channels find a pathway for therapeutic success. Proceedings of the National Academy of Sciences of the United States of America. 113 (20), 5472-5474 (2016).
- Hauser, A. S., Attwood, M. M., Rask-Andersen, M., Schioth, H. B., Gloriam, D. E. Trends in GPCR drug discovery: new agents, targets and indications. Nature Reviews Drug Discovery. 16 (12), 829-842 (2017).
- Lakowicz, J. R. Principles of fluorescence spectroscopy. 3rd edn. , Springer. (2006).
- Higgins, C. F., Linton, K. J. The ATP switch model for ABC transporters. Nature Structural & Molecular Biology. 11 (10), 918-926 (2004).
- Toyoshima, C., Cornelius, F. New crystal structures of PII-type ATPases: excitement continues. Current Opinion in Structural Biology. 23 (4), 507-514 (2013).
- Craven, K. B., Zagotta, W. N. CNG and HCN channels: two peas, one pod. Annual Review of Physiology. 68, 375-401 (2006).
- Csanady, L., Vergani, P., Gadsby, D. C. Strict coupling between CFTR's catalytic cycle and gating of its Cl- ion pore revealed by distributions of open channel burst durations. Proceedings of the National Academy of Sciences of the United States of America. 107 (3), 1241-1246 (2010).
- Vedovato, N., Ashcroft, F. M., Puljung, M. C. The Nucleotide-Binding Sites of SUR1: A Mechanistic Model. Biophysical Journal. 109 (12), 2452-2460 (2015).
- Burnstock, G. Introduction to the Special Issue on Purinergic Receptors. Advances in Experimental Medicine and Biology. 1051, 1-6 (2017).
- Puljung, M., Vedovato, N., Usher, S., Ashcroft, F. Activation mechanism of ATP-sensitive K(+) channels explored with real-time nucleotide binding. Elife. 8, 41103(2019).
- Usher, S. G., Ashcroft, F. M., Puljung, M. C. Nucleotide inhibition of the pancreatic ATP-sensitive K+ channel explored with patch-clamp fluorometry. Elife. 9, 52775(2020).
- Chatterjee, A., Guo, J., Lee, H. S., Schultz, P. G. A genetically encoded fluorescent probe in mammalian cells. Journal of the American Chemical Society. 135 (34), 12540-12543 (2013).
- Gordon, S. E., Senning, E. N., Aman, T. K., Zagotta, W. N. Transition metal ion FRET to measure short-range distances at the intracellular surface of the plasma membrane. Journal of General Physiology. 147 (2), 189-200 (2016).
- Heuser, J. The production of 'cell cortices' for light and electron microscopy. Traffic. 1 (7), 545-552 (2000).
- Schmied, W. H., Elsasser, S. J., Uttamapinant, C., Chin, J. W. Efficient multisite unnatural amino acid incorporation in mammalian cells via optimized pyrrolysyl tRNA synthetase/tRNA expression and engineered eRF1. Journal of the American Chemical Society. 136 (44), 15577-15583 (2014).
- Lin, C. Y., et al. Enhancing Protein Expression in HEK-293 Cells by Lowering Culture Temperature. PloS One. 10 (4), 0123562(2015).
- Usukura, J., et al. Use of the unroofing technique for atomic force microscopic imaging of the intra-cellular cytoskeleton under aqueous conditions. Journal of Electron Microscopy. 61 (5), 321-326 (2012).
- Colquhoun, D. Binding, gating, affinity and efficacy: the interpretation of structure-activity relationships for agonists and of the effects of mutating receptors. British Journal of Pharmacology. 125 (5), 924-947 (1998).
- Mannuzzu, L. M., Moronne, M. M., Isacoff, E. Y. Direct physical measure of conformational rearrangement underlying potassium channel gating. Science. 271 (5246), 213-216 (1996).
- Zheng, J., Zagotta, W. N. Gating rearrangements in cyclic nucleotide-gated channels revealed by patch-clamp fluorometry. Neuron. 28 (2), 369-374 (2000).
- Zheng, J., Zagotta, W. N. Patch-clamp fluorometry recording of conformational rearrangements of ion channels. Science's STKE. 2003 (176), 7(2003).
- Biskup, C., et al. Relating ligand binding to activation gating in CNGA2 channels. Nature. 446 (7134), 440-443 (2007).
- Kusch, J., et al. Interdependence of receptor activation and ligand binding in HCN2 pacemaker channels. Neuron. 67 (1), 75-85 (2010).
- Wu, S., et al. State-dependent cAMP binding to functioning HCN channels studied by patch-clamp fluorometry. Biophysical Journal. 100 (5), 1226-1232 (2011).
- Cha, A., Bezanilla, F. Characterizing voltage-dependent conformational changes in the Shaker K+ channel with fluorescence. Neuron. 19 (5), 1127-1140 (1997).
- Kalstrup, T., Blunck, R. Dynamics of internal pore opening in K(V) channels probed by a fluorescent unnatural amino acid. Proceedings of the National Academy of Sciences of the United States of America. 110 (20), 8272-8277 (2013).
- Kalstrup, T., Blunck, R. S4-S5 linker movement during activation and inactivation in voltage-gated K(+) channels. Proceedings of the National Academy of Sciences of the United States of America. 115 (29), 6751-6759 (2018).
- Dai, G., Aman, T. K., DiMaio, F., Zagotta, W. N. The HCN channel voltage sensor undergoes a large downward motion during hyperpolarization. Nature Structural & Molecular Biology. 26 (8), 686-694 (2019).
- Liu, C., et al. Patch-clamp fluorometry-based channel counting to determine HCN channel conductance. Journal of General Physiology. 148 (1), 65-76 (2016).
- Hwang, T. C., et al. Structural mechanisms of CFTR function and dysfunction. Journal of General Physiology. 150 (4), 539-570 (2018).
- Puljung, M. C. Cryo-electron microscopy structures and progress toward a dynamic understanding of KATP channels. Journal of General Physiology. 150 (5), 653-669 (2018).
- Kasuya, G., et al. Structural insights into the competitive inhibition of the ATP-gated P2X receptor channel. Nature Communications. 8 (1), 876(2017).
- Virginio, C., Robertson, G., Surprenant, A., North, R. A. Trinitrophenyl-substituted nucleotides are potent antagonists selective for P2X1, P2X3, and heteromeric P2X2/3 receptors. Molecular Pharmacology. 53 (6), 969-973 (1998).
- Bhargava, Y., Nicke, A., Rettinger, J. Validation of Alexa-647-ATP as a powerful tool to study P2X receptor ligand binding and desensitization. Biochemical and Biophysical Research Communications. 438 (2), 295-300 (2013).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved