
Method Article
قياسات القوة المباشرة للميكانيكا دون الخلوية في الحبس باستخدام ملاقط بصرية
In This Article
Summary
هنا ، نقدم بروتوكولا للتحقيق في الخواص الميكانيكية داخل الخلايا لخلايا الزرد الجنينية المعزولة في الحبس ثلاثي الأبعاد مع قياس القوة المباشرة بواسطة مصيدة بصرية.
Abstract
أثناء تطور كائن حي متعدد الخلايا ، تنقسم خلية مخصبة واحدة وتؤدي إلى أنسجة متعددة ذات وظائف متنوعة. يسير مورفوجين الأنسجة جنبا إلى جنب مع التغيرات الجزيئية والهيكلية على مستوى الخلية الواحدة التي تؤدي إلى اختلافات في الخواص الميكانيكية دون الخلوية. ونتيجة لذلك ، حتى داخل نفس الخلية ، تقاوم العضيات والمقصورات المختلفة بشكل مختلف الضغوط الميكانيكية. ويمكن لمسارات النقل الميكانيكي أن تنظم بنشاط خصائصها الميكانيكية. وبالتالي فإن قدرة الخلية على التكيف مع البيئة الدقيقة لمكانة الأنسجة ترجع جزئيا إلى القدرة على الشعور بالضغوط الميكانيكية والاستجابة لها. لقد اقترحنا مؤخرا نموذجا جديدا للإحساس الميكانيكي حيث يمكن التشوه النووي وتحديد المواقع الخلية من قياس بيئة 3D المادية ويمنح الخلية إحساسا بالاستقبال العميق لفك تشفير التغيرات في شكل الخلية. في هذه المقالة ، نصف طريقة جديدة لقياس القوى وخصائص المواد التي تشكل نواة الخلية داخل الخلايا الحية ، والتي تتمثل في الخلايا الملتصقة والخلايا المحصورة ميكانيكيا. يمكن إجراء القياسات بشكل غير جراحي باستخدام مصائد بصرية داخل الخلايا ، ويمكن الوصول إلى القوى مباشرة من خلال الكشف الخالي من المعايرة عن زخم الضوء. وهذا يسمح بقياس ميكانيكا النواة بشكل مستقل عن تشوهات سطح الخلية والسماح بتشريح مسارات النقل الميكانيكي الخافتة والمتقبلة. الأهم من ذلك ، يمكن دمج تجربة الاصطياد مع المجهر الضوئي للتحقيق في الاستجابة الخلوية والديناميكيات دون الخلوية باستخدام التصوير الفلوري للهيكل الخلوي أو أيونات الكالسيوم أو المورفولوجيا النووية. الطريقة المقدمة سهلة التطبيق ، ومتوافقة مع الحلول التجارية لقياسات القوة ، ويمكن توسيعها بسهولة للتحقيق في ميكانيكا المقصورات تحت الخلوية الأخرى ، على سبيل المثال ، الميتوكوندريا ، وألياف الإجهاد ، والاندوسومات.
Introduction
تشكل الأنسجة هو عملية معقدة يتم فيها تنسيق الإشارات الكيميائية الحيوية والقوى الفيزيائية مكانيا وزمانيا. في الجنين النامي ، تملي تدرجات عوامل الإشارات الكيميائية الحيوية مواصفات المصير وتضمن نمط الأنسجة الصحيح1,2. في الوقت نفسه ، تلعب القوى الجوهرية والخارجية دورا في بناء بنية الجنين3,4. تمت دراسة تأثير ميكانيكا قشرة الخلية في هذا السياق على نطاق واسع5,6. يعتمد الترابط الوثيق بين العمليات الميكانيكية والكيميائية أثناء التشكل على خصائص الخلايا المفردة لاستشعار القوى الميكانيكية في بيئتها الدقيقة للأنسجة والاستجابة لها. الخلايا، وبالتالي فك تشفير الإشارات الميكانيكية من خلال وجود عناصر دون خلوية وجزيئية حساسة للقوة التي تنقل المعلومات الميكانيكية إلى مسارات إشارات محددة تتحكم في سلوك الخلية، ومصير الخلية، وميكانيكا الخلايا.
السمة المميزة للعمليات التنموية هي أن الخلايا تنظم كمجموعات لبناء هياكل متعددة الخلايا. وعلى هذا النحو، نادرا ما تعيد الخلايا المفردة ترتيبها وتتحرك بمفردها ولكنها ترتبط ببيئة اجتماعية ضيقة تظهر فيها سلوكا جماعيا مثل الهجرة فوق الخلوية7 أو انتقالات التشويش8,9 أو ضغط الكيسة الأريمية10. تعمل القوى الميكانيكية المتولدة داخل الخلايا وفيما بينها كإشارات مهمة لتوجيه ديناميكيات الخلايا الجماعية7،11. ولكن حتى عندما تتحرك الخلايا بمفردها، مثل الخلايا السلفية التي تشق طريقها بين صفائح الأنسجة أو منافذ الأنسجة الضيقة، فإنها تواجه قوى ميكانيكية متباينة الخواص واسعة النطاق عند التنقل في بيئة ثلاثية الأبعاد. هذه الضغوط الميكانيكية على الخلايا لها عواقب وخيمة على السلوك الخلوي12,13. تم التحقيق في العديد من الآليات التي تتلاقى على النواة كعنصر نقل ميكانيكي رئيسي14,15 ، كعنصر ميكانيكي سلبي أو نشط أثناء الهجرة داخل بيئة نسيجية ثلاثية الأبعاد كثيفة15,16.
اقترحنا مؤخرا آلية تزود الخلايا بقياس تشوهات الشكل باستخدام النواة كمقياس ميكانيكي مرن داخل الخلايا12. تخضع النواة ، كونها أكبر عضية في الخلية ، لتشوهات كبيرة عندما تستقطب الخلايا أو تهاجر أو تغير شكلها تحت التمدد الميكانيكي أو الحبس أو الإجهاد الأسموزي 16،17،18،19. وجدنا أن تمدد الغلاف النووي جنبا إلى جنب مع تحديد المواقع داخل الخلايا للنواة يزود الخلايا بمعلومات عن حجم ونوع تشوه الخلية (مثل ضغط الخلايا مقابل تورم الخلايا). يرتبط تمدد النواة بتكشف الغشاء النووي الداخلي (INM) ، الذي يعزز نشاط الليباز cPLA2 المعتمد على الكالسيوم (الفوسفوليباز الخلوي A2) في INM متبوعا بإطلاق حمض الأراكيدونيك (AA) والتنشيط السريع للميوسين الثاني في قشرة الخلية. وهذا يؤدي إلى زيادة انقباض الخلايا وهجرة الخلايا الأميبية فوق عتبة الانقباض القشري6. تحدث الاستجابة الحساسة الميكانيكية لتشوه الخلايا في أقل من دقيقة ويمكن عكسها عند إطلاق الحبس ، مما يشير إلى أن النواة تعمل كمقياس إجهاد للاستحسان الخلوي الذي ينظم سلوك الخلية التكيفي في ظل ظروف الإجهاد الميكانيكي. ويظهر أن هذا المسار الحساس للميكانيكية نشط في الخلايا الجذعية السلفية المشتقة من أجنة الزرد، سواء في الخلايا متعددة القدرات أو الملتزمة بالنسب12 ويتم حفظه في أنواع وخطوط خلايا مختلفة20.
بالإضافة إلى الخصائص النووية كمحن ميكانيكي للخلايا، يتم تنظيم البنية النووية والميكانيكا بشكل جوهري أثناء التطوير واستجابة لمواصفات مصير الخلية21، وبالتالي ضبط الحساسية الميكانيكية الخلوية22,23. وقد تكون النتيجة تغيرا في الامتثال النووي يسمح بالتغيرات والتحولات المورفولوجية من حالة ما قبل الهجرة إلى حالة الهجرة والعكس بالعكس8.
تم تطبيق العديد من التقنيات لقياس ميكانيكا نواة الخلية، مثل مجهر القوة الذرية24,25، وشفط الأنابيب الدقيقة26،27، وتكنولوجيا الموائع الدقيقة28، والوخز بالإبر الدقيقة29. ومع ذلك ، فإن العديد من هذه التقنيات غازية بمعنى أنه يجب تشويه الخلية بأكملها ، مما يحد من قياس الخصائص الميكانيكية والاستجابات المعتمدة على القوة للنواة نفسها. للتحايل على التشوه المتزامن لسطح الخلية وقشرة الخلايا الحساسة ميكانيكيا30 ، تمت دراسة النوى المعزولة في سياقات مختلفة31,32. ومع ذلك، لا يمكن استبعاد أن العزل النووي يرتبط بتغير في خصائص النواة الميكانيكية وتنظيمها (المرجع 24 والملاحظات الخاصة غير المنشورة).
الملقط البصري (OTs) هي تقنية متعددة الاستخدامات سمحت بمجموعة كبيرة من التجارب في علم الأحياء الميكانيكية الخلوية وكان لها دور فعال في فهمنا لكيفية تحويل الآلات الجزيئية للطاقة الكيميائية إلى طاقة ميكانيكية33,34. تستخدم الملقط البصري شعاع ليزر مركز بإحكام لممارسة قوى بصرية على الجسيمات العازلة التي لها معامل انكسار أعلى من الوسط المحيط33. يمكن أن تكون هذه القوى في حدود مئات البيكو نيوتن وتؤدي إلى حبس فعال للجسيم داخل تركيز مصيدة الليزر ، مما يتيح التلاعب بالجسيم المحاصر في ثلاثة أبعاد. استخدام الضوء له ميزة مهمة في أنه يمكن إجراء القياس بشكل غير جراحي داخل الخلايا الحية. تقتصر التلاعبات البصرية أيضا على تركيز المصيدة لشعاع الليزر. وبالتالي ، يمكن إجراء التلاعب دون تحفيز الأغشية الخلوية المحيطة ولا يزعج قشرة الأكتين أو العمليات الحساسة الميكانيكية في الغشاء البلازمي ، مثل التنشيط المعتمد على القوة للقنوات الأيونية.
تكمن صعوبة نهج الملقط البصري في التحديد الدقيق للقوى المطبقة على المجهر باستخدام الأساليب الكلاسيكية التي تعتمد على معايرة القوة غير المباشرة استنادا إلى نظرية التقسيم المتساوي أو استخدام قوى سحب ستوكس المحددة لقياس قوة هروب تعتمد على طاقة الليزر35. في حين أن هذه الطرق سهلة التنفيذ في تجربة في المختبر ، إلا أنه لا يمكن ترجمتها عادة إلى بيئة خلوية. تم إدخال العديد من الاستراتيجيات في هذا المجال التي تعتمد على معايرة القوة المباشرة، المستمدة من المبادئ الأولى للحفاظ على الزخم36,37. على عكس أساليب التحليل الطيفي للقوة الأخرى، يتم استنتاج قياسات القوة من تبادل محلي لزخم الضوء مع الجسيم المحاصر على شكل تعسفي38,39. في إعدادنا التجريبي، يتم قياس التغيرات في زخم الضوء الناشئة عن القوى البصرية مباشرة دون الحاجة إلى معايرة المصيدة في الموقع40،41،42،43. وبالتالي ، تصبح القياسات ممكنة في بيئة لزجة مثل الجزء الداخلي من الخلية أو حتى داخل الأنسجة ، ويمكن قياس القوى بسهولة إلى مستوى pN.
في هذا البروتوكول ، نصف فحصا لمعالجة العضيات أو الهياكل داخل الخلايا ميكانيكيا وتقييم خصائصها الميكانيكية كميا بواسطة إعداد ملاقط بصري. تم دمج هذا الإعداد في مجهر الفلورسنت القرص الدوار الذي يتيح التصوير المتوازي للسلوك الخلوي أو الديناميكيات داخل الخلايا. يسمح الفحص بتوصيف الخواص الميكانيكية لمقصورات خلوية محددة ، مثل النواة ، مع دراسة الميكانوريسبونس المحتمل وتنشيط مسارات الإشارات الجزيئية في نفس الوقت نتيجة للتشوه نفسه. وعلاوة على ذلك، فإن الاصطياد البصري للميكروبات المحقونة داخل الخلايا يسمح بزيادة قوة المسافة البادئة بفضل معامل انكسار أعلى بكثير من حبة البوليسترين (n = 1.59) مقارنة بالتباين الانكساري الداخلي44 للنواة (n ~ 1.35) مقابل السيتوبلازم (n ~ 1.38). يمكن تكييف الاستراتيجية المقدمة بسهولة مع دراسة الهياكل والعضيات الأخرى داخل الخلايا ، بالإضافة إلى النهج الأخرى التي تنطوي على علم الأحياء الدقيقة النشط ، واستخدام مصائد بصرية متعددة لفحص نفس الهياكل دون الخلوية / المختلفة في وقت واحد ، والقياسات التي تستهدف علم الأحياء الميكانيكية للخلايا في الجنين الحي.
Protocol
تمت الموافقة على جميع البروتوكولات المستخدمة من قبل اللجنة المؤسسية لأخلاقيات رعاية واستخدام الحيوانات (PRBB-IACUEC) وتنفيذها وفقا للوائح الوطنية والأوروبية. تم إجراء جميع التجارب وفقا لمبادئ 3Rs. تم الحفاظ على الزرد (دانيو ريريو) كما هو موضح سابقا.
1. تحضير الخلايا الجذعية السلفية الجنينية الجنينية المعزولة
- الأنابيب الدقيقة وإعداد الأغاروز
ملاحظة: للحصول على بروتوكول حقن دقيق كامل لجنين الزرد، انظر المرجع 45.- باستخدام مجتذب الأنابيب الدقيقة، اسحب شعرية زجاجية مقاس 1.0 مم للحصول على إبرتين45. قم بتخزين الإبر غير المستخدمة في طبق بتري مقاس 150 مم متصل بوسادة عجين اللعب أو في حلقة شريط مختبر من الداخل إلى الخارج لحماية الطرف الرفيع من التلف أثناء النقل.
- تذوب الأغاروز فائق النقاء بنسبة 1٪ في E3 (5 mM NaCl ، 0.17 mM KCl ، 0.33 mM CaCl2 ، 0.33 mM MgSO4) في ميكروويف المطبخ / المختبر القياسي لمدة 10 ثوان. سخني المزيج مرارا وتكرارا لفترات قصيرة من الزمن (بضع ثوان) حتى يذوب الأغاروز.
- عندما يذوب الأغاروز تماما ، اتركه يبرد لفترة وجيزة ، ثم اسكبه في طبق بتري 10 سم. أضف ببطء قالب الحقن المجهري الثلاثي (انظر جدول المواد) في الجزء العلوي من الأغاروز لتجنب ظهور الفقاعات. لا تدفع القالب ، مما يضمن بقاءه على سطح الأغاروز.
- عندما يتصلب الأغاروز تماما ، قم بإزالة القالب الثلاثي ببطء شديد عن طريق ممارسة قوة لطيفة لتجنب أي فواصل في الأغاروز. يمكن تخزين اللوحة رأسا على عقب عند 4 درجات مئوية لمدة 2-4 أسابيع.
- قبل 30 دقيقة من الحقن المجهري ، أخرج الطبق من الثلاجة وأضف E3 مسبقا إلى 28 درجة مئوية للسماح له بالاستقرار في درجة حرارة الغرفة.
- إعداد مزيج الحقن
- لتحضير خليط الحقن ، قم بتخفيف 1 ميكروبيدات ميكرومتر (البوليسترين ، غير الفلورسنت) بنسبة 1: 5 في الماء الخالي من RNase.
- تحضير الحمض النووي الريبوزي المرسال للتعبير العابر عن علامات الفلورسنت أو التعبير عن التركيبات الجينية المؤتلفة و / أو الحقن المشترك للمورفولينو بالتركيز المطلوب.
ملاحظة: خليط حقن نموذجي للحقن المشترك للميكروبات مع 100 جزء من الحمض النووي الريبوزي المرسال لكل جنين لتسمية ، على سبيل المثال ، النواة مع H2A-mCherry هي: 1 ميكرولتر من الخرز + 1 ميكرولتر من الحمض النووي الريبي المرسال (تركيز المخزون هو 1 ميكروغرام / ميكرولتر) + 2.5 ميكرولتر من الماء الخالي من الحمض النووي الريبي + 0.5 ميكرولتر من الفينول الأحمر (محلول المخزون 0.5٪ ، الفينول الأحمر ليس إلزاميا ؛ يتم استخدامه لتصور أفضل للقطرة المحقونة ولكن الحقن غير الموسوم قطرة مرئية أيضا لمجرب من ذوي الخبرة). يمكن أن يكون حقن الحمض النووي الريبي مفيدا أيضا لاختيار الأجنة المحقونة. يمكن حقن الميكروبيدات الفلورية ، بدلا من غير الفلورسنت ، لتصورها.
- تحميل ومعايرة إبرة الحقن المجهري
- قم بتشغيل الحاقن الدقيق باستخدام خيار Time-Gated . هذا الإعداد مهم جدا لمعايرة حجم الحقن بشكل صحيح. اضبط وقت البوابة على حوالي 500 مللي ثانية.
- قم بتحميل 3 ميكرولتر من خليط الحقن في الإبرة باستخدام ماصة محمل صغير.
- أدخل الإبرة في جهاز المعالجة الدقيقة وأغلقها بإحكام. تحقق مما إذا كان المعالجة الدقيقة في وضع جيد ولديه حرية كافية للتحرك في اتجاه x-y على لوحة الحقن.
- قم بقياس حجم القطرة باستخدام شريحة ميكرومتر (5 مم / 100 قسم) مع قطرة من الزيت المعدني في الأعلى45 وإخراج قطرة من مزيج الحقن مباشرة في الزيت المعدني.
- اقتصاص الإبرة بملقط حاد بزاوية حادة لتوليد طرف مدبب حاد. اضبط حجم القطرة على 0.1 مم ، أي ما يعادل 0.5 نانولتر من المواد المحقونة.
ملاحظة: إذا تم تجاوز هذا الحجم عن طريق قطع الإبرة، فمن المستحسن إعادة إجراء المعايرة باستخدام إبرة جديدة. يمكن ضبط وقت البوابة للحاقن الدقيق قليلا ليتناسب مع حجم السقوط ؛ ومع ذلك ، فإن أوقات البوابة القصيرة تتوافق مع قطر إبرة كبير ، مما قد يؤدي إلى تلف الأجنة.
- الحقن المجهري لأجنة الزرد في مرحلة الخلية الواحدة
- اجمع أجنة الزرد بعد فترة وجيزة من الإخصاب للحقن المجهري لخليط الخرز مباشرة في جنين مرحلة الخلية الواحدة (الزيجوت) قبل حدوث الانقسام الخلوي الأول.
ملاحظة: يضمن ذلك التوزيع السليم للميكروسفير وإنتاجية عالية بما فيه الكفاية من الأرومات المعزولة مع ميكروسفير واحد على الأقل لكل خلية في مراحل النمو اللاحقة التي يتم فيها إجراء التجارب (مرحلة الأرومة الغازية). لا يزال من الممكن إجراء تجارب المسافة البادئة إذا كان هناك مجالان داخل الخلية ، ولكن يجب استبعاد الخلايا التي لا تحتوي على حبات (على الرغم من أن المسافة البادئة بدون كرات ممكنة). تم استخدام سلالات AB البرية في هذا البروتوكول ، ولكن يمكن استخدام أي سلالة أخرى ، على سبيل المثال ، TL. - ضع أجنة المرحلة ذات الخلية الواحدة (الزيجوت) في قالب أغاروز مثلث الشكل 1٪ ، كما هو موضح في الشكل 1A ، باستخدام ماصة باستور بلاستيكية.
- قم بإزالة الوسط الإضافي بنفس الماصة لتجنب الأجنة العائمة. ادفع الأجنة برفق إلى القالب المثلث عبر فرشاة. احتفظ ببعض المساحة بين الأجنة لتسهيل الاتجاه الصحيح (الشكل 1B).
- قم بمحاذاة الأجنة بلطف باستخدام فرشاة بحيث يتم توجيه الأجنة أفقيا ، مع رؤية الخلية الواحدة من الزيجوت بوضوح ، كما هو موضح في الشكل 1B. يتم الوصول إلى الاتجاه المثالي للحقن المجهري عندما تواجه خلية واحدة من الجنين اتجاه الإبرة (الحقن عبر القطب الحيواني للجنين) أو في الاتجاه المعاكس الذي يواجه خلية صفار البيض (الحقن عبر القطب النباتي للجنين) ، كما هو موضح في الشكل 1C.
- أمسك الطبق بيد واحدة واستخدم اليد الأخرى لوضع طرف الإبرة باستخدام وحدة التحكم في المعالجة الدقيقة. اخفض طرف الإبرة نحو الأجنة.
- اخترق المشيمة وأدخل الجنين أحادي الخلية بالإبرة أثناء مراقبة الإجراء من خلال المجهر المجهري. تأكد من الموضع الصحيح للإبرة ، وبعد الحقن ، الموقع الصحيح للقطرة المحقونة كما هو موضح في الشكل 1C.
- كرر ذلك لجميع الأجنة: حرك الإبرة لأعلى ، وحرك الطبق مع الأجنة حتى يتم توسيط الجنين التالي ، واخفض الإبرة ، وحقنها.
- بمجرد حقن المجموعة الكاملة من الأجنة ، قم بإزالة الأجنة من قالب الأغاروز / طبق بتري عن طريق مسح بعض E3 ووضعها في طبق بتري جديد باستخدام ماصة باستور بلاستيكية. يوصى بوضع وسائط كافية على صفيحة الحقن لتجنب جفاف الأجنة أثناء إجراء الحقن المجهري.
- كرر الإجراء حتى يتم حقن العدد المطلوب من الأجنة. يجب أن تكون الأجنة في مرحلة خلية واحدة لضمان الانتشار الأقصى والمتجانس للخرز.
ملاحظة: تم تحسين هذا الإجراء لأجنة الأرومة المبكرة ومن المحتمل أن يحتاج إلى تحسين إذا كان يجب التحقيق في مراحل النمو المختلفة. - ضع الأجنة المحقونة داخل حاضنة عند 28-31 درجة مئوية لمدة 4 ساعات تقريبا أو حتى المرحلة المطلوبة (الشكل 1D) قبل الشروع في بروتوكول زراعة الخلايا الأولية.
ملاحظة: اختياريا، دع الأجنة تتطور إلى ما بعد مرحلة الأرومة (أو نقطة القياس الزمنية المطلوبة) لضمان البقاء على قيد الحياة واستبعاد القطع الأثرية السامة. في مراحل اليرقات ، قم بتركيب يرقات مخدرة مع tricaine في 0.75 ٪ من الأغاروز وصور توزيع الميكروسفير في الأنسجة المختلفة. لصنع محلول مخزون ، امزج: 400 ملغ من مسحوق التريكايين في 97.9 مل من الماء المقطر ، وحوالي 2.1 مل من 1 M TRIS-base (الرقم الهيدروجيني 9) ، واضبط على الرقم الهيدروجيني 7. يمكن تخزين هذا الحل عند 4 درجات مئوية. لاستخدام التريكايين كمخدر ، قم بتخفيف 4.2 مل من محلول المخزون في 100 مل من وسط البيض (أو الوسائط المطلوبة) ؛ في هذه الحالة ، تم استخدام E3. راجع المرجع46 للحصول على التفاصيل.
- اجمع أجنة الزرد بعد فترة وجيزة من الإخصاب للحقن المجهري لخليط الخرز مباشرة في جنين مرحلة الخلية الواحدة (الزيجوت) قبل حدوث الانقسام الخلوي الأول.
2. إعداد خلية واحدة وتلطيخ
- ضع أجنة المرحلة الكروية (4 hpf ، ساعات بعد الإخصاب) في طبق زجاجي باستخدام ماصة باستور بلاستيكية. حدد الأجنة الإيجابية لإشارة الخرز المحقون ، والتي تعبر عن بروتين الفلورسنت في حالة حقن الحمض النووي الريبوزي المرسال. قد تظهر بعض الأجنة تجمعا عاليا للخرز ويمكن استبعادها.
- قم بإزالة الأجنة يدويا باستخدام الملقط. انقل ما يقرب من 10-15 جنينا إلى حاويات تفاعل سعة 1.5 مل باستخدام ماصة باستور زجاجية.
ملاحظة: عندما يتم نزع الأجنة ، فإنها تلتصق بالبلاستيك ، ويلزم استخدام الأواني الزجاجية. كبديل للصفيحة الزجاجية ، يمكن استخدام طبق بتري بلاستيكي بطبقة رقيقة من 1٪ من الأغاروز. يجب تفضيل التطهير اليدوي على علاج البروناز الأنزيمي لمنع تلف البروتين البروتيني لبروتينات سطح الخلية والتغيرات المحتملة في خصائص الخلايا والأنسجة الميكانيكية ، مما يمنع أوقات الاسترداد الممتدة47.
- قم بإزالة الأجنة يدويا باستخدام الملقط. انقل ما يقرب من 10-15 جنينا إلى حاويات تفاعل سعة 1.5 مل باستخدام ماصة باستور زجاجية.
- قم بإزالة وسائط E3 وأضف 500 ميكرولتر من وسط زراعة الأنسجة المستقل عن CO2 الذي تم تسخينه مسبقا (DMEM-F12 ؛ مع L-glutamine و 15 mM HEPES ، بدون بيكربونات الصوديوم والفينول الأحمر المكمل ب 10 وحدات من البنسلين و 10 ملغم / لتر من الستربتومايسين).
ملاحظة: لا تستخدم الوسائط المعتمدة على CO2 ما لم يتم استخدام حاضنة مجهر. استخدام ، على سبيل المثال ، RPMI في الظروف المخزنة مؤقتا للكربونات يسبب تغيرات في درجة الحموضة في الوسائط ويمكن أن يؤثر على بقاء الخلية. جانب رئيسي آخر هو تجنب وسائل الإعلام الثقافية التي تحتوي على المصل. قد يحتوي المصل على حمض الليسوفوسفاتيديك (LPA)، وهو منشط قوي لمسار Rho/ROCK، قادر على التحكم في الانقباض الخلوي والحركة في الخلايا الجذعية السلفية6. يجب الحفاظ على أسمولية الوسط عند 300 mOsm لتجنب التحديات الأسموزية التي يمكن أن تتداخل مع المورفولوجيا النووية أو الميكانيكا 12. - قم بفصل الخلايا يدويا عن طريق هز الأنبوب بلطف. تأكد من أن محتويات الأنبوب تصبح عكرة مع عدم وجود قطع كبيرة مرئية بالعين. تجنب تكوين فقاعات لتقليل تلف وفقدان الخلايا.
- جهاز طرد مركزي عند 200 × جم لمدة 3 دقائق. يجب أن تكون الكرية مرئية بوضوح.
- قم بإزالة supernatant واتبع إحدى الخطوات المفصلة أدناه.
- إذا لم تكن هناك حاجة إلى تلطيخ ، أضف 500 ميكرولتر من DMEM. أعد تعليقه بلطف باستخدام ماصة سعة 200 ميكرولتر عن طريق استهداف نفاثة سائلة على الكريات. لا تمارس قوة القص المفرطة على الخلايا. تشير الرغوة إلى تلف الخلايا.
- لوضع العلامات على النواة بأصباغ الحمض النووي مثل Hoechst ، امزج 0.5 ميكرولتر من DNA-Hoechst (مخزون 2 ملغ / مل) في 1000 ميكرولتر من DMEM للحصول على 1 ميكروغرام / مل من التركيز النهائي. أضف 500 ميكرولتر من محلول التلطيخ هذا إلى الخلايا وأعد تعليقه بلطف. احتضن لمدة 7 دقائق في الظلام.
- لتلطيخ الخلايا بمؤشر الكالسيوم الكيميائي الفلورسنت Calbryte-520 ، أضف Calbryte-520 إلى تركيز 5 ميكرومتر في DMEM. احتضن لمدة 20 دقيقة في الظلام.
ملاحظة: تم تحسين البروتوكولات المشار إليها في الخطوتين 2.5.2 و2.5.3 لهذه المنتجات المحددة. يمكن إجراء تلطيخ آخر باستخدام البروتوكولات المشار إليها من قبل الشركة المصنعة.
- جهاز الطرد المركزي مرة أخرى باستخدام نفس الإعدادات كما في الخطوة 2.4 ؛ قم بإزالة supernatant ، وأعد تعليق الخلايا بلطف (لتجنب تكوين مجموعات) في 50 ميكرولتر من DMEM للعينات المعلقة أو 20 ميكرولتر من DMEM للخلايا في الحبس.
3. إعداد غرف الاصطياد البصري باستخدام تباعد البولي ديميثيل سيلوكسان (PDMS)
ملاحظة: تتطلب قياسات القوة البصرية المستندة إلى اكتشاف زخم الضوء التقاط كل الضوء الخارج من المصائد البصرية40. من أجل متانة عامل المعايرة الثابت α (pN/V)، يجب أن يحمل توزيع الضوء في المستوى البؤري الخلفي (BFP) لمستشعر القوة البصرية تطابقا دقيقا مع زخم الفوتون. يحدد هذا المسافة من سطح عدسة التجميع إلى مستوى الملائمة إلى حوالي 2 مم ، وهو الحد الأقصى لارتفاع غرف الملائمة البصرية.
- PDMS طلاء الدوران من # 1.5 أطباق القاع الزجاجي.
ملاحظة: يتم توفير الوصفة التالية لحوالي 40 طبقا. سيكون للغرفة الصغيرة الناتجة ارتفاعات مختلفة اعتمادا على ما إذا كان سيتم إجراء التجارب على الخلايا المعلقة أو المحصورة (الشكل 1D).- امزج 9 مل من البوليمر الأساسي PDMS و 1 مل من عامل معالجة PDMS في أنبوب مخروطي سعة 50 مل. امزج المنتجين بنشاط لضمان التوزيع السليم لعامل المعالجة.
- قم بإزالة الغاز من الخليط لتجنب الفقاعات باستخدام مضخة فراغ. أدخل الأنبوب المخروطي في زجاجة فراغ وقم بإخلاء الغرفة. انتظر حتى لا توجد فقاعات في الخليط.
ملاحظة: افتح الفراغ ببطء لمنع رغوة وانسكاب PDMS خارج أنبوب الصقر. - ضع الطبق السفلي الزجاجي على ظرف المعطف الدوار (الشكل 2A). كن لطيفا حتى لا تخدش الطبق أو بصمته أو تتسخ. حماية مربع معطف الدوران من تسرب PDMS مع رقائق الألومنيوم.
- بالنسبة لغرف OT للتجارب على الخلايا المعلقة ، أضف حوالي 250 ميكرولتر من خليط PDMS في وسط الطبق السفلي وقم بتدويره عند 750 دورة في الدقيقة لمدة 1 دقيقة. سيكون ارتفاع طبقة PDMS 50 ميكرومتر تقريبا48.
- بالنسبة لغرف OT للتجارب على الخلايا المحصورة ، أضف قطرة صغيرة من PDMS (حوالي 50 ميكرولتر) وقم بتدويرها عند 4000 دورة في الدقيقة لمدة 5 دقائق. سيكون ارتفاع طبقة PDMS 10 ميكرومتر تقريبا. للحصول على بروتوكول مفصل حول كيفية الحصول على سماكات PDMS مختلفة، راجع المرجع 48.
- علاج الأطباق الزجاجية السفلية المغلفة PDMS عند 70 درجة مئوية لمدة 1 ساعة.
- قطع مربع 1 × 1 سم على طبقة PDMS بمشرط وقشره بملاقط (الشكل 2C). في حالة الخلايا المحصورة ، اغسل حطام PDMS بالأيزوبروبانول.
- طلاء الغرفة للتجارب مع الخلايا المتصلة بخفة في التعليق
- أضف 100 ميكرولتر من Concanavalin A (ConA) عند 0.5 ملغم / مل لتغطية كامل سطح التجويف المربع واتركه يحتضن لمدة 30 دقيقة.
ملاحظة: ConA هو الليكتين الذي يرتبط بالسكريات السطحية للخلايا ويقرن الخلايا الفردية على سطح الغطاء الزجاجي. - قم بإزالة قطرة ConA واشطف السطح بعناية باستخدام وسط DMEM دون خدش السطح المعالج ب ConA.
- أضف 30 ميكرولتر من العينة المعدة مسبقا (الخطوة 2.6) إلى البئر وأعيد تعليقها بلطف للتخلص من أي مجموعات خلوية.
- أغلق التجويف عن طريق وضع زجاج غطاء 22 × 22 مم # 1.5 برفق فوق حواف PDMS (تجنب تركه يسقط فجأة ، واستخدم الملقط إن أمكن ، الشكل 2B ، C).
ملاحظة: أي سمك coverslip ستعمل مع الغطاء الزجاجي العلوي (العدسة التجميع لديها مسافة عمل 2 مم).
- أضف 100 ميكرولتر من Concanavalin A (ConA) عند 0.5 ملغم / مل لتغطية كامل سطح التجويف المربع واتركه يحتضن لمدة 30 دقيقة.
- إعداد الغرفة للتجارب مع الخلايا في الحبس
- ضع قطرة 10 ميكرولتر من المحلول الذي يحتوي على خلايا (الخطوة 2.6) في التجويف المربع (الشكل 2B).
- بلطف شديد ، شطيرة العينة مع زجاج غطاء 22 × 22 مم بحيث ينتشر القطرة في المنطقة بأكملها ولا يتم ملاحظة أي فقاعات. مرة أخرى ، من المريح استخدام الملقط ، كما هو موضح في الشكل 2C ، لمنع زجاج الغطاء من السقوط فجأة.
4. خيارات بديلة لتباعد غرف OT
ملاحظة: يمكن اتباع هذه الخطوات في حالة عدم توفر ورشة تصنيع دقيق أو معطف غزل.
- إعداد الغرفة للتجارب مع الخلايا المعلقة
ملاحظة: في حالة عدم توفر معطف الدوران ، يمكن عمل فاصل باستخدام شريط سكوتش عادي على الوجهين (يبلغ ارتفاعه حوالي 100 ميكرومتر).- اقطع قطعة من الشريط الاسكتلندي على الوجهين بفتحة مربعة تبلغ مساحتها حوالي 10 سم × 10 سم في الوسط (نفس الأبعاد كما في PDMS ، الشكل 2B).
- قم بإزالة إحدى الطبقات الواقية من الشريط عن طريق تقشيره ووضع الجانب المكشوف من الشريط في وسط طبق زجاجي سفلي # 1.5 H. اضغط برفق للحصول على كل السطح ملتصقا بالزجاج مع تجنب فقاعات الهواء ، ثم قم بإزالة الطبقة الواقية المتبقية من الشريط عن طريق تقشيره.
- اتبع الإرشادات الواردة في الخطوة 3.2.
- إعداد الغرفة للتجارب مع الخلايا في الحبس
ملاحظة: لحصر الخلايا بدقة، يمكن استخدام الجسيمات الدقيقة أحادية التشتت ذات القطر المعروف كفواصل بين نظارتي الغطاء.- أضف 10 ميكرومتر من حبات البوليسترين إلى الخلايا العالقة بتركيز 104 حبات / ميكرولتر.
- ضع قطرة 10 ميكرولتر من المحلول الذي يحتوي على خلايا وخرز على زجاج غطاء 22 × 60 مم.
- بلطف شديد ، شطيرة العينة مع زجاج غطاء آخر 22 × 60 مم بحيث ينتشر الانخفاض في المنطقة بأكملها ولا يتم ملاحظة أي فقاعات. لوضع زجاج الغطاء العلوي بلطف (تجنب سقوطه فجأة) ، من المريح استخدام الملقط.
- نظرا لأن العينة يمكن أن تجف ، فمن المستحسن إجراء التحضير بسرعة.
5. إعداد المصيدة البصرية للقياسات داخل الخلايا
ملاحظة: تم تحسين الخطوات التالية لمنصة ملاقط بصرية تجارية تتألف من وحدة معالجة دقيقة بصرية تعتمد على الانحراف الصوتي البصري (AOD) ومستشعر قوة بصرية يعتمد على الكشف المباشر عن تغيرات زخم الضوء (الشكل 2، المرجع 12،40،49). يمكن العثور على التفاصيل والمكونات البصرية للإعداد في الشكل 2F. لمراقبة التشوه الناجم عن القوة أثناء التلاعب بالملقط البصري ، يتم إقران المجهر البؤري للقرص الدوار Nipkow في المنفذ الأيسر للمجهر المقلوب للتصوير الفلوري المزدوج اللون. بدون نقص العمومية ، يمكن تطبيق هذا البروتوكول مع أي نظام OTs ديناميكي مجهز بقياسات القوة المباشرة بناء على اكتشاف زخم الضوء. تتوفر إجراءات مفصلة خطوة بخطوة لإنشاء مصائد تدرج بصرية محلية الصنع للتطبيقات في الجسم الحي50. تلك القائمة على تعديل AOD تبرز للتجارب النهائية مع الفخاخ المتعددة والقياسات السريعة51,52. توجد العديد من البروتوكولات لبناء أداة قائمة على زخم الضوء في الأدبيات36,39,40,53 ، ويمكن استخدام أي طريقة تصوير أخرى (تباين التداخل التفاضلي ، التألق الواسع ، إلخ).
- بدء تشغيل الملقط البصري
- من أجل تحسين استقرار طاقة الإخراج، قم بتشغيل الليزر بطاقة عالية إلى حد كبير (على سبيل المثال، 3 واط) قبل 30 دقيقة على الأقل من التجربة.
- قم بتشغيل وحدة الإلكترونيات الخاصة بوحدات المعالجة الدقيقة البصرية وقياس القوة.
ملاحظة: تطبيق جميع تدابير السلامة بالليزر واستخدام المعدات المعتمدة من قبل مجلس إدارة المؤسسة فقط. لا تستخدم أبدا عدسات المجهر الضوئي عندما يكون الليزر قيد التشغيل. استخدم دائما نظارات حماية الأشعة تحت الحمراء المعتمدة (OD7 في نطاق 950-1080 نانومتر) ، وقم بحظر ضوء ليزر الأشعة تحت الحمراء باستخدام الغالق في منفذ التألق 2 ، ولا تنفذ برنامج الملائمة البصرية حتى الانتهاء من محاذاة مستشعر القوة البصرية بعد الخطوة 5.3. بشكل عام ، لا تستخدم عينة عاكسة للغاية ، لأن الانعكاس الخلفي يمكن أن يسبب تلفا لليزر. - تحكم في طاقة المصيدة باستخدام HWP الدوار (الشكل 2F) عند مدخل وحدة المعالجة الدقيقة البصرية.
ملاحظة: تتضمن وحدة المعالجة الدقيقة البصرية التجارية المستخدمة في هذا البروتوكول بالفعل هذه الميزة. بالنسبة لأنظمة الاصطياد البصري محلية الصنع ، قم بدمج هذه الأداة للتحكم في الطاقة بحيث يمكن استخدام قوى ليزر أعلى وأكثر استقرارا.
- استخدام غرفة صغيرة فارغة للمعايرة
- اقطع مربعا بحجم 1 × 1 سم على شريط سكوتش على الوجهين وقم بإرفاقه بشريحة مجهر بسماكة 1 مم.
- أضف الماء إلى المربع وأغلقه من الأعلى باستخدام زجاج غطاء # 1.5 (22 × 22 مم). ينصح بإضافة كمية أكبر قليلا من الماء ، على سبيل المثال ، 30-40 ميكرولتر لتجنب الفقاعات داخل الغرفة المغطاة. امسح غرفة المعايرة برفق في حالة انسكاب الماء منها.
- محاذاة مستشعر القوة البصرية
- ضع قطرة من الماء على هدف الغمر بالماء 60x / 1.2. ضع غرفة المعايرة على المسرح مع زجاج الغطاء رقم 1.5 الذي يواجه الهدف. ركز على السطح السفلي ، حيث ستكون عينات الخلية في النهاية.
- أضف قطرة من زيت الغمر فوق الشريحة الزجاجية العلوية التي تغطي العينة (الشكل 2D). اخفض عدسة التجميع الخاصة بوحدة مستشعر القوة بعناية حتى تتلامس مع قطرة الزيت.
ملاحظة: يجب أن تكون القطرة كبيرة بما يكفي بحيث تغطي العدسة بأكملها التي تجمع ضوء الليزر الخارج من الفخاخ. عادة ، 200 ميكرولتر يكفي لتغطية السطح بأكمله وتوفير اتصال غمر مستقر. كن متحفظا وتجنب الملء الزائد لأنه قد يتسرب إلى العينة. - باتباع بروتوكول الشركة المصنعة لمحاذاة مستشعر القوة البصرية، انظر إلى صورة مستوى العينة على الكاميرا الإضافية التي سيتم استخدامها لوضع OTs (AUX، الشكل 2F). بلطف شديد، اخفض مستشعر القوة البصرية حتى يظهر توقف المجال (FS، الشكل 2F-G) مقترنا بمستوى العينة. وسيضمن ذلك إجراء قياسات مناسبة للقوة المباشرة من خلال الكشف الثابت للعينات عن تغيرات زخم الضوء40.
ملاحظة: أغلق FS بما فيه الكفاية بحيث تصبح صورته أصغر من مجال الرؤية (FOV)، وبالتالي، مرئية. كن حذرا للغاية ولا تدفع عدسة التجميع الخاصة بمستشعر القوة البصرية ضد العينة. يمكن بدلا من ذلك تحديد الموضع الرأسي لمستشعر القوة البصرية من تحليل توزيع ضوء الملائمة في BFP للمخاريط الضوئية ذات الفتحة العددية المحددة (NA). - تأكد من عدم وجود فقاعات هواء في قطرة الزيت ؛ هذه يمكن أن تؤثر بشكل مباشر على قياسات القوة. للتحقق من وجود فقاعات هواء، ضع عدسة برتراند في مكانها (BL، الشكل 2G) وراقب مسار التصوير من خلال العدسة. إذا كانت أي أوساخ أو فقاعات هواء مرئية أو كانت هناك حاجة إلى مزيد من الزيت (الشكل S1A)، فقم بتنظيف العدسة والغرفة بأنسجة عدسة خالية من الغبار وكرر الإجراء في الخطوتين 5-3-2 و5-3-3. يصور الشكل S1B مسارا بصريا دون عائق.
- باستخدام البراغي الجانبية الموضوعة على حامل مستشعر القوة البصرية ، قم بتوسيط FS في FOV. للحصول على الدقة، افتح FS بحيث تملأ FOV المرئي على الكاميرا الإضافية تقريبا (AUX، الشكل 2F).
6. تحسين الملقط البصري
ملاحظة: يعتمد قياس القوة المباشرة فقط على تغير زخم الضوء الناشئ عن القوة المؤثرة على الجسيم المحاصر، وبالتالي، على النقيض من الطرق غير المباشرة، لا يلزم معايرة صلابة المصيدة قبل كل تجربة. تتم معايرة التحويل الخاص بالأداة لعامل الانحراف / القوة (α ؛ pN / V ، المرجع 41) من قبل الشركة المصنعة وبالتالي فهو ثابت للتجربة. ومع ذلك ، نظرا لأن بقعة الليزر يتم التلاعب بها على مساحة 70 ميكرومتر × 70 ميكرومتر ، فإن الخطوات 6.2-6.5 ضرورية لضمان الملائمة المثلى واستقرار الطاقة. يتم توفير الخطوات التالية في برنامج الشركة المصنعة بحيث يتم تحسين OTs على منطقة العمل بطريقة شبه تلقائية.
- قم بتشغيل برنامج OTs وبرنامج الاستحواذ على كاميرا AUX.
- اطرح خط الأساس الأولي للجهد بالنقر فوق الخطوة 1: خطوة إزاحة الإلكترونيات في القائمة الفرعية لمعايرة النظام في برنامج قيادة الملقط البصري.
- لإجراء تسطيح طاقة المصيدة عبر منطقة عمل OT، اضبط طاقة المصيدة على نصف الحد الأقصى لها عن طريق تدوير HWP وفقا لذلك. لا تقم بتغيير طاقة المصيدة عن طريق تغيير خرج الليزر ، ولكن باستخدام HWP الدوار (الشكل 2F). انقر فوق الخطوة 2: الطاقة لبدء الروتين التلقائي لتسطيح طاقة الملائمة.
ملاحظة: هذه خطوة حاسمة للتعويض عن تباين طاقة الملائمة عبر منطقة عمل OTs (الشكل S1D). يؤدي الروتين الناجح إلى انخفاض تباين طاقة المصيدة إلى 2٪ عبر منطقة عمل OTs ويتقارب بعد دقيقتين. - لإجراء معايرة موضع الملائمة، قم بإزالة مرشح الأشعة تحت الحمراء بحيث يكون الضوء المنبعث من الليزر مرئيا على الكاميرا. أوجد بقعة الأشعة تحت الحمراء عن طريق ضبط مستوى الصورة مركزا على السطح السفلي للغرفة الصغيرة. احصل على أصغر بقعة ممكنة للأشعة تحت الحمراء عن طريق ضبط مستوى الصورة (الموضع الموضوعي) وتباين الرسم البياني في برنامج اكتساب AUX للكاميرا. إذا لزم الأمر ، قلل من طاقة المصيدة البصرية عن طريق تدوير HWP (الشكل 2F). انقر فوق الخطوة 3: الموضع لبدء معايرة تحديد المواقع الروتينية أو الملائمة.
ملاحظة: يتيح هذا الروتين التطابق الدقيق لإحداثيات موضع OT في كاميرا AUX مع زوايا توجيه AOD. يقوم الروتين الناجح بإنشاء رسم الخرائط من زاوية إلى موضع في عدة ثوان. - تعويض الزخم الأولي
ملاحظة: تؤدي حركة المصيدة البصرية عبر العينة إلى اختلافات في توزيع زخم الضوء عند BFP (الشكل S1E، F). يؤدي ذلك إلى تغييرات في الإشارة المستقلة عن القوة المتعلقة بموضع الليزر فوق منطقة العمل ، على الرغم من أن قوة المصيدة قد تم تسطيحها كما في الخطوة 6.3. والنتيجة هي اختلاف في خط الأساس للقوة بسبب الموضع (بغض النظر عن قوة فعلية تؤثر على الخرزة المحاصرة بصريا) التي تحتاج إلى تصحيح قبل كل تجربة.- اضبط قوة المصيدة التي سيتم استخدامها في التجارب ، عن طريق تدوير HWP (الشكل 2F).
- انقر على خيار الإزاحة العالمية في القائمة الفرعية الأدوات. سيؤدي ذلك إلى فتح مساعد Offset Cancel الخاص ببرنامج الملقط البصري الذي يصحح خط الأساس الأولي للزخم.
- انقر على | الإزاحة تعويض لتصحيح الزخم الأولي المتغير للموضع.
ملاحظة: إذا لم يؤثر أي تعديل على المسار البصري خلال الأسابيع الجارية، فستظل خرائط تسطيح طاقة الملائمة (الخطوة 6.3) والموضع (الخطوة 6.4) ثابتة. لذلك نوصي دائما باستخدام نفس المزيج من العناصر البصرية (المرايا ثنائية اللون ، والمرشحات ، وما إلى ذلك) التي قد تؤثر على مسار مصيدة الليزر أو لتنفيذ روتين جديد لتسطيح قوة المصيدة. فيما يتعلق بتعويض الزخم الأولي (الخطوة 6.5) ، توفر الشركة المصنعة لمنصة OTs معايرة فورية يجب تغييرها لكل قوة محاصرة جديدة وجلسة تجريبية. يجب تنفيذ الخطوتين 6.3 و6.4 على شريحة المعايرة الفارغة الموضحة في الخطوة 5.2. في عينة تحتوي على خلايا أو كائنات أخرى، يجب تنفيذ الخطوة 6.5 خالية من الكائنات التي قد تغير تشتت الضوء في منطقة عمل OTs.
- اختياريا، قم بحبس المجهر وحرك المصيدة بسرعة معروفة أثناء تسجيل إشارة القوة. على سبيل المثال ، اضبط المصيدة لإجراء تذبذب مثلث: ستكون إشارة القوة المسجلة إشارة مربعة.
ملاحظة: يجب أن تزداد قيمة القوة خطيا مع السرعة، وفقا لقوة السحب المؤثرة على الخرزة. يعمل هذا الاختبار كعنصر تحكم إيجابي في أن قياسات القوة يتم إجراؤها بشكل صحيح38. وبدلا من ذلك، يمكن استخدام مستشعر القوة البصرية للحصول على صلابة الملائمة البصرية، κ [pN/μm]، وعامل معايرة الموضع، β [μm/V]، من التحليل الطيفي للقدرة35. تحت المحاذاة الصحيحة ، يكون عامل المعايرة الثابت الذي توفره الشركة المصنعة هو α = κ· β [pN / V].- ابدأ قراءة القوة في الوقت الفعلي بالنقر فوق Plot 1 في القائمة الفرعية التدابير في برنامج الشركة المصنعة. سيوفر هذا قراءة لقوة وقوة الاصطياد البصري الحالية.
- افتح مربع الحوار معلمات التذبذب من القائمة الفرعية أدوات . قم بتعيين شكل موجي ثلاثي الفضاء في حلقتي محدد الشكل والنوع، على التوالي. على سبيل المثال ، اضبط سعة 10 ميكرومتر وتردد 3 هرتز. سيؤدي ذلك إلى قوة لزجة تبلغ حوالي 1 pN على ميكروبيد يبلغ قطره 1 ميكرومتر 38.
- في نافذة AUX الخاصة بالكاميرا، انقر بزر الماوس الأيمن فوق الميكروبيد وحدد بدء التذبذب. ستصبح قراءة القوة إشارة قوة مربعة مع هضاب عند ±1 pN.
- انقر بزر الماوس الأيمن على الميكروبيد وحدد إيقاف التذبذب.
7. المجهر البؤري القرص الغزل
- قم بتشغيل المجهر البؤري للقرص الدوار ومعدات الملحقات ومحركات الليزر المدمجة وكاميرات الاقتناء.
- قم بتشغيل برنامج التصوير.
- تعيين قنوات التصوير لتلطيخ Hoechst للنواة و GFP لغشاء البلازما الخلوي.
- قم بتنشيط خطوط ليزر الإثارة 405 نانومتر و 488 نانومتر.
- أضف ثنائي اللون متعدد النطاقات ليعكس الإثارة إلى العينة ويسمح للضوء المنبعث بالمرور إلى الكاميرات.
- قسم انبعاث التألق باستخدام مرآة ثنائية اللون ذات حافة تمرير طولها 500 نانومتر.
- استخدم مرشحات الانبعاثات DAPI/BFP (~ 445 نانومتر) وGFP (~ 521 نانومتر) أمام كاميرتي الاقتناء، على التوالي. ارجع إلى الشكل 2F,G.
- اضبط وقت التعرض الضوئي على 100 مللي ثانية لكل قناة.
- اضبط انبعاث الليزر للحصول على طاقة قدرها 5 ميجاوات في مستوى العينة. لقياس الطاقة، استخدم عداد طاقة تجاري.
- قم بتعيين بروتوكول التصوير. لتجنب النزيف الطيفي من قناة Hoechst إلى قناة GFP ، يجب تصوير الصبغتين بالتتابع.
ملاحظة: في حالة وجود مزامنة للأجهزة بين AODs الخاصة بالمصيدة البصرية واكتساب الكاميرا، تأكد من إعداد قطبية المشغل بشكل صحيح. إذا كنت في شك ، فاستشر مدير منشأتك أو الشركة المصنعة للمجهر.
8. إجراء تجارب المسافة البادئة للنواة
ملاحظة: قم دائما بإيقاف تشغيل المصائد البصرية - سواء باستخدام البرامج أو إغلاق الغالق على منفذ إبيفلورسنشن 2 - عند رفع وحدة مستشعر القوة وتغيير العينة. إذا لم يكن الأمر كذلك ، فقد يحدث ضرر جسيم للعناصر البصرية والمجرب. كن حذرا مع المسافة الجانبية بين حامل العدسة وحافة الطبق السفلية عند البحث عن الخلايا لتجنب اصطدام العدسة بطبق المرحلة / الثقافة (الشكل 2).
- ضع العينة في المجهر واتبع الخطوة 5.3 من هذا البروتوكول.
- باستخدام HWP الدوار (الشكل 2F) ، اضبط طاقة المصيدة على 200 mW كقيمة بداية إذا لم تكن صلابة النواة أو البنية داخل الخلايا التي تم فحصها معروفة. ترجمة منطقة عمل OTs (باستخدام مرحلة المجهر) إلى مكان خال من الخلايا من أجل التعويض عن خط الأساس الأولي للزخم من خلال الخطوة 6.5.
ملاحظة: اعتمادا على صلابة البنية دون الخلوية، يجب ضبط قيمة طاقة الملائمة إلى قيم أقل أو أعلى للحصول على عمق مسافة بادئة مماثل. - باستخدام وحدة التحكم في برنامج مرحلة المجهر ، ابحث عن خلية بها حبة واحدة أو حبتان من خلال الفحص المجهري الساطع المنقول (الشكل 3A).
- حدد مسار الملائمة.
- افتح مربع حوار المسار في القائمة الفرعية أدوات واختر الإزاحة في حلقة محدد نوع المسار .
- في الورقة العددية ، اكتب الإزاحة والوقت لكل خطوة مسار لاحقة. فيما يلي مثالان.
- بالنسبة لتجربة استرخاء الإجهاد ، قم ببرمجة الأحمال شبه المنحرفة ، كما هو موضح في الشكل 3B. وفي الجدول دال-1، طبقت مسافة بادئة شبه منحرفة بمسافة سفر قدرها 5 ميكرومتر؛ سرعة 5 ميكرومتر / ثانية ؛ وقت الانتظار قبل التراجع: 10 ثانية.
- بالنسبة لتجربة المسافة البادئة المتكررة بسرعة ثابتة للحصول على روتين مثلث دون وقت سكن على النواة ، اضبط سعة المسار ، على سبيل المثال ، 5 ميكرومتر ، ووقت الخطوة ، على سبيل المثال ، 2 s لسرعة 2.5 ميكرومتر / ثانية. في الجدول S2 ، يتم تطبيق هذا ثماني مرات بنفس السرعة.
ملاحظة: يجب تحديد هذه القيم لكل نوع خلية وتجربة، ولكن المعلمات التالية للروتين شبه المنحرف تلتقط أهم الديناميكيات في التجربة المعروضة هنا. يجب أن يكون وقت الانتظار كافيا حتى تظهر النواة استرخاء الإجهاد الكامل بعد المسافة البادئة
- محاصرة المجهر
- اضبط مستوى الصورة فوق الخرزة قليلا باستخدام وحدة التحكم في برنامج مرحلة المجهر.
- قم بتنشيط الفخاخ باستخدام برنامج OTs وانقر على الخرزة في نافذة تصوير AUX للكاميرا (تمت معايرتها باتباع الخطوة 6.4). الحبس الناجح للحبة بواسطة المصيدة البصرية سيقلل بشدة من حركة الخرزة.
- انقر واسحب الخرزة عبر السيتوبلازم وضعها على مسافة ~ 2 ميكرومتر من الغلاف النووي (الشكل 3A). تأكد من تعيين المسار بحيث تكون المسافة البادئة للحرزة عمودية على الغشاء النووي.
- اختياريا، إذا لزم الأمر لقياسات موضع الخرزة بالنسبة إلى المصيدة، قم بمسح المصيدة ضوئيا عبر الخرزة لتحديد صلابة الملائمة، k [pN/μm]54، وبالتالي Δxbead = -F/k (انظر المناقشة). تحتوي وحدة المعالجة الدقيقة البصرية المستخدمة في هذا البروتوكول على روتين مدمج لهذا الغرض.
- افتح مربع الحوار مسح الجسيمات في القائمة الفرعية أدوات .
- حدد الملائمة التي تريد مسحها ضوئيا والتردد العالي كطريقة المسح الضوئي. حدد الاتجاه (x أو y) لمسار المسافة البادئة لقياس مسح الخرزة.
- ستظهر نافذة مع قياس صلابة الملائمة. في الرسم البياني، اسحب المؤشرين لتحديد مساحة الملائمة الخطية المقابلة ل F = -kx. سيتم تحديث الملاءمة الخطية لجزء البيانات المحدد تلقائيا.
ملاحظة: اضبط الموضع الأولي للحبة بعيدا عن غشاء الخلية (~ 5 ميكرومتر)، حيث تؤثر انحرافات زخم الضوء في واجهة الخلية المتوسطة على مدى ملاءمة قياسات القوة. إذا كانت النواة قريبة جدا من غشاء الخلية، فحاول وضع مسافة بادئة للنواة من الموقع المقابل. تجاهل الخلية إن لم يكن ذلك ممكنا.
- ابدأ في الحصول على الصور بالنقر فوق زر الاقتناء في برنامج التصوير.
- ابدأ في توفير بيانات موضع الملائمة وقوة قياس البيانات بالنقر فوق | البيانات حفظ في نافذة قراءة القوة في الوقت الحقيقي (مفتوحة كما في الخطوة 6.6.1).
ملاحظة: تم تجهيز المصيدة البصرية بإدخال مشغل يمكن توصيله بمخرج توقيت الكاميرا. وبالتالي ، فإن بيانات الصورة والقوة متزامنة مع الأجهزة والإلكترونية قادرة على تعيين دورات المصيدة مع عدد إطارات الصور أثناء الاقتناء. - ابدأ المسار الذي تم تحميله مسبقا بالنقر بزر الماوس الأيمن فوق الخرزة وتحديد مسار البدء.
- انتظر حتى يتم الانتهاء من المسار ويستقر النظام.
- إيقاف حفظ بيانات قياس قوة الملائمة. سيظهر مربع حوار لتوفير البيانات.
ملاحظة: لتحسين تخزين البيانات، يمكن تدمير البيانات عن طريق تحديد المعلمة التدميرية في مربع الحوار هذا (10 أو 100 أو 1000). - توقف عن الحصول على الصور ورسم النتائج في برنامج المعالجة اللاحقة الذي يختاره المستخدم.
- إذا فقدت الميكروسفير أثناء الروتين ولا يمكن وضع مسافة بادئة للنواة (الشكل S2) ، فتجاهل القياس وقم بزيادة الطاقة. لاحظ أنه يجب تكرار الخطوة 6.5. في أيدينا ، يتم الانتهاء من 95٪ على الأقل من الروتين بنجاح دون أن تفقد الخرزة من الفخ.
النتائج
الحقن المجهري لخرز الاصطياد:
تنتشر الميكروسفيرات التي يتم حقنها في جنين الزرد أحادي الخلية على الغطاء الحيواني بأكمله أثناء التشكل. للحصول على تصور أكثر وضوحا ، كررنا بروتوكول الحقن باستخدام الميكروبيدات الفلورية الحمراء والتقطنا صورا حجمية باستخدام المجهر البؤري في مراحل النمو المختلفة. في الشكل 4A-D ، يتم تصور الخرز المحقون في سيتوبلازم الخلايا الجذعية السلفية في الجسم الحي عند 5 hfp. في وقت لاحق ، ظهرت الميكروسفير منتشرة على الجنين بأكمله بسرعة 24 حصانا (الشكل 4E). تطورت الأجنة في كلتا المرحلتين بشكل طبيعي وكانت معدلات البقاء على قيد الحياة قابلة للمقارنة مع الأجنة غير المحقونة أو المحقونة الوهمية (انظر الشكل S3). وهذا يتفق مع الدراسات الأخرى التي تفيد ببقاء سمك الزرد عن طريق الحقن بالخرز دون إزعاج لمدة تصل إلى 5 أيام بعد الإخصاب55.
يتوافق المجهر البؤري للقرص الدوار مع المجهر الفلوري متعدد القنوات. في الشكل 5A ، نظهر خلايا جذعية معزولة مع حبة واحدة أو اثنتين في السيتوبلازم. يمكن استخدام ملصقات فلورسنت متعددة للتحقيق في جوانب مختلفة من الخلية (الشكل 5B). يمكن تتبع المورفولوجيا النووية باستخدام صبغة Hoechst أو باستخدام تعبير H2A::mCherry mRNA ، في حين يمكن تحليل الغشاء النووي الداخلي باستخدام Lap2b-eGFP12. يمكن ملاحظة ديناميكيات قشرة الأكتوميوسين ، وكذلك مستويات الكالسيوم داخل الخلايا ، مع خط My12.1::eGFP المعدل وراثيا56 وحضانة Calbryte-520 ، على التوالي. يهدف البروتوكول الذي تم وصفه هنا إلى مقارنة ميكانيكا نواة الخلية للخلايا البرية المجمدة على ركائز لاصقة (يشار إليها لاحقا باسم التعليق) وفي الحبس الميكانيكي. أظهرت الخلايا الجذعية المعزولة المحصورة في غرف صغيرة يبلغ ارتفاعها 10 ميكرومتر تكشفا جزئيا للغشاء النووي الداخلي (INM) وزيادة لاحقة في انقباض الأكتوميوسين12. في الشكل 5C ، تظهر الخلايا المحصورة مع حبة واحدة أو اثنتين في السيتوبلازم. سيكون الحبس الناجح مرئيا عبر خلايا مسطحة وموسعة مع مقطع عرضي أوسع من النواة. يتم الكشف عن الغشاء النووي بشكل أكبر في الخلايا المحصورة ويجب أن يبدو ناعما مقارنة بالخلايا المعلقة (الشكل 5C).
تحليل وقت القوة وتشوه القوة
يعتمد تحليل النتائج التي تم الحصول عليها بشدة على العينة التي تم التحقيق فيها ومسألة الاهتمام وبالتالي لا يمكن تعميمها هنا. على سبيل المثال، تتمثل إحدى الطرق الشائعة لتحليل قياس المسافة البادئة في استخراج معامل يونغ عن طريق تركيب نموذج هيرتز المعدل مع بيانات المسافة البادئة للقوة57. ومع ذلك ، فإن افتراض مثل هذا العلاج يحتاج إلى تقييم دقيق وقد لا يكون دائما مبررا بشكل صحيح (مثل أن تكون البنية التي تم التحقيق فيها متساوية الخواص ، متجانسة ، مع مرونة خطية ومسافات بادئة أصغر من نصف قطر الخرزة). وبالتالي فإننا ننظر فقط في القياسات المستقلة النموذجية هنا التي تسمح بمقارنة السلوك الميكانيكي للبنية التي تم التحقيق فيها بين السيناريوهات التجريبية المختلفة.
كنقطة بداية، يوفر قياس ميل منحنى إزاحة القوة عند عمق مسافة بادئة معينة مقياسا لنموذج صلابة هيكلية مستقلة58 للنواة. يمكن بعد ذلك جمع هذه القيمة من عينات متعددة ومقارنتها بين الإعدادات التجريبية المختلفة واضطرابات العينة.
قياس المسافة البادئة
في السطور التالية ، نركز على الاستجابة الميكانيكية لنواة الخلية أثناء تشوه الخلية في الحبس. عادة ما تؤدي التجارب في الخطوة 8 من هذا البروتوكول إلى قمم قوة تصل إلى 200 pN لأعماق المسافة البادئة التي تبلغ حوالي 2-3 ميكرومتر. ومع ذلك ، يمكن أن تكون هذه القيم مختلفة إلى حد كبير ، اعتمادا على نوع الخلية والظروف التجريبية ، مع نوى أكثر ليونة تؤدي إلى انخفاض القوة لمسافة بادئة معينة. وبالتالي هناك حاجة إلى قياس التشوه النووي بدقة ، جنبا إلى جنب مع القوة ، من أجل توصيف ميكانيكي دقيق لنواة الخلية. في هذا القسم ، سنحصل على الصلابة النووية للخلية من قياسات المسافة البادئة للقوة التمثيلية.
في الشكل 6، نبين تشوهات الجانبين البعيد والقريب من النواة في خلية معلقة ومحصورة. يمكن ملاحظة سلوك ميكانيكي غني. في خلية معلقة نموذجية على ركيزة لاصقة ، كانت النواة بادئة بقوة بواسطة الخرزة ، ولكنها أيضا نزحت قليلا عند أحداث الدفع المتكررة. قمنا بقياس المسافة البادئة للحبة على النواة من خلال تحليل الكيموغراف الذي تم الحصول عليه من التصوير الفلوري لنوى الخلايا الملطخة ب Hoechst. تم حساب Kymographs بسهولة باستخدام المكون الإضافي Multi Kymograph في فيجي على طول اتجاه المسافة البادئة (الشكل 6A ، B) وتم استيراده إلى Matlab (الإصدار 2021 ، Mathworks) لمزيد من المعالجة. تم تركيب وظيفة خطوة على ملف تعريف الكثافة الخام بهدف تتبع الحواف المحددة للنواة على طول مسار روتين المسافة البادئة. وكما يمكن أن نرى، فإنه يحمل معلومات دقيقة عن التغير النووي في الشكل (الشكل 6 والشكل S2). استخدمنا المنحنى السيني المزدوج التالي كنسخة تحليلية لدالة الخطوة:
 (المعادلة 1)
(المعادلة 1)
هنا ، يشير x1 و x2 إلى الحواف البعيدة والقريبة للنواة ، في حين أن A و B هما القيمتان الرماديتان القصوى والخلفية للقناة الزرقاء (صبغة Hoechst) للصورة (الشكل 6B). تم النظر في عرض الحافة (e0 = 0.25 مم). في حين أن حافة النواة القريبة (x2) البادئة تتبع المسار المطبق بواسطة روتين المصيدة البصرية بعد ملامسة المجهر والنواة، فإن الحافة البعيدة المعاكسة (x1) تعرض ديناميكيات الاسترخاء كما هو متوقع لمادة لزجة مرنة مثل السيتوبلازم (الشكل 6D). وعلى النقيض من ذلك، فإن النوى في الخلايا المحصورة في غرف صغيرة بارتفاع 10 ميكرومتر لا تظهر مثل هذا السلوك الانتقالي للنواة عند المسافة البادئة داخل الخلية (الشكل 6B، D). كما هو موضح في الشكل 6D ، تظل الحواف الخلفية للنوى دون تغيير بسبب دفع الخرزة من الجانب القريب ، على الأرجح بسبب القوى الأقوى الناشئة عن انقباض الخلية والاحتكاك الذي يعمل ضد قوة المسافة البادئة. من أجل الحصول على عمق التشوه الصحيح ، تم طرح الإزاحة x1 من المقياس البادئ x2: Δx = x2 - x1 (انظر أيضا الشكل 6D).
تحليل بيانات القوة
تم قياس القوة المسببة للتشوه النووي من التغير في زخم الضوء الذي نشأ في الميكروبيد المحبوس بصريا (الشكل 7A). زادت القوة عند تطبيق مسارات شبه منحرف (الخطوة 8.4.3 ، الشكل 7B) في البداية خطيا حتى توقف المصيدة عن الحركة ، ولكن بعد ذلك ارتخت إلى قيمة حالة ثابتة. أشار هذا السلوك إلى وجود مادة لزجة مرنة تظهر معامل الفقد والتخزين. مباشرة بعد حدث المسافة البادئة ، وصلت القوة إلى قيمة الذروة ، Fp ، تليها استرخاء الإجهاد (الشكل 7C):
 (المعادلة 2)
(المعادلة 2)
حيث F0 هي القوة المخزنة للمكون المرن و f(t) هي دالة استرخاء بدون أبعاد. لقد قمنا بتحليل هذا السلوك بثلاث طرق:
1. النظر في مادة صلبة خطية قياسية مع استرخاء إجهاد أسي ، أي f(t) = e-t / τ ، ممثلة تخطيطيا في الشكل 7C inset.
2. استخدام الاضمحلال العام الأسي المزدوج:
F(t) = A + B1e-t/τ1 + B2e-t/τ2.
3. استخدام قانون القدرة متبوعا باضمحلال أسي59:
f(t) = t-pe-t/τ، مثبتة في الشكل 7C.
في حين يمكن تنفيذ ملاءمة النموذج 1 بشكل مباشر ، نوصي بتقدير التخمينات الأولية ل (τ1 ، τ2) و (p ، τ) للنموذجين 2 و 3 ، على التوالي. ويمكن القيام بذلك، على التوالي، عن طريق تركيب خطوط على البيانات في المقاييس اللوغاريتمية مقابل الخطية (الشكل 7D، اليسار) واللوغاريتمية مقابل اللوغاريتمية (الشكل 7D، اليمين). يلخص الجدول S3 نتائج المثال الذي تم تحليله في الشكل 7. في القسم التالي، سننظر في الجمع بين قانون القدرة والقانون الأسي لتوصيف ميكانيكا نواة الخلية.
علاقة إزاحة القوة
وبالمثل ، يمكن استخدام الإعداد التجريبي الموصوف للحصول على علاقة إزاحة القوة لأحداث المسافة البادئة المتعددة. من خلال تنفيذ الإجراءات الروتينية الثلاثية (الخطوة 8.4.4 ، الشكل 8A) ، من الممكن ربط القوة بالتشوه ورسم منحنى المسافة البادئة للقوة. ويظهر الشكل 8 باء نتيجة مثالية، حيث يتغير الميل بسلاسة إلى خط أساس مسطح بمجرد ملامسة الخرزة للنواة. ويمثل تحديد نقطة الاتصال الحقيقية في البيانات الصاخبة تحديا، ويجب توخي الحذر لمعرفة ما إذا كانت منطقة التلامس مناسبة للنماذج المرنة60. في هذه التجربة بالذات، يمكن أيضا ملاحظة أن المسافات البادئة اللاحقة تؤدي إلى منحنيات ذات نقاط اتصال أعمق، مما يدل على انتعاش الشكل النووي البطيء للغاية بعد تراجع الخرزة وتغيير في الدورة الهستيرية المحددة بخصائص المواد اللزجة المرنة للنواة (الشكل 8C). وبالتالي ، يجب أن يكون الباحث على دراية بما إذا كان هذا يحدث وأن يدمج هذا في خط الأنابيب التحليلي ، أو يحد من عدد القياسات اللاحقة بحيث لا يعدل هذا التأثير القياس.
ميكانيكا النواة في الخلايا المعلقة وتحت 10 ميكرومتر الحبس
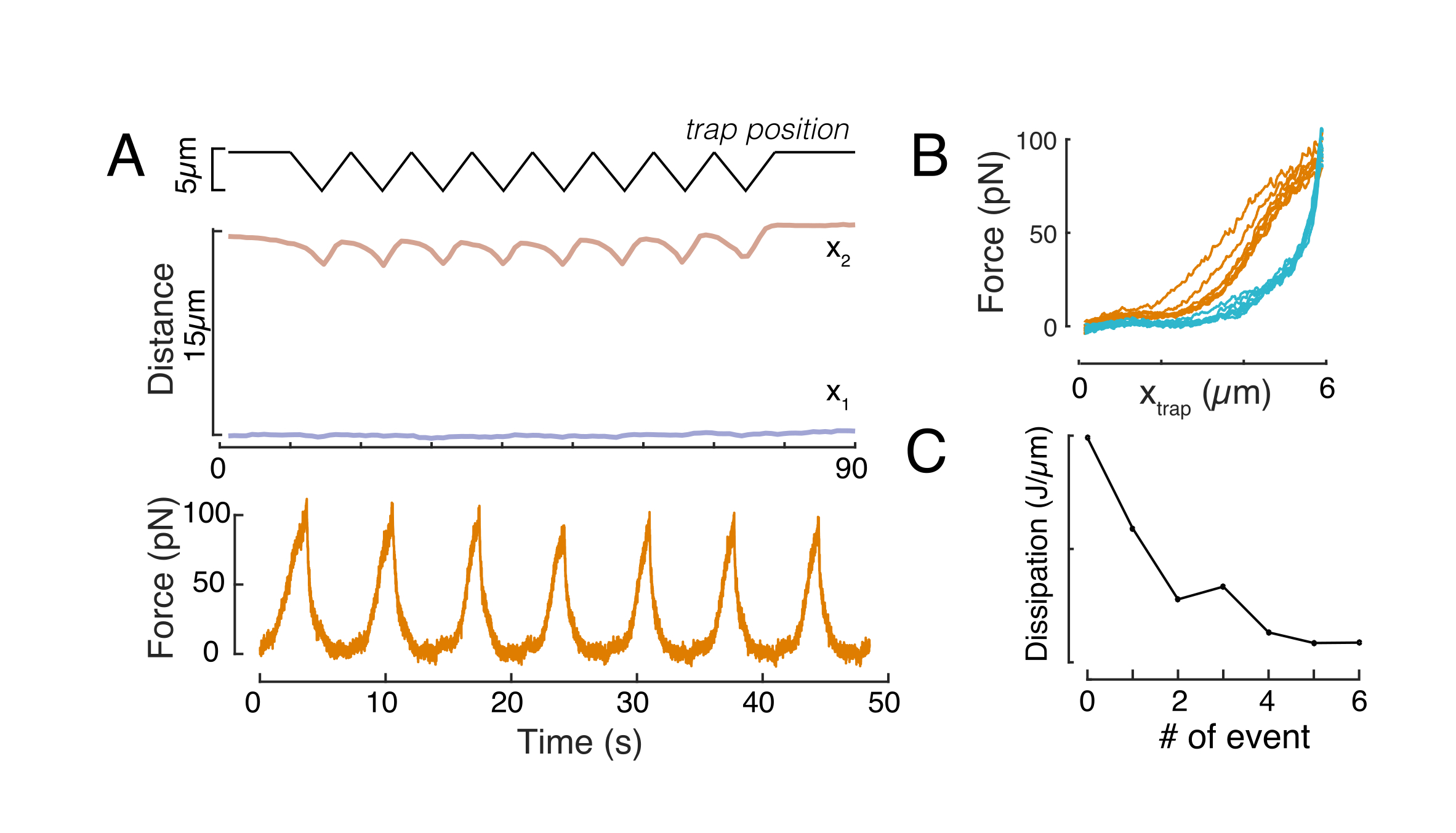
تم استخدام النهج المذكور أعلاه لتحليل ديناميكيات استرخاء إجهاد النواة في الخلايا المعلقة على ركائز لاصقة وخلايا محصورة. تظهر نتائجنا أن الحبس يؤدي إلى توسع في المساحة المتوقعة (الشكل 9 أ) ، ولكن تغير ضئيل في الصلابة النووية (الشكل 9 ب). قمنا بقياس استرخاء مماثل مع τ = 6.08 ± 1.1 s (غير محصور) و τ = 4.00 ± 0.6 s (الحبس) ، مما يشير إلى تبديد سريع للمرونة اللزجة ، تليها قيمة قوة مخزنة تتوافق مع معامل المرونة للنواة. من أجل حساب الاختلافات التجريبية ، والتي قد تنتجها ظروف أولية مختلفة في إجراءات المسافة البادئة ، تم تطبيع القوى المخزنة المقاسة إلى عمق المسافة البادئة ، مثل  . تمثل هذه المعلمة صلابة النواة وتصف القوة ، أو الإجهاد ، اللازمة لمبادئة معينة. حصلنا على صلابة مماثلة تحت الحبس وفي الخلايا غير المحصورة: = 20.1 ± 12.6 pN/ μm و = 24.6 ± 13.6 pN/μm (متوسط الانحراف المعياري ±)، على التوالي.
. تمثل هذه المعلمة صلابة النواة وتصف القوة ، أو الإجهاد ، اللازمة لمبادئة معينة. حصلنا على صلابة مماثلة تحت الحبس وفي الخلايا غير المحصورة: = 20.1 ± 12.6 pN/ μm و = 24.6 ± 13.6 pN/μm (متوسط الانحراف المعياري ±)، على التوالي.

الشكل 1: الحقن المجهري لأجنة الزرد في مرحلة الخلية الواحدة (الزيجوت). (أ) لوحة الحقن: يتم استخدام لوحة حقن مثلثة الشكل للحقن. اللوحة مصنوعة من 1٪ من الأغاروز فائق النقاء في E3 (وسط البيض). تظهر طرق العرض العلوية والجانبية على اليمين. (ب) تحديد موضع الجنين: قم بتوجيه الأجنة بلطف باستخدام فرشاة وتوجيهها بحيث تكون الخلية الواحدة مرئية بوضوح ويمكن الوصول إليها بسهولة باستخدام الإبرة. نقترح توجيه الأجنة مع الخلية الموجودة في الجانب الآخر من الإبرة ، كما هو موضح في الرسم. (ج) إجراء الحقن في الجنين في مرحلة الخلية الواحدة: اخترق المشيمة المحيطة بالجنين والخلية الواحدة بالإبرة. تأكد من أن طرف الإبرة داخل الخلية وحرر الضغط للحقن. (د) احتضان الأجنة عند 28-31 درجة مئوية حتى تتطور حتى مرحلة الأرومة (الكرة) (4 حصان). قم بإجراء بروتوكول عزل الخلايا وتلطيخ الخلايا (الخطوة 2) وقم بإعداد غرفة الاصطياد البصري مع الخلايا المعزولة في التعليق و / أو الحبس جنبا إلى جنب مع طلاء سطح الركيزة المقابل (الخطوة 3). يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 2: تحضير جهاز الملقط البصري. (أ) طبقات الطلاء الدوار من PDMS مع ارتفاع محدد على الأطباق ذات القاع الزجاجي. سوف ينتشر انخفاض PDMS بالتساوي بسبب قوة الطرد المركزي. (ب) إعداد غرفة العينة من طبقة PDMS. 1: قطع مربع مع مشرط ، 2: تغطية البئر الداخلية مع concanavalin A (ConA) ، وغسل وخلايا البذور. 3: تغطية مع شريحة زجاجية أو غطاء زلة لإغلاق البئر. (ج) صورة للقطع المربع بمشرط وإزالة PDMS جيدا بالملقط. (د) تركيب عدسة التجميع الخاصة بمستشعر القوة البصرية فوق غرفة الاصطياد. تعمل قطرة من زيت الغمر كوسيط غمر بين عدسة التجميع والغطاء الزجاجي العلوي. تخطيطي ليس على نطاق واسع. كن حذرا أثناء خفض عدسة التجميع حتى لا تلمس الغطاء الزجاجي لطبق العينة. (ه) صورة لوحدة كشف القوة الملامسة للعينة. (و) مخطط الإعداد التجريبي. تستخدم وحدة المعالجة الدقيقة البصرية شعاع ليزر موجي مستمر (5W ، λ = 1064 نانومتر) مع التحكم في الطاقة من خلال لوحة نصف موجة (HWP) وفاصل شعاع استقطاب (BS). بعد تعديله بزوج من AODs ، يقترن بمنفذ التألق العلوي للمجهر المقلوب. ثم ينعكس شعاع الليزر بواسطة مرآة ثنائية اللون قصيرة التمرير 950 نانومتر (IR-DM) ، مما يسمح بنفاذية الإثارة الفلورية والانبعاثات. يتم توجيه ليزر الاصطياد إلى المنفذ الخلفي للتألق في المجهر (البرج العلوي). يتم إنشاء OTs في المستوى البؤري لعدسة هدف الغمر بالماء (60x ، NA = 1.2). يتعرض مستشعر القوة البصرية بواسطة برج المجهر ويلتقط ضوء الليزر الخارج من OTs باستخدام عدسة عالية NA ، غمر الزيت. في الوقت نفسه ، يتيح مستشعر القوة إضاءة المجال الساطع. تقترن وحدة البؤرية للقرص الدوار بالمنفذ الأيسر. وهي مجهزة بمحركي ليزر مدمجين (ILE) يتحكمان في سبعة ليزر إثارة فلورية وكاميرتين sCMOS مضاءتين من الخلف ، مما يتيح التصوير الفلوروفوري المزدوج بالتوازي Abb: TI ، Transilluminator. خ م، توقف ميداني؛ AOD ، الانحراف acustooptical ؛ HWP ، لوحة نصف موجة ؛ CAM ، الكاميرا (G) صورة فوتوغرافية لمعدات الاصطياد البصري. تشير الدائرة الحمراء إلى عدسة برتراند ، والتي يمكن تحويلها إلى المسار البصري يدويا. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
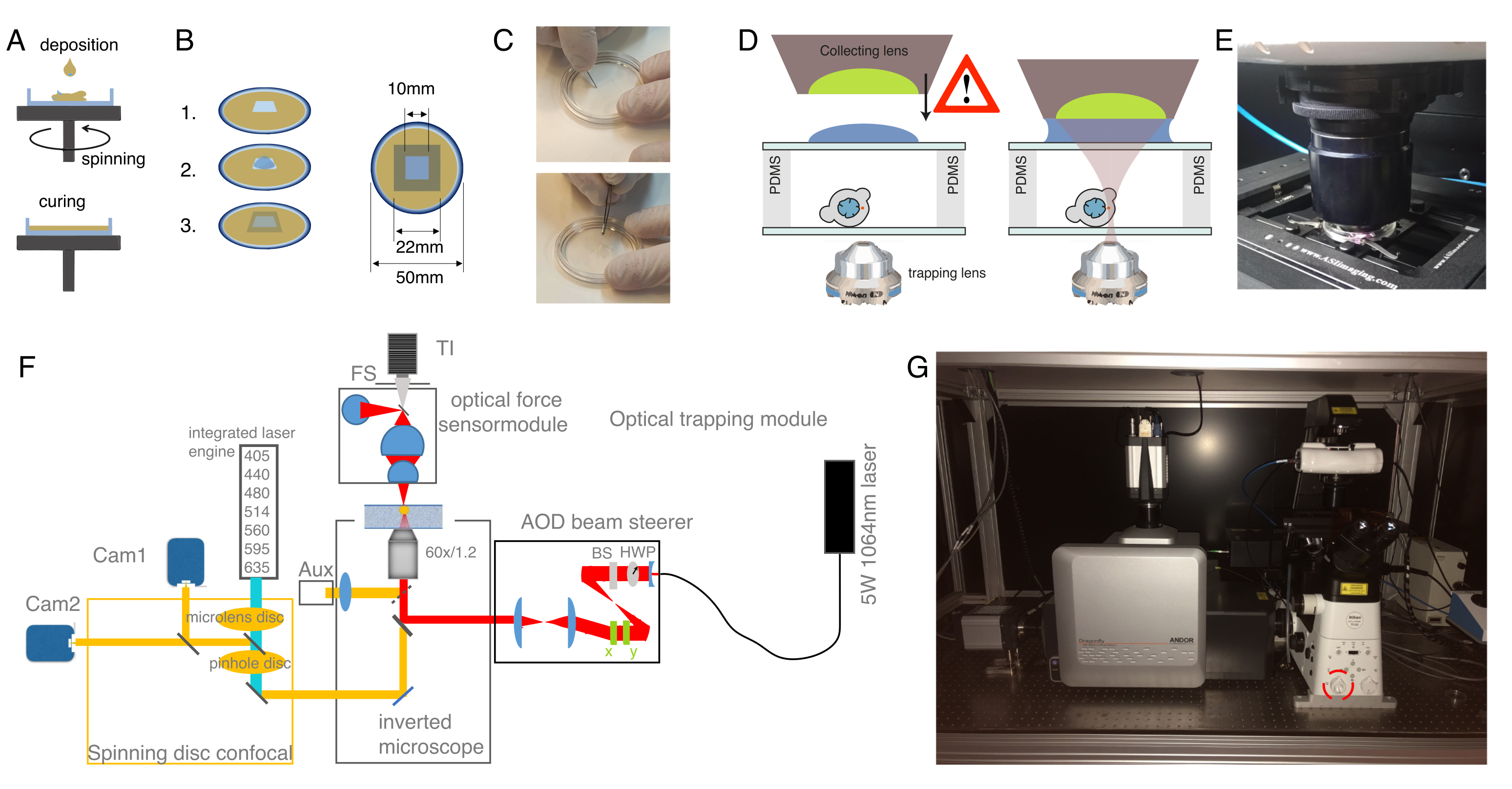{kind=link}

الشكل 3: اختيار العينات والمعلمات الصحيحة. (أ) صورة تمثيلية لخلية جذعية سلف الزرد المعزولة مع وجود ميكروسفير واحد بالقرب من النواة لإجراء تجربة المسافة البادئة. شريط المقياس = 10 ميكرومتر (ب) مسار المصيدة المثالي؛ عمق المسافة البادئة 5 ميكرومتر ؛ سرعة المسافة البادئة = 5 ميكرومتر / ثانية ؛ وقت الاسترخاء 10 ثانية يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
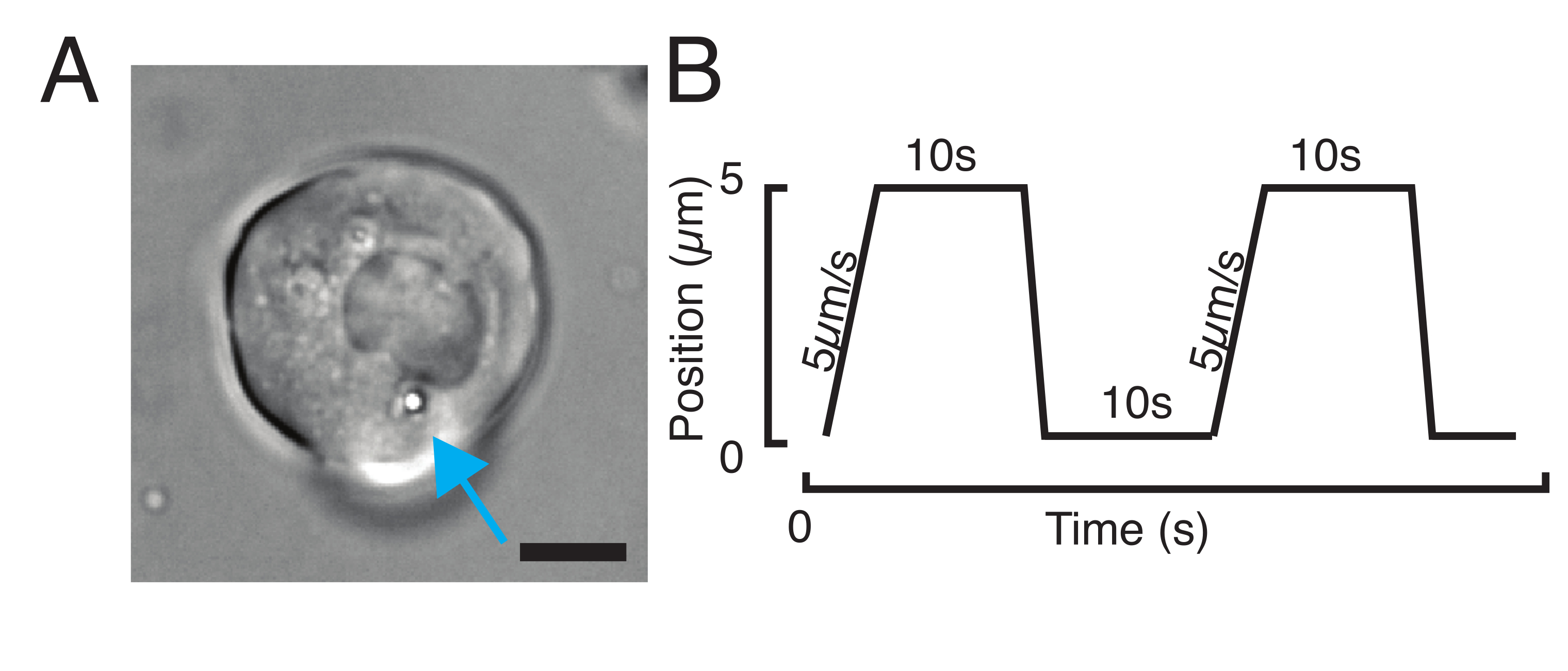{kind=link}

الشكل 4: توطين الميكروبيد داخل أجنة الزرد أثناء النمو. يتم حقن 0.5 نانولتر من حبات الفلورسنت الحمراء 1 ميكرومتر مع GPI-GFP mRNA (100 pg / embryo ، غشاء البلازما) في أجنة WT لتصور توطين الخرز. (ألف - دال) توزيع الميكروسفير 5 ساعات بعد الحقن داخل الجنين المركب في 0.75٪ من الأغاروز. (أ) صورة برايتفيلد والتألق. تنتشر الخرز بشكل متجانس عبر أنسجة الجنين كما هو موضح في المجهر البؤري. (ب) الإسقاط الأقصى للفلور البؤري البؤري z-stack. يتم ترميز الخرز بالألوان من الأرجواني إلى الأصفر وفقا لموضعها على شكل حرف z في مكدس الصور. يتوافق اللون الأرجواني / الأرجواني مع معظم الخرز / الخلايا الخارجية (EVL ؛ طبقة مغلفة ظهارية ؛ أو الخلايا الجذعية السلفية الموجودة بالقرب من سطح EVL) ، الأصفر يتوافق مع الخرز الداخلي (الخلايا العميقة السلف) ، كما هو موضح في الرسم على اليمين. (ج) قطع والحد الأقصى لإسقاط كومة فرعية من (B) المقابلة للمنطقة في المربع البرتقالي: جزء كبير من الخلايا العميقة يحتوي على 1-2 حبات. (د) قطع والحد الأقصى لإسقاط كومة فرعية من (B) المقابلة لمربع أرجواني: تحتوي بعض خلايا EVL على 1-2 حبة. (ه) صورة برايتفيلد والحد الأقصى لإسقاط كومة z لجنين بقوة 24 حصانا مثبتا في أغاروز بنسبة 0.75 في المائة ومخدرا بالتريكايين. تم تحضين الأجنة مسبقا باستخدام التريكايين لمدة 15 دقيقة. من اليسار إلى اليمين: الميكروسفير (قطرها 1 ميكرومتر) ، GPI-GFP وتداخل الصورة. الخرز موزعة على كامل جسم الجنين. البعد مقياس المشار إليه في كل لوحة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
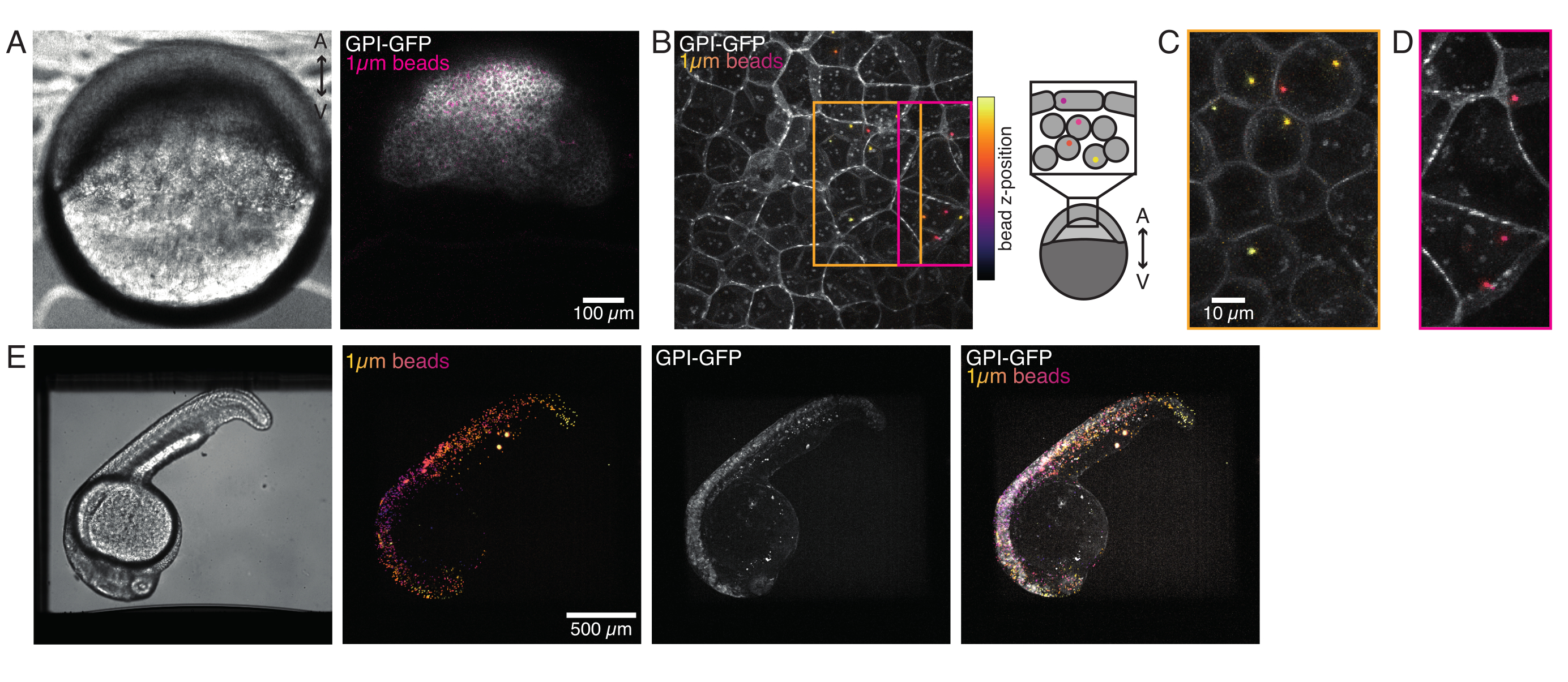{kind=link}

الشكل 5: الخلايا الجذعية السلفية المعزولة لسلف الزرد مع وضع علامات مختلفة. (أ) صورة المجهر الضوئي الناقل للخلايا المعلقة مع 1 (أعلى) أو 2 (أسفل) حبات حقن. الأسهم السماوية تشير إلى الخرز. (ب) صور الفلورسنت البؤرية للخلايا المعلقة ذات البقع المختلفة. أعلى اليسار: Lap2b-eGFP (الغشاء النووي الداخلي، 80 بيكوغرام/جنين) و H2A-mCherry. أعلى اليمين: GPI-GFP (غشاء البلازما ، 100 pg / الجنين) و DNA-Hoechst (ملطخ كما هو موضح في القسم 2). أسفل اليسار: MyI12.1-eGFP (الخط المعدل وراثيا) و DNA-Hoechst. أسفل اليمين: Calbryte488 و DNA-Hoechst (ملطخة كما هو موضح في القسم 2). (ج) صورة المجهر الضوئي الناقل للخلايا المحصورة مع 1 (أعلى) أو 2 (أسفل) حبات حقن. الأسهم السماوية تشير إلى الخرز. أشرطة المقياس = 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 6: تقدير التشوه النووي من أفلام القرص الدوار.(أ،ب) الفاصل الزمني لتجربة المسافة البادئة للنواة في (أ) خلية معلقة و (ب) خلية محصورة. شريط المقياس 10 ميكرومتر. يتم عرض لقطات تمثيلية لنوى تحمل علامة Hoechst قبل 5 ثوان وأثناءها و 5 ثوان بعد المسافة البادئة باستخدام ميكروسفير محاصر بصريا (رأس سهم أبيض). Kymographs على طول جزء المسافة البادئة (الخط الأحمر ، اللوحة اليمنى). x1 و x2 هما الحدود البعيدة والقريبة (بالقرب من الخرزة) للنواة أثناء تجربة المسافة البادئة المستخرجة من ملاءمة ملف تعريف الكثافة إلى المعادلة 1. (ج) ملامح الكثافة على طول مقطع المسافة البادئة لثلاثة إطارات مختلفة (قبل المسافة البادئة وأثناءها وبعدها) ومثبتة في المعادلة 1 لتقييم المواضع البعيدة ، x1 ، والقريبة ، x2 ، لحواف النواة. (د) المسارات التمثيلية ل x1 (t) باللون الأزرق و x2 (t) في العنبر أثناء تجربة المسافة البادئة للخلايا المعلقة والمحصورة (10 ميكرومتر). تشير المناطق المظللة إلى المسافة البادئة ، وتشير المسافة بين x1 و x2 إلى قطر النواة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
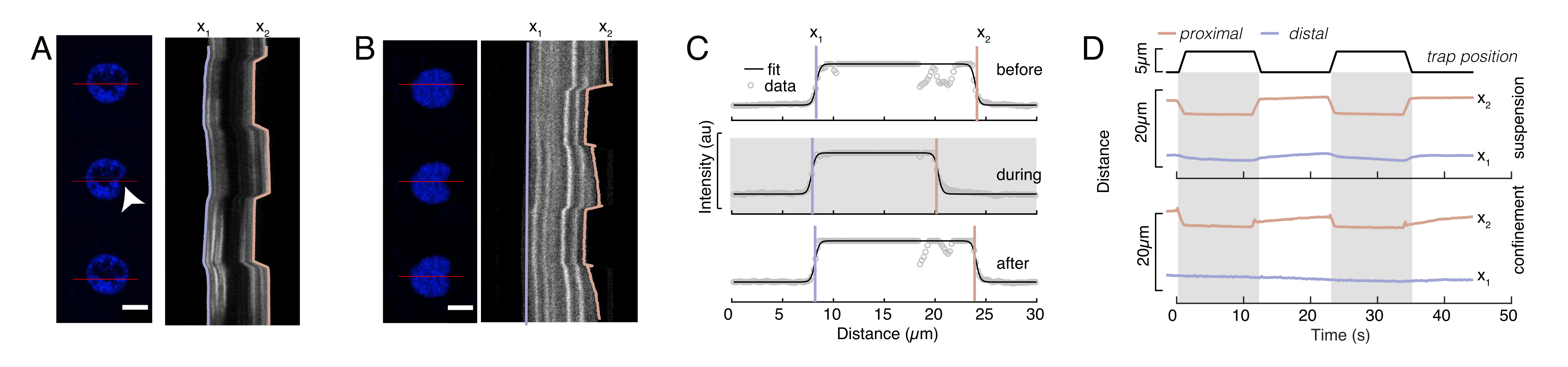{kind=link}

الشكل 7: معالجة إشارات القوة. (أ) مخطط لميكروسفير محاصر بصريا يشوه نواة الخلية عند المسافة البادئة. يشار إلى الغشاء النووي والقوى البصرية بواسطة الأسهم السوداء. يشار إلى التغير في زخم الحزمة بواسطة السهم الأخضر Pout. (ب) مسار المصيدة (أعلى) والقوة (أسفل) التي يعاني منها المجهر المحاصر بصريا أثناء تجربة المسافة البادئة النووية المتكررة. (ج) اضمحلال استرخاء القوة بعد ذروة القوة عند أقصى عمق المسافة البادئة. يظهر Inset مخططا للصلب الخطي القياسي الذي تقترب ديناميكياته من الملاحظات الظاهرية هنا. (د) اليسار: لوغاريتم القوة المعيارية مقابل الزمن. تشير المساحات المظللة إلى جزء البيانات المستخدم لاحتواء الاضمحلال الأسي المزدوج (الخطوط الحمراء). على اليمين: لوغاريتم القوة المعيارية مقابل لوغاريتم الزمن. تشير المنطقة المظللة إلى جزء البيانات المستخدم ليناسب قانون الطاقة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 8: روتين المسافة البادئة للقوة مع إزاحات المصيدة المثلثية. (أ) المسار التمثيلي ل x1 (t) باللون الأزرق و x2 (t) باللون الكهرماني خلال تجربة المسافة البادئة المثلثة التي أجريت على خلية في ارتفاع حبس 10 ميكرومتر. أعلى: موضع الفخ. الوسط: تحليل شكل النواة. تشير المسافة بين x1 و x2 إلى قطر النواة. أسفل: إشارة القوة. (ب) موضع القوة مقابل المصيدة لثمانية مسافات بادئة متتالية. (ج) تطور التبديد، المستمد من التباطؤ بين الاقتراب وجزء الانسحاب من منحنى f-d، للنواة لكل حدث مسافة بادئة لاحق. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 9. الخصائص النووية للخلايا المعلقة (سطح لاصق) والحبس من الروتين شبه المنحرف. (أ) المساحة المتوقعة للنواة من الخلايا المعلقة والأقل من 10 ميكرومتر. يمثل الشريط الأسود الوسيط. (ب) التصلب النووي للزنزانات المعلقة وتحت الحبس. يمثل الشريط الأسود الوسيط. قيم P المشتقة من اختبار Kruskal-Wallis باستخدام MatLab. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
الجدول التكميلي 1: المسار شبه المنحرف المحدد بواسطة برنامج الملقط البصري. الصف الأول (الثاني) هو المسافة x (y) التي سيتم إزاحة المصيدة خطيا. في الصف الثالث ، يتم تعيين مدة خطوة معينة بالثواني. يتكون هذا المسار من سبع نقاط ويتوافق مع شبه المنحرف الذي تم تحميله مرتين ضد النواة في الشكل 7B. يرجى النقر هنا لتنزيل هذا الجدول.
الجدول التكميلي 2: المسار الثلاثي المحدد بواسطة برنامج الملقط البصري. وعلى غرار الجدول 2، يتكون هذا المسار من 16 نقطة، تقابل ثمانية أحداث مسافة بادئة على عمق 5 ميكرومتر وسرعة 2.5 ميكرومتر/ثانية .
الجدول التكميلي 3: ملاءمة المعلمات للبيانات الواردة في الشكل 7. IG: التخمين الأولي. يرجى النقر هنا لتنزيل هذا الجدول.
الشكل التكميلي S1: محاذاة مستشعر القوة البصرية وتعويض خط الأساس للزخم. (أ) التوقف الميداني المصور في الكاميرا المساعدة (AUX، الشكل 2) من خلال عدسة برتراند. تظهر فقاعة الهواء مرئية في زيت الغمر ، والذي لا يمكن رؤيته من خلال العدسة. (ب) المسار البصري النظيف. للحصول على محاذاة دقيقة، افتح نقطة التوقف الميدانية واجعلها تتزامن مع مخروط الضوء NA = 1.2. (ج) صورة لمستوى العينة. يشير المربع الأحمر إلى منطقة عمل OT. شريط المقياس: 20 ميكرومتر (D) قوة المصيدة المقاسة عبر FOV ، على طول الأسهم المزدوجة البيضاء المشار إليها في C. باللون الأحمر، تباين طاقة الملائمة عند عدم تطبيق أي تصحيح. باللون الأزرق، تم تصحيح طاقة الملائمة على كامل مجال الرؤية. (ه) المكون X من خط الأساس للزخم على طول نفس النطاق. باللون الأحمر، تتبع غير مصحح. باللون الأزرق، تم تصحيح التتبع للحصول على طاقة المصيدة. باللون الأخضر، تم تصحيح التتبع لخط الأساس للزخم باستخدام تعويض الإزاحة العالمي في برنامج الشركة المصنعة. (و) نفس الشيء الوارد في E، بالنسبة للمكون Y. لاحظ أنه في ظل التشغيل العادي ، يتم استخدام المكونات المظللة لقياسات الميكانيكا والقوة ، على سبيل المثال ، مكون القوة x أثناء الحركة على طول الإحداثي x ومكون القوة y أثناء الحركة على طول المحور y. بعد تنفيذ جميع التصحيحات ، يتم الحصول على ضوضاء RMSD من <0.5 pN. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S2: روتين فاشل بسبب الفخاخ الضعيفة. (أ) Kymograph تظهر المسافة البادئة للنواة من روتين فاشل. فقط التشوهات القصيرة والعابرة مرئية بسبب هروب الخرزة من الفخ. الأهم من ذلك ، لا يزال ليزر الملائمة يتحرك بدون حبة لإكمال المسار المحدد مسبقا (الخط الأخضر المنقط). شريط المقياس = 10 ميكرومتر (B) أعلى: موضع المصيدة مقابل الوقت. الوسط: نتيجة تتبع الحافة لحافة النواة القريبة والبعيدة ذات المسافة البادئة. لاحظ أن الحافة البعيدة لا تتحرك بدون المسافة البادئة كما هو ملاحظ عادة للإجراءات الروتينية المكتملة على الخلايا المعزولة على ركائز لاصقة. القاع: القوة مقابل الوقت الذي يظهر فقدان المجهر المشار إليه بانخفاض في الضوضاء الحرارية وانخفاض مفاجئ إلى قوة الصفر. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S3: بقاء الأجنة المحقونة. تمت مقارنة الأجنة التي تم حقنها بخرز 1 ميكرومتر و 100 بيكوغرام / جنين من الحمض النووي الريبي المرسال بتركيزات موضحة في البروتوكول بالأجنة غير المخبوزة ولم تظهر أي اختلافات كبيرة 24 ساعة بعد الإخصاب. متوسط الانحراف المعياري لثلاث تجارب مستقلة مع N > 21 جنينا لكل حالة لكل تجربة. يرجى النقر هنا لتنزيل هذا الملف.
Discussion
في هذا البروتوكول ، نصف طريقة فريدة لاستجواب الخواص الميكانيكية لنواة الخلية داخل الخلايا الحية. وخلافا لتقنيات التحليل الطيفي للقوة الأخرى، سمح لنا الاصطياد البصري غير الغازي بفصل مساهمة غشاء الخلية والهيكل الخلوي عن الصلابة النووية للخلية. الأهم من ذلك ، أن المعالجة الدقيقة البصرية متوافقة مع المجهر متعدد الوسائط ، مما سيسمح للمجرب بدراسة العمليات المختلفة التي ينطوي عليها علم الأحياء الميكانيكية النووية للخلايا. كنتيجة تمثيلية ، استخدمنا تلطيخ DNA-Hoechst لقياس تشوه النواة عند المسافة البادئة التي تقوم بها قوى من ترتيب عدة مئات من picoNewton.
التطبيقات المحتملة لطريقتنا تتجاوز الأمثلة الموضحة في هذا البروتوكول
إن إمكانية استخراج المعلومات الميكانيكية الكمية من القياسات داخل الخلايا الحية دون اضطرابات خارجية تمكن من مجموعة كبيرة من الفرص غير المسبوقة التي بدأت للتو في استكشافها. وبالتالي ، يمكن توسيع البروتوكول المقدم لمنصة المعالجة الدقيقة البصرية الخاصة بنا إلى تجارب أكثر تعقيدا مع تنوع كبير. يمكن للانحرافات الصوتية البصرية (AOD) توليد مصائد بصرية متعددة لقياسات القوة المتزامنة عبر مواقع الخلايا المختلفة ، وكذلك يمكن استخدامها في علم الموجات الدقيقة النشط في نطاق تردد واسع51,61. كما ذكرنا ، يمكن لاستجابة القوة عند المسافة البادئة التغلب على أقصى قوة محاصرة ، مما يؤدي إلى هروب الخرزة من المصيدة البصرية. في هذه الحالة ، يمكن تكوين تغذية مرتدة للقوة باستخدام AOD من أجل تثبيت القوة البصرية. الكل في الكل ، يمكن الحصول على مناهج ميكروريولوجية متعددة ، مثل تخفيف الإجهاد الموصوف في هذا البروتوكول ، ولكن أيضا علم الأنسجة الدقيقة النشط أو الامتثال الزحف ، تجريبيا باستخدام هذه المنصة وتحليلها بدقة بواسطة حزم برامج جديدة61،62،63،64،65 . وعلاوة على ذلك، فإن تطبيق القوى لا يقتصر على النواة بل يمكن من حيث المبدأ القيام به لقياس الهياكل المتنوعة داخل الخلايا وفي الأنسجة المعقدة كما هو موضح في حبس خلايا الدم الحمراء المتدفقة داخل الأوعية الدموية السليمة666,67 أو حبس وتشويه البلاستيدات الخضراء والميتوكوندريا68 . تكون معايرة زخم الضوء مستقلة عن شكل وحجم الجسم المحاصر، مما يتيح إجراء قياسات القوة المباشرة على أي مسبار قوة ذي شكل تعسفي38,39. سمح لنا استخدام الميكروسفيرات المحقونة بتطبيق قوى عالية على النواة ذات طاقة ليزر منخفضة نسبيا مقارنة بالتلاعب المباشر بالهياكل الخلوية69,70,71. ومع ذلك ، نظرا لاختلاف معامل الانكسار المرتفع بما فيه الكفاية ، لا يلزم إجراء مسبار قوة مطبق خارجيا ويمكن معالجة العضيات داخل الخلايا مباشرة دون حقن الخرز (ملاحظات غير منشورة ومرجع 70).
التعديلات المحتملة على طريقتنا لتوسيع التطبيقات
يمكن حقن أحجام مختلفة من الميكروبيدات اعتمادا على التجربة ، ولكن يجب إجراء الضوابط النسبية. على سبيل المثال ، لدراسة الخلايا في مراحل لاحقة ، يمكن حقن حبات أصغر. سيؤدي ذلك إلى تقليل القوة القصوى التي يمكن ممارستها بواسطة المصيدة البصرية (كما هو موضح في المرجع 55). يمكن حقن حبات أكبر لممارسة قوى أعلى، ولكنها قد تؤثر على نمو الجنين اعتمادا على حجمها أو مرحلة اهتمامها. في التجارب التي لا يكون فيها حقن الميكروبيد خيارا، لا يزال من الممكن معالجة العضيات المختلفة التي تظهر اختلافات في مؤشرات الانكسار مقارنة بالسيتوبلازم بصريا، مما يؤدي إلى ظهور قوى بصرية قابلة للقياس من تغيرات زخم الضوء42. كما ذكر أعلاه ، تم استخدام هذه الطرق من قبل Bambardekar et al. لتشويه تقاطعات الخلايا الخلوية في جنين ذبابة الفاكهة 70. وبالمثل، فإن نواة الخلية لديها معامل انكسار أقل من الوسط المحيط44، مما يسمح بالمسافة البادئة الخالية من الخرز (الملاحظات غير المنشورة والمرجع 72) على الرغم من أن قوة الاصطياد أقل. وبالتالي ، لا يمكن محاصرة النواة بسهولة وتهرب من الفخ.
يتم تصنيع فاصل PDMS المطلي بالدوران عبر طريقة مريحة وسريعة ولكن قد يكون بعيدا عن متناول المختبرات دون الوصول إلى منشأة تصنيع متناهية الصغر / نانوية أو مختبرات هندسية. وبالتالي ، يمكن تجميع الفاصل بسهولة من شريط المختبر أو parafilm (الخطوة 4). يمكن أيضا تكييف البروتوكول عن طريق تصنيع قنوات الموائع الدقيقة التي تعمل على أتمتة توصيل الخلايا المفردة إلى آبار قياس محددة مسبقا أو إلى غرفة ذات ارتفاع محدد لتقدير تأثير الحبس داخل نفس العينة. ومع ذلك ، يجب تصميم هذه الأجهزة الموائع الدقيقة بحيث تناسب المسافة بين هدف المجهر وعدسة التجميع لمستشعر القوة البصرية ، حوالي 2 مم (انظر الخطوة 3). لاحظ أنه يجب وضع مستشعر القوة البصرية على الارتفاع المناسب بحيث لا تؤثر أي انحرافات بصرية عن إلغاء التركيز البؤري على قياس زخم الفوتون.
ويمكن أن تشمل التعديلات الأخرى تغيير المراسلين البيولوجيين. وجدنا أن Hoechst الفلورية تنزف طيفيا في قناة GFP ، وبالتالي فإننا نفضل الجمع مع هيستون الموسوم ب mCherry كعلامة نووية للقياس المتزامن في قناتين فلوريتين. بدلا من ذلك ، يمكن بسهولة تتبع التشوه النووي باستخدام ملصق يستهدف الغشاء النووي الداخلي مثل Lap2b-GFP (الشكل 2).
كانت المسافة البادئة على نواة الخلية في حدود 2-3 ميكرون ، والتي يمكننا قياسها بدقة عن طريق تحليل صورة المجهر البؤري المحدود للقرص الغزل المحدود الحيود. بالنسبة لحالة النوى الأكثر صلابة أو القوى الأصغر ، فإن المسافة البادئة ستكون بالكاد قابلة للقياس باستخدام هذا النهج. ومع ذلك، يمكن أيضا معايرة الملقط البصري المطلق المعاير للقوة لقياسات موضع الخرزة المحاصرة في الموقع باستخدام قياس التداخل BFP بدقة نانومتر51. باستخدام هذا النهج، يمكن ترجمة إشارة الجهد ومستشعر القوة البصرية إلى موضع المسبار المحاصر من خلال المعلمة β [نانومتر/فولت]، في حين أن المعلمة الثابتة α [pN/V] تنتج قيم القوة من خلال معايرة زخم الضوء المذكورة أعلاه41 (انظر أدناه للحصول على التفاصيل).
استكشاف الاخطاء
وجدنا أن التحديات التالية يمكن أن تحدث أثناء التجربة:
لا يتم تشكيل مصيدة مستقرة ويهرب المجهر بسهولة
أي أوساخ على هدف المجهر أو طوق تصحيح منحرف يمكن أن يؤدي إلى فشل فخ مستقر. إذا لم يتم العثور على حل فوري ، فقم بقياس وظيفة انتشار النقطة للعدسة الموضوعية. إذا كانت العينة ذات الاهتمام عميقة داخل نسيج كثيف بصريا ، فقد يواجه تركيز الليزر انحرافات بصرية شديدة تؤدي إلى محاصرة غير مستقرة (عادة ما يكون هذا التأثير ضئيلا في الخلايا المعزولة ولكنه يصبح أكثر وضوحا في الأنسجة السميكة). بالنسبة للصلابة العالية ، يمكن أن تتجاوز قوة استعادة النواة قوة الهروب من المصيدة ، بحيث يتم فقدان المجهر وفشل روتين المسافة البادئة. في البداية ، بالكاد يتم وضع مسافة بادئة لحافة الغشاء النووي القريبة من المصيدة البصرية (الشكل S2A). عندما يحدث هذا ، لم يعد ليزر الاصطياد يتأثر بالقوة والحركة البراونية ، مما يؤدي إلى انخفاض القوة إلى الصفر وانخفاض ضوضاء الإشارة (الشكل S2B). في حالة حدوث ذلك ، يمكن زيادة طاقة الليزر للحصول على مصيدة أقوى ، ويمكن تقليل سعة المسار شبه المنحرف الذي يدفع الخرزة إلى النواة ، أو يمكن وضع الموضع الأولي للميكروبيد المحاصر بعيدا عن النواة.
تتحرك الخلية أثناء التحفيز
إذا لم تكن الخلايا متصلة بما فيه الكفاية ، فإن مصيدة التدرج البصري ستحرك الخلايا أثناء إجراء روتين المسافة البادئة داخل الخلايا ، بحيث تكون القوى والميكانيكا الأساسية للنواة مصطنعة. لمنع إزاحة الخلية بأكملها ، نوصي بزيادة تركيز جزيئات التصاق الخلية على السطح ، على سبيل المثال ، ConA.
تعويض الزخم الأولي
إذا لم يكن روتين تعويض الزخم الأولي متاحا في منصة OTs (الخطوة 6.5) ، فيجب تصحيح إشارة خط الأساس المصطنعة المستقلة عن القوة. هذا مرئي كمنحدر على منحنى القوة حتى مع عدم وجود حبة محاصرة (الشكل S1E). للقيام بالتصحيح ، يجب إجراء نفس المسار بدون خرزة ، خارج الخلية في نفس الموضع بالضبط. لهذا، حرك الخلية بعيدا عن الملائمة باستخدام عنصر تحكم المرحلة. كمرجع ، تتغير قوة الإزاحة 5 pN عبر FOV عند 200 mW في نظامنا ؛ وبالتالي ، يصبح لا يكاد يذكر للمسارات القصيرة. بدلا من ذلك ، يمكن استخدام مرحلة مسح بيزو لتحريك الخلايا الموجودة على العينة ، مع ترك موضع الليزر ثابتا.
الخطوات الحاسمة للبروتوكول المقدم
يجب حقن الميكروسفير في المرحلة اليمنى ، 1 خلية لضمان أقصى توزيع على الجنين. لا ينبغي أن تكون الخرز فلورية بحيث لا يتسرب الضوء إلى قنوات الفلورسنت المستخدمة في التصوير. على سبيل المثال ، حتى الخرز الأحمر الفلورسنت النموذجي مرئي بوضوح في القناة الزرقاء المستخدمة لتصوير نواة الخلية بعد تلطيخ Hoechst بسبب سطوعها (الإثارة: 405 نانومتر ؛ الانبعاثات: 445 نانومتر). يعد التعلق المستقر للخلية بالركيزة أمرا بالغ الأهمية لمنع الإزاحة الجانبية أثناء روتين المسافة البادئة. إذا تحركت الخلية أثناء الروتين ، يتم التقليل من شأن القوى. في حالة حدوث ذلك بشكل متكرر ، قم بتحسين بروتوكول المرفق. بالنسبة لخلايا زراعة الأنسجة ، تؤدي بروتينات التصاق الخلايا الأخرى ، مثل الفيبرونيكتين أو الكولاجين أو البولي إل ليسين إلى ارتباط مرض (ملاحظات غير منشورة). أثناء الحبس ، تتعرض الخلايا لإجهاد ميكانيكي مفاجئ وشديد. هذا يمكن أن يسبب تلفا للخلايا وغالبا ما يواجه المجرب خلايا منفجرة إذا لم يتم تنفيذ الإجراء بعناية. أيضا ، إذا كان ارتفاع الحبس صغيرا جدا ، فستعاني جميع الخلايا من كسر المغلف النووي أو تلف لا رجعة فيه. للتخفيف من حدتها، قم بخفض زلة الغطاء العلوية ببطء أكبر و/أو قم بزيادة التباعد بين زلة الغطاء.
قيود التقنية والاقتراحات للتغلب عليها
هناك قيود واضحة على هذه التقنية وهي تغلغل ضوء الليزر في أقسام عميقة من الأنسجة ، مما يؤدي إلى انحرافات ومحاصرة غير مستقرة. وبالتالي ، يعتمد الحد الأدنى لعمق الاختراق على وضوح العينة ، وتصحيح الانحراف الذي يمكن استخدامه73 وقوة الليزر المطبقة. يجب أن يؤخذ في الاعتبار أن طاقة الليزر الأعلى تؤدي إلى الإثارة الحرارية للعينة بالقرب من المجهر. ومع ذلك ، يتم تقليل تسخين العينة التي نشأت عن بقعة الليزر ذات الطول الموجي 1064 نانومتر لتجنب الإجهاد المعقول المرتبط بالحرارة على عيناتنا البيولوجية74.
قيد آخر هو الحد الأقصى للقوة التي يمكن قياسها. على الرغم من أن الكشف المباشر عن زخم الضوء يمكن من قياس القوة إلى ما هو أبعد من نظام الاستجابة الخطية للمصيدة البصرية40,41 ، فإن القوة القصوى المطبقة هي في حدود بضع مئات من بيكو نيوتن. وهذا محدود بقوة الليزر وما يترتب على ذلك من عتبة الضرر للمواد البيولوجية اللينة والاختلافات في معامل الانكسار، والتي لا تزيد عادة عن 0.1 أو 0.344. وقد اقترحت عدة طرق لزيادة حد الكشف عن القوة، على سبيل المثال، استخدام الضوء المنظم75، والميكروسفير المغلف المضاد للانعكاس76، والجسيمات ذات معامل الانكسار العالي77 أو النقاط الكمومية شديدة المنشطات78.
يمكن استخدام OTs لقياسات موضع النانومتر من خلال قياس التداخل BFP ، بحيث يكون موضع الخرزة داخل المصيدة هو Δx = β Sx ، حيث Sx هي إشارة الجهد للمستشعر ، ويمكن معايرة β [μm / V] أثناء التنقل باتباع بروتوكولات مختلفة35,54. بالنسبة لمستشعر القوة البصرية ، يمكن إثبات أن عامل التحويل الثابت من الجهد إلى القوة α [pN / V] يرتبط مباشرة β وصلابة المصيدة ، k [pN / μm] ، من خلال α = kβ 37) في التجارب مع إزاحات الخرز الصغيرة جدا التي لا يمكن اكتشافها من التصوير البصري ، يمكن استخدام هذه الاستراتيجية لاستكمال قياسات القوة مع الكشف عن موضع صغير. مثال على ذلك هو تطبيق الروتين التجريبي المعروض هنا على نوى شديدة الصلابة ، والتي لا تكفي القوى ذات قوى الليزر المعقولة (200-500 ميجاوات) للحث على قيم المسافة البادئة الكبيرة بما فيه الكفاية. في هذه الحالة ، يجب ملامسة الخرزة للنواة ويجب معايرة صلابة المحاصرة قبل القياس (الخطوة 8.6). يمكن تحديد المسافة البادئة d للنواة كدالة للقوة بشكل غير مباشر على النحو التالي:
d = xtrap - F/k
حيث xtrap هو موضع المصيدة. يختلف العامل β [μm/V] عن عامل زخم الضوء الثابت α [pN/V]، حيث يحتاج إلى معايرة قبل كل تجربة لأنه يعتمد على العديد من المتغيرات المحلية التي تحدد ديناميكيات الملائمة، مثل حجم الجسيمات، وحجم بقعة المصيدة البصرية، ومؤشرات الانكسار النسبية.
Disclosures
وليس لدى صاحبي البلاغ ما يكشفان عنه.
Acknowledgements
تعترف MK بالدعم المالي المقدم من وزارة الاقتصاد والقدرة التنافسية الإسبانية من خلال الخطة الوطنية (PGC2018-097882-A-I00) ، FEDER (EQC2018-005048-P) ، برنامج سيفيرو أوتشوا لمراكز التميز في البحث والتطوير (CEX2019-000910-S; RYC-2016-21062) ، من مؤسسة Privada Cellex ، و Fundació Mir-Puig ، ومن Generalitat de Catalunya من خلال CERCA وبرنامج الأبحاث (2017 SGR 1012) ، بالإضافة إلى التمويل من خلال ERC (MechanoSystems) و HFSP (CDA00023/2018). تعترف V.R. بالدعم المقدم من وزارة العلوم والابتكار الإسبانية لشراكة EMBL ، ومركز Excelencia Severo Ochoa ، وخطة MINECO الوطنية (BFU2017-86296-P ، PID2020-117011GB-I00) و Generalitat de Catalunya (CERCA). تعترف V.V. بالدعم المقدم من برنامج الدكتوراه ICFOstepstone الممول من برنامج البحث والابتكار Horizon 2020 التابع للاتحاد الأوروبي بموجب اتفاقية منحة Marie Skłodowska-Curie 665884. نشكر أرناو فاري على قراءته النقدية للمخطوطة. ماريا مارسال للمساعدة في تصوير وتركيب الجنين 24 hpf و ؛ سيندا خيمينيز-ديلغادو للحصول على الدعم مع الحقن المجهري لسمك الزرد.
Materials
| Name | Company | Catalog Number | Comments |
| #1.5 22 mm cover glasses | Ted Pella | 260148 | |
| #1.5 22x60 mm Coverglasses | Ted Pella | 260152 | |
| #1.5H glass bottom dishes | Willco | GWST-5040 | |
| 10-um beads | Supelco | 72986 | |
| 1-mm glass capillaries | Harvard Apparatus | 30-0020 GC100F-15 | |
| 1-um polystyrene microbeads | Sigma | 89904 | |
| 1-um red-fluorescent beads | ThermoFisher | F8816 | |
| Agar | ThermoFisher | 16500500 | |
| Aqcuisition cameras sCMOS | Andor | Sona-4BV11-UNI | |
| Auxiliary camera (Figure 3, AUX) | Blackfly, FLIR | BFS-U3-200S6M-C | |
| Calbryte 520 | AAT Bioquest | 520 AM | |
| Centrifuge | Eppendorf | 5453000011 | |
| Concanavalin A | Sigma | C5275 | |
| DMEM | Sigma | D2906 | |
| DNA-Hoechst | ThermoFisher | 33342 | |
| Double scotch tape | Biesse Adesivi | ||
| E3 | 5 mM NaCl. 0.17 mM KCl. 0.33 mM CaCl2. 0.33 mM MgSO4 | ||
| Eclipse Ti2 | Nikon | ||
| Forceps | Fine Science Tools | 11252-20 | |
| GPI-GFP | |||
| H2A-mCh | |||
| Image acquisition software | Fusion-Andor | ||
| Immersion Oil | Cargille | Type B: 16484 | |
| IR protection googles | Thorlabs | LG1 | |
| Lap2b-eGFP | |||
| Micro loader pipette | Eppendorf | GELoader | |
| Microinjector | World Precision Instruments | SYS-PV820 | |
| MicroManager 2.0 | |||
| Micromiter slide | ID5243 GXMGRAT-5 5mm/100 divisions | ||
| Mineral oil | Sigma | M3616 | |
| Motorized stage | ASI | ||
| Needle puller | Sutter instrument Co. | Model P-97 | |
| Optical tweezers platform | Impetux Optics | Sensocell | |
| OTs software (LightAce) | Impetux Optics | ||
| PDMS | Sigma | Sylgard 184 | |
| PDMS Curing agent | Sigma | Sylgard 184 | |
| Post processing software (Matlab) | Mathworks | ||
| RNAse free water | Thermofisher | AM9937 | |
| Short-pass dichroic mirror (Figure 3, IR-F) | Semrock | FF01-950/SP-25 | |
| Spin-coater | Specialty Coating Systems | Spincoat G3P-8 | |
| Spinning-disk confocal microscope | Andor | DragonFly 502 | |
| Stereomicroscope | Leica M80 | ||
| Triangular microinjection mold | Adaptive Science Tools | TU1 | |
| Universal oven | Memmert | UNB 200 | |
| Water immersion objective | Nikon | MRD07602 |
References
- Chan, C. J., Heisenberg, C. P., Hiiragi, T. Coordination of Morphogenesis and Cell-Fate Specification in Development. Current Biology. 27 (18), 1024-1035 (2017).
- Heller, E., Fuchs, E. Tissue patterning and cellular mechanics. Journal of Cell Biology. 211 (2), 219-231 (2015).
- Heisenberg, C. P., Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell. 153 (5), 948-962 (2013).
- Petridou, N. I., Spiró, Z., Heisenberg, C. P. Multiscale force sensing in development. Nature Cell Biology. 19 (6), 581-588 (2017).
- Krieg, M., et al. Tensile forces govern germ-layer organization in zebrafish. Nature Cell Biology. 10 (4), 429-436 (2008).
- Ruprecht, V., et al. Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell. 160 (4), 673-685 (2015).
- Shellard, A., Mayor, R. Supracellular migration - Beyond collective cell migration. Journal of Cell Science. 132 (8), (2019).
- Mongera, A., et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. 561 (7723), 401-405 (2018).
- Atia, L., et al. Geometric constraints during epithelial jamming. Nature Physics. 14 (6), 613-620 (2018).
- Turlier, H., Maître, J. -. L. Mechanics of tissue compaction. Seminars in Cell & Developmental Biology. 47-48, 110-117 (2015).
- Ladoux, B., Mège, R. M. Mechanobiology of collective cell behaviours. Nature Reviews Molecular Cell Biology. 18 (12), 743-757 (2017).
- Venturini, V., et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science. 370 (6514), (2020).
- Charras, G., Sahai, E. Physical influences of the extracellular environment on cell migration. Nature Reviews Molecular Cell Biology. 15 (12), 813-824 (2014).
- Kirby, T. J., Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nature Cell Biology. 20 (4), 373-381 (2018).
- Lee, H. P., et al. The nuclear piston activates mechanosensitive ion channels to generate cell migration paths in confining microenvironments. Science Advances. 7 (2), (2021).
- Friedl, P., Wolf, K., Lammerding, J. Nuclear mechanics during cell migration. Current Opinion in Cell Biology. 23 (1), 55-64 (2011).
- Versaevel, M., Riaz, M., Grevesse, T., Gabriele, S. Cell confinement: Putting the squeeze on the nucleus. Soft Matter. 9 (29), 6665-6676 (2013).
- Zuela-Sopilniak, N., et al. Measuring nucleus mechanics within a living multicellular organism: Physical decoupling and attenuated recovery rate are physiological protective mechanisms of the cell nucleus under high mechanical load. Molecular Biology of the Cell. 31 (17), 1943-1950 (2020).
- Kim, D. H., Wirtz, D. Cytoskeletal tension induces the polarized architecture of the nucleus. Biomaterials. 48, 161-172 (2015).
- Lomakin, A. J., et al. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science. 370 (6514), (2020).
- Hampoelz, B., et al. Microtubule-induced nuclear envelope fluctuations control chromatin dynamics in Drosophila embryos. Development. 138 (16), 3377-3386 (2011).
- Heo, S. J., et al. Differentiation alters stem cell nuclear architecture, mechanics, and mechano-sensitivity. eLife. 5, 1-21 (2016).
- Cosgrove, B. D., et al. Nuclear envelope wrinkling predicts mesenchymal progenitor cell mechano-response in 2D and 3D microenvironments. Biomaterials. 270, 120662 (2021).
- Liu, H., et al. In situ mechanical characterization of the cell nucleus by atomic force microscopy. ACS Nano. 8 (4), 3821-3828 (2014).
- Hobson, C. M., et al. Correlating nuclear morphology and external force with combined atomic force microscopy and light sheet imaging separates roles of chromatin and lamin A/C in nuclear mechanics. Molecular Biology of the Cell. 31 (16), 1788-1801 (2020).
- Pajerowski, J. D., Dahl, K. N., Zhong, F. L., Sammak, P. J., Discher, D. E. Physical plasticity of the nucleus in stem cell differentiation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15619-15624 (2007).
- Rowat, A. C., Lammerding, J., Ipsen, J. H. Mechanical properties of the cell nucleus and the effect of emerin deficiency. Biophysical Journal. 91 (12), 4649-4664 (2006).
- Davidson, P. M., et al. High-throughput microfluidic micropipette aspiration device to probe time-scale dependent nuclear mechanics in intact cells. Lab on a Chip. 19 (21), 3652-3663 (2019).
- Lombardi, M., Zwerger, M., Lammerding, J. Biophysical assays to probe the mechanical properties of the interphase cell nucleus: Substrate strain application and microneedle manipulation. Journal of Visualized Experiments: JoVE. (55), (2011).
- Luo, T., Mohan, K., Iglesias, P. A., Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nature Materials. 12 (11), 1064-1071 (2013).
- Dahl, K. N., Engler, A. J., Pajerowski, J. D., Discher, D. E. Power-law rheology of isolated nuclei with deformation mapping of nuclear substructures. Biophysical Journal. 89 (4), 2855-2864 (2005).
- Guilluy, C., et al. Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. Nature Cell Biology. 16 (4), 376-381 (2014).
- Bustamante, C. J., Wang, M. D. Optical tweezers in single-molecule biophysics. Nature Reviews Methods Primers. , 1-29 (2021).
- Svoboda, K., Block, S. M. Force and velocity measured for single kinesin molecules. Cell. 77 (5), 773-784 (1994).
- Berg-Sørensen, K., Flyvbjerg, H. Power spectrum analysis for optical tweezers. Review of Scientific Instruments. 75 (3), 594-612 (2004).
- Smith, S. B., Cui, Y., Bustamante, C. Optical-trap force transducer that operates by direct measurement of light momentum. Methods in Enzymology. 361 (1994), 134-162 (2003).
- Farré, A., Montes-Usategui, M. A force detection technique for single-beam optical traps based on direct measurement of light momentum changes. Optics Express. 18 (11), 11955 (2010).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Extending calibration-free force measurements to optically-trapped rod-shaped samples. Scientific Reports. 7, 1-10 (2017).
- Bui, A. A. M., et al. Calibration of force detection for arbitrarily shaped particles in optical tweezers. Scientific Reports. 8 (1), 1-12 (2018).
- Farré, A., Marsà, F., Montes-Usategui, M. Beyond the hookean spring model: Direct measurement of optical forces through light momentum changes. Methods in Molecular Biology. 1486, (2017).
- Farré, A., Marsà, F., Montes-Usategui, M. Optimized back-focal-plane interferometry directly measures forces of optically trapped particles. Optics Express. 20 (11), 12270 (2012).
- Jun, Y., Tripathy, S. K., Narayanareddy, B. R. J., Mattson-Hoss, M. K., Gross, S. P. Calibration of optical tweezers for in vivo force measurements: How do different approaches compare. Biophysical Journal. 107 (6), 1474-1484 (2014).
- Mas, J., Farré, A., Sancho-Parramon, J., Martín-Badosa, E., Montes-Usategui, M. Force measurements with optical tweezers inside living cells. Optical Trapping and Optical Micromanipulation XI. 9164, (2014).
- Schürmann, M., Scholze, J., Müller, P., Guck, J., Chan, C. J. Cell nuclei have lower refractive index and mass density than cytoplasm. Journal of Biophotonics. 9 (10), 1068-1076 (2016).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments: JoVE. (25), (2009).
- Westerfield, M. . The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish. (Danio rerio), 5th Edition. , (2007).
- Schubert, R., et al. Assay for characterizing the recovery of vertebrate cells for adhesion measurements by single-cell force spectroscopy. FEBS Letters. 588 (19), 3639-3648 (2014).
- Koschwanez, J. H., Carlson, R. H., Meldrum, D. R. Thin PDMS films using long spin times or tert-butyl alcohol as a solvent. PLoS One. 4 (2), 2-6 (2009).
- Das, R., et al. Mechanical stretch inhibition sensitizes proprioceptors to compressive stresses. bioRxiv. , (2021).
- Chardès, C., Clement, R., Blanc, O., Lenne, P. F. Probing cell mechanics with bead-free optical tweezers in the drosophila embryo. Journal of Visualized Experiments: JoVE. (141), (2018).
- Staunton, J. R., Blehm, B., Devine, A., Tanner, K. In situ calibration of position detection in an optical trap for active microrheology in viscous materials. Optics Express. 25 (3), 1746 (2017).
- Bola, R., Treptow, D., Marzoa, A., Montes-Usategui, M., Martin-Badosa, E. Acousto-holographic optical tweezers. Optics Letters. 45 (10), 2938-2941 (2020).
- Thalhammer, G., Obmascher, L., Ritsch-Marte, M. Direct measurement of axial optical forces. Optics Express. 23 (5), 6112 (2015).
- Vermeulen, K. C., et al. Calibrating bead displacements in optical tweezers using acousto-optic deflectors. Review of Scientific Instruments. 77 (1), 1-6 (2006).
- Dzementsei, A., Barooji, Y. F., Ober, E. A., Oddershede, L. B. Foregut organ progenitors and their niche display distinct viscoelastic properties in vivo during early morphogenesis stages. bioRxiv. , 1-35 (2021).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Krieg, M., et al. Atomic force microscopy-based mechanobiology. Nature Reviews Physics. , (2018).
- A-Hassan, E., et al. Relative microelastic mapping of living cells by atomic force microscopy. Biophysical Journal. 74 (3), 1564-1578 (1998).
- Khalilgharibi, N., et al. Stress relaxation in epithelial monolayers is controlled by the actomyosin cortex. Nature Physics. 15, (2019).
- Crick, S. L., Yin, F. C. Assessing micromechanical properties of cells with atomic force microscopy: importance of the contact point. Biomechanics and Modeling in Mechanobiology. 6 (3), 199-210 (2007).
- Hurst, S., Vos, B. E., Betz, T. Intracellular softening and fluidification reveals a mechanical switch of cytoskeletal material contributions during division. bioRxiv. , 425761 (2021).
- Kaplan, J. L., Bonfanti, A., Kabla, A. RHEOS.jl - A Julia package for rheology data analysis. arXiv. 4, 1-5 (2020).
- Bonfanti, A., Kaplan, J. L., Charras, G., Kabla, A. Fractional viscoelastic models for power-law materials. Soft Matter. 16 (26), 6002-6020 (2020).
- Rivas-Barbosa, R., Escobedo-Sánchez, M. A., Tassieri, M., Laurati, M. i-Rheo: determining the linear viscoelastic moduli of colloidal dispersions from step-stress measurements. Physical Chemistry Chemical Physics: PCCP. 22 (7), 3839-3848 (2020).
- Tassieri, M., et al. i-Rheo: Measuring the materials' linear viscoelastic properties "in a step". Journal of Rheology. 60 (4), 649-660 (2016).
- Zhong, M. C., Wei, X. B., Zhou, J. H., Wang, Z. Q., Li, Y. M. Trapping red blood cells in living animals using optical tweezers. Nature Communications. 4, 1767-1768 (2013).
- Harlepp, S., Thalmann, F., Follain, G., Goetz, J. G. Hemodynamic forces can be accurately measured in vivo with optical tweezers. Molecular Biology of the Cell. 28 (23), 3252-3260 (2017).
- Bayoudh, S., Mehta, M., Rubinsztein-Dunlop, H., Heckenberg, N. R., Critchley, C. Micromanipulation of chloroplasts using optical tweezers. Journal of Microscopy. 203 (2), 214-222 (2001).
- Favre-Bulle, I. A., Stilgoe, A. B., Rubinsztein-Dunlop, H., Scott, E. K. Optical trapping of otoliths drives vestibular behaviours in larval zebrafish. Nature Communications. 8 (1), 630 (2017).
- Bambardekar, K., Clément, R., Blanc, O., Chardès, C., Lenne, P. F. Direct laser manipulation reveals the mechanics of cell contacts in vivo. Proceedings of the National Academy of Sciences of the United States of America. 112 (5), 1416-1421 (2015).
- Ferro, V., Chuai, M., McGloin, D., Weijer, C. J. Measurement of junctional tension in epithelial cells at the onset of primitive streak formation in the chick embryo via non-destructive optical manipulation. Development (Cambridge). 147 (3), (2020).
- Hörner, F., et al. Holographic optical tweezers-based in vivo manipulations in zebrafish embryos. Journal of Biophotonics. 10 (11), 1492-1501 (2017).
- Zhong, M. -. C., Wang, Z. -. Q., Li, Y. -. M. Aberration compensation for optical trapping of cells within living mice. Applied Optics. 56 (7), 1972 (2017).
- Català, F., Marsà, F., Montes-Usategui, M., Farré, A., Martín-Badosa, E. Influence of experimental parameters on the laser heating of an optical trap. Scientific Reports. 7 (1), 1-9 (2017).
- Taylor, M. A., Waleed, M., Stilgoe, A. B., Rubinsztein-Dunlop, H., Bowen, W. P. Enhanced optical trapping via structured scattering. Nature Photonics. 9 (10), 669-673 (2015).
- Bormuth, V., et al. Optical trapping of coated microspheres. Optics Express. 16 (18), 13831-13844 (2008).
- Sudhakar, S., et al. Germanium nanospheres for ultraresolution picotensiometry of kinesin motors. Science. 371 (6530), (2021).
- Shan, X., et al. Optical tweezers beyond refractive index mismatch using highly doped upconversion nanoparticles. Nature Nanotechnology. 16 (5), 531-537 (2021).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved