
Method Article
在共同的文化整合模式中GABA能神经元的刺和HEK293细胞抑制性突触形成稳定表达GABA
摘要
个体发育过程中统筹抑制GABA能突触形成的分子机制知之甚少。为了研究这些方法中,我们已经开发出一种共培养模型系统,该系统采用了与稳定转染人胚胎肾293(HEK293)表达功能性GABA A受体的细胞一起培养的胚胎中型多棘GABA能神经元。
摘要
Inhibitory neurons act in the central nervous system to regulate the dynamics and spatio-temporal co-ordination of neuronal networks. GABA (γ-aminobutyric acid) is the predominant inhibitory neurotransmitter in the brain. It is released from the presynaptic terminals of inhibitory neurons within highly specialized intercellular junctions known as synapses, where it binds to GABAA receptors (GABAARs) present at the plasma membrane of the synapse-receiving, postsynaptic neurons. Activation of these GABA-gated ion channels leads to influx of chloride resulting in postsynaptic potential changes that decrease the probability that these neurons will generate action potentials.
During development, diverse types of inhibitory neurons with distinct morphological, electrophysiological and neurochemical characteristics have the ability to recognize their target neurons and form synapses which incorporate specific GABAARs subtypes. This principle of selective innervation of neuronal targets raises the question as to how the appropriate synaptic partners identify each other.
To elucidate the underlying molecular mechanisms, a novel in vitro co-culture model system was established, in which medium spiny GABAergic neurons, a highly homogenous population of neurons isolated from the embryonic striatum, were cultured with stably transfected HEK293 cell lines that express different GABAAR subtypes. Synapses form rapidly, efficiently and selectively in this system, and are easily accessible for quantification. Our results indicate that various GABAAR subtypes differ in their ability to promote synapse formation, suggesting that this reduced in vitro model system can be used to reproduce, at least in part, the in vivo conditions required for the recognition of the appropriate synaptic partners and formation of specific synapses. Here the protocols for culturing the medium spiny neurons and generating HEK293 cells lines expressing GABAARs are first described, followed by detailed instructions on how to combine these two cell types in co-culture and analyze the formation of synaptic contacts.
引言
GABA是一个在胚胎大脑发现最早的神经递质,最丰富的兴奋性神经递质谷氨酸盐1前面。在开发过程中,GABA的去极化,并激发未成熟的神经元,在调节细胞增殖,迁移和形成的神经元网络的未经诱导的兴奋性中毒的关键作用。在成人脑,反转电位为γ-氨基丁酸A受体通道被移动到更负的电位,由于在氯化物的胞内浓度的降低。这个移位是通过上调钾氯化物共转运体(KCC2),其输送氯化物出细胞引起的,并且,并行地下调的钠 - 钾 - 氯化物转运体(NKCC1),其具有相反的效果2。
在大脑中,GABA主要结合两种GABA A和GABA B受体介导快或慢突触抑制,相应的LY。 GABA A卢比是一类受体也被称为异五离子型或配体门控半胱氨酸环离子通道。需要进行活化的受体,它是可渗透的氯离子,并在较小程度,碳酸氢根离子的两个分子的GABA。在氯化物电导的增加而降低的激活突触后神经元3去极化,兴奋性活动的有效性。
对GABA A卢比结构的多样性一直被认为是决定其广泛的功能和药理特性的关键因素。天然GABA A卢比是杂五聚体亚单位具有多个同种型分类为组成:α(1-6),β(1-3),γ(1-3),δ,ε,π和θ3,具有一个共同的跨膜拓扑结构包括一个大的N-末端胞外结构域,4个跨膜结构域(TM的),和记忆库3和4 4之间的一个主要的胞内结构域的β3和γ2亚基的突触抑制和生物生存所必需的,因为出生后5,6小鼠基因敲除这些亚基死亡。与此相反,α亚基的单个异构体可用于在具有不同的行为,如焦虑,镇静,觉醒,以及其他相关的大脑特定突触连接的功能是重要的,但不是单独地,对于生活7-9必不可少的。 GABA A卢比的作用,适用于各种药物具有强效镇静,催眠,抗焦虑和抗惊厥的作用,如苯二氮卓类,巴比妥类,甾体和麻醉药7,10,11的主要场所。
突触的 GABA A卢比通常包含γ2亚单位,2个β亚单位(最常见的β2或β3)和两个α亚基(α1,α2,α3或α5)12,13。主要的类外突触受体包含在组合δ亚基带有两个α亚基(α4或α6),和两个β亚基(β2或β3)14。对GABA A卢比到轴突,树突或细胞体,而进入细胞膜的插入的亚细胞定位是依赖于β亚基15,16的存在。然而,不同的GABA A R的选择性结合亚型成不同类型的突触具有特定的α亚基(α1,α2,α3或α5)7,17,18存在很好的相关性。重要的是,小鼠缺失α1和α2亚基会导致在抑制性突触19超微结构的改变。这表明,GABA A RS本身可能在调节突触的形成起到直接的作用。
有证据表明,GABA能突触的发展是一个精确协调的事件序列,其中两个神经元靶接触的不同类型的抑制轴突,并且聚集在受体每一类抑制性突触是选择性和功能上切合17,20-22。特异性的GABA能突触这一基本原则提出了一个问题,就是突触接触的过程中开始前和突触后的合作伙伴是如何认出对方。
在体外共培养测定法已成功地应用于研究一些突触形成的机理,并测试在这个过程中个体突触间隙跨度蛋白质的作用。一个常见的跨突触相互作用蛋白组合的功能双向调解突触的形成和成熟,是Neurexins(Nrxns)和neuroligin的小鼠(NLS)。 Nrxns是突触前蛋白,表现出它们的层粘连蛋白neurexin-性激素结合蛋白结构域之内的选择性剪接,以许多不同的异构体23引起。而Nrxns也与其他蛋白相互作用,NLS被认为是他们的无处不在的突触后巴rtners 24。总之这些蛋白质有助于保持突触前和突触后膜的紧密和严格的并置25。两种最丰富的亚型是NL-1和NL-2,它是存在于兴奋性和抑制性突触,分别为26。一个最早的共培养模型系统,旨在探讨跨突触蛋白相互作用,所使用的不同类型的非神经元细胞,最常用的永生化细胞系,例如人胚胎肾(HEK)293细胞,过表达NL- 2。当这些细胞与桥脑的神经元培养的,突触前蛋白质在接近所述HEK细胞的表面上的积累,观察,表明形成突触样接触。除了 可溶性β-neurexin这些共培养物的抑制接触的形成,这表明Nrxns和NLS之间跨突触的相互作用是必需的突触形成接触27。此外,瞬时表达β-neurexin在COS(℃的V-1(猿猴) 为 O rigin,并携有S V40的遗传物质)的细胞共培养解离的海马谷氨酸能和GABA能神经元的GABA A R亚基的突触蛋白gephryin和诱导表达的γ2和α2在这两种细胞类型28之间的接触点。用于研究突触形成的共培养模型的另一例所涉及的HEK293细胞中,瞬时转染的GABA A R亚基α2/β3/γ2和NL-2,和下丘脑神经元29的混合种群。该研究的结论是,NL-2的表达是为了形成抑制性突触的绝对要求。
然而,在最近的共培养物的研究中,稳定转染的α1/β2/γ2 的 GABA A在HEK293细胞卢比被认为是足以诱导功能性突触时共培养与GABA能MED鎓多棘神经元,而不需要附加的跨突触或突触后粘附蛋白。然而,在突触形成和强度的显着增加,观察当NL-2共表达与GABA A卢比30。这表明该共培养模型系统具有过前面描述的模型系统中,最明显地增加的灵敏度和突触接触检测的可靠性的优点。有助于在检测的突触联系的整体改善的两个重要因素是:i)使用稳定转染的HEK293细胞系中含有单个细胞的表面高的和一致的表达GABA A R亚基。这种一致性有利于不同共培养条件之间的定量比较。二)使用GABA能中型多棘神经元的纯人口从胚胎31纹状体培养删除并发症及因使用混合性神经元群并产生歧义异体ws的,例如,选择最合适的突触后的GABA A R的类型,可以彼此在突触的形成进行比较。
突触的形成被认为涉及前和突触后的细胞粘附复合物中的许多跨突触信号。由于突触信号和细胞粘附分子的绝对数量的双向性质,很难鉴定参与突触形成的关键组件。因此,转染的单个细胞粘附蛋白进入非神经元细胞(在这种情况下, 在体内的GABA能中型多棘神经元的两个最常见的突触后的目标,α1/β2/γ2或α1/β3/γ2 的 GABA A卢比32)大大减少了可在突触后表面跨突触信号的复杂性,并允许这种蛋白质在促进突触形成的功效的精确的定量分析。
研究方案
SD大鼠或BAB / C近交系小鼠(Harlan,UK;用于怀孕的女性人数是30),是根据英国内政部[和1986年11月24日欧共体理事会指令安置和牺牲(609分之86/ EEC) ]指导原则。该项目正式通过药剂伦理委员会的UCL学校批准。
1.准备仪器,培养基和菜名
- 打开和清洁层流罩用70%的乙醇,以便在任何时候都在无菌条件下工作。
- 制备的HEK293细胞培养基,含有Dulbecco改良的Eagle培养基pH为7.4(DMEM,500毫升),L-谷氨酰胺(2毫摩尔),青霉素(50单位/毫升),链霉素(50微克/毫升),和胎牛血清(10 %)。
注:青霉素和链霉素的刺激。 - 制备无血清的神经元培养基,含有Neurobasal培养基pH为7.4(500毫升)中,B27添加物(25毫升),L-谷氨酰胺(2毫摩尔),青霉素(50单位/毫升),链霉素(50微克/毫升),和葡萄糖(6毫米)。
- 制成500毫升之的HEPES缓冲盐溶液(HBSS)中含有的HBSS 10倍的库存(50毫升),HEPES(1μM)(5毫升)和水(445毫升),pH为7.4。
- 高压釜的磷酸盐缓冲生理盐水溶液(PBS,pH 7.4中; 1L),水(1L),玻璃盖玻片(13毫米直径)和玻璃巴斯德移液管进行消毒它们。
表达α1/β3/γ2-GABA A卢比2.制备HEK293稳定细胞系
- 板2×10 6的HEK293细胞进10cm的无菌组织培养板,在37℃С为了过夜达到70-90%汇合培养在湿润的5%二氧化碳(CO 2)的气氛中(CO 2培养箱)。
- 第二天,转染的HEK293细胞与GABA A Rα1亚单位的cDNA中掺入G418二硫酸盐( 表1)抗性基因的PCDN 3.1(+)表达载体与GABA A Rβ3亚基的cDNA在表 达载体结合的腐草霉素D 1( 表1)抗性基因(两者下一个人巨细胞病毒立即早期(CMV)启动子的调节),使用阳离子脂质体制剂,该复合物与带负根据制造商的协议带电的核酸分子( 表1)。
- 简言之,加入500μl低血清培养基(pH为7.4; 表1)和7.5微克每cDNA的构建到一个无菌的15毫升离心管中,接着加入15μl脂质体转染缓冲试剂,和前在室温下放置,轻轻混匀5分钟。
- 向此混合物中,加入8.75微升脂质体转染试剂和前在室温下放置30分钟,轻轻混匀。在此之后,添加3毫升HEK293细胞的培养基(无抗生素),吸液管的离心管中的内容上下两次,transfe- [R滴加到细胞在10cm的组织培养皿中生长,并孵育48小时,在37°С在湿润的5%CO 2的气氛中(CO 2培养箱)。
- 用无菌PBS,pH 7.4中轻轻洗HEK293细胞,并稀释转染的HEK293细胞进入新10厘米组织培养皿中,在下面的比率:1:3,1:5,1:7,1:10 1:15和1:20。这确保了细胞将不会变得过分汇合。
- 通过将每种抗生素选择标记,G418,和腐草霉素D1的800微克/毫升,加入到培养基中启动选择的HEK293细胞中同时表达GABA A R亚基。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换含抗生素的培养基中(10毫升)中,每2天。
- 当小的白色菌落开始形成(通常在约7天),小心地从每个菜肴选择单个菌落并使用无菌P1000收集它枪头。其传送含有500μl的培养基中,并小心地通过吸取培养基上下重悬在24孔组织培养板的一个井。确保只有一个菌落被转移到每个孔中(共5-20菌落建议)。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换含抗生素的培养基中,每2天。
- 一旦菌落变成70-80%汇合时,轻轻地吸取培养基上下,从24孔组织培养板的底部移去的细胞。转移,分化在6孔组织培养板2的孔之间的细胞的悬浮液。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换含抗生素的培养基中,每2天。
- 一旦70% - 80%汇合时,从1的2孔中含有细胞来自同一单个结肠始发的收集细胞y和准备蛋白裂解。简单地说,洗涤细胞2倍的PBS,pH 7.4中,并且在PBS,pH 7.4中添加200μl的2%十二烷基硫酸钠(SDS)中。收集裂解物,并将其转移到微量离心管中。测量用BCA蛋白试剂的蛋白质浓度根据制造商的方案( 见表1)。分析的GABA A受体通过SDS / PAGE的α1和β3亚基的表达和使用亚基特异性抗体(兔抗α1特异性和兔抗β3特异性的GABA A R抗体的免疫印迹, 见表1对这些信息抗体)。
- 移去并从剩余孔仅传送阳性克隆到较大的6厘米组织培养皿。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换含抗生素的培养基中,每2天。
- 逐步扩大细胞u的殖民地Lander(国家)的抗生素选择通过它们转移到10厘米的组织培养皿中,并最终以组织培养瓶(T-75瓶)。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换含抗生素的培养基中,每2天。
- 板从上盖玻片各菌落70000电池(直径13mm)和固定细胞的免疫荧光分析中的GABA A R亚基在细胞表面的表达和共定位。
- 选择HEK293细胞中表达高水平的GABA A受体,板2×10 6个细胞的两个α1和β3亚基的阳性克隆入10cm的无菌组织培养盘中,在湿润的5%CO 2的气氛中温育,在37°С( CO 2的培养箱)过夜。确保细胞在任何时间下培养在含有抗生素(G418和腐草霉素D1)的平台。
- 第二天,转染的HEK293细胞的GABA A Rγ2s亚基的cDNA中的pCDNA™3.1(+),结合使用非脂质体的脂质转染试剂( 表1)潮霉素B抗性基因的表达载体。
注:此转染法可以更好地生存和外源蛋白的生长缓慢的稳定细胞系都在不断的选择与抗生素高表达效率。 - 在无菌的15毫升离心管中,加入250μl的增强剂和DNA缩合缓冲液( 表1)和γ2sGABA 1.4微克A受体亚单位的cDNA。之前,在室温下放置5分钟加入11.2微升强化剂,涡旋1秒。
- 之前在室温下放置10分钟,加入35微升的非脂质体的脂质转染试剂和旋涡10秒。加入3毫升含D1 G418 /腐草霉素培养基中并吸取的离心管中的内容上下两次BEF矿转让其逐滴到细胞中有10厘米的组织培养板中成长。在潮湿的5%CO 2的气氛中(CO 2培养箱中)孵育细胞48小时,在37°С。
- 用无菌PBS轻轻洗涤细胞,并稀释成新10厘米组织培养皿中,在下面的比率:1:3,1:5,1:7,1:10,1:15,1:20。
- 通过添加的抗生素选择标记潮霉素的800微克/毫升的G418 /腐草霉素的D1-含有培养基开始选择α1的表达γ2s亚基/β3-HEK293细胞。更换旧媒体与新鲜G418 /腐草霉素D1 /含潮霉素B介质(10毫升),每2天。
- 重复G418 步骤2.7-2.12,在不断的选择/腐草霉素D1 /潮霉素B含细胞的培养液中。
- 在不含抗生素的细胞培养基和10%二甲亚砜(DMSO)为将来使用存储的阳性克隆在-140℃。
- 测试水平对α1,β3表达和在下面除霜每个克隆γ2sGABA A R亚基免疫印迹和免疫荧光,因为表达式可以改变由于细胞在抗生素选择存活率降低。
3.维护的HEK293细胞系
- 除霜控制的HEK293细胞的小瓶或那些表达或者α1/β2/γ2-GABA A卢比( 表1)或α1/β3/γ2-GABA A卢比(如上所述)到10ml细胞培养基在15ml的无菌离心管中。离心机在440×g离心5分钟以除去过量的DMSO。
- 在1ml新鲜的细胞培养基中除去上清,重悬细胞。
- 先添加9毫升的新鲜细胞培养基中,以10cm的组织培养皿中涂有聚-D-赖氨酸(0.1毫克/毫升)中加入1ml重悬浮细胞。轻轻搅动,一边到另一侧,以分散细胞并在湿润的温育,在37°С5%CO 2的气氛中(CO 2培养箱)。
- 第二天,吸出,以除去任何细胞碎片,并替换为10ml新鲜的HEK293细胞培养基的培养基。
- 选择稳定的细胞系用抗生素,以消除任何细胞,不表达GABA A R亚基,因此也缺乏抗生素抗性标记物的表达。为α1/β2/γ2的稳定细胞系,用含有G418(800微克/毫升)的新鲜细胞培养基替换正常培养基。为α1/β3/γ2细胞系,替换与含有新鲜细胞培养基的G418(800微克/毫升),腐草霉素D1(800微克/毫升)和潮霉素B(800微克/毫升)。 小心!G418为刺激性潮霉素B具有腐蚀性,毒性和刺激性。
- 通道中的细胞分化成由播种的新组织培养皿中以较低的密度,一旦达到> 70%汇合。抽吸10毫升细胞培养基和两次简要洗的PBS,pH 7.4中。加入1 ml的胰蛋白酶-EDTA溶液中,所述蛋白酶的胰蛋白酶溶液(0.05%胰蛋白酶)和钙离子螯合剂EDTA(0.02%)的PBS,pH为7.4,分离从培养皿的细胞。 小心!胰蛋白酶-EDTA解决方案是一种刺激物。
- 添加含有正确的抗生素10毫升细胞培养基,向培养皿并吸出细胞。离心将细胞在440×g离心5分钟,重悬它们在5毫升细胞培养基中的。
- 通过使用1:10稀释到含有新鲜细胞培养基和正确的抗生素新的组织培养瓶(T-75烧瓶)中的细胞。孵育细胞,在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱中)并更换培养基每两天。通过细胞时> 70%汇合(见步骤2.8)。
4.准备中GABA能神经元刺文化
- 在无菌条件下制备24我们LL板用聚L-赖氨酸(0.1毫克/毫升)包被的盖玻片(13毫米直径),并在湿润的5%CO 2的气氛中(CO 2培养箱中)孵育,在37°С。
- 第二天,吸出物用移液管的过量聚-L-赖氨酸和洗涤盖玻片用两个简短10秒和2 5分钟长的洗涤液用无菌水。加层粘连蛋白(0.01毫克/毫升)过夜,并在湿润的5%CO 2的气氛中(CO 2培养箱中)孵育,在37°С。
- 消毒用70%乙醇的解剖区域和聚集的解剖工具,例如曲线和直线组织钳,剪刀和镊子阵列,并放入70%乙醇,充分消毒的解剖仪器。
- 将怀孕的大鼠/小鼠安乐死,用CO 2在它的后面,并清洁在其腹部用70%乙醇的皮肤。捏用镊子皮肤和切断周围的腹部穿过皮肤,肌肉,腹膜,以显示内部器官和子宫与胚胎清晰。提取子宫中的胚胎(E16-17),并将其放置在培养皿冷冻PBS。
- 将胚胎在层流罩和斩首它们,收集头中一个新的培养皿用冷HBSS中。
- 在解剖显微镜下,解剖出使用了弯曲和直镊子的大脑。把大脑变成含冷冻HBSS一个新的培养皿。
- 分开的两个大脑半球,小心地取出脑膜。沿着海马的线切割和剥离背面的皮质以显示纹状体。观察纹状体的白色条纹状结构,在半球的前。
- 解剖纹状体切成非常小的片(1 - 2毫米的直径),并使用一个火抛光的巴斯德吸液管将材料收集到无菌的15ml离心管中的1ml的总体积。火抛光确保材料被收集而不被破坏。
- 用火抛光钛巴斯德移液管的对,吸出细胞,并释放它们8 - 10倍。取一个新的移液管具有其原始直径(1毫米)的大约30%的火抛光头,磨碎的溶液再进行4 - 6次,直至出现均匀。
- 使用100微米的尼龙细胞过滤到一个新的无菌离心管中过滤细胞。
- 计数细胞,用血细胞计数器和板70000细胞向500微升每孔的神经元培养基中在24孔组织培养板中。搅动井从左到右和在湿润的5%CO 2的气氛中(CO 2培养箱)14天的体外 (DIV),孵育在37℃С。
- 7天后,检查神经细胞的纯度,如果神经胶质细胞存在,加胞嘧啶β-D-阿糖胞苷(阿糖胞苷; 5微米)到孔阻止其扩散。要做到这一点,从每个孔取出250微升的神经元培养液(pH值7.4)中,加入250微升的新鲜培养基含有ING阿糖胞苷。 小心!阿糖胞苷是一种有刺激性。
5.共培养制备
- 对神经元培养的第11天,外套一个6孔板用多聚-D-赖氨酸(0.1毫克/毫升)在无菌条件下过夜孵育,在37°С在湿润的5%CO 2的气氛中(CO 2培养箱)。
- 翌日(神经元培养的第12天),吸去多余的聚-D-赖氨酸和用少量加入新鲜的细胞培养基(无抗生素),以涂层的孔中之前5分钟,用无菌水洗涤2次孔简要和2x的血清的培养基。
- 吸出从组织培养瓶(T-75)的培养基中。加入1 ml的Ca 2+前冲洗细胞,用PBS两次/镁离子螯合剂乙二胺四乙酸(0.48毫米),它轻轻地和非解离酶的细胞。搅动细胞从组织烧瓶底部分离它们。
- 加入10 mL的细胞培养培养基(不含抗生素,因为这可以与转染干扰),吸在无菌的离心管中的细胞和地点。沉淀细胞,在440×g离心使用低速台式离心机5分钟。
- 去除上清并将细胞重新悬浮在1ml新鲜的细胞培养基中。使用血细胞计数器,计数细胞和板以每孔3×10 5个细胞于6孔板中的密度。轻轻搅动,并在湿润的5%CO 2的气氛中(CO 2培养箱中)孵育24小时,在37°С。
- 翌日(神经元培养的第13天),瞬时利用脂质体转染试剂转染中的pcDNA3表达构建体的HEK293细胞与mCherry的cDNA。简要地说,在无菌的离心管中,加入500μl的低血清培养基和5微克mCherry的cDNA。加入5微升脂质体缓冲试剂和前在室温下放置5分钟,轻轻混匀。
- 添加8.75微升脂质体转染REAG之前,在室温下放置30分钟耳鼻喉科,轻轻混匀。吸取微量离心管中的内容上下两次,转印逐滴向每孔中的6孔组织培养板,孵育在37℃С在湿润的5%CO 2的气氛中(CO 2培养箱)。
- 翌日(神经元培养的第14天),吸从每个6孔的HEK293细胞的培养基并用PBS(pH 7.4)中洗涤各孔两次短暂。添加300微升的Ca 2+ / Mg 2 +的螯合试剂的EDTA(0.48毫摩尔)和200μl的胰蛋白酶-EDTA(0.02 - 0.48毫摩尔)的溶液,以每孔中并孵育37°С5分钟。
- 加入1 ml,每孔新鲜的HEK293细胞培养基(此猝灭胰蛋白酶)和吸出分离的细胞到无菌的15ml离心管中。离心将细胞在440×g离心5分钟,在室温下,去除上清。重悬在500μl的神经元培养液(pH7.4)中的沉淀。
- 使用血细胞计数器,计数细胞和种子以每孔30,000个细胞在含有神经元的24孔组织培养板的密度。搅动板以分散细胞,并在湿润的5%CO 2的气氛中温育共培养物,在37°С(CO 2培养箱中)放置24小时。
突触联系及其活动6.分析
- 经过23小时的共培养,研究了γ-氨基丁酸能中型多棘神经元,并使用抗突触管腔结构域特异性抗体缀合了荧光染料的活性依赖性摄取HEK293细胞之间"有效"接触地层(Cy5的, 见表1) 。
注:该抗体只获得对突触结合蛋白的腔结构域,获得与其结合时存在的突触小泡腔和外空间之间的连续性。该神经递质释放过程中具体情况,制定本antibODY活性的突触前末梢的良好标志物。 - 首先漂洗共培养与神经元培养基(神经基础的培养基,pH值为7.4; 参见表1),并添加Cy5标记的小鼠抗突触结合蛋白抗体,在神经元的培养基以1:50稀释(的Neurobasal的培养基,pH7.4)中,向培养物,进行30分钟。在这段时间内,在37°С在湿润的5%CO 2的气氛中(CO 2培养箱中)孵育细胞。
- 以除去抗体的访问,洗净共培养简要三次:第一次用冷正常的PBS(pH 7.4)中,第二用冷的PBS(pH7.4)中含有200mM NaCl和第三冷正常的PBS(pH 7.4)中
- 固定的细胞用300μl的4%多聚甲醛/ 4%蔗糖的PBS中(PFA,pH7.4)中10分钟,搅拌。简要地两次,用PBS(pH7.4)中,然后用两个长10分钟的洗涤洗细胞。
- 添加甘氨酸(0.3 M),向每孔中10分钟,在搅拌下,淬灭的PFA。
- 洗细胞brieflÝ两次,用PBS(pH7.4)中,然后用两个长10分钟,洗涤之前,加入封闭溶液(1%(重量/体积)牛血清白蛋白(BSA)的PBS,pH 7.4)中,以减少非特异性结合的300微升抗体。
- 吸去封闭溶液并添加豚鼠抗GABA A R-γ2抗体针对γ2N-末端结构域33(1:3000于PBS中,pH 7.4)中4℃过夜°С。
- 第二天,吸出从孔中的第一抗体,用PBS简要洗细胞两次,然后用两个长10分钟的洗涤。
- 透化在封闭溶液中30分钟,在室温下使用的Triton X-100(0.1%)的细胞。
- 简要地两次,用PBS(pH7.4)中,然后用两个长10分钟的洗涤添加任一所述的小鼠抗谷氨酸脱羧酶之前洗涤细胞(GAD)65抗体(1:4000, 表1)或小鼠抗突触蛋白I抗体( 1:1000, 表1)为120分钟,在室温吨emperature。
- 然后用PBS(pH 7.4)中简短地洗涤细胞两次有两个长10分钟,洗涤,然后加入封闭液中30分钟,在室温下进行。
- 离心机的适当的二抗(典型的山羊抗豚鼠IgG缀合Cy5标记山羊抗小鼠IgG缀合的Alexa Fluor 488山羊抗小鼠IgG的Alexa Fluor 405,所有在2微克/毫升)以除去抗体的聚集体在21910×g离心10分钟,并加入抗体(1:750),以封闭液。适用于适当的孔中1小时,并用铝箔覆盖,以保护荧光团由曝光和随后的光漂白。
- 最后,用PBS(pH 7.4)中简短地洗细胞两次,然后用两个长10分钟的洗涤以除去任何未结合的第二抗体,并装入用固定试剂(延长黄金, 表1)每盖玻片大约10微升的盖玻片。允许24小时,设定为室温,同时转印到4前避光°С用于长期存储。
- 分析使用激光扫描共聚焦显微镜与63X油浸物镜样本。保证光线水平和检测器的增益被调整,以避免饱和。
- 观察潜在突触样接触的突触前末梢的正面为GAD65,突触蛋白I或Cy5标记的抗突触结合蛋白和突触后的HEK293细胞通过DIC或由mCherry荧光指示剂可视化之间的共定位的区域。
- 计数采用Z堆栈系列的光学部分(8 - 10)潜在的突触样接触过的4深度 - 每个单元5微米用图像处理软件。
结果
该协议为这个神经元的HEK293细胞共培养模型系统已被细微调整,以允许最佳的细胞存活。在该系统中,形成的突触样接触和它们的分析依赖于对所有三种的GABA A R亚基其中组装成一个功能性受体稳定和一致的表达。因此重要的是,以将其添加到神经元培养物之前,在HEK293细胞中的表面用免疫细胞化学分析,以测试对于亚单位的表达。在这些实验中,使用了结合于这些亚基的胞表位的亚基特异性抗体检测的α1,β2和γ2亚基( 图1A),或α1,β3和γ2亚基( 图1B)的细胞表面表达。这些亚单位之间的共定位在HEK293细胞表面的高度被证明。
确认GABA A的表面表达和共定位后在HEK293细胞,共培养ř亚基采用HEK293细胞表达α1/β2/γ2GABA A R亚基和中型多棘神经元的14天(14天体外(DIV))培养的准备。在共培养的细胞孵育24小时,固定,用免疫细胞化学和共焦显微镜分析。触头的分析表明,谷氨酸脱羧酶阳性GABA能轴突末梢仅形成与对照的HEK293细胞( 图2A,2B)零星的接触。在4小时检测出的联系人的数量为7.3±0.9的HEK293细胞,并且这个数目被添加的HEK293细胞的培养的神经元后降低至5.5±0.5的连接(平均值±SEM)的每HEK293细胞在24小时左右。相比之下,GAD65阳性GABA能轴突末梢形成HEK293细胞表达GABA A卢比无数突触样联系。加入HEK293细胞后,在4小时获得联系人的数量为28.3±4.7 HEK293细胞,而这一数字进一步上升d,来52.1±6.3(平均值±SEM)的每HEK293细胞在24小时共培养( 图2A,2B)。
为了确定这些突触样接触是否是'活性', 即支持囊泡递质释放,一个泡囊的腔结构域特异性抗突触Cy5偶联的抗体加入到共同培养介质23小时的孵育后。该抗体只结合到突触前神经末梢当突触小泡腔和神经递质的释放过程中,在突触间隙中的细胞外流体之间的孔的形式。以下释放,孔关闭,而使突触Cy5偶联荧光抗体附着于突触小泡内。在这种方式中,只有积极参与神经递质释放的囊泡标记抗体。在这些实验中几乎没有的HEK293细胞和对照之间有任何接触的中型多棘神经元的终端是'活性’如由缺乏突触前谷氨酸脱羧酶/突触荧光和mCherry荧光在HEK293细胞( 图3A)之间的共定位。与此相反,许多"有效"接触者的中型多棘神经元终端和α1/β2/γ2表达的HEK293细胞中,所揭示的GAD 65 /突触结合蛋白和mCherry之间的共定位的程度高,特异性表达在HEK293细胞(间形成图3B)。
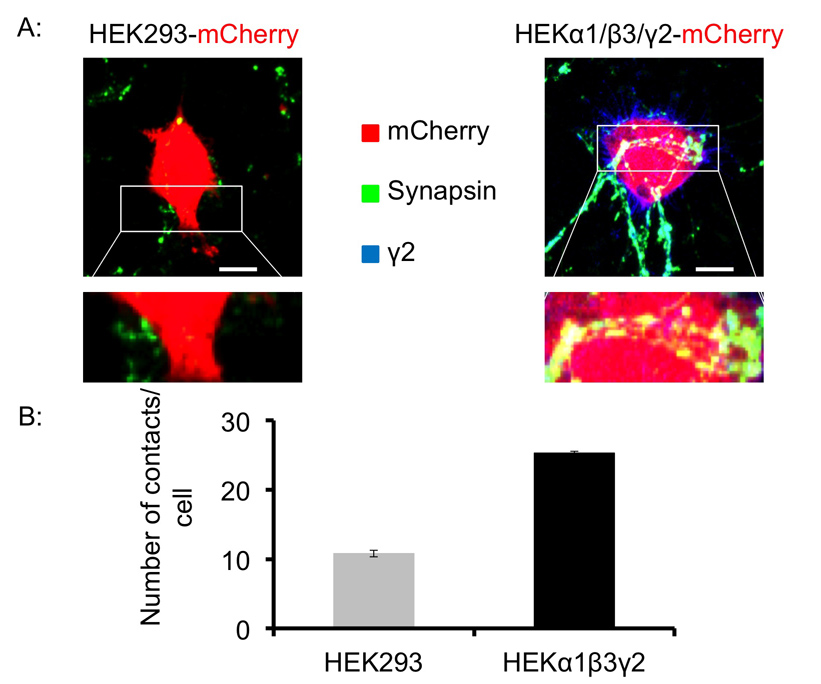
为了测试是否不同亚型GABA A R也可以促进突触样形成的影响,我们已经共同培养α1/β3/γ2表达HEK293细胞,中型多棘神经元。再次,控制HEK293细胞很少接受与突触蛋白阳性的突触前末梢到达触点10.8±0.48(平均值±SEM)的每HEK293细胞接触24小时,在共培养物( 图4A 左 ,4B)之后。然而,HEK293细胞表达α1/β3/γ2 的 GABA A卢比形式显著更具有中等棘状神经元到达每HEK293细胞25.3±0.27(平均值±SEM)的接触24小时,在共培养后的突触蛋白阳性的突触前末梢( 图4A 右突触样接触图4B)。这表明,α1/β3/γ2 的 GABA A卢比表达在HEK293细胞中也能促进突触形成接触,尽管它们的效力比α1/β2/γ2含GABA A Rs的效力较低。
这些实验表明,在我们的实验室中研制的共培养模型系统允许突触接触形成的影响的定量分析的 GABA A卢比在这个过程中的不同亚型的效力以及评价。这些实验还表明,GABA A Rs时,除了是GABA能突触的关键功能部件,可发挥关键作用在识别和形成的抑制性神经元的独立的其他突触粘附蛋白和相应的神经细胞的靶细胞,间突触联系的过程。

图1.免疫细胞的GABA A Rα1表达的分析/β2/γ2或α1/β3/γ2在稳定的HEK293细胞系。抗体识别的GABA A R亚基的胞外结构域被用来表达在细胞表面标记的受体。( A)的 HEK293细胞系表达α1(的Alexa Fluor 488),β2(的Alexa Fluor 555)和γ2(Cy5标记),在高的水平。(B)的 HEK293细胞系表达α1(的Alexa Fluor 488),β2(的Alexa Fluor 555)和γ2 (Cy5的)亚基在高水平。比例尺:10微米,请点击这里查看该图的放大版本。
{kind=link}

图2. GABA能中型多棘神经元形成具有α1/β2/γ2-在共培养物表达的HEK293细胞突触样接触。 (A)与抗GAD65抗体的突触前末梢的荧光标记(绿色)和HEK293细胞与mCherry(红色, 左 )或与GABA A Rγ2亚基(蓝色, 右 ),表明它们之间的共定位的点标记指示后在共培养4或24小时,形成的突触样接触。比例尺:10μm以下(B)的突触样共定量分析 ntacts。 HEK293细胞根据它们的形状被确定为揭示由DIC的成像和/或mCherry表达,GAD-65阳性泪点(绿色)和HEK293细胞的表面之间的接触的数目是由眼睛的各光学部分在计数Z堆叠系列(9 - 10),每使用成像软件细胞,并表示为接触/细胞的数量。该图显示后在共培养4小时和24小时的中型多棘神经元与对照的HEK293细胞(浅灰色)或α1/β2/γ2-HEK293细胞(黑)之间的接触的数目(平均值±SEM,每组中的每个8从两个独立的实验条件)。这个数字已经被修改Fuchs 等人(2013年)30。 请点击这里查看该图的放大版本。
{kind=link}
广告/ 52115 / 52115fig3highres.jpg"WIDTH ="600"/>
图3. GABA A卢比推广 24小时中型多棘神经元的终端呈阳性GAD65间共培养后所形成突触样接触(的Alexa Fluor 405青色)和(A)的控制HEK293细胞形成积极的突触联系。免疫标记,或(B)中的HEK293-α1/β2/γ2细胞,无论瞬时转染mCherry构造(红色)。有源触点由囊泡腔结构域特异性抗突触结合蛋白抗体(Cy5标记)和GAD 65特异性抗体都在突触前末梢之间的共定位标识,mCherry表达在HEK293细胞中。比例尺:10微米,请点击这里查看该图的放大版本。
{kind=link}
igure 4"FO:内容宽度="6英寸"的src ="/文件/ ftp_upload / 52115 / 52115fig4highres.jpg"宽度="600"/>
图4. GABA能中型多棘神经元形成具有α1/β3/γ2-亚基表达的HEK293细胞在共培养突触样接触。 (A)与抗突触蛋白I抗体(绿色),和控制HEK293细胞(左),或α1突触前末梢的荧光标记/β3/γ2表达HEK293细胞中,无论是瞬时转染mCherry(红色),显示的分这些标志物,指示24小时共培养后形成的突触样接触之间的共定位。比例尺:10μm的(B)中 。 突触样接触的定量分析。将HEK293细胞以mCherry表达进行了鉴定,并且突触蛋白I阳性泪点(绿色)和HEK293细胞的表面之间的接触的数目是由眼睛的Z堆叠系列,每个光学部分在计数(8 - 10)每使用成像软件细胞,并表示为接触/细胞的数量。图中示出了24小时,在共培养后中型多棘神经元与对照的HEK293细胞(浅灰色)或α1/β3/γ2-HEK293细胞(黑)之间的接触的数目(平均值±SEM,每组8-12细胞中每个条件,由两个独立的实验)。 请点击这里查看该图的放大版本。
{kind=link}
讨论
虽然这个协议不是技术上难以进行,有必须遵循以达到最准确和可重复的共培养测定几个关键步骤。首先,培养的中型多棘神经元必须在最佳密度接种。如果接种过疏,神经元倾向于发展非常缓慢和存活大大降低。另一方面,如果晶种太密,神经细胞倾向于聚集其中损害与HEK293细胞接触的分析。其次,建议以瞬时表达荧光报道,GFP或mCherry在HEK293细胞中稳定表达的 GABA A Rs时,platting成共培养之前。这允许可靠识别的HEK293细胞,其可以通过相似于这些细胞,并在培养的神经元存活的罕见神经胶质细胞之间的形状和尺寸会受到影响的。为了实现高效的转染GFP或mCherry的cDNA,将HEK293细胞系必须是在指数生长期和接种在6孔板的适当的密度。稀疏接种随后转染会引起细胞生长不良,而在接种将防止细胞从占用该cDNA。理想情况下,电池应该接种,使他们70之间 - 对转染当天90%汇合。第三,转染,必须为每个使用的细胞系进行优化,因为一些细胞系是比其他人更敏感。这是因为构GABA在HEK293细胞中阿 R的表达减少的细胞存活和细胞转染后,以恢复的能力。此外,生存取决于所表达的HEK293细胞的 GABA A Rs中的类型,其中一些细胞系是显著比其他人更敏感。采用脂质体转染试剂是用于表达外源蛋白在快速增长的细胞系,同时提供了较高的转染效率和表达水平的最佳方法。然而,这种试剂会导致生长缓慢的细胞系造成太大的损失,为这是我们经常使用的非脂质体转染试剂。这个工作在一个类似的方式向脂质体试剂但DNA所需的有效转染的量显著降低。这允许更大的细胞存活(约80 - 90%的脂质体试剂60%的使用相比),但具有较低的转染效率(60%)。最后,控制的HEK293或α1数/β2/γ2的HEK293表达加入到神经元培养细胞需要被优化。添加太少细胞损害HEK293细胞和神经元之间的联系的分析成功,是因为他们变得非常罕见。相反,加入过多的HEK293细胞导致在数小时内神经元细胞死亡。
胚胎中型多棘神经元培养理想情况下应该使用纹状体组织从胚胎15岁解剖准备 - 17。然而,经常发生胚胎是不是最佳的年龄稍微年轻或年长的。在这种情况下,神经元的数目在培养接种将需要进行改变。组织是年龄小于E15可能需要在稍低的密度被接种,而组织中超过E17中,可能需要以更高的密度进行接种,以使最佳的细胞存活。此外,阿糖胞苷(阿糖胞苷)可能需要被添加到旧的培养物,以防止生长的神经胶质细胞的,这是在较旧的组织更丰富。
当建立共培养物,它在板转染的HEK293或α1/β2/γ2的HEK293表达细胞的最佳数量是很重要的,如上所述。然而,可能有必要确定,因为在他们的存活差异这为每个单独的细胞系。典型的30,000个细胞中的50微升的最大体积应该被添加到24孔皿中,其中已含有500μl的神经元培养基中的每个孔中,因为这确保了空调的神经介质没有稀释太多,该条件在每个孔中保持相对稳定, 如 的生长因子的浓度。添加量超过50微升,每孔通常会杀死神经元。
一对共培养技术的主要缺点之一是,神经元培养物从生长为单层,这意味着该神经元已经从它们的正常微环境取出并不能确定其正常的解剖组织离解的细胞产生。因此,它们缺乏适当的连接,输入和分泌的分子来自其他小区可影响突触发育的最初阶段。例如, 在体内中型多棘神经元的密集支配由谷氨酸能输入从皮 质,丘脑和其他脑区域34,然而,在我们的培养的神经元谷氨酸突触不会形成,因为这些输入的纹状体组织的解剖过程中损坏。如何缺乏功能谷氨酸突触培养中型多棘神经元AFFECTS他们形成GABA能突触彼此和/或HEK293细胞表达GABA A卢比的能力仍然是一个悬而未决的问题。这个问题可以通过中型多棘神经元与皮质谷氨酸能神经元,从而使他们能够形成功能性突触35,然后加入HEK293细胞一起培养很容易地得到解决。另一种方法是设计一个共培养模型系统的基础上器官切片文化,以维持其中一些可能是成熟和突触形成的重要的细胞结构。然而,器官切片文化有密集的,异构的神经纤维,可危及在这里进行的分析。使用共培养测定的另一个重要缺点是GABA A卢比表达在HEK293细胞中未被聚类,因为它们是在神经细胞的表面上,尽管这似乎不是必需给予足够高的表面表达30突触的形成。例如,在第rodent大脑和海马文化中,α1GABA A R亚基在锥体细胞突触后的所有领域最GABA能突触中。然而,α2是专门设在对胞体突触和树突的一个子集,但高度浓缩的轴突起始段,所揭示的免疫荧光显微镜和电子显微镜36。鉴于突触形成的共培养物仍可以可靠地检测和分析30,这表明的GABA A的Rs的HEK293细胞的细胞表面上的密度可以类似于或甚至高于内突触这些受体的密度集群中的神经元。这可以解释,至少部分地,为何突触粘附蛋白如人类neuroligin,和突触后密度蛋白,如gephyrin时,没有必要对突触形成的共培养物中,如果适当地组装的 GABA A卢比存在于足够密度。
这是有据可查的GABA A卢比在结构和功能上异构,并且该受体亚基组成决定了他们的亚细胞定位和药理特性。例如,2亚基的结合被称为是对GABA A卢比突触本地化的先决条件,而亚基几乎全部存在于突触外GABA A卢比。结合了只αβ组合的受体也认为是主要定位于突触外结构域12-14。此特异性是否保持在我们的共同培养系统可通过瞬时转染2或亚单位的cDNA转染HEK293细胞系中稳定表达α和β亚基组成,将它们加入到神经元培养物之前,可以很容易地测试。我们使用这种方法的初步实验已经表明,突触联系仅在2亚基的存在下容易地形成,表明该特定ficity 体内观察到有可能在体外被保留(数据未显示)。
此外,GABA A卢比结合不同的α亚基选择性地定位于特定类型的突触前神经元形成突触联系。例如,在苍白球中,α1-GABA A卢比通常在striatopallidal(海峡-GP)和palliopallidal(GP-GP)的突触,它们位于树突和中等棘状神经元的体细胞的区域,分别找到。该α3-GABA A卢比位于中型多棘神经元perisomatic地区和当地的GP轴突侧支接触,而α2-GABA A卢比位于这些神经元的树突远端和纹状体32主要通过接触输入。在不同类型的突触,并在不同的神经元的隔间具体α亚单位的表达也已经证明在其他脑区如HIPPocampus 21和新皮质18,20。这些发现提出的问题是,如何在特定抑制性突触形成在大脑中。不突触前末梢的一个特定类型的附着引起的特定的GABA A R亚型插入的接触点?是受体贩卖到特定的亚细胞定位,根据自己的亚基组成,他们的细胞膜的插入是针对特定的原产地终末的附着力的先决条件?迄今为止,这些问题仍然没有答案。采用简化模型系统,如共培养模型系统,让我们开始回答这个复杂的问题,因为该系统很容易服从的DNA结构和应用试剂转染,更重要的,它是适合活细胞成像分析30。因此,使用这个模型系统,我们可以开始测试单个分子,包括不同类型的GABA A卢比的作用,知道电量为出席突触联系。另一个优点是,突触在该模型系统迅速形式,在几分钟之内,以小时,减少了实验的持续时间。类似的共培养模型系统被成功地使用,在过去筛选新颖的突触发生的分子27,37,38。
理解中枢神经系统如何发展,成熟和神经元以便错综复杂控制,例如,行为或认知之间形成连接,具有根本的重要性。这个遥远的目标将仅由划定支配发育过程中识别和细胞 - 细胞通信的各个步骤的分子机制来实现。由于纯粹的复杂性,这些多细胞相互作用的分子机制目前可以仅在减少系统的研究与精度。然而,能够通过表达蛋白质的多种组合和研究如何提高这些系统的复杂性它们相互作用具有与,例如,基因缺失的方法相比有一些优点。这是因为单个基因缺失的影响的准确的解释通常是由与屏蔽原始病变的影响,尤其是在发育中的大脑代偿机制相关的变化受到损害。这里所描述的简单而翔实的共培养技术已经允许发现的GABA A卢比在突触形成的结构性作用,开辟了可能性,研究如何GABA A R和其他细胞粘附分子和/或突触基质蛋白互相交流在突触。突触基质蛋白是因为它们最近被证实发挥谷氨酸能突触形成39了关键的作用特别令人感兴趣的。共培养模型的进一步发展是非常重要的,因为它们具有潜在的推进我们哪个引导"正常和分子机制的知识#8217;脑发育,从而提高对这些机制是怎样改变的许多神经发育疾病,如癫痫,精神分裂症,孤独症谱系障碍和其他许多人理解。
披露声明
The authors declare that they have no competing financial interests.
致谢
我们要感谢来自英国MRC(G0800498)的资金支持。我们还要感谢JM Fritschy教授,苏黎世大学,提供的GABA A -R亚基特异γ2抗体和R·哈维教授,伦敦大学学院药剂学院,提供含有抗生素抗性的pCDNA 3.1(+)表达载体基因生产的稳定转染的HEK293细胞系。
材料
| Name | Company | Catalog Number | Comments |
| DMEM (Dulbecco's Modified Eagle Medium) | Life Technologies | 11960-044 | Warm in water bath at 37 °C before use |
| L-Glutamine | Life Technologies | 25030-024 | |
| Penicillin/Streptomycin | Life Technologies | 15070-063 | Danger: irritant |
| FBS (Fetal Bovine Serum) | Life Technologies | 10106-169 | |
| Neuralbasal | Life Technologies | 21103-049 | Warm in water bath at 37 °C before use |
| B27 Supplement | Life Technologies | 17504-044 | |
| Glucose | Sigma-Aldrich | G8769 | |
| HBSS (10X) | Life Technologies | 14180-046 | |
| HEPES (1M) | Life Technologies | 15630-056 | |
| PBS (1X) | Life Technologies | 10010-031 | |
| pcDNA 3.1(+) Mammalian Expression Vector (Geneticin-selection) | Life Technologies | V790-20 | |
| pcDNA 3.1(+) Mammalian Expression Vector (Zeocin-selection) | Life Technologies | V860-20 | |
| pcDNA 3.1(+) Mammalian Expression Vector (Hygromycin-selection) | Life Technologies | V870-20 | |
| Stable HEKα1β2γ2 line | Sanofi-Synthelabo, Paris | ||
| Poly-D-lysine | Sigma-Aldrich | P1149 | |
| G418 disulfate salt (Geneticin) | Sigma-Aldrich | G5013 | Danger: irritant |
| Phleomycin D1 (Zeocin) | Life Technologies | R25001 | |
| Hygromycin B | Life Technologies | 10687-010 | Danger: toxic, irritant and corrosive |
| Trypsin-EDTA | Life Technologies | 25300-054 | Warm in water bath at 37 °C before use. Danger: irritant. |
| Poly-L-lysine | Sigma-Aldrich | P6282 | |
| Laminin | Sigma-Aldrich | L2020 | |
| 100 μm Nylon Cell Strainer | VWR | 734-0004 | |
| Cytosine β-D-arabinofuranoside (Ara-C) | Sigma-Aldrich | C1768 | Danger: Irritant |
| Chelating agent (Versene) | Life Technologies | 15040-033 | |
| Liposomal transfection reagent (Lipofectamine LTX) with liposomal transfection buffering reagent (PLUS reagent). | Life Technologies | 15338-100 | Alternative transfection method: Effectene Reagent |
| Non-liposomal transfection reagent (Effectene reagent) | Qiagen | 301425 | |
| Reduced serum medium (Opti-MEM) | Life Technologies | 11058-021 | |
| Mouse anti-Synaptotagmin antibody conjugated to Cy5 | Synaptic Systems | 105311C5 | |
| Neurobasal A | Life Technologies | 10888-022 | |
| Sodium Chloride (NaCl) | VWR | 27810.364 | |
| Glycine | Sigma-Aldrich | G1726 | |
| BSA (Bovine Serum Albumin) | Sigma-Aldrich | A3294 | |
| Guinea pig anti-γ2 GABAA receptor antibody | Prof. Jean Marc Fritschy | N/A | (Institute of Zurich, Switzerland) Fritschy, JM and Mohler, H. J. Comp. Neurol. 359 (1), 154-194 (1995). |
| Triton X-100 | Promega | H5141 | |
| Mouse anti-Glutamate Decarboxylase (GAD)65 antibody | Merck Millipore | MAB351 | |
| Mouse anti-synapsin antibody | Synaptic Systems | 106-011 | |
| Mouse anti-β2/3 antibody (BD17) | Merck Millipore | MAB341 | |
| Rabbit anti-α1 GABAA receptor antibody | Professor Anne Stephenson | N/A | (UCL School of Pharmacy, London) FA Stephenson et al. J. Comp. Neurol. 416 (2), 158-172 |
| Goat anti-guinea pig conjugated to Cy5 antibody | Merck Millipore | AP1085 | |
| Goat anti-mouse Alexa Fluor 488 antibody | Merck Millipore | AP124S | |
| Goat anti-mouse Alexa Fluor 405 antibody | Life Technologies | A31553 | |
| Goat anti-mouse Alexa Fluor 555 antibody | Life Technologies | A21422 | |
| Goat anti-rabbit Alexa Fluor 488 antibody | Life Technologies | A11008 | |
| Mounting reagent (Prolong Gold) | Life Technologies | P36930 | Use at room temperature |
参考文献
- Wang, D. D., Kriegstein, A. R. Defining the role of GABA in cortical development. J Physiol. 587, 1873-1879 (2009).
- Ben-Ari, Y., Khalilov, I., Kahle, K. T., Cherubini, E. The GABA excitatory/inhibitory shift in brain maturation and neurological disorders. The Neuroscientist : a review journal bringing neurobiology, neurology and psychiatry. 18, 467-486 (2012).
- Structure Sieghart, W. pharmacology, and function of GABAA receptor subtypes. Adv Pharmacol. 54, 231-263 (2006).
- Unwin, N. Neurotransmitter action: opening of ligand-gated ion channels. Cell. 72, 31-41 (1993).
- Homanics, G. E., et al. Mice devoid of gamma-aminobutyrate type A receptor beta3 subunit have epilepsy, cleft palate, and hypersensitive behavior. Proceedings of the National Academy of Sciences of the United States of America. 94, 4143-4148 (1997).
- Gunther, U., et al. Benzodiazepine-insensitive mice generated by targeted disruption of the gamma 2 subunit gene of gamma-aminobutyric acid type A receptors. Proceedings of the National Academy of Sciences of the United States of America. 92, 7749-7753 (1995).
- Mohler, H. GABA(A) receptor diversity and pharmacology. Cell Tissue Res. 326, 505-516 (2006).
- Low, K., et al. Molecular and neuronal substrate for the selective attenuation of anxiety. Science. 290, 131-134 (2000).
- Rudolph, U., et al. Benzodiazepine actions mediated by specific gamma-aminobutyric acid(A) receptor subtypes. Nature. 401, 796-800 (1999).
- Rudolph, U., Knoflach, F. Beyond classical benzodiazepines: novel therapeutic potential of GABAA receptor subtypes. Nat Rev Drug Discov. 10, 685-697 (2011).
- Hulst, C., Atack, J. R., Kooy, R. F. The complexity of the GABAA receptor shapes unique pharmacological profiles. Drug Discov Today. 14, 866-875 (2009).
- Essrich, C., Lorez, M., Benson, J. A., Fritschy, J. M., Luscher, B. Postsynaptic clustering of major GABAA receptor subtypes requires the gamma 2 subunit and gephyrin. Nat Neurosci. 1, 563-571 (1998).
- Schweizer, C., et al. The gamma 2 subunit of GABA(A) receptors is required for maintenance of receptors at mature synapses. Mol Cell Neurosci. 24, 442-450 (2003).
- Belelli, D., et al. Extrasynaptic GABAA receptors: form, pharmacology, and function. The Journal of neuroscience : the official journal of the Society for Neuroscience. 29, 12757-12763 (2009).
- Connolly, C. N., Wooltorton, J. R., Smart, T. G., Moss, S. J. Subcellular localization of gamma-aminobutyric acid type A receptors is determined by receptor beta subunits. Proceedings of the National Academy of Sciences of the United States of America. 93, 9899-9904 (1996).
- Connolly, C. N., Krishek, B. J., McDonald, B. J., Smart, T. G., Moss, S. J. Assembly and cell surface expression of heteromeric and homomeric gamma-aminobutyric acid type A receptors. J Biol Chem. 271, 89-96 (1996).
- Klausberger, T., Roberts, J. D., Somogyi, P. Cell type- and input-specific differences in the number and subtypes of synaptic GABA(A) receptors in the hippocampus. The Journal of neuroscience : the official journal of the Society for Neuroscience. 22, 2513-2521 (2002).
- Thomson, A. M., Jovanovic, J. N. Mechanisms underlying synapse-specific clustering of GABA(A) receptors. Eur J Neurosci. 31, 2193-2203 (2010).
- Fritschy, J. M., Panzanelli, P., Tyagarajan, S. K. Molecular and functional heterogeneity of GABAergic synapses. Cell Mol Life Sci. 69, 2485-2499 (2012).
- Ali, A. B., Thomson, A. M. Synaptic alpha 5 subunit-containing GABAA receptors mediate IPSPs elicited by dendrite-preferring cells in rat neocortex. Cereb Cortex. 18, 1260-1271 (2008).
- Thomson, A. M., Bannister, A. P., Hughes, D. I., Pawelzik, H. Differential sensitivity to Zolpidem of IPSPs activated by morphologically identified CA1 interneurons in slices of rat hippocampus. Eur J Neurosci. 12, 425-436 (2000).
- Nyiri, G., Freund, T. F., Somogyi, P. Input-dependent synaptic targeting of alpha(2)-subunit-containing GABA(A) receptors in synapses of hippocampal pyramidal cells of the rat. Eur J Neurosci. 13, 428-442 (2001).
- Treutlein, B., Gokce, O., Quake, S. R., Sudhof, T. C. Cartography of neurexin alternative splicing mapped by single-molecule long-read mRNA sequencing. Proceedings of the National Academy of Sciences of the United States of America. 111, 1291-1299 (2014).
- Siddiqui, T. J., Craig, A. M. Synaptic organizing complexes. Current opinion in neurobiology. 21, 132-143 (2011).
- Tanaka, H., et al. Higher-order architecture of cell adhesion mediated by polymorphic synaptic adhesion molecules neurexin and neuroligin. Cell reports. 2, 101-110 (2012).
- Krueger, D. D., Tuffy, L. P., Papadopoulos, T., Brose, N. The role of neurexins and neuroligins in the formation, maturation, and function of vertebrate synapses. Current opinion in neurobiology. 22, 412-422 (2012).
- Scheiffele, P., Fan, J., Choih, J., Fetter, R., Serafini, T. Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons. Cell. 101, 657-669 (2000).
- Graf, E. R., Zhang, X., Jin, S. X., Linhoff, M. W., Craig, A. M. Neurexins induce differentiation of GABA and glutamate postsynaptic specializations via neuroligins. Cell. 119, 1013-1026 (2004).
- Dong, N., Qi, J., Chen, G. Molecular reconstitution of functional GABAergic synapses with expression of neuroligin-2 and GABAA receptors. Mol Cell Neurosci. 35, 14-23 (2007).
- Fuchs, C., et al. GABA(A) receptors can initiate the formation of functional inhibitory GABAergic synapses. Eur J Neurosci. 38, 3146-3158 (2013).
- Ventimiglia, R., Lindsay, R. . Culturing nerve cells: Rat striatal neurons in low-density, serum-free culture. , 371-393 (1998).
- Gross, A., et al. Differential localization of GABA(A) receptor subunits in relation to rat striatopallidal and pallidopallidal synapses. Eur J Neurosci. 33, 868-878 (2011).
- Fritschy, J. M., Mohler, H. GABAA-receptor heterogeneity in the adult rat brain: differential regional and cellular distribution of seven major subunits. J Comp Neurol. 359, 154-194 (1995).
- Doig, N. M., Moss, J., Bolam, J. P. Cortical and thalamic innervation of direct and indirect pathway medium-sized spiny neurons in mouse striatum. The Journal of neuroscience : the official journal of the Society for Neuroscience. 30, 14610-14618 (2010).
- Lalchandani, R. R., Vicini, S. Inhibitory collaterals in genetically identified medium spiny neurons in mouse primary corticostriatal cultures. Physiological reports. 1, (2013).
- Nusser, Z., Sieghart, W., Benke, D., Fritschy, J. M., Somogyi, P. Differential synaptic localization of two major gamma-aminobutyric acid type A receptor alpha subunits on hippocampal pyramidal cells. Proceedings of the National Academy of Sciences of the United States of America. 93, 11939-11944 (1996).
- Linhoff, M. W., et al. An unbiased expression screen for synaptogenic proteins identifies the LRRTM protein family as synaptic organizers. Neuron. 61, 734-749 (2009).
- Pettem, K. L., Yokomaku, D., Takahashi, H., Ge, Y., Craig, A. M. Interaction between autism-linked MDGAs and neuroligins suppresses inhibitory synapse development. J Cell Biol. 200, 321-336 (2013).
- Wit, J., et al. Unbiased discovery of glypican as a receptor for LRRTM4 in regulating excitatory synapse development. Neuron. 79, 696-711 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。