
Method Article
用于学习和记忆测试的低成本、气味奖励关联任务
摘要
气味奖励、联想学习任务用于研究生理作对长期和短期记忆的不同影响。
摘要
对于对记忆的细胞和分子机制感兴趣的研究人员来说,稳健而简单的食欲、联想记忆行为范式至关重要。在本文中,描述了一种有效且低成本的小鼠行为方案,用于检查生理作(例如注入药物)对气味奖励记忆的学习率和持续时间的影响。一项研究酪氨酸激酶受体活性在短期 (STM) 和长期记忆 (LTM) 中的不同作用的研究提供了代表性结果。雄性小鼠被条件反射将奖励(糖丸)与两种气味中的一种相关联,并在 2 或 48 小时后测试它们对关联的记忆。在训练之前,将酪氨酸激酶 (Trk) 受体抑制剂或载体输注输送到嗅球 (OB) 中。虽然输注对学习率没有影响,但 OB 中 Trk 受体的阻断选择性损害了 LTM (48 h),而不是短期记忆 (STM; 2 h)。LTM 损伤归因于通过挖掘时间长度测量的气味选择性降低。该实验结果的高潮表明,OB 中的 Trk 受体激活是嗅觉记忆巩固的关键。
引言
联想记忆形成的机制以前主要基于一次试验的恐惧条件反射研究进行研究。然而,许多平凡的任务通常具有更复杂的获取模式,并且依赖于重复的遭遇。该协议的目标是提供一种具有成本效益的啮齿动物行为范式,用于理解多次试验食欲学习和记忆的细胞和分子机制。
依赖于主要嗅球 (OB) 的气味学习为多次试验食欲记忆的研究提供了几个优势。首先,OB依赖性记忆具有不同的持续时间(STM、LTM和中期记忆1),并且依赖于与大脑其他部位相同的分子2,3和结构机制,包括神经调控4、长期增强5和成体神经发生6,7,8.其次,与海马体等高阶区域相比,OB 依赖性记忆允许观察到实验者对感知环境的作与负责学习的神经回路的变化之间更直接的对应关系 8,9,10,11 .在本文中,详细介绍了一种可用于研究一般分子和结构机制的 OB 依赖性联想学习和记忆范式。它的开发是为了让研究人员能够利用嗅觉学习的优势来研究记忆的细胞和分子机制。
在我们最近的出版物3 中,此处描述的协议用于证明食欲气味学习的巩固取决于 OB 内的 Trk 受体激活。在下面的协议中,还讨论了可以根据不同实验需求调整行为范式的领域。
本研究共使用了 27 只成年雄性 CD-1 小鼠,插管时为 8 周龄。有关精确的组分布和气味集的使用,请参阅我们之前出版物3 的方法部分。雄性小鼠被用来避免雌激素水平的大幅波动,因为之前的研究12 表明雌激素水平的增加会增强嗅觉记忆。这些小鼠始终保持 12:12 小时的反向光照/黑暗循环,并且可以获得水。在行为实验期间,限制小鼠的饮食以将其维持在自由摄食体重的 ~90%。饮食限制在行为实验开始前 3 天开始。如下所述,同一组小鼠具有不同的气味集,以达到适当的统计功效水平,同时最大限度地减少动物使用。统计分析部分显示了如何解释由此可能引入的随机方差。
研究方案
以下协议遵守厄勒姆学院 IACUC 的动物护理指南。
1. 嗅球插管
注意:这些手术不需要无菌技术,因为它们不需要做大切口。但是,每个机构的要求可能有所不同。如果实验者对免疫功能低下的小鼠品系进行这种手术,则可能需要额外的考虑。总而言之,鼓励实验者在使用前与他们的兽医和动物护理团队讨论此方案,并在每次手术之间清洁和消毒所有工具。
- 在设置过程中,将套管和螺钉浸泡在含有 32% 洗必泰的小烧杯或培养皿中以保持消毒。
- 在纯氧中用气态 4% 异氟醚麻醉小鼠,并固定在立体定位装置中。在手术过程中,确保小鼠维持在通过鼻锥供应的 1.5-2% 异氟醚麻醉下。在整个手术过程中监测呼吸。使用眼药膏以防止麻醉时眼睛干燥。
- 在鼠标被固定并且不再对后足的用力捏有反应后,使用 32% 洗必泰摩擦头顶以清洁切口表面。
- 接下来,用利多卡因(局部镇痛剂)沿喙部向尾部摩擦头顶。
- 用力向下按压,用干净的手术刀刀片在中线上做一个切口。
- 使用连接到立体定位装置的钻头,使用坐标 AP +5.0 mm,ML +/-0.75 mm(相对于前囟)在嗅球上钻两个孔作为导向套管 (26 G)。
- 在小脑结构上钻两个孔。
注意: 这些螺钉的位置不需要精确,确保它们沿中线对称。 - 将螺钉放入小脑结构上方的两个孔中,并使用组织粘合剂将这些螺钉固定到颅骨上。
- 使用立体定位将导向套管 (26 G) 插入嗅球上钻出的孔中(步骤 1.6)。将套管 DV 降低 1.0 毫米。
- 在培养皿中混合牙科水泥。用一个小金属勺慢慢地将牙科粘固剂堆在套管周围。让它干燥 5 秒。然后,取下立体定位装置的臂,注意在此过程中不要将导管拉出。继续将牙科粘固剂堆在整个切口上,直到形成一个小盖子(见 图 1A)。
- 此时将假塞放入导向套管中,以防止堵塞。
- 手术后立即注射酮洛芬 (0.2 mg/kg) 和生理盐水 (200 μL) 以减轻疼痛和补水。手术后为小鼠提供软化的食物或水凝胶。在小鼠恢复意识以维持胸骨卧位之前,不要让它们无人看管。
注意:从这一点开始,老鼠也是单独饲养的。 - 手术后两天每天注射生理盐水 (200 μL) 和酮洛芬 (0.2 mg/kg) 一次。
- 手术后 2 天至最多 5 天(根据需要),称量小鼠并监测其体重。如果体重在两三天内没有恢复到手术前的水平,请咨询兽医关于适当的喂养方法。
- 在开始行为训练之前,让小鼠恢复至少 7 天。
2. 联想判别任务
- 注入
- 如下对小鼠进行酪氨酸激酶受体抑制剂 K252a(50 μM;5% DMSO 盐水溶液)或载体(5% DMSO 盐水溶液)的 OB 特异性输注。
- 将输液双侧输送到小鼠的 OB 中。使用双进样泵,以 0.2 μL/min 的输注速率和 10 分钟的总输注时间,每个灯泡注入 2.0 μL 最终体积。
注意:作的时间可以根据正在进行的行为研究的类型进行调整。确切的输注步骤特定于每个注射泵,并在制造商手册中提供。 - 确保在分娩后将注射器留在套管内约 5 分钟,以抑制回流并促进扩散。(即,计划每次输注总共 15 分钟)。
- 将输液双侧输送到小鼠的 OB 中。使用双进样泵,以 0.2 μL/min 的输注速率和 10 分钟的总输注时间,每个灯泡注入 2.0 μL 最终体积。
- 如下对小鼠进行酪氨酸激酶受体抑制剂 K252a(50 μM;5% DMSO 盐水溶液)或载体(5% DMSO 盐水溶液)的 OB 特异性输注。
- 气味集。
- 使用基于蒸气压的预先计算的比率,将所有加臭剂在轻质矿物油中稀释至 1.0 Pa 的分压(表 1)。
- 使用 表 1 中的 5 个单独的气味对(表中的数字表示在 50 mL 矿物油中混合 1.0 Pa 的体积(以 μL 为单位)。
- 为了准备在行为步骤(第 2.3 节和第 3 节)中使用的香味沙,每 100 克游戏沙混合 400 μL 步骤 2.2.2 中的 1.0 Pa 加臭剂。
- 塑造
注意:小鼠应进行 10 天的塑形,如下所述。- 将小鼠带入手术室,并在手术恢复后的前两天每天处理它们 10 分钟。
- 第 3 天,将一个装满 (+) - 柠檬烯香味沙的培养皿放入小鼠的家笼中,并装满约 10 个蔗糖沉淀,每个颗粒的质量为 5 毫克。
注:使用 1.0 Pa (+)-柠檬烯(在 50 mL 矿物油中混合 102 μL)作为奖励气味,使用普通矿物油(测试气味的稀释剂)作为无奖励气味。还建议选择单分子气味剂,因为这些气味剂很可能对小鼠来说是新的。 - 在第 4 天补充沙子和颗粒。
- 在第 5 天和第 6 天,通过将小鼠放入装置中并让它们探索空间,使小鼠适应定制的行为装置(图 1B、C)。使用标准家用笼子制作设备,并使用聚(甲基丙烯酸甲酯)构建两个盖子和一个黑色中心隔板。确保盖子和中央隔板都比家用笼子大 1-2 厘米。
- 准备一个培养皿,里面有柠檬烯香味的沙子,另一个培养皿里有含有矿物油的沙子。在第 5 天和第 6 天驯化后,将两个沙盘放入测试室中,并将 10 个蔗糖颗粒混合到柠檬烯香味的盘中作为奖励。将每只鼠标放入测试室 10 分钟,并允许自由探索和消耗奖励颗粒。
- 在第 7 天,通过将含有柠檬烯香味和矿物油香味沙子的培养皿放入行为装置中,将小鼠引入最终测试程序的简化版本。这次包括中心分隔线。
- 在柠檬香味的沙子上放一个奖励,然后将鼠标放入休息室。
- 将鼠标放入静置室后,提起中央隔板,让鼠标进入测试室,调查和挖掘装满沙子的培养皿。在鼠标取回奖励颗粒后或经过 5 分钟后,将鼠标放回休息室。
- 重复此过程,每只小鼠总共进行 10 次试验。通过使用随机数生成器来平衡奖励菜品在左侧或右侧的放置。试验之间没有休息时间。
- 在第 8 天,重复第 7 天的试验,但也逐渐将颗粒埋在沙子中越来越深。
注意:大多数小鼠应该在第 8 天的第 10次 试验之前挖掘看不见的奖励颗粒。 - 在第 9 天,将每只小鼠的试验次数增加到 20 次试验,将蔗糖沉淀完全深埋,并将小鼠引入测试室,每次试验 1 分钟。让老鼠在两个盘子里挖来获得奖励。
- 在第 10 天,对每只小鼠重复 20 次试验,但如果它们在挖有奖励的培养皿之前先挖无奖励的培养皿,然后开始下一次试验。让首先在奖励(柠檬烯香味)培养皿中挖洞的老鼠取回奖励颗粒,然后再将它们送回休息室。
3. 训练和测试
注意:一旦小鼠开始可靠地挖掘看不见的、有气味提示的奖励颗粒,就可以开始实验了。
- 训练
注意:训练阶段在整形完成后两天开始,包括每只小鼠的 20 次试验。在培训之前, 立即进行球内药物/载体输注(输注详情见第 2.1 节),并在输注后立即开始培训。- 将鼠标放入静置室。
- 将两个带有新气味对香味的沙子盘放入测试室中,其中一个盘子中埋有奖励颗粒。
- 测试室准备就绪后,掀开不透明屏障并将鼠标引入测试室。如果鼠标先在未奖励的盘子中挖洞,请立即将鼠标放回休息室(将这些试验记录为“0”)。如果老鼠先挖掘出有益的气味,请让它取回颗粒并将其送回休息室。将这些试验记录为 “1”。如果试用持续 1 分钟而鼠标没有取回奖励,请将鼠标送回休息室。
- 清洗并重新装满餐具,然后开始下一次试用。对 20 次试验重复相同的作。
- 测试
注意:记忆测试可以在研究人员感兴趣的任何持续时间进行。在该实验中,鉴于对 K252a 对 STM 和 LTM 的不同影响感兴趣,在训练后 2 小时 (STM) 或 48 小时 (LTM) 对两组独立的小鼠进行了测试。- 使用与培训中描述的相同的气味和程序进行测试(第 3.1 节)。
注意:根据研究问题,研究人员可能需要包括对照组。例如,在之前发表的实验中,研究了 Trk 受体阻断对记忆巩固的影响3。因此,对照组在 48 小时测试前输注 K252a,以表明效果不是由于干扰检索造成的。
- 使用与培训中描述的相同的气味和程序进行测试(第 3.1 节)。
- 统计分析
注意:每个步骤的 SPSS 22.0 语法作为 补充文件 作为示例提供。- 使用线性混合效应分析执行统计分析。与方差分析不同,线性混合效应模型可以更好地解释随机效应和重复测量。
- 计算相关度量:“ratio correct”。回想一下 3.1.3 中,鼠标首先挖掘有益气味的试验为“1”,如果鼠标首先挖掘无益的气味,则为“0”。平均每 5 次试验创建 4 个试验组(TB;例如,试验块 1 或 TB1 是试验 1-5 的平均值,试验块 2 或 TB 2 是试验 6-10 的平均值,依此类推)。
- 将自变量或固定效应设置为药物组(K252a 或载体;Section 2.1.1) 和 Trial Block(从 3.3.1 开始)。在下面的代表性结果中,指定了用于每个分析的变量。
- 将嵌套在鼠标中的单个鼠标和气味组作为“随机效应”包括在内,以补偿小鼠的内在行为差异以及使用多个气味组的任何影响。
- 对 Ratio correct 执行 logit 变换。
注意:比例正确不是连续的、未绑定的因变量。因此,它违反了线性模型的两个假设。因此,将执行 logit 变换。 - 使用估计的边际均值对完整模型识别的重要交互作用进行事后检验;必须在事后测试中更正多个成对比较。通常使用 Bonferroni 或 Šidák。
结果
如前所述,该协议允许研究人员评估某些作对学习、STM 和 LTM 的影响。此处介绍了 Tong et al, 20183 的样本结果。结果支持 Trk 受体阻断选择性抑制 LTM 而不是学习或 STM 的假设。
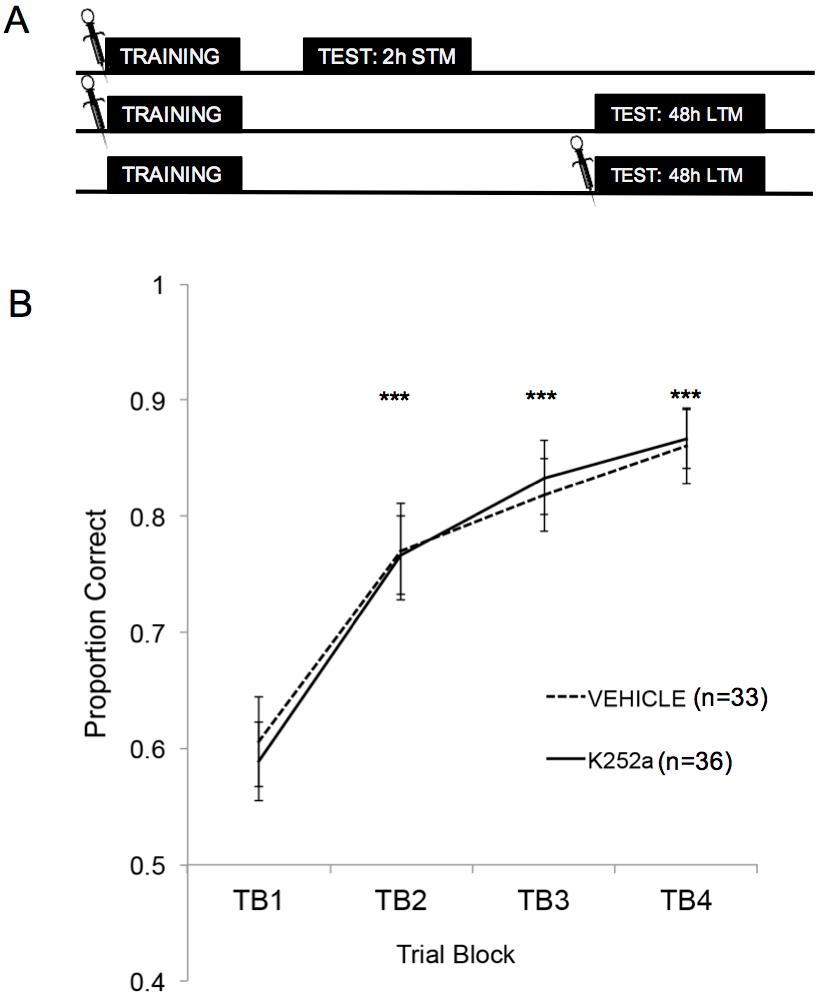
图 2A 显示了训练、 STM 测试和 LTM 测试的示意图。首先,结果表明 K252a 输注不会影响气味-奖励关联的学习率。 图 2B (第 3.1 节)显示了 K252a 和车辆组从 Training 中学习的速率。使用线性混合模型进行统计分析,包括两个固定效应,药物组和试验区间 (TB);嵌套在鼠标内的 Mouse 和 Odor Set 是随机效果。来自测试的数据(第 3.2 节)不包括在分析中。试验阻滞观察到显著的主效应 (F(3, 183.692) = 43.735, p < 0.001),但药物组没有影响 (F(1, 85.685) = 0.132, p = 0.717) 并且没有显著的交互作用 (F(3, 183.692) = 0.111, p = 0.954)。使用 Šidák 调整的事后测试证实,在训练期间,K252a 和载体组在任何试验块上都没有差异(所有比较的 p > 0.05)。在所有比较中,TB2、TB3 和 TB4 均显著高于 TB1 (在所有情况下 p ≤ 0.001),表明两组在 20 次训练试验结束时都成功学习了气味-奖励关联。
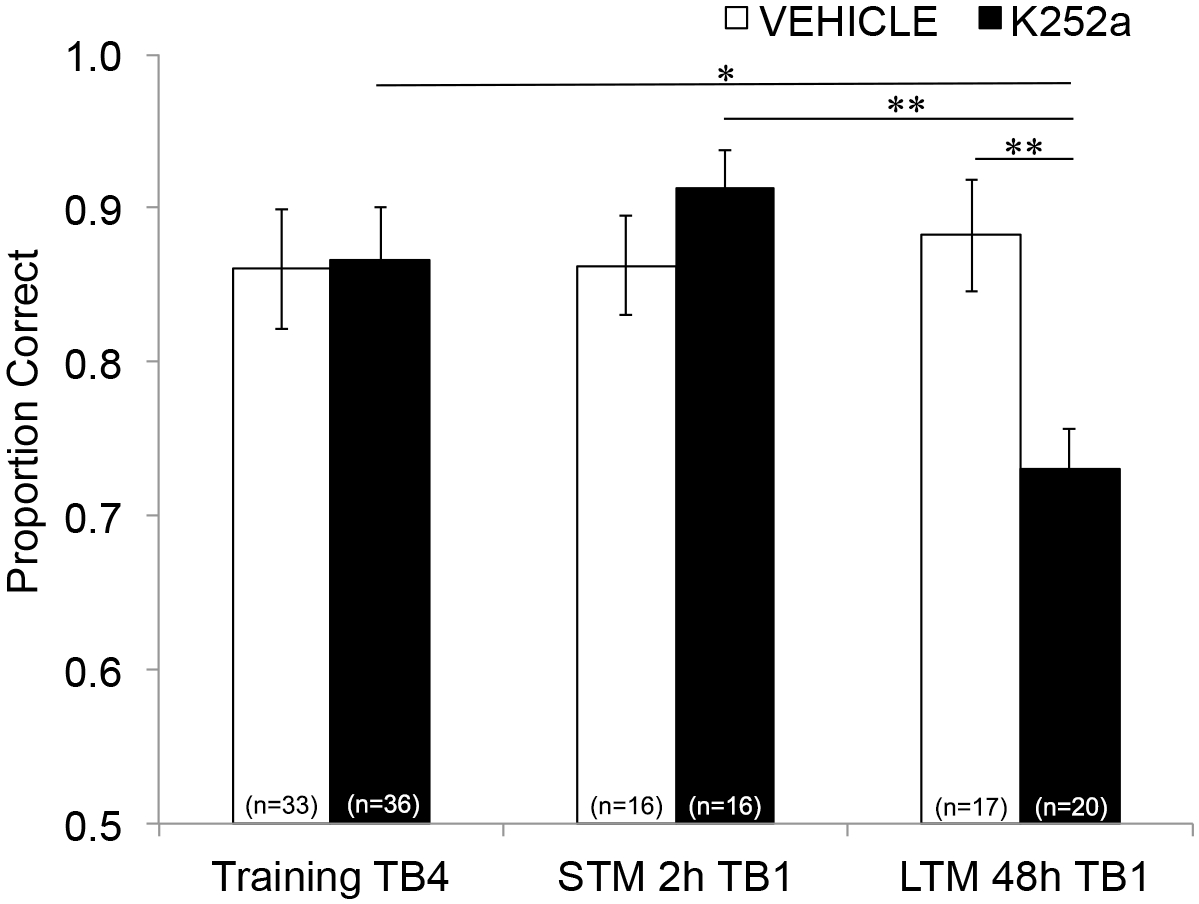
接下来,为了检查输注对 STM 和 LTM 的影响,进行了相同的分析并包括来自测试的数据(第 3.2 节)。结果显示药物组与试验块之间存在显著交互作用 (F(2, 77.558) = 4.043, p = 0.021),药物组 (F(1, 55.629) = 1.438, p = 0.236) 或试验组 (F(2, 69.979) = 1.360, p = 0.263) 无显著主效应。为了专门检查记忆,事后与 Šidák 校正进行成对比较,比较了训练的最后一个试验块(第 3.1 节)和测试的第一个试验块(第 3.2 节)之间的差异,在 2 小时 (STM) 或 48 小时 (LTM) 之后。对于注入载体的小鼠,比较显示训练后 2 小时和 48 小时联想记忆的保留(对于与训练表现的所有比较,p > 0.05)。对于注入 K252a 的小鼠,2 小时测试 (STM) 的第一个试验块与训练的最后一个试验块 (p > 0.05) 没有差异;然而,他们的记忆性能在 48 小时后显着降低 (p = 0.018)。此外,与 2 小时测试的记忆 (p = 0.009) 相比,48 小时测试的记忆显著降低,与 48 小时测试时车辆组的性能 (p = 0.006) 相比。载体和 K252a 输注小鼠之间的 STM 没有差异 (p = 0.356)。总之,结果表明 K252a 抑制嗅球中 Trk 受体会选择性地破坏长期而不是短期的气味记忆(图 3)。
| 气味套装 | 气味 1 | 气味 2 |
| 1 | 戊酸 | 丁酸 |
| 225.1 | 63.6 | |
| 2 | 己醛 | 庚醛 |
| 11.1 | 35.3 | |
| 3 | Propyl acetate 乙酸丙酯 | Butyl acetate 乙酸丁酯 |
| 3.1 | 10.9 | |
| 4 | 2-辛酮 | 2-庚酮 |
| 87.4 | 28.7 | |
| 5 | 戊醇 | 己醇 |
| 37.2 | 127.3 |
表 1:气味集的混合体积。 每行显示两种气味,它们可以用作行为步骤的一对。例如,要使用第一个“气味组”,请制作五酸和丁酸混合物。表中的数字表示在 50 mL 矿物油中混合每种气味浓度为 1.0 Pa 时的体积(以 μL 为单位)。在训练和测试期间,一个培养皿会用戊酸闻香,另一个培养皿会用丁酸闻香。

图 1:插管放置和行为装置。 (A) 显示套管、牙科粘固帽和螺钉与鼠标头部的相对位置。请注意,套管的针头伸入两个嗅球,基座本身嵌入牙科粘接帽中。螺钉被放置在小脑形成上方的颅骨中钻入的两个孔中。螺钉不接触大脑本身,但它们充当牙科粘接帽的尾部锚。该图显示了制作水泥帽的相对尺寸。(B) 显示组装的行为装置。身体是典型的老鼠家笼。带有空气孔的盖子由有机玻璃制成。中央隔板也由黑色有机玻璃制成。盖子应由足够重的有机玻璃制成,以便它们可以作为在试验期间抬起和放下中心隔板的通道。(C) 显示行为装置的一侧。请注意,中心隔板比笼子略高,以便于抬起。沙子培养皿可以沿边缘放置。 请单击此处查看此图的较大版本。
{kind=link}

图 2:研究设计和学习结果。 (A) 显示研究设计的示意图。请注意,STM 和 LTM 组是独立的(即不同的小鼠组)。块开头的符号表示输液的时间。(B) 显示训练期间试验 1-20 的正确比例。结果表明,K252a 和车辆组的学习率(线的斜率)没有差异。误差线表示均值的标准误差 (SEM)。星号显示载体和 K252a 队列的正确比例与 TB1 相比显着增加(所有比较的 p ≤ 0.001)。此图改编自 Tong 等人,2018 年,经许可3。 请单击此处查看此图的较大版本。
{kind=link}

图 3:Trk 受体阻断对 STM 和 LTM 的不同影响。 显示 STM 和 LTM 测试的训练试验块 4 和试验块 1 的正确比例。也就是说,使用线性混合模型来比较训练阶段最后一个试验块期间的正确比例(图 2;Training-TB4) 到短期(2 小时测试)和长期(48 小时测试)记忆测试的第一个试验块 (Testing-TB1) 期间的人。线性混合模型有两个固定效应:药物组和试验区间 (Training-TB4 、 STM-TB1 、 LTM-TB1 )。随机效应是嵌套在鼠标内的鼠标和气味集。事后比较发现 K252a 小鼠的 LTM 显著受损(与训练 TB4; p = 0.018),但不是 STM (p > 0.05)。K252a 输注小鼠的 LTM 性能也显著低于 K252a 输注小鼠的 STM (p = 0.009),低于载体小鼠的 LTM (p = 0.006)。误差线代表 SEM。此图改编自 Tong 等人,2018 年,经许可3。 请单击此处查看此图的较大版本。
{kind=link}

补充文件:用于统计分析的语法。 请点击此处下载此文件。
讨论
一次性恐惧条件反射是研究记忆的分子和细胞动力学的强大行为协议,但大部分自然学习是渐进的,最好通过上述范式进行建模。OB 中 Trk 受体的抑制阻止了嗅觉记忆在多次试验、食欲学习范式中的巩固,正如我们之前第3 组所示。这一发现为研究神经营养因子等分子机制在食欲学习和厌恶学习中的不同时间开辟了新的途径。
该实验包括两个关键部分:(1) 插管和 (2) 联想判别任务(细分为整形、训练和测试)。实验者可以根据他们的特定研究问题调整此方案。例如,我们主要对 OB 感兴趣,这个建立的方案可以很容易地应用于其他 OB 研究。对于其他感兴趣区域的实验者,在试点研究中验证输注部位非常重要。实验者可能还需要考虑他们所注入的物质的扩散速率、空间渗透率和生物活性持续时间。
该协议中概述的整形步骤已被该协议的作者广泛使用。如前所述遵守它们似乎对于让小鼠及时学习任务很重要。使用其他时间线,作者观察到小鼠对任务的熟悉程度存在更多差异,这意味着需要进行额外的训练,以使所有小鼠都达到测试实验气味的标准。对于培训和测试,研究人员可以根据他们的研究兴趣、试验次数、实验气味的浓度以及气味之间的相似性进行灵活调整。正如我们所描述的,我们建议尽可能使用多个气味组,以减少用于实验的动物数量。有关如何在最终分析中考虑多个阶次集使用的指导,请参阅 Statistical Analysis 部分。原则上,气味对的相似性可以改变以调整鉴别的难度。在我们第3 组之前发表的研究中,气味对由具有相同官能团但彼此相差一个碳长度的两种气味剂组成。这些区分比相差两个或多个碳长度的对更困难,但比对映异构体(例如,(+)-柠檬烯和 (-)-柠檬烯)更容易。来自不同官能团的加臭剂在感知上存在很大差异。Cleland 等人13 讨论了更多的刺激变化及其对特定学习参数的影响。
该协议的一个主要限制是,与可以并行测试多个动物的自动联想学习任务相比,它需要更长的时间来执行。对于给定的测试,一名研究人员至少需要 20 分钟才能完成一只小鼠的 20 次试验。然而,正是这种缺乏自动化意味着该协议在财务上更容易获得,这是许多机构的首要任务。重要的是,在这个实验的情况下,已经发现该协议对于培训对行为神经科学感兴趣的本科生研究人员非常易于处理和有效。特别是,这些学生除了获得参与研究的通常好处外,还培养了强大的动物处理技能。
有兴趣采用此范式的研究人员可以改变几个参数。最明显的是,机制的药理学作是多种多样的,这种行为方案可以与化学遗传学技术或纵分子和细胞途径的各种其他方式(例如 光遗传学)。范式本身可以进行调整,以改变所测试的学习和记忆类型。例如,研究人员可以调整呈现的两种气味的相似性,以控制学习率。在我们的研究3 中,气味对由两种具有相同官能团但彼此相差一个碳长度的气味剂组成。这些区分比相差两个或多个碳长度的对更困难,但比对映异构体(例如 (+)-柠檬烯和 (-)-柠檬烯)更容易。来自不同官能团的加臭剂在感知上存在很大差异。Cleland 等人13 讨论了更多的刺激变化及其对特定学习参数的影响,得出的结论是,更相似的气味更难区分,因此需要更长的时间来学习14。这些作也会影响记忆的强度。沿着这个思路,研究人员可能有兴趣在学习后的不同时间点测试记忆。例如,两项研究15,16 检查了 BDNF 在单次试验厌恶性学习任务的 LTM 持久性中的作用。BDNF 是 TrkB 的配体。研究表明,学习后 12 小时在海马体中输注抗 BDNF 反义寡核苷酸可在 7 天后阻断 LTM,但在 2 天后则不阻断。这项研究表明,初始学习后分子机制的时间尺度在 LTM 中起着有趣但尚未理解的作用。本文描述了一种允许调查这些时间尺度的行为协议。未来应用的其他相关参数包括使用的小鼠模型。例如,在未来的研究中,用雌性小鼠代替雄性小鼠(神经生物学研究得更好)17 以检查学习率、STM 和 LTM 的变化会很有趣,因为雌性哺乳动物对气味的敏感性和选择性高于雄性哺乳动物18。当然,疾病的啮齿动物模型也可以与该协议一起有效使用。
披露声明
本文的作者没有相互竞争的经济利益。
致谢
该项目得到了 Scantland Summer Collaborative Research Gift 和 Stephen and Sylvia Tregidga Burges Endowed Research Fund 的支持。作者要感谢厄勒姆学院心理学系和神经科学项目的同事们的支持和指导。
材料
| Name | Company | Catalog Number | Comments |
| Double guide cannula | PlasticsOne | C235GS-5-1.5/SPC | Custom order |
| (-)-limonene | Sigma-Aldrich | 218367-50G | |
| (+)-limonene | Sigma-Aldrich | 183164-100ML | |
| 2-hetanone | Sigma-Aldrich | 537683 | |
| 2-octanone | Sigma-Aldrich | O4709 | |
| 5mg sucrose pellets | Test Diet | 1811560 | Custom size. Used for rewards |
| Butanoic acid | Sigma-Aldrich | B103500 | |
| butyl acetate | Sigma-Aldrich | 402842 | |
| Dental Cement Powder (Coral) | A-M Systems | 525000 | |
| Dental Cement Solvent | A-M Systems | 526000 | |
| Double connector assembly | PlasticsOne | C232C | |
| Double dummy cannula | PlasticsOne | C235DCS-5/SPC dummy dbl | Custom order |
| Double injector | PlasticsOne | C235IS-5/SPC | Custom order |
| Drill | Kopf Instruments | Model 1474 High Speed Stereotaxic Drill | This drill requires an additional "adaptor" piece in order to fit certain drill bits. We get by this problem by wrapping the drill bit with lab tape to increase the circumference of the drill it to fit. This may not be an option for surgeries requiring sterile technique. |
| Eye Ointment | Purchase from local pharmacy | ||
| Figure 1 illustration software | BioRender | ||
| heptanal | Sigma-Aldrich | W254002 | |
| hexanal | Sigma-Aldrich | 115606 | |
| hexanol | Sigma-Aldrich | H13303 | |
| Infusion pump model 11 | Harvard Apparatus | 4169D | Used pumps available via American Instrument Exchange |
| Isoflurane | Santa Cruz Animal Health | sc-363629Rx | Vet prescription needed for order |
| K252a | Sigma-Aldrich | K2015 | Mixed to 50uM in DMSO (5%) |
| Ketoprofen | Allivet | 25920 | Vet prescription needed for order |
| Lidocaine | Aspercreme | Purchased from Amazon | |
| Mounting Screws | PlasticsOne | 00-96 X 3/32 | |
| Mouse Anesthesia Mask | Kopf Instruments | Model 907 Mouse Anesthesia Mask | Used with the stereotaxic to allow oxygen and anesthesia while mouse in stereotax |
| Mouse Nose Adaptor | Kopf Instruments | Model 926 Mouse Adaptor | Used with the stereotaxic to allow for head of mouse to be secured. |
| Novalsan | Jeffers | 41375 | |
| Pentanoic acid | Sigma-Aldrich | 240370 | |
| pentanol | Sigma-Aldrich | 138975 | |
| Petri dish glass bottoms | VWR | 10754-804 | |
| Polycarbonate Café bottoms | Ancare | N10PCSEC | Use normal housing cages and custom fit a track in the middle to act as the track for an opaque plexiglass divider |
| propyl acetate | Sigma-Aldrich | 537438 | |
| Quikrete Premium Play Sand | Purchase from local hardware store | ||
| Saline | Insight Needles | N/A | Sterile saline for drug mixing |
| Stereotaxic apparatus | Kopf Instruments | Model 902 Small Animal Stereotaxic Instrument | |
| Testing chamber | Ancare | N10PCSEC | Our testing chambers are modified using the regular mouse housing cage. The manuscript details what was done. |
| Vetbond Tissue Adhesive | 3M | Purchased from Amazon |
参考文献
- Grimes, M. T., Smith, M., Li, X., Darby-King, A., Harley, C. W., Mclean, J. H. Mammalian Intermediate-Term Memory: New Findings in Neonate Rat. Neurobiology of Learning and Memory. 95 (3), 385-391 (2011).
- Grimes, M. T., Harley, C. W., Darby-King, A., Mclean, J. H. PKA Increases in the Olfactory Bulb Act as Unconditioned Stimuli and Provide Evidence for Parallel Memory Systems: Pairing Odor with Increased PKA Creates Intermediate- and Long-Term, but not Short-Term, Memories. Learning and Memory. 19 (3), 107-115 (2012).
- Tong, M. T., Kim, T. Y. P., Cleland, T. A. Kinase Activity in the Olfactory Bulb is Required for Odor Memory Consolidation. Learning & Memory. 25 (5), 198-205 (2018).
- Devore, S., Lee, J., Linster, C. Odor Preferences Shape Discrimination Learning in Rats. Behavioral Neuroscience. 127 (4), 498-504 (2013).
- Gao, Y., Strowbridge, B. W. Long-Term Plasticity of Excitatory Inputs to Granule Cells in the Rat Olfactory Bulb. Nature Neuroscience. 12 (6), 731-733 (2009).
- Bath, K. G., Mandairon, N., et al. Variant Brain-Derived Neurotrophic Factor (Val66Met) Alters Adult Olfactory Bulb Neurogenesis and Spontaneous Olfactory Discrimination. Journal of Neuroscience. 28 (10), 2383-2393 (2008).
- Lazarini, F., Lledo, P. M. Is Adult Neurogenesis Essential for Olfaction. Trends in Neurosciences. 34 (1), 20-30 (2011).
- Mandairon, N., Peace, S., Karnow, A., Kim, J., Ennis, M., Linster, C. Noradrenergic Modulation in the Olfactory Bulb Influences Spontaneous and Reward-Motivated Discrimination, but not the Formation of Habituation Memory. European Journal of Neuroscience. 27 (5), 1210-1219 (2008).
- Guérin, D., Peace, S. T., Didier, A., Linster, C., Cleland, T. A. Noradrenergic Neuromodulation in the Olfactory Bulb Modulates Odor Habituation and Spontaneous Discrimination. Behavioral Neuroscience. 122 (4), 816-826 (2008).
- Moreno, M. M., Bath, K., Kuczewski, N., Sacquet, J., Didier, A., Mandairon, N. Action of the Noradrenergic System on Adult-Born Cells Is Required for Olfactory Learning in Mice. Journal of Neuroscience. 32 (11), 3748-3758 (2012).
- Vinera, J., Kermen, F., Sacquet, J., Didier, A., Mandairon, N., Richard, M. Olfactory Perceptual Learning Requires Action of Noradrenaline in the Olfactory Bulb: Comparison with Olfactory Associative Learning. Learning and Memory. 22 (3), 192-196 (2015).
- Dillon, T. S., Fox, L. C., Han, C., Linster, C. 17β-estradiol Enhances Memory Duration in the Main Olfactory Bulb in CD-1 Mice. Behavioral Neuroscience. 127 (6), 923 (2013).
- Cleland, T. A., Narla, V. A., Boudadi, K. Multiple Learning Parameters Differentially Regulate Olfactory Generalization. Behavioral Neuroscience. 123 (1), 26 (2009).
- Cleland, T. A., Morse, A., Yue, E. L., Linster, C. Behavioral Models of Odor Similarity. Behavioral Neuroscience. 116 (2), 222-231 (2002).
- Bekinschtein, P., Cammarota, M., Igaz, L., Bevilaqua, L., Izquierdo, I., Medina, J. Persistence of Long-Term Memory Storage Requires a Late Protein Synthesis- and BDNF- Dependent Phase in the Hippocampus. Neuron. 53 (2), 261-277 (2007).
- Bekinschtein, P., et al. BDNF is Essential to Promote Persistence of Long-Term Memory Storage. Proceedings of the National Academy of Sciences. 105 (7), 2711-2716 (2008).
- Andreano, J., Cahill, L. Sex influences on the neurobiology of learning and memory. Learning and Memory. 16 (2009), 248-266 (2009).
- Kass, M. D., Czarnecki, L. A., Moberly, A. H., Mcgann, J. P. Differences in Peripheral Sensory Input to the Olfactory Bulb Between Male and Female Mice. Scientific Reports. 7 (1), (2017).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。