
Method Article
通过定量染色体形成捕获(4C)鉴定胚胎体增强剂-促进者接触
摘要
我们报告定量染色体构象捕获的应用,随后在胚胎干细胞生成的胚胎体中进行高通量测序。该技术允许在胚胎干细胞分化期间识别和定量特定基因的假定增强剂和启动器区域之间的接触。
摘要
在哺乳动物发育过程中,通过建立调控网络来确定细胞命运,这些网络定义了基因表达的特异性、时间和空间模式。多能干细胞衍生的胚胎体(EB)是研究主要三个胚层分化并在细胞命运规范中定义调控回路的流行模型。虽然众所周知,组织特异性增强剂通过与启动者互动在这些网络中扮演着重要的角色,但将它们分配给相关的靶基因仍然具有挑战性。为了使这一成为可能,需要采取定量方法,研究增强剂促进者接触及其在开发过程中的动态。在这里,我们调整了4C方法,在EB分化模型中定义增强剂及其与同构促进器的接触。该方法使用频繁切割限制酶、声波和与商业DNA库制备试剂盒兼容的嵌套连接介导PCR协议。随后,4C 库进行高通量测序,并进行生物信息分析,从而能够检测和定量与所选启动器接触的所有序列。生成的测序数据还可用于获取有关增强剂-促进器在分化期间接触动力学的信息。EB 分化模型描述的技术易于实现。
引言
在小鼠中,3.5天胚胎的内细胞质量(ICM)含有胚胎多能干细胞。ICM在第4.5天进一步发展成表外爆炸,产生异母细胞、中皮细胞和内皮细胞,这是胚胎中主要的三个生殖层。虽然ICM中的多能细胞只是暂时的存在于体内,但它们可以通过建立小鼠胚胎干细胞(mESCs)1、2、3,2,3在培养中捕获。mESCs仍然处于无差别状态,无限扩散,但在内在和外在刺激时,它们也能够退出多能状态,产生三个发育性胚芽层22、44的细胞。有趣的是,当在小液滴悬浮中培养时,mESC形成三维聚合体(即EB),分化成所有三个胚芽层5。EB地层测定是研究早期系系规范过程的重要工具。
在谱系规范中,每个生殖层的细胞获得特定的基因表达程序4。基因的精确时空表达受多种调节元素的调节,包括核心促进剂、增强剂、消音器和绝缘体66、7、8、9。7,8,9增强剂,调节DNA段通常跨越几百个碱基对,协调组织特异性基因表达8。增强剂通过约束转录因子和调节局部染色质结构88,1010的配合机激活或沉默。识别假定增强剂的常用技术是全基因组染色质免疫沉淀,然后是测序(ChIP-seq)和利用测序(ATAC-seq)技术进行转波酶可获取的染色质的测定。因此,主动增强剂的特点是特定的活性组蛋白标记和增加局部DNA可及性11,12,13,14。11,12,13,14此外,发育增强剂被认为需要与他们的认知促进者88,99物理互动。事实上,已经表明,增强剂变种和删除,扰乱增强剂促进者接触可能导致发育畸形15。因此,需要采用新技术,为识别控制发育基因表达的功能增强剂提供额外信息。
自染色体构象捕获(3C)技术16开发以来,染色体接触的映射被大量用于评估调节元素之间的物理距离。重要的是,最近开发了3C技术的高通量变体,为染色质片段17之间的固定、消化、结扎和恢复提供了不同的策略。其中,原位高C已成为一种流行的技术,允许测序3C连接产物全基因组18。然而,达到适合分析增强剂-促进器触点的分辨率所需的高测序成本使得该技术难以用于研究特定位点。因此,,开发了替代方法,以分析第19、20、21、22,20,21号决议的靶点。22这些方法之一,即 4C,称为一个与所有策略,允许检测所有与选定为视点的站点的序列。然而,标准 4C 技术的缺点是所需的反向 PCR,它放大了大小不同的片段,有利于小产品,并在高通量测序后偏置定量。最近,UMI-4C,一种使用唯一分子标识符(UMI)的4C技术的新变种,已经开发用于定量和有针对性的染色体接触分析,绕过这个问题23。这种方法使用频繁的刀具、声波和嵌套连接介导的PCR协议,从而涉及长度分布相对均匀的DNA片段的扩增。这种同质性减少了PCR偏好放大过程中的偏差,从而缩短了序列,并可实现空间连接分子/片段的有效恢复和精确计数。
在这里,我们描述了一种协议,该协议使UMI-4C技术适应EB分化期间系系指导性转录因子的促进因素和增强剂之间的染色质接触。
研究方案
1. 小鼠胚胎干细胞胚胎体生成
- 制备mESC无血清培养培养基:DMEM/F12和神经基础介质以1:1的比例混合。培养基辅以MEM非必需氨基酸溶液(1倍)、丙酮钠(1 mM)、L-谷氨酰胺(2 mM)、青霉素-链霉素(100 U/mL)、β-梅尔卡托 乙醇(50 μM)、N2和B27补充剂(1倍)、PD0325901(1 μM)、CHIR99021(3 μM)和白血病抑制因子(LIF)(1,000 U/mL)。
- 制备EB分化介质:DMEM补充10%胎儿牛血清(FBS)、MEM非必需氨基酸(1倍)、丙酮钠(1 mM)、L-谷氨酰胺(2 mM)、青霉素-链霉素(100 U/mL)、β-甲醇(50μM)。
- 在 10 厘米塑料菜肴上培养 mESC,在 mESC 培养无血清介质中预涂 0.1% (w/v) 明胶。
- 当 mESC 达到 60% 的汇合度时,去除培养介质,用 2 mL 消毒 PBS 轻轻洗涤 1x。
- 完全取出PBS,并加入2 mL的细胞分离介质。在37°C孵育培养皿5分钟。
- 通过在盘中加入8 mL的EB分化介质来灭活反应。
- 通过上下移液15-20次分离mESC菌落,以获得单细胞悬浮液。
- 在室温 (RT) 下以 300 x g 的 300 x g离心 5 分钟,并小心地取出上清液。
- 计算细胞(例如,使用血细胞计)。
- 用EB分化介质重新悬浮细胞颗粒,并将浓度调整到2 x 104细胞/mL。
- 反转 15 厘米培养盘的盖子,并使用 200 μL 多通道移液器在盖子上沉积 20 μL 的重新悬浮细胞(+400 细胞/滴)。
- 小心地将盖子倒置到底部腔室上,在37°C下用悬挂液滴孵育,5%CO2和95%湿度为3天。
- 用 10 mL PBS 轻轻清洗盖子,将含 EB 的悬浮液转移到 50 mL 塑料管中,收集 EB。
- 将管置于 RT 上 30 分钟,以便 EB 通过重力沉入底部。小心地取出上清液。
- 用10 mL的新鲜EB分化介质轻轻重新悬浮EB,并转移到10厘米细菌培养皿中。
- 3-6 天后使用倒置显微镜检查 EB 形成。生成的 EB 应是圆形和均匀的尺寸。
- 在37°C下孵育培养物,同时进行5%的CO2和95%的湿度。EB将继续分化成三个胚芽层,并可在不同的时间点收集进行分析。
2. EB 的分离
- 将 2 到 3 个 10 厘米的 PB 收集到 50 mL 塑料管中。在 RT 时将 EB 在 300 x g下离心 5 分钟,然后小心地取出上清液。
- 使用 10 mL PBS 重新挂起 EB。在 RT 时以 300 x g的离心 EB 3 分钟,并去除上清液。
- 添加 2 mL 胰蛋白酶-EDTA (0.25%)到颗粒,在37°C孵育管15分钟。 移液器上下每3分钟获得单细胞悬浮液。
- 加入8 mL的EB分化介质,停止胰蛋白酶反应。在显微镜下检查EB分离并计数细胞。
3. 固定
- 以 1 x 106细胞/mL 的速度重新悬浮新鲜 EB 培养基中的细胞。对于 50 mL 管,在 45 mL 介质中最多使用 4.5 x 107个单元。
- 将甲醛从37%的库存(不超过6个月)添加到1%的最终浓度。
注意:在处理甲醛时,遵循适当的健康和安全规定,因为它是一种危险化学品。 - 在旋转下 RT 孵育 10 分钟。
- 通过将甘氨酸加入到0.125 M的最终浓度中来淬火甲醛。
- 在旋转下的 RT 孵育 5 分钟。
- 从现在起,将固定细胞转移到冰上,并在4°C下保持寒冷。
- 在冷藏离心机中以 300 x g的速度将细胞切碎 5 分钟。
- 丢弃上清液,在冷PBS中重新悬浮颗粒(1 mL,用于 5 x 106个细胞),然后转移到 1.5 mL 安全锁管中。
- 在4°C下以300 x g将细胞颗粒5分钟,丢弃上清液,并在液氮中捕捉冷冻颗粒。储存在-80°C或继续下面的协议。
4. 细胞裂酶和限制性酶消化
- 将细胞颗粒轻轻重新悬浮在新鲜制备的冰冷赖解缓冲液(10 mM Tris-HCl pH = 8.0、10 mM NaCl、0.2% Igepal CA630 和 1x 蛋白酶抑制剂)中,每 2-5 x 106个细胞。要准备 5 mL 的 lysis 缓冲区,请参阅表 1。
- 在冰上孵育细胞15分钟。
- 在 1,000 x g下离心 5 分钟,在 4 °C 下。丢弃上清液,并保留含有核的颗粒。
- 用500μL的冷水清缓冲液清洗颗粒核。
- 在 1x 缓冲 2 中以 50 μL 0.5% SDS 的 50 μL SDS 轻轻重新悬浮颗粒,然后在 62 °C 的加热块中孵育 10 分钟。
- 从加热块中取出管,加入170μL的消化缓冲液,含有25μL的10%三吨X-100,以淬火SDS。通过移液混合良好,避免过度发泡。
- 在37°C孵育15分钟。
- 加入25μL的消化缓冲液,通过反转混合,并采取8μL作为一个未消化的控制。将未消化的控制样品保持在 -20°C。将 100 U MboI 限制酶(25 U/μL 库存的 4 μL)添加到剩余核中,并在旋转 37°C 下消化染色质 2 小时。再添加另一个 100 U 的 MboI,再孵育 2 小时。
- 再加入100 U的MboI,并在37°C下一夜旋转孵育。
- 第二天,再加入100 U的MboI,在旋转37°C下孵育3小时。
- 以 8 μL 作为消化控制样品。通过添加80 μL的TE缓冲液(10 mM Tris pH = 8,1 mM EDTA)和10μL蛋白酶K(10mg/mL),从步骤4.8中分离消化消化控制样品和未消化的控制样品。在65°C孵育1小时。
- 在0.6%凝胶上运行20μL等位物,以检查消化效率。成功的消化主要显示3.0-0.5 kb范围内的碎片。
5. 接近连接和交联反转
- 在加热块中孵育65°C的MboI消化样品20分钟,使MboI失去活性,然后冷却至RT。
- 在 RT 时以 1,000 x g将管离心 5 分钟,去除上清液,并将颗粒溶解在 200 μL 的新鲜联体缓冲液中。
- 在每个样品中加入 1,000 μL 的结扎母混合物。要准备连接母体组合的 1,000 μL,请参阅表 2。
- 通过反转和孵育在RT一夜与缓慢旋转(9 rpm)。
- 加入100μL蛋白酶K(10mg/mL)和10μL的RNase A(10mg/mL),消除RNA和蛋白质残留物。在55°C孵育样品45-60分钟。
- 在 65 °C 下继续孵育样品,再孵育 4 小时。
6. DNA剪切和尺寸选择
- 冷却管到RT。
- 在 4 °C 下以 1,000 x g的离心机 5 分钟。
- 将样品分成2 mL管中的三个400μL等位,在每个管中加入2μL的糖原(20mg/m)、40μL的醋酸钠(3 M,pH = 5.2),以及2.5倍体积(1 mL)100%乙醇。在-80°C下进行反转和孵育45-60分钟混合。
- 在4°C下在16,000 x g下离心25分钟。旋转后将管子放在冰上,并通过移液小心地去除上清液。
- 通过在70%乙醇的800μL中重新悬浮来洗涤DNA颗粒。在 4 °C 下在 16,000 x g下离心 5 分钟。
- 去除上清液,用800μL的70%乙醇再次进行洗涤。
- 将颗粒溶解在 1x Tris 缓冲液(10 mM Tris-HCl,pH = 8)的 130 μL 中,并在 37 °C 孵育 15 分钟以完全溶解 DNA。如有必要,使用移液重新悬浮任何沉淀物。
- 测量DNA产量;2.5-5 μg 染色质可望为 1 x 106细胞。通过在 0.6% 的琼脂胶凝胶上运行 3C 产品的 ± 200 ng 来检查结扎。成功的结扎主要显示DNA片段 > 3 kb。将样品存放在 -20 °C。
- 在 0.65 mL 管中稀释样品,该管适用于在 100 μL 的 1x Tris 缓冲体积(每管 1 μg)中声波到 10 纳克/μL。用于库制备的标准量为3μg,因此如有必要,在三个独立的管中执行声波。
- 使用声波器上的以下参数将DNA剪切到150-700 bp(平均=400-500 bp)的大小:循环:6-8,20 s-60 s关闭。这将使DNA适合使用光明场测序仪的高通量测序库制备。
- 将拉线DNA转移到一个正常的新的安全锁管。从同一样本池多个声波。
- 在RT加热一瓶DNA纯化珠。从现在开始,使用低绑定提示。
- 将 1.8 倍的珠子添加到 DNA 管中,轻轻重新悬浮。
- 在RT孵育5分钟。
- 用磁性架收集珠子。用 1 mL 新鲜制备的 80% 乙醇清洗珠子 2x,同时将管保持在磁性机架中。
注:清除所有乙醇,包括残留液滴。 - 在 RT 时将珠子短暂(2-3 分钟)空气干燥。
注:请勿干燥珠子超过 5 分钟。这将降低DNA产量。 - 用 90 μL 的 1x Tris 缓冲液(10 mM Tris-HCl,pH = 8)重新悬浮珠子,以释放 DNA。
- 测量DNA产量,并在1.5%凝胶上分析5μL等位。与预音速率相比,损失应该很小。
7. 图书馆为测序做准备
- 从库制备套件中加入 15 μL 的主混合。要修复胶合DNA的末端,将10μL的10倍端修复反应缓冲液和5μL的端修复酶混合物相结合。
- 在RT孵育30分钟。
- 加入1.1倍的DNA纯化珠,轻轻重新悬浮。
- 在RT孵育5分钟。
- 用磁性架收集珠子。用1 mL新鲜制备的80%乙醇清洗珠子两次,同时将管子保持在磁性架中。去除乙醇。
- 在RT时将珠子干2-3分钟。用42μL的1x三分缓冲液(10 mM Tris-HCl,pH = 8)重新悬浮珠子,以释放DNA。
- 在每个样品中加入8μL的dA尾接主混合物。要准备 dA 尾控主混合,请组合 5 μL 的 10x dA 尾矿反应缓冲液和 3 μL 的 Klenow 片段外减。
- 在37°C孵育30分钟。
- 加入2μL小牛肠道碱性磷酸酶(CIP)以脱磷脱脂DNA。
- 在37°C孵育30分钟,然后在50°C孵育60分钟。
- 加入1.1倍的DNA纯化珠,轻轻重新悬浮。
- 在RT孵育5分钟。
- 用磁性架收集珠子。用 1 mL 新鲜制备的 80% 乙醇清洗珠子 2x,同时将管保持在磁性机架中。
- RT. 重新悬浮珠时短暂(2-3 分钟),具有 35 μL 的 1x Tris 缓冲液(10 mM Tris-HCl,pH = 8),以释放 DNA。
- 执行适配器连接反应。使用表 3中提到的降低适配器/ligase 浓度。
- 在20°C孵育15分钟。
- 加入3 μL的尿素DNA糖酶和DNA糖酶酶酶酶VII(例如,USER)酶的混合物,通过移液混合,并在37°C孵育15分钟。
- 用水将体积增加到100μL,在96°C下煮5分钟,然后将样品放在冰上。
- 加入1.1倍的DNA纯化珠,轻轻重新悬浮。
- 在RT孵育5分钟。
- 用磁性架收集珠子。用 1 mL 新鲜制备的 80% 乙醇清洗珠子 2x,同时将管保持在磁性机架中。
- 在RT时将珠子干2-3分钟。用50μL的1x三分缓冲液(10 mM Tris-HCl,pH = 8)重新悬浮珠子,以释放DNA。
8. 4C 染色质相互作用库扩增和纯化
- 使用 10 μL 库放大 4C 库以执行第一个 PCR。PCR 设置和程序可在表 4中找到。
- 执行嵌套 PCR。嵌套 PCR 设置和程序可在表 5中找到。
- 为每个库池 PCR 产品,并用 1.1 倍 DNA 纯化珠进行纯化。
- 测量DNA产量,并分析1.5%凝胶上的5μL等位。
- 调整库浓度并对库进行排序。如果编制索引,则可以在排序之前池化库。
结果
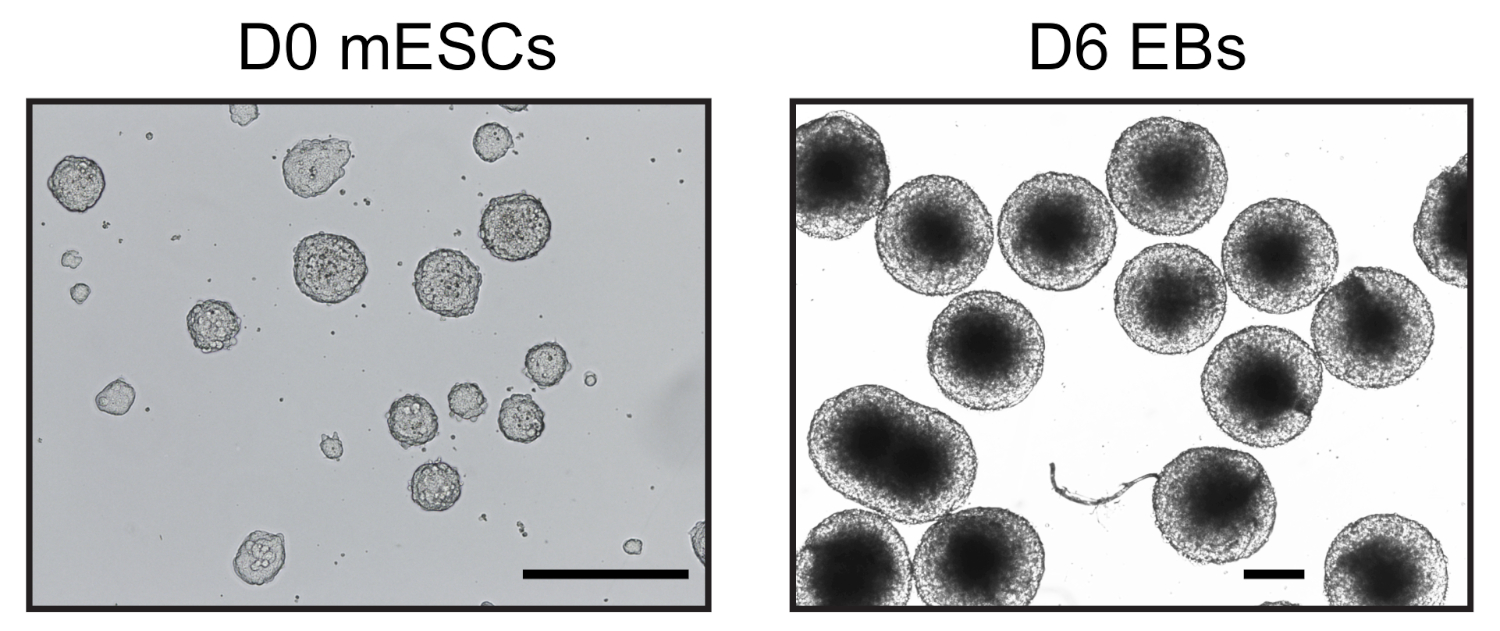
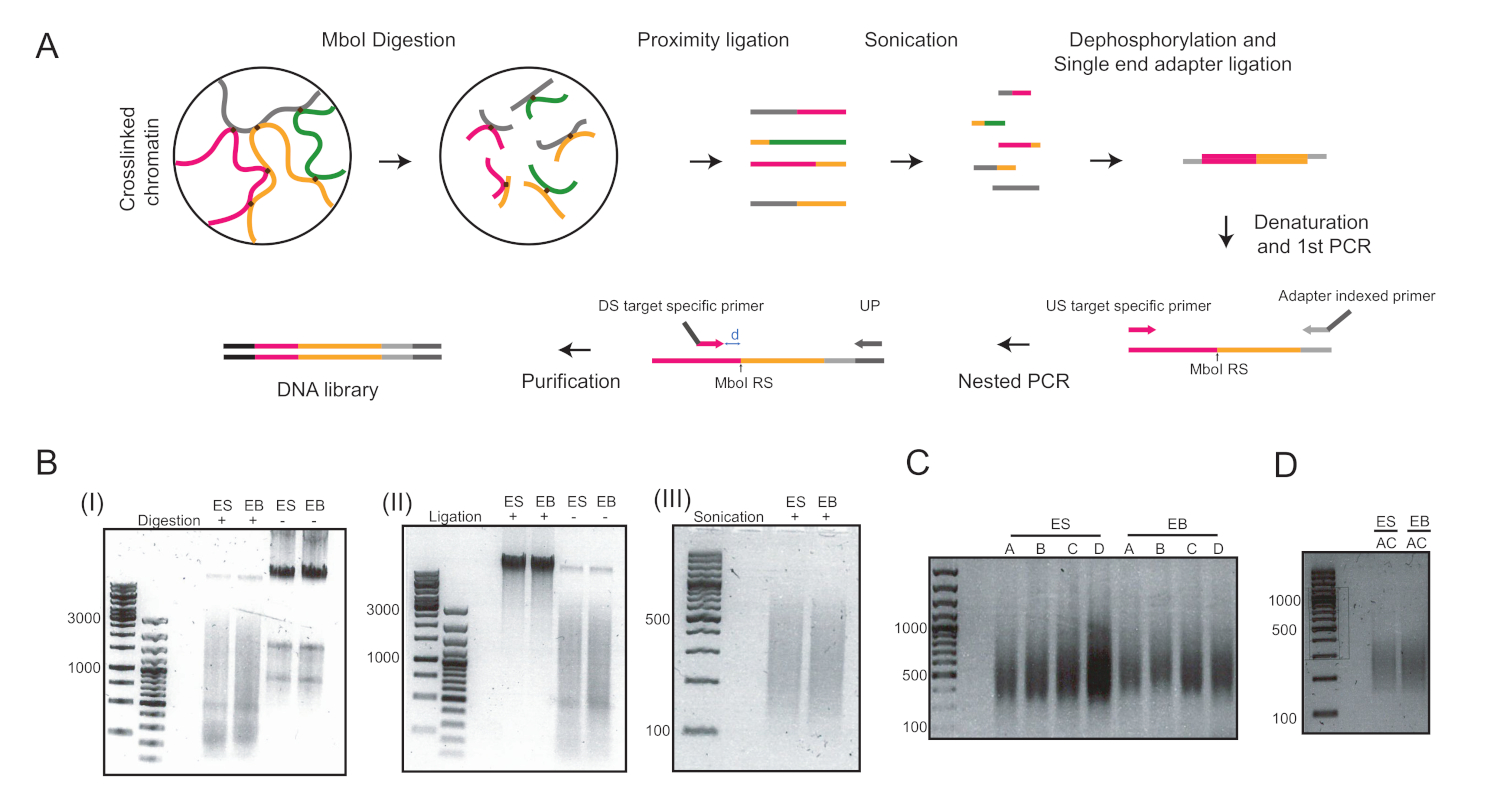
在悬挂液滴中引入ESC分化六天后,我们获得了用于进一步分析的EBs的同质种群(图1)。我们调整了UMI-4C方法23,以量化EBs24中系谱特定基因的促进者的特定染色质相互作用。图2A显示了不同步骤中具有代表性的质量控制凝胶的协议的原理图概述。进行了第一次质量控制,以确定MboI限制酶消化的效率。有效消化显示片段大小小于3 kbp(图2B)。值得注意的是,mESC和EB染色质消化是困难的,有时残留未消化的染色质持续存在。第二次质量控制是在结扎后进行的,以验证大多数碎片现在为 > 3 kbp(图 2B)。然后,通过凝胶电泳分析声波后获得的染色质片段。碎片尺寸预计为400-500bp(图2B)。
脱磷和单端适配器连接后,进行了两轮PCR,以放大感兴趣的目标。嵌套方法用于为每个位点设计一组两个底漆。这有助于提高特异性。每个目标分别被放大,用两个不同的引底对来优化PCR条件(即,Pou5f1位点的底漆对A和B,T位点的底漆对C和D),并导致DNA涂片约400bp(图2C)。或者,进行了多路 PCR 以同时放大目标 A 和 C(图 2D),并在纯化后产生类似的片段大小(图 2D)。用于4C库制备的引素(Pou5f1和T的孔)见表6。
在数据分析方面,原始测序读取首先与 refence mm10 小鼠基因组对齐,全部重复,并且删除了低质量 (< 20) 读取。对于每个诱饵,通过计算读取片段的数量获得每个限制片段的信息,并获取原始接触配置文件。接下来,感兴趣区域被定义为所有限制片段,距离诱饵2 kbp和250 kbp。通过按顺序聚合相邻的限制片段来平滑轮廓,直到在感兴趣区域达到原始接触总数的 5% 的阈值,从而增加了每个限制片段的大小。为了确保集成复制和比较条件,我们在限制片段级别上包括斜率和随机截取。每个条件的平均轮廓和它们之间的折叠变化被绘制,如图3所示。在EB分化过程中,增强剂和多能基因Pou5f1的促进者之间的接触减少,而mesendoderm谱系指导转录因子T的增强剂-促进者接触增加(图3),为这些发育增强剂提供了功能见解。

图1:mESC和衍生胚胎体的代表性图像。第0天mESC在无血清条件下(左)和均质第6天EB(右)中由倒置显微镜观察。刻度条 = 500 μm。请点击此处查看此图形的较大版本。
{kind=link}

图 2:协议的 4C 工作流和协议主要步骤的代表性图像。(A) 定量 4C 的原理图工作流。RS = 限制站点;美国 = 上游;DS = 下游;UP = 通用引底;D = RS 和 DS 之间的距离理想情况下应为 5-15 bp. (B) MboI 消化染色质 (I)、 核内联扎染色质 (II) 和声波染色质 (III) 的示例。左侧的数字表示由每个样本运行的 DNA 梯确定的 DNA 大小。(C) 两个位点的 PCR 扩增示例:Pou5f1(引质 A 和 B)和T(底漆 C 和 D)。(D) 使用引物 A 和 C. ES = 胚胎干细胞在Pou5f1和T位点进行多路 PCR 扩增的示例;EB = 胚胎体。请点击此处查看此图形的较大版本。
{kind=link}

图 3:4C 配置文件的示例。定量4C配置文件的诱饵位于Pou5f1和T基因促进器在mESC和第6天EBS中测定。顶部面板显示了由两个独立的生物复制产生的平均接触图;底部面板显示第 6 天 EB 与 mESC 的平均接触折叠变化(两个复制的平均)。浅蓝色框指示增强剂在分化过程中动态变化的位置。图改编自田等人24。请点击此处查看此图形的较大版本。
{kind=link}
| 对于 5mL | |
| 1M 三轮车-HCl,pH8.0 | 50 μL |
| 5M NaCl | 10 μL |
| 10% Igepal CA630 | 100 μL |
| 50倍罗氏全蛋白酶抑制剂 | 100 μL |
| 米利Q水 | 4.74 mL |
表 1:Lysis 缓冲区。
| 用于 1000μL | |
| 米利Q水 | 869 μL |
| 10X NEB T4 DNA 加糖缓冲器 | 120 μL |
| 20mg/mL 牛血清白蛋白 | 6 μL |
| 2000 U/μL T4 DNA联利塞 | 5 μL |
表2:连接母体混合制备。
| 用于 15 μL | |
| 5X 快速连接反应缓冲器 | 10 μL |
| NEBNext 适配器 | 3 μL |
| 快速 T4 DNA 连接 | 2 μL |
表3:适配器连接反应。
| PCR 设置 | |
| 适配器在死时连接库 | 10 μL |
| PCR 级水 | 20.25 μL |
| 10 μM 目标特定引底器 | 3.75 μL |
| 10 μM NEB 指数底漆 | 3.75 μL |
| 赫库拉塞 II 5X 缓冲器 | 10 μL |
| 10 毫米 dNTP | 1.25 μL |
| 赫库雷酶II聚合酶 | 1 μL |
| 总容量 | 50 μL |
| PCR 程序 | |
| 第 1 步: 98 °C - 2 分钟 | |
| 第 2 步:98 °C - 20s | |
| 第 3 步:65 °C - 30s | |
| 第4步:72 °C - 45s | |
| 步骤 5:转到步骤 2,总共进行 15-18 个周期 | |
| 步骤 6: 72 °C - 3 分钟 | |
| 步骤 7: 4 °C + 保持 |
表4:4C染色质相互作用库扩增,首次PCR。
| 嵌套 PCR 设置 | |
| 第一个PCR的DNA片段 | 10 μL |
| PCR 级水 | 20.25 μL |
| 10 μM 特异性引底器_P5 光明素底漆 | 3.75 μL |
| 10 μM P7 光明素底漆 | 3.75 μL |
| 赫库拉塞 II 5X 缓冲器 | 10 μL |
| 10 毫米 dNTP | 1.25 μL |
| 赫库雷酶II聚合酶 | 1 μL |
| 总容量 | 50 μL |
| 嵌套 PCR 程序 | |
| 第 1 步: 98 °C - 2 分钟 | |
| 第 2 步:98 °C - 20s | |
| 第 3 步:65 °C - 30s | |
| 第4步:72 °C - 45s | |
| 步骤 5:转到步骤 2,总共进行 15-18 个周期 | |
| 步骤 6: 72 °C - 3 分钟 | |
| 步骤 7: 4 °C + 保持 |
表5:4C染色质相互作用库扩增,嵌套PCR。
| 名字 | 序列 (5'-3') |
| DS-Oct4-A | 阿加塔格格加加加查查查委员会 反恐委员会CTCTCTCTCTCTCTAAAGAAA委员会 |
| 美国-10月4-A | TCTCTTGCAAGAACTAAGCACGCC |
| DS-10月4-B | 阿加塔格格加加加查查查委员会 CTCTCCGCTGTGGGGGGCTGGGGCGGCT |
| 美国-10月4-B | 阿高格格格格格格格格卡格格格茨 |
| DS-T-C | 阿加塔格格加加加查查查委员会 反恐委员会,CTCTCTGGTGCCTCCGCCAAGGC |
| 美国-T-C | 关贸总协定 |
| DS-T-D | 阿加塔格格加加加查查查委员会 CTCTCCGCTCTCTCTG加加格加加加加格格格格格格格格格格格格格格格格格格格格 |
| 美国-T-D | GCTGAGGCTG加加格加加加加加加加加加 |
| UP-4C | 卡阿加加加CGGCATAC加加 |
| 阿达普-伊1 | 卡阿加加加加察察加加茨格特格格特格加茨加格塔加加加加 CGTGTGCTCTCGATC |
| 阿达普-伊2 | 卡阿加加加加察加察加塔加察察茨格格格加茨塔加加 CGTGTGCTCTCGATC |
| 阿达普-伊3 | 卡阿加加加加察加茨加加茨加塔格格加茨加加加加加 CGTGTGCTCTCGATC |
| 阿达普-伊4 | 卡阿加加加加察加察加加加加格格格格格格格加茨加茨加加加加加加 CGTGTGCTCTCGATC |
表6:用于4C库制备的入门。
讨论
吊落培养法不需要额外的生长因子或细胞因子,可从预定数量的mESCs5中产生均匀的EBs种群。在这里,我们描述了一个定量4C协议,该协议从UMI-4C方法改编而成,用于量化EB分化模型中系系特定转录因子的增强剂-促进者接触。在EB分化过程中,我们确定了以动态方式接触Pou5f1和T基因启动子的染色质区域。Pou5f1 在 EB 分化期间受到抑制,Pou5f1 促进剂与其远端增强剂之间的接触频率降低。相反,T在EB分化期间得到增强,我们确定了三个增强剂,其接触频率与启动器的接触频率降低(图3)。为了确认鉴定,染色质免疫沉淀(ChIP)对活性组蛋白标记H3K27ac的测定可以进行24,因为这个组蛋白标记已被证明与增强剂激活相关,增强剂在失活11期间失去这个标记。
一种标准的4C技术被广泛用于调查特定基因组位点的染色质接触剖面25。然而,这种方法很难定量解释,即使在广泛的规范化26,27,28,由于PCR片段大小的异质性和无法区分PCR重复的异质性引入。26,27,28我们的定量4C方法与UMI-4C技术基本相同,该技术允许使用声波和嵌套连接介导的PCR步骤对单个分子进行定量,以绕过经典4C方法23的限制。然而,与使用唯一分子标识符的UMI-4C不同,我们的定量4C协议允许根据声波步骤产生的特定DNA断裂对单个分子进行定量。它使我们的协议与商业DNA库制备试剂盒兼容,无需使用具有唯一分子标识符的引素。
我们的协议涉及几个应考虑的关键步骤。与经典的4C方法28一样,我们协议的关键因素是3C分子制备过程中的消化和结扎效率。消化/结扎效率低会大大降低与感兴趣的片段相互作用的复杂性,从而降低分辨率。如前所述,该协议的另一个关键步骤是为库扩增设计引素。第二个PCR反应引漆应位于5-15 nt从询问限制位点。在读取 75 nt 排序中,这允许至少 40 nt 剩余捕获长度进行映射。第一次PCR反应中使用的引漆应设计在第二个引底漆的上游,没有重叠,并且两者应足够具体,以确保有效的DNA扩增。对于复用,引素应独立设计,目标是熔融温度 (Tm) 60-65 °C。此外,对于其他3C技术,定量4C方法的分辨率由协议25中使用的限制性酶决定。该协议使用具有4bp识别位点(MboI)的限制酶。这种酶的最大分辨率约为500bp,但这是高度依赖位,很少实现。另一个限制是,位于同一限制片段中的元素之间发生的交互是无法检测到的。此外,在一个限制站点的距离处发生的交互无法与未消化的背景区分开来。在连接之前使用填充步骤可能允许检测这些相互作用。
定量4C非常适合查询目标位点的染色质触点。但是,特定的 PCR 放大步长限制了可同时调查的位点数。增加目标位点数的一种方法是多路复用 PCR 步骤以同时放大多个目标,但这需要所用引素的兼容性,并在实现之前测试每个引素对。如果全球需要促进器的染色质结构变化,那么全基因组方法(如Hi-C、PC Hi-C或HiChIP)将更适合29、30、31。30,3129
披露声明
作者没有什么可透露的。
致谢
我们要感谢勒迪利、斯塔德胡德和格拉夫实验室成员的建议和讨论。G.S.由玛丽·斯克洛多斯卡-居里研究金(H2020-MSCA-IF-2016,miRStem)支持,T.V.T由胡安·德拉西尔瓦博士后研究金(MINECO,FJCI-2014-22946)支持。这项工作得到了欧洲研究理事会根据第7框架方案FP7(ERC协同赠款4D-Genome,授予协议609989T.),西班牙经济,工业和竞争力部(MEIC)EMBL伙伴关系,塞韦罗奥乔亚中心2013-2017年和CERCA方案总统一的加泰罗尼亚。
材料
| Name | Company | Catalog Number | Comments |
| 0.1% EmbryoMax gelatin | EMD Millipore | ES-006-B | Cell culture |
| 0.25% Trypsin-EDTA | 25200072 | ||
| AMPure XP | Beckman Coulter | 10136224 | 4C/DNA purification |
| B27 supplement | Gibco | 17504044 | Cell culture |
| Beta-mercaptoethanol | Gibco | 31350010 | Cell culture |
| Bioruptor Pico | Diagencode | B01060010 | 4C/sonication |
| BSA | NEB | B9000S | 4C |
| CHIR99021 | Selleck Chemicals | S1263 | Cell culture |
| CIP | NEB | M0212 | 4C |
| cOmplete Protease Inhibitor Cocktail | Roche | 4693116001 | 4C |
| DMEM/F12 medium | Gibco | 11320033 | Cell culture |
| dNTP | NEB | N0447S | 4C |
| ESGRO Leukaemia Inhibitory Factor (LIF) | EMD Millipore | ESG1107 | Cell culture |
| Formaldehyde solution (37%) | Sigma | 252549-25ML | 4C |
| Glycin | Sigma | GE17-1323-01 | 4C |
| Glycogen | ThermoFischer | R0551 | 4C |
| Herculase II Fusion DNA polymerase | Agilent | 600675 | 4C |
| IGEPAL CA-630 | Sigma | I3021-50ML | 4C |
| Knockout DMEM | 10829018 | ||
| L-glutamine | Gibco | 25030081 | Cell culture |
| MboI | NEB | R0147M | 4C |
| MEM non-essential amino acids | Gibco | 11140050 | Cell culture |
| N2 supplement | Gibco | A1370701 | Cell culture |
| NEBNext DNA Library prep | NEB | E6040 | 4C |
| NEBuffer 2.1 | NEB | B7202S | 4C/digestion |
| Neurobasal medium | Gibco | 21103049 | Cell culture |
| PD0325901 | Selleck Chemicals | S1036 | Cell culture |
| Penicillin Streptomycin | Gibco | 15140122 | Cell culture |
| Proteinase K | NEB | P8107S | 4C |
| Qubit 4 Fluorometer | ThermoFischer | Q33238 | 4C |
| Qubit dsDNA HS Assay Kit | ThermoFischer | Q32851 | 4C |
| RNase A | ThermoFischer | EN0531 | 4C |
| Sodium pyruvate solution | Gibco | 11360070 | Cell culture |
| StemPro Accutase Cell Dissociation Reagent | Gibco | A1110501 | Cell culture |
| T4 DNA Ligase Reaction Buffer | NEB | B0202S | 4C |
| T4 DNA Ligase Reaction Buffer | NEB | M0202M | 4C |
参考文献
- Evans, M. J., Kaufman, M. H. Establishment in culture of pluripotential cells from mouse embryos. Nature. 292 (5819), 154-156 (1981).
- Martello, G., Smith, A. The nature of embryonic stem cells. Annual Review Cell and Developmental Biology. 30, 647-675 (2014).
- Martin, G. R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proceedings of the National Academy of Science U. S. A. 78 (12), 7634-7638 (1981).
- Loh, K. M., Lim, B., Ang, L. T. Ex uno plures: molecular designs for embryonic pluripotency. Physiological Reviews. 95 (1), 245-295 (2015).
- Sheridan, S. D., Surampudi, V., Rao, R. R. Analysis of embryoid bodies derived from human induced pluripotent stem cells as a means to assess pluripotency. Stem Cells International. 2012, 738910(2012).
- Gaszner, M., Felsenfeld, G. Insulators: exploiting transcriptional and epigenetic mechanisms. Nature Reviews in Genetics. 7 (9), 703-713 (2006).
- Lenhard, B., Sandelin, A., Carninci, P. Metazoan promoters: emerging characteristics and insights into transcriptional regulation. Nature Reviews in Genetics. 13 (4), 233-245 (2012).
- Long, H. K., Prescott, S. L., Wysocka, J. Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution. Cell. 167 (5), 1170-1187 (2016).
- Schoenfelder, S., Fraser, P. Long-range enhancer-promoter contacts in gene expression control. Nature Reviews in Genetics. 20 (8), 437-455 (2019).
- Spitz, F., Furlong, E. E. Transcription factors: from enhancer binding to developmental control. Nature Reviews in Genetics. 13 (9), 613-626 (2012).
- Creyghton, M. P., et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proceedings of the National Academy of Sciences U. S. A. 107 (50), 21931-21936 (2010).
- Heintzman, N. D., et al. Histone modifications at human enhancers reflect global cell-type-specific gene expression. Nature. 459 (7243), 108-112 (2009).
- Klemm, S. L., Shipony, Z., Greenleaf, W. J. Chromatin accessibility and the regulatory epigenome. Nature Reviews in Genetics. 20 (4), 207-220 (2019).
- Rada-Iglesias, A., et al. A unique chromatin signature uncovers early developmental enhancers in humans. Nature. 470 (7333), 279-283 (2011).
- Lettice, L. A., et al. Development of five digits is controlled by a bipartite long-range cis-regulator. Development. 141 (8), 1715-1725 (2014).
- Dekker, J., Rippe, K., Dekker, M., Kleckner, N. Capturing chromosome conformation. Science. 295 (5558), 1306-1311 (2002).
- de Wit, E., de Laat, W. A decade of 3C technologies: insights into nuclear organization. Genes and Development. 26 (1), 11-24 (2012).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Simonis, M., et al. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nature Genetics. 38 (11), 1348-1354 (2006).
- Splinter, E., de Wit, E., van de Werken, H. J., Klous, P., de Laat, W. Determining long-range chromatin interactions for selected genomic sites using 4C-seq technology: from fixation to computation. Methods. 58 (3), 221-230 (2012).
- Stadhouders, R., et al. Multiplexed chromosome conformation capture sequencing for rapid genome-scale high-resolution detection of long-range chromatin interactions. Nature Protocols. 8 (3), 509-524 (2013).
- van de Werken, H. J., et al. Robust 4C-seq data analysis to screen for regulatory DNA interactions. Nature Methods. 9 (10), 969-972 (2012).
- Schwartzman, O., et al. UMI-4C for quantitative and targeted chromosomal contact profiling. Nature Methods. 13 (8), 685-691 (2016).
- Tian, T. V., et al. Whsc1 links pluripotency exit with mesendoderm specification. Nature Cell Biology. 21 (7), 824-834 (2019).
- Chen, H., et al. Dynamic interplay between enhancer-promoter topology and gene activity. Nature Genetics. 50 (9), 1296-1303 (2018).
- Apostolou, E., et al. Genome-wide chromatin interactions of the Nanog locus in pluripotency, differentiation, and reprogramming. Cell Stem Cell. 12 (6), 699-712 (2013).
- de Wit, E., et al. The pluripotent genome in three dimensions is shaped around pluripotency factors. Nature. 501 (7466), 227-231 (2013).
- Krijger, P. H. L., Geeven, G., Bianchi, V., Hilvering, C. R. E., de Laat, W. 4C-seq from beginning to end: A detailed protocol for sample preparation and data analysis. Methods. , (2019).
- Mumbach, M. R., et al. HiChIP: efficient and sensitive analysis of protein-directed genome architecture. Nature Methods. 13 (11), 919-922 (2016).
- Rao, S. S., et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell. 159 (7), 1665-1680 (2014).
- Schoenfelder, S., Javierre, B. M., Furlan-Magaril, M., Wingett, S. W., Fraser, P. Promoter Capture Hi-C: High-resolution, Genome-wide Profiling of Promoter Interactions. Journal of Visualized Experiments. (136), e57320(2018).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。