
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
DNA系留RNA聚合酶用于可编程体外转录和分子计算
摘要
我们描述了一种新型DNA系留T7 RNA聚合酶的工程设计,以调节体外转录反应。我们讨论了蛋白质合成和表征的步骤,验证了概念验证转录调控,并讨论了其在分子计算、诊断和分子信息处理中的应用。
摘要
DNA纳米技术使核酸的可编程自组装成用户规定的形状和动态,适用于各种应用。这项工作表明,DNA纳米技术的概念可用于编程噬菌体衍生的T7 RNA聚合酶(RNAP)的酶活性,并构建可扩展的合成基因调节网络。首先,通过表达N末端SNAP标记RNAP以及随后SNAP标签与苄基胍宁(BG)修饰的寡核苷酸的化学偶联来设计寡核苷酸系留的T7 RNAP。接下来,核酸链置换用于按需编程聚合酶转录。此外,辅助核酸组装体可用作"人工转录因子",以调节DNA程序化的T7 RNAP与其DNA模板之间的相互作用。这种体外转录调节机制可以实现各种电路行为,如数字逻辑、反馈、级联和多路复用。这种基因调控架构的可组合性促进了设计抽象、标准化和缩放。这些功能将使体外遗传设备的快速原型设计成为可能,用于生物传感、疾病检测和数据存储等应用。
引言
DNA计算使用一组设计的寡核苷酸作为计算介质。这些寡核苷酸被编程为序列,以根据用户指定的逻辑动态组装并响应特定的核酸输入。在概念验证研究中,计算的输出通常由一组荧光标记的寡核苷酸组成,可以通过凝胶电泳或荧光板读数器进行检测。在过去的30年中,已经证明了越来越复杂的DNA计算电路,例如各种数字逻辑级联,化学反应网络和神经网络1,2,3。为了帮助制备这些DNA回路,数学模型已被用于预测合成基因回路4,5的功能,并且已经开发了用于正交DNA序列设计6,7,8,9,10的计算工具。.与硅基计算机相比,DNA计算机的优势包括它们能够直接与生物分子连接,在没有电源的情况下在溶液中运行,以及它们的整体紧凑性和稳定性。随着下一代测序的出现,合成DNA计算机的成本在过去二十年中一直在以比摩尔定律11更快的速度下降。这种基于DNA的计算机的应用现在开始出现,例如用于疾病诊断12,13,用于为分子生物物理学提供动力14,以及作为数据存储平台15。

图1:脚趾介导的DNA链位移的机制。 δ,脚趾是部分双工上的自由、未绑定序列。当在第二条链上引入互补域(δ*)时,自由δ域充当杂交的支架,允许链的其余部分(ɑ*)通过称为链迁移的压缩/解压缩可逆反应缓慢地取代其竞争对手。随着δ长度的增加,正向反应的ΔG减小,位移更容易发生。 请点击此处查看此图的放大版本。
{kind=link}
迄今为止,大多数DNA计算机都利用动态DNA纳米技术领域公认的基序,称为脚趾介导的DNA链置换(TMDSD,图1)16。该基序由部分双链DNA(dsDNA)双链组成,显示短的"脚趾"悬垂(即7至10个核苷酸(nt))。核酸"输入"链可以通过脚趾与部分双链相互作用。这导致其中一条链从部分双工移出,然后这个释放的链可以作为下游部分双工的输入。因此,TMDSD可实现信号级联和信息处理。原则上,正交TMDSD基序可以在解决方案中独立运行,从而实现并行信息处理。TMDSD反应存在许多变化,例如脚趾介导的DNA链交换(TMDSE)17,具有双长结构域18的"无泄漏"脚趾,序列不匹配的脚趾19和"手持"介导的链位移20。这些创新的设计原则允许更精细地调整TMDSD能量学和动力学,以提高DNA计算性能。
合成基因回路,如转录基因回路,也能够计算21、22、23。这些回路由蛋白质转录因子调节,蛋白质转录因子通过与特定的调节DNA元件结合来激活或抑制基因的转录。与基于DNA的电路相比,转录电路具有几个优点。首先,酶转录比现有的催化DNA回路具有更高的周转率,因此每个输入拷贝产生更多的输出拷贝,并提供更有效的信号放大手段。此外,转录电路可以产生不同的功能分子,如适配子或编码治疗蛋白的信使RNA(mRNA)作为计算输出,可以用于不同的应用。然而,当前转录电路的一个主要限制是它们缺乏可扩展性。这是因为基于正交蛋白质的转录因子集非常有限,并且新蛋白质转录因子的从头设计在技术上仍然具有挑战性且耗时。

图2:"系绳"和"笼式"聚合酶复合物的提取和机理(A和B)寡核苷酸系绳通过SNAP标签反应酶被酶标记为T7聚合酶。由具有系绳补体悬垂的"人造"T7启动子组成的笼子使其能够与系绳杂交并阻断转录活性。(C)当操作者(a*b*)存在时,它结合到寡核苷酸系绳(ab)上的脚趾并取代笼子的b*区域,从而允许转录发生。这个数字是从Chou和Shih27修改而来的。缩写:RNAP = RNA 聚合酶。请点击此处查看此图的放大版本。
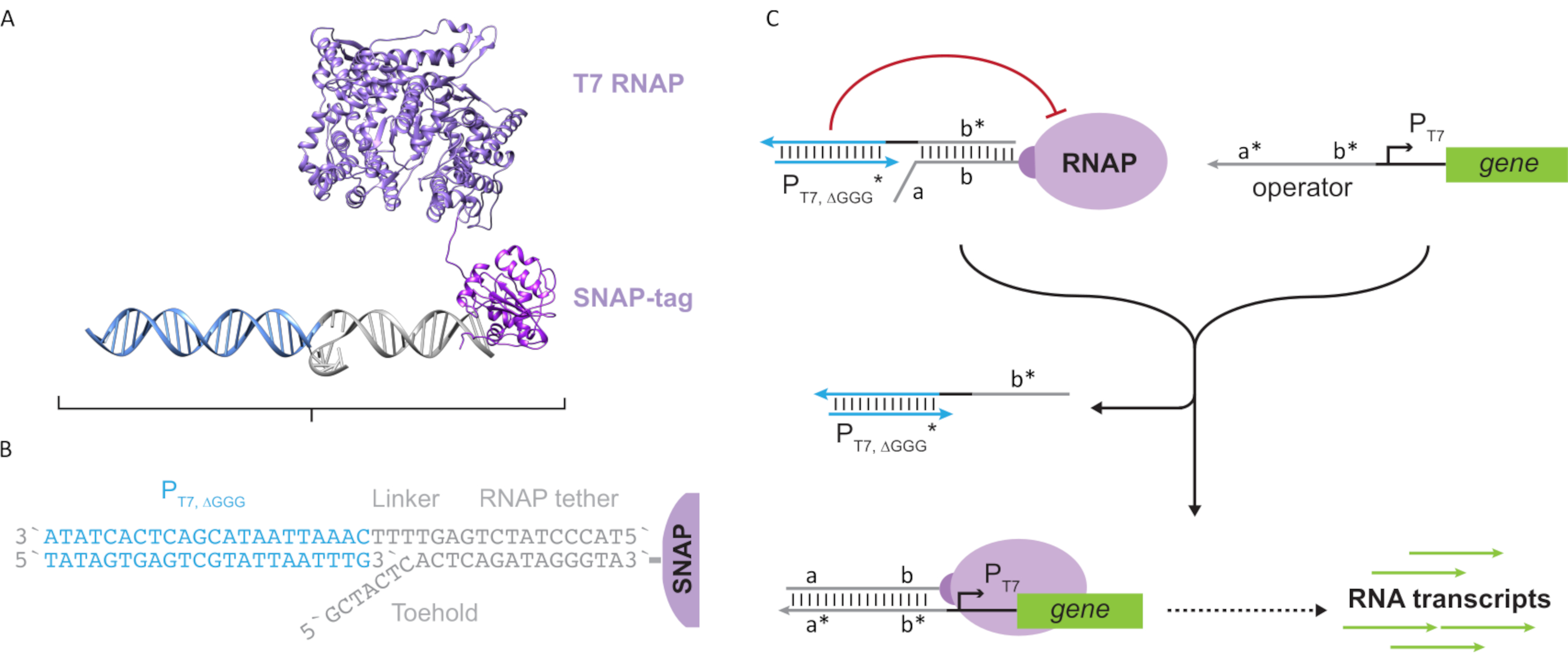{kind=link}
本文介绍了一种用于分子计算的新型构建块,它将转录电路的功能与基于DNA的电路的可扩展性相结合。该构建块是与单链DNA系绳共价连接的T7RNAP(图2A)。为了合成这种DNA系留的T7 RNAP,将聚合酶融合到N端SNAP标签24中并在大肠杆菌中重组表达。然后将SNAP标签与与BG底物功能化的寡核苷酸反应。寡核苷酸系绳允许分子来宾通过DNA杂交定位在靠近聚合酶的位置。其中一位客人是一种被称为"笼子"的竞争性转录阻滞剂,它由一个"人造"T7启动子DNA双链体组成,下游没有基因(图2B)。当通过其寡核苷酸系绳与RNAP结合时,笼子通过与其他DNA模板竞争RNAP结合来阻止聚合酶活性,从而使RNAP处于"OFF"状态(图2C)。
为了将聚合酶激活到"ON"状态,设计了具有基因T7启动子上游的单链"操作员"结构域的T7 DNA模板。操作结构域(即结构域a*b*图2C)可以设计为通过TMDSD将笼子从RNAP取代,并将RNAP定位在基因T7启动子的近端,从而启动转录。或者,还设计了DNA模板,其中操作序列与称为"人工转录因子"的辅助核酸链(即图3A中的TFA和TFB链)互补。当两条链都被引入反应中时,它们将在操作员现场组装,从而创建一个新的伪连续域a*b*。然后,该结构域可以通过TMDSD取代笼子以启动转录(图3B)。这些链可以外源供应或生产。

图3:通过三组分开关激活剂对聚合酶活性进行选择性编程。 (A)当转录因子(TFA 和TFB)存在时,它们与启动子上游的操作员结构域结合,形成一个伪单链序列(a*b*),能够通过脚趾介导的DNA置换来置换笼子。(B) 这个 a*b* 结构域可以通过 TMDSD 取代笼子以启动转录。这个数字是从Chou和Shih27修改而来的。缩写: TF = 转录因子;RNAP = RNA聚合酶;TMDSD = 脚趾介导的 DNA 链置换。 请点击此处查看此图的放大版本。
{kind=link}
使用基于核酸的转录因子进行体外转录调控允许可扩展地实现复杂的电路行为,如数字逻辑、反馈和信号级联。例如,人们可以通过设计核酸序列来构建逻辑门级联,使得来自上游基因的转录本激活下游基因。利用该技术所提出的级联和多路复用能力的一个应用是开发更复杂的分子计算电路,用于便携式诊断和分子数据处理。此外,集成分子计算和从头RNA合成功能可以实现新的应用。例如,分子电路可以设计用于检测用户定义的RNA的一种或组合,作为输入和输出治疗性RNA或mRNA编码功能肽或蛋白质的mRNA,用于床旁医疗应用。
研究方案
1. 缓冲液制备
注意:蛋白质纯化缓冲液的制备可以在任何一天进行;在这里,它是在开始实验之前完成的。
- 制备含有50mM三(羟甲基)氨基甲烷(Tris),300mM氯化钠(NaCl),5%甘油和5mM β巯基乙醇(BME)的裂解/平衡缓冲液,pH 8。在50 mL离心管中加入1.5 mL 1M Tris,1.8 mL 5M NaCl,1.5 mL甘油,25.2mL去离子水(ddH 2 O),并在使用前加入10.5μL14.2 M BME。
注意:Tris可引起急性毒性;因此,避免吸入其灰尘,并避免皮肤和眼睛接触。BME是有毒的,只能在通风橱中使用。重要的是在重悬和细胞裂解之前最后添加BME。裂解缓冲液式见表1。 - 准备含有50 mM Tris,800 mM NaCl,5%甘油,5 mM BME和20 mM咪唑的洗涤缓冲液(pH 8)。将 1.5 mL 1 M Tris、4.8 mL 5 M NaCl、1.5 mL 甘油和 22.2 mL ddH2O 加入 50 mL 离心管中。在使用前,将7μL14.2 M BME和200μL2M咪唑加入20mL上述溶液中。
注意:为防止咪唑引起的急性毒性,请使用个人防护装备。重要的是最后添加BME和咪唑,就在将蛋白质从柱中洗出之前。洗涤缓冲液配方见表2。 - 制备含有50mM Tris,800mM NaCl,5%甘油,5mM BME和200mM咪唑的洗脱缓冲液(pH8)。将 0.5 mL 1 M Tris、1.6 mL 5 M NaCl、0.5 mL 甘油和 6.4 mL ddH2O 加入 15 mL 离心管中。在使用前,将3.5μL的14.2 M BME和1mL的2M咪唑加入10mL上述溶液中。
注意:重要的是最后添加BME和咪唑,就在将蛋白质从柱中洗脱出来之前。洗脱缓冲液配方见表3。 - 准备2x储存缓冲液(与甘油1:1混合),其中含有100mM Tris,200mM NaCl,40mM BME和2mM乙二胺四乙酸(EDTA),0.2%的非离子表面活性剂(见 材料表)。通过将5 mL 1 M Tris,2 mL 5 M NaCl,42.56 mL ddH2O,200μL0.5 M EDTA,100μL非离子表面活性剂加入50mL离心管,制备50mL储存缓冲液。混合直至溶液均匀,通过0.2μm注射器过滤器过滤储存缓冲液,并在使用前向上述溶液中加入140.8μL BME。
注意:为避免EDTA引起的急性毒性,请避免吸入其灰尘,并避免皮肤和眼睛接触。重要的是最后添加BME,并在储存纯化的蛋白质之前将整个储存缓冲液以1:1的比例与甘油混合。有关储存缓冲液公式,请参见表 4。
2. 过夜培养:第1天
- 通过将500mg卡那霉素溶解在10mL的ddH2O中来制备1,000x卡那霉素储备。
注意:使用个人防护装备,以防止卡那霉素引起的急性毒性。 - 将20μL1,000x卡那霉素储备液加入20mL溶菌汤中。使用无菌移液器吸头,戳翻变形的BL21 大肠杆菌 甘油原液,然后通过将尖端引入生长培养基汤中来接种培养物。

图4:SNAP T7 RNAP的质粒图谱。 质粒在pQE-80L骨架上的lac抑制剂(lacI)下编码含有N端组氨酸标签(6x His)和SNAP标签结构域(SNAP T7 RNAP)的T7 RNAP。其他特征包括卡那霉素耐药性(KanR)和氯霉素耐药性(CmR)基因。缩写:RNAP = RNA 聚合酶。 请点击此处查看此图的放大版本。
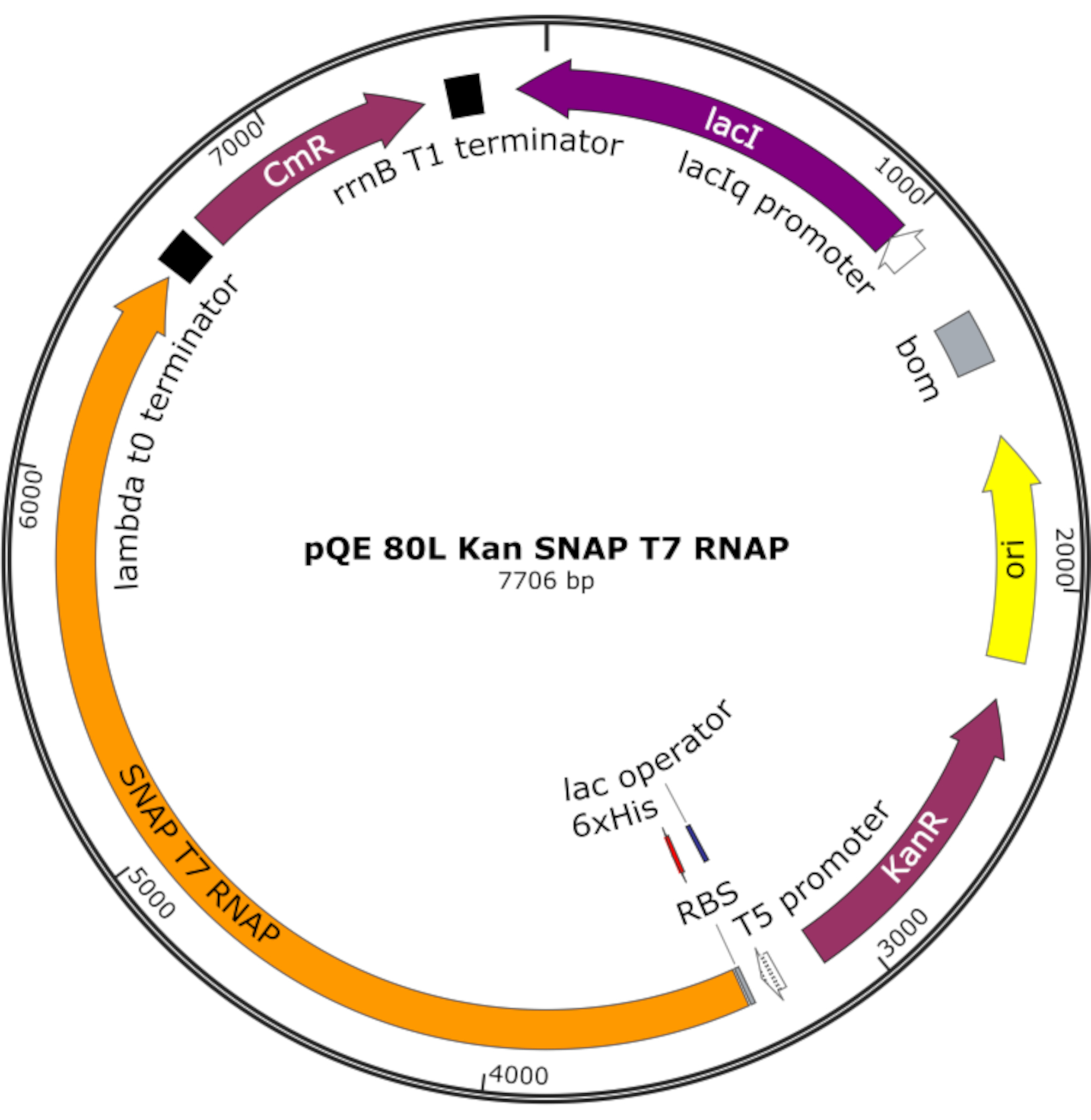{kind=link}
注:质粒编码含有N端组氨酸标签和SNAP标签结构域(SNAP T7 RNAP)的T7 RNAP,以及pQE-80L骨架下的卡那霉素抗性基因(图4)25。
- 再次,将20μL1,000x卡那霉素储备液加入含有20mL溶菌汤的单独培养瓶中,并将其作为对照孵育。
- 将两个样品(从步骤2.2和2.3)在37°C下孵育12-18小时,同时以 10×g旋转 。
3. 细胞生长和诱导:第2天
- 接种400mL含有400μL卡那霉素储备液的溶菌肉汤和步骤2.4的4mL过夜生长培养物。将培养瓶孵育在37°C,同时以10×g旋转。
- 一旦培养物达到约0.5的600nm处的光密度(OD),从生长瓶中取出1mL样品作为对照。将对照样品储存在4°C。
- 通过每100mL培养物加入40μL1M IPTG,用异丙基β-D-1-硫代乳糖吡喃糖苷(IPTG)诱导细胞,以达到0.4mM IPTG的最终浓度。将样品在37°C下孵育3小时,以10×g旋转,然后以8,000×g旋转诱导培养物10分钟以沉淀细胞。 除去上清液,并将沉淀储存在-20°C直至进一步使用。
注意:为避免IPTG引起的急性毒性,请避免吸入其灰尘,并避免皮肤和眼睛接触。如有必要,您可以在此处暂停实验,并在第二天继续。
4.细胞裂解,蛋白质纯化:第3天
- 将储存的细胞沉淀与10mL裂解缓冲液重悬于冰上,并轻轻旋转以确保整个沉淀重悬。然后,将1 mL样品移入10个1.5mL管中,这些管子保持在冰上。
- 以"1"的幅度设置对每个样品进行超声处理,脉冲2 s,占空比为50%,持续时间为30 s。在每个样品之前和之后,用70%乙醇和ddH2O清洁超声处理尖端。在超声处理期间和之后将所有样品保持在冰上。
注意:保持70%乙醇远离热源和明火。 - 将带镍的硝酸三乙酸(Ni-NTA)纯化离心柱平衡至4°C的工作温度。 将色谱柱置于4°C,并在使用过程中保持在冰上。
- 将10个1mL样品在4°C下以15,000×g离心20分钟。 小心地移出含有重组RNAP的上清液,而不会干扰沉淀。如有必要,使用额外的平衡缓冲液将总体积调节至≥6 mL。
- 轻轻地从Ni-NTA旋转柱上取下底部卡舌,以允许流过柱。将色谱柱放入离心管中,并保持在冰上。
注:使用 50 mL 离心管和 3 mL Ni-NTA 离心柱。 - 将色谱柱在700×g和4°C下离心2分钟以除去储存缓冲液。通过向色谱柱中加入6 mL平衡缓冲液来平衡色谱柱。让缓冲液完全进入树脂床。
- 通过在700×g和4°C下离心2分钟,从色谱柱中除去平衡缓冲液。在将准备好的细胞提取物添加到色谱柱之前,请在色谱柱上放置底部塞子以避免丢失任何产品。然后,将细胞提取物加入色谱柱中,并在4°C下在轨道振荡器上混合30分钟。
- 从色谱柱上取下底塞,并将色谱柱置于标有流经的50 mL离心管中。将柱以700×g离心2分钟以收集流经。
- 向色谱柱中加入6 mL洗涤缓冲液以洗涤树脂。将色谱柱以700×g离心2分钟,以在标有洗涤剂1的新离心管中收集馏分。重复此步骤两次,总共3个单独的馏分,并将馏分收集在单独的离心管中(洗涤2和洗涤3)。
- 加入3mL洗脱缓冲液以洗脱树脂中的His标记蛋白质。将色谱柱以700×g离心2分钟,以在标有洗脱液1的新离心管中收集馏分。重复此步骤两次,总共3个单独的馏分,并将馏分收集到单独的离心管中(洗脱液2和洗脱液3)。
- 结合洗脱液并进行脱盐以除去蛋白质溶液中的盐。
- 移液器 15 mL 0.05 % w/v 聚山梨醇酯 20 在 100 kDa 离心过滤装置上。以4,000×g离心40分钟,并丢弃流出物。
- 使用包衣过滤器将洗脱液1,2和3(9mL总蛋白质洗脱液+ 6mL储存缓冲液)浓缩至~1,500μL。将过滤器以3,220×g离心20分钟,并轻轻移液洗膜以防止沉淀。
- 用储存缓冲液将样品稀释至15 mL。通过重复步骤 4.11.2 两次,使用存储缓冲区 1:1,000 执行缓冲区交换。
- 通过测量280nm处馏分的吸光度来量化纯化的蛋白质。用储存缓冲液(4°C下2x储存缓冲液)清空分光光度计。轻轻混合混合混合洗脱液的样品并测量其吸光度。
注意:在蛋白质样品的1x,10x和50x稀释液中执行三个单独的读数,以平均和定量蛋白质。在储存缓冲液中稀释样品。 - 使用2x储存缓冲液将蛋白质样品调节至100μM。用100%甘油以1:1的体积稀释调整后的样品。将所得蛋白质溶液储存在-80°C。
5. 十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)蛋白质产物分析:第3天
- 运行SDS-PAGE凝胶进行蛋白质分析。将9μL样品与3μL四倍十二烷基硫酸锂(LDS)蛋白上样染料混合。将样品在95°C下加热10分钟。
- 将样品上样到4-12%双三SDS-PAGE凝胶设置中。将蛋白质分子量标准品加载到孔1中,然后用样品(从左到右):流过,洗涤1,洗涤2,洗涤3,洗脱1,洗脱2,洗脱3和总脱盐洗脱。
注: 表5 包含SDS-PAGE凝胶的样品上样表。 - 将加载的凝胶样品在2-(N-吗啉)乙烷磺酸(MES)缓冲液中在200 V下运行35分钟,使用200 mL ddH 2 O在干净的托盘中冲洗凝胶三次,每次10分钟,轻轻搅拌以从凝胶基质中除去任何SDS。
注意:穿戴个人防护装备,以避免由于MES引起的急性毒性。 - 用20 mL考马斯蓝染色凝胶,并在室温下温和搅拌凝胶孵育过夜。用200 mL ddH 2 O对凝胶进行两次去污,每次1小时,并在轨道振荡器上轻轻搅拌。
注意:长时间洗涤凝胶或频繁更换水将增强灵敏度。此外,将折叠的精密擦拭组织放入容器中以吸收多余的染料将加速去污过程。
6. 通过体外转录对SNAP T7 RNAP进行功能验证
注意:该协议使用DNA模板,该模板编码荧光西兰花RNA适配子,并允许使用荧光来监测荧光板读数仪上转录的动力学。
- 设置三种体外转录(IVT)反应,以比较来自商业来源和纯缓冲液对照的SNAP T7 RNAP与野生型(WT)T7 RNAP的活性。将每个反应的体积调节至20μL。
- 通过混合2μL的10x转录缓冲液,0.4μL的25mM核糖核苷三磷酸(rNTP)混合物,5μL的500nM DNA模板,2μL的500nM SNAP T7 RNAP和10.6μL的ddH 2 O来制备SNAP T7IVT反应。
- 通过混合2μL10x转录缓冲液,0.4μL25mM rNTP混合物,5μL500nM DNA模板,2μLWT T7 RNAP和10.6μLdH2O来制备WT RNAP IVT反应。
- 通过混合2μL10x转录缓冲液,0.4μL25mM rNTP混合物,5μL500nM DNA模板和12.6μLdH2O来制备仅缓冲液IVT反应。
注意:最后加入RNAP,将样品保持在冰上,直到其引入。表6、表7和表8包含IVT反应式。
- 在荧光板读数器上监测转录动力学,在37°C下以2分钟的间隔使用470nm的激发波长和512nm的发射波长。
7. BG修饰寡核苷酸的制备:第1天
- 用3'-胺修饰在ddH2O中溶解寡核苷酸至终浓度为1mM。将此 S1标记为 .
- 将25μL1M碳酸氢钠(NaHCO 3),284μL100%二甲基亚砜(DMSO),125μLS1(寡核苷酸储备)和66μL50mM的BG-N-羟基琥珀酰亚胺(NHS)酯(BG-GLA-NHS)用DMSO稀释,将体积调节至500μL,并在室温下以100×g孵育过夜。
注意:保持DMSO远离热源和火焰,因为它是可燃液体。 表9 包含BG与寡核苷酸偶联的反应式。
- 将25μL1M碳酸氢钠(NaHCO 3),284μL100%二甲基亚砜(DMSO),125μLS1(寡核苷酸储备)和66μL50mM的BG-N-羟基琥珀酰亚胺(NHS)酯(BG-GLA-NHS)用DMSO稀释,将体积调节至500μL,并在室温下以100×g孵育过夜。
8. BG-寡核苷酸偶联物乙醇/丙酮沉淀:第2天
- 离心步骤7.1.1的产物。在13,000×g下5分钟。小心地将上清液转移到新鲜试管中并丢弃任何沉淀的BG.将反应分成两个相等的250μL等分试样以防止溢出,并在两个等分试样上执行以下步骤。
- 加入3M乙酸钠体积的 1/10(25μL),然后在100%乙醇(625μL)中加入体积的2.5倍。在-80°C孵育1小时。
注意:在处理醋酸钠(可能对眼睛、皮肤、消化道和呼吸道造成刺激)和乙醇(极易燃,接触时会引起刺激)时,请使用个人防护装备。如有必要,请在此处暂停实验,并在第二天继续。 - 将离心管放入离心机中,并在外边缘标记。在4°C下以17,000×g离心管30分钟。
注意:寡核苷酸沉淀将出现在管的标记边缘。 - 在不干扰沉淀的情况下,丢弃上清液。加入750μL冷藏的70%乙醇,并在4°C下以17,000×g离心10分钟。
- 在不干扰沉淀的情况下,丢弃上清液。加入750μL100%丙酮,并在4°C下以17,000×g旋转10分钟。
注意:处理丙酮时使用个人防护装备,因为它非常易燃,接触时会引起刺激。 - 打开管盖,风干5分钟,通过蒸发除去任何多余的丙酮。将寡核苷酸重新溶解在250μL的1x Tris-EDTA(TE)缓冲液中,以产生〜850μM BG寡核苷酸溶液。
- 重复步骤8.2至8.6,并在70μL的1x TE缓冲液中重新溶解。将此 S2标记为 。
9. 通过凝胶过滤色谱法清除BG-寡核苷酸
- 通过多次大力反转柱子来暂停矩阵;取下顶盖并折断柱底。将色谱柱置于1.5mL离心管中,并在室温下以1,000×g离心管1分钟。 丢弃洗脱的缓冲液和收集管。
注意:防止真空形成很重要。立即使用准备好的色谱柱。 - 将包装好的色谱柱置于干净的1.5 mL离心管中。向柱床中心加入300μL1x TE缓冲液,并以1,000×g离心2分钟以交换缓冲液。 再次,丢弃洗脱的缓冲液和收集管。
- 将缓冲液交换的色谱柱置于干净的1.5 mL离心管中。将多达75μL样品施用于床的中心。以1,000×g旋转4分钟。
注意:请勿打扰床或触摸柱子的侧面;凝胶介质的最高点应指向外转子。 - 从收集管中收集洗脱液,因为它含有纯化的核酸。为了量化样品,测量其在260nm处的吸光度;标记此 S3。
注意:注意测量中使用的路径长度,并使用比尔-朗伯定律计算浓度。
10. BG-寡核苷酸偶联物的变性页面分析
- 浇注18%的三硼酸乙二胺四乙酸酯(TBE)尿素PAGE凝胶。将4.8克尿素,4.5毫升40%丙烯酰胺(19:1)和1毫升10x TBE溶解在2.8毫升ddH2O中;加入5μL四甲基乙二胺(TEMED)并充分混合。用100μL10%过硫酸铵(APS)重复。将溶液倒入空的凝胶盒中,并使其聚合40分钟。
注意:处理尿素(对眼睛和皮肤造成刺激)、丙烯酰胺(有毒和致癌)和TEMED(有毒、易燃、腐蚀性)时,请使用适当的个人防护装备。 表10 包含18%TBE-脲聚丙烯酰胺凝胶的反应式。 - 微波500mL TBE缓冲液(0.5x)2分钟和30秒或直到〜70°C,然后倒入凝胶设备中。制备含有95%甲酰胺+ 1mM EDTA和溴酚蓝的甲酰胺(变性)加载染料。将上样染料与每个样品混合,并将混合物上样到聚丙烯酰胺凝胶上。
注:处理甲酰胺时,请使用适当的个人防护装备,因为它具有致癌性。 表11 包含样品凝胶上样表。 - 将凝胶在270 V下运行35分钟,或直到染料前端迁移到末端。将凝胶置于凝胶盒中,在成像前用菁染料在室温下染色15分钟核酸。
注意:处理菁染料时,请使用适当的个人防护装备,因为它是可燃的。
11. 寡核苷酸与SNAP T7 RNAP的偶联和PAGE分析
- 准备试剂,用于BG-寡核苷酸与SNAP T7 RNAP的分析级偶联:用ddH2O制备9个稀释的单链DNA(ssDNA)寡核苷酸,以产生5:1至1:5的寡核苷酸比值。将蛋白质储备稀释至50μM。
注:示例比率见 表12;这些比率使用50μM的RNAP浓度计算。 - 对于每个稀释的ssDNA寡核苷酸,使10μL含有2μLSNAP缓冲液,4μLBG寡核苷酸和4μLSNAP的反应混合物。
注: 表13 包含SNAP标签标记反应的反应公式。- 再制备两个对照样品:1)通过用ddH2O代替BG寡核苷酸来对照RNAP;2)通过将SNAP T7 RNAP替换为ddH2O(对于SNAP T7 RNAP的最低寡核苷酸浓度)进行DNA对照。将所有样品在室温下孵育1小时,并保存在冰上直到需要。
- 通过将每个样品的2μL加入4μLSNAP缓冲液和2μL蛋白质上样染料中,并在70°C下加热10分钟,设置11个10μL反应。将每个样品的2μL倒入4-12%双-Tris蛋白凝胶上,并在200 V的冰上进行凝胶电泳35分钟。
注: 表14 包含凝胶上样样品的反应公式。- 通过在摇摇杯上进行3次水交换来冲洗SDS,每次洗涤持续10分钟。在成像前用菁染料染色核酸15分钟。使用20 mL考马斯蓝染色剂再次染色凝胶1小时。在成像前用 ddH2O 去污 1 小时(或过夜)。
注意:在凝胶中,其中一种反应将产生最紧密的系留聚合酶以及最少量的过量游离BG-寡核苷酸;这是最佳比率。
- 通过在摇摇杯上进行3次水交换来冲洗SDS,每次洗涤持续10分钟。在成像前用菁染料染色核酸15分钟。使用20 mL考马斯蓝染色剂再次染色凝胶1小时。在成像前用 ddH2O 去污 1 小时(或过夜)。
- 制备试剂,用于制备规模偶联BG-寡核苷酸与SNAP T7 RNAP。以分析尺度中发现的最佳比率进行偶联反应。
注意:在不使用时,将蛋白质放在冰上,尽量减少蛋白质暴露在室温下。
12. 使用离子交换柱纯化寡核苷酸系留SNAP-T7
- 如果与此处列出的说明不同,请遵循制造商的管子设置说明。制备pH值高于蛋白质等电点的纯化缓冲液。
注意:对于该方案中的示例蛋白质,使用10mM磷酸钠缓冲液(pH 7)的纯化缓冲液。- 制备1,000μL洗脱缓冲液,含有最终浓度为50mM Tris和0.5M NaCl。混合 50 μL 1 M Tris、100 μL 5 M NaCl 和 850 μL ddH2O。
注: 表15 包含洗脱缓冲液的反应式。
- 制备1,000μL洗脱缓冲液,含有最终浓度为50mM Tris和0.5M NaCl。混合 50 μL 1 M Tris、100 μL 5 M NaCl 和 850 μL ddH2O。
- 将色谱柱置于2mL离心管中,并用纯化缓冲液以2,000×g洗涤15分钟,或直到所有缓冲液洗脱。丢弃洗脱的缓冲液。
- 用纯化缓冲液以3:1的纯化缓冲液:样品比例稀释每个样品,并将样品一次400μL加载到柱中。以2,000×g旋转10分钟,或直到所有缓冲液都洗脱。收集流出并将其标记为流出。
- 将400μL纯化缓冲液加入柱中心。以2,000×g旋转15分钟,或直到所有缓冲液都洗脱。收集流出物并将其标记为 wash 1。重复两次洗涤2和洗涤3。
- 将50μL洗脱缓冲液加入色谱柱的中心。以2,000×g旋转5分钟,或直到所有缓冲液都洗脱。收集流出物并将其标记为洗脱液 1。对洗脱液2和洗脱3重复两次。
- 池洗脱液1,2和3(标记此 总洗脱液),为每个洗脱液留下一小部分用于凝胶,并测量260nm(A260)和280nm(A280)处的吸光度。测量后,以1:1的比例加入甘油,并储存在-20°C直至进一步使用。
- 使用离心过滤装置(0.5 mL;30 kDa)用2x储存缓冲液(~1:100)缓冲交换 总洗脱液 (标记本 产品)。再次测量 A260/280。以1:1的比例加入甘油,储存在-20°C直至进一步使用。
- 将每个洗脱液上样:流经,洗涤1-3,总洗脱液和产品在4-12%双-三SDS-PAGE凝胶中,以及蛋白质分子量标准品。在200 V下运行35分钟,或直到染料前端迁移到末端。
13. 系留RNA聚合酶活性按需控制的演示
- 准备5x退火缓冲液,其中包含25mM Tris,5mM EDTA和25mM氯化镁(MgCl2)。将每个模板的2.4μL(1μM)与5μL退火缓冲液和14.2μLdH2O混合,形成25μL的1μM dsDNA笼。将该溶液在75°C孵育2分钟。类似地,退火启动子和孔雀石绿色适配体DNA模板的意义和反义链。准备1mM的孔雀石绿草酸盐溶液。
注: 表16 包含5x退火缓冲液的反应式, 表17 包含用于退火的反应式两个ssDNA模板。 - 将系留的SNAP T7 RNAP与dsDNA笼在室温下以1:5摩尔比孵育15分钟至终浓度为500 nM RNAP。保持冰敷,直到需要。
- 将读板器预热至37°C。 在冰上设置三个 25 μL IVT 反应
- 建立含有笼状SNAP T7RNAP与核酸转录因子的反应。在10μLddH2 O中混合2.5μL10x IVT缓冲液,1μL25mM rNTP混合物,1μL1mM孔雀石绿,2.5μLRNA笼混合物,1μM转录因子A和B寡核苷酸链各2.5μL,以及3μL1mM孔雀石绿色适配子模板。
- 建立含有不含核酸转录因子的笼状SNAP T7RNAP的反应。将2.5μL10x IVT缓冲液,1μL25mM rNTP混合物,1μL1mM孔雀石绿,2.5μLRNAP笼混合物和3μL1mM孔雀石绿色适配子模板在15μLddH2O中混合。
- 设置仅包含缓冲液的反应。将2.5μL10x IVT缓冲液,1μL25mM rNTP混合物,1μL1mM孔雀石绿和3μL1mM孔雀石绿色适配子模板混合在17.5μLdH2O中。
注: 表18 包含体外转录反应的一般参考。
- 将每个反应转移到384孔板中。在荧光板读数器上监测孔雀石绿色适配体的转录,在37°C下,以610nm激发和655nm发射。完成后,将盘子放在冰上,直到需要。
- 微波0.5x TBE缓冲液2分钟30秒或直至~70°C。 在变性12%TBE聚丙烯酰胺凝胶中,在280 V下加热的0.5x TBE缓冲液中运行每个孔的RNA产物20分钟,或直到染料前端到达终点。在成像之前,用菁染料核酸染色在眼眶振荡器上染色凝胶10分钟。
注: 表19 包含变性12%TBE-尿素PAGE凝胶的反应公式。
结果

图5:SNAP T7 RNAP表达和体外转录测定的SDS-PAGE分析。(A)SNAP T7 RNAP蛋白纯化分析,SNAP T7 RNAP分子量:119.4kDa。FT=从柱中流出,W1=含有杂质的洗涤缓冲液的洗脱级分,E1-3=含有纯化产物的洗脱级分,并且DE=10倍稀释的脱盐总洗脱。4-12%预制双-Tris蛋白凝胶,染色:考马斯蓝...
讨论
本研究展示了一种受DNA纳米技术启发的方法,通过N末端SNAP标记的重组T7 RNAP与BG功能化的寡核苷酸共价耦合来控制T7 RNA聚合酶的活性,随后用于编程TMDSD反应。根据设计,SNAP标签位于聚合酶的N端,因为野生型T7 RNAP的C端埋在蛋白质结构核心内并与DNA模板28进行重要接触。先前修改聚合酶C末端的尝试已导致酶活性完全丧失,除非引入其他补偿性突变。29,30
披露声明
作者中没有任何竞争性经济利益需要声明。
致谢
L.Y.T.C感谢来自研究探索新前沿(NFRF-E),加拿大自然科学与工程研究委员会(NSERC)发现补助金以及多伦多大学医学设计倡议的慷慨支持,该倡议获得加拿大第一研究卓越基金(CFREF)的资助。
材料
| Name | Company | Catalog Number | Comments |
| 0.5% polysorbate 20 (TWEEN 20) | BioShop | TWN510.5 | |
| 0.5M ethylenediaminetetraacetic acid (EDTA) | Bio Basic | SD8135 | |
| 10 mM sodium phosphate buffer (pH 7) | Bio Basic | PD0435 | Tablets used to make 10 mM buffer |
| 10% ammonium persulfate (APS) | Sigma Aldrich | A3678-100G | |
| 100 kDa Amicon Ultra-15 Centrifugal Filter Unit | Fisher Scientific | UFC910008 | |
| 100% acetone | Fisher Chemical | A18P4 | |
| 100% ethanol (EtOH) | House Brand | 39752-P016-EAAN | |
| 10x in vitro transcription (IVT) buffer | New England Biolabs | B9012 | |
| 10x Tris-Borate-EDTA (TBE) buffer | Bio Basic | A0026 | |
| 1M Isopropyl β- d-1-thiogalactopyranoside (IPTG) | Sigma Aldrich | I5502-1G | |
| 1M sodium bicarbonate buffer | Sigma Aldrich | S6014-500G | |
| 1M Tris(hydroxymethyl)aminomethane (Tris) | Sigma Aldrich | 648311-1KG | |
| 1X Tris-EDTA (TE) buffer | ThermoFisher | 12090015 | |
| 2M imidazole | Sigma Aldrich | 56750-100G | |
| 2-mercaptoethanol (BME) | Sigma Aldrich | M3148 | |
| 3M sodium acetate | Bio Basic | SRB1611 | |
| 40% acrylamide (19:1) | Bio Basic | A00062 | |
| 4x LDS protein sample loading buffer | Fisher Scientific | NP0007 | |
| 5M sodium chloride (NaCl) | Bio Basic | DB0483 | |
| 5mM dithiothreitol (DTT) | Sigma Aldrich | 43815-1G | |
| 6x gel loading dye | New England Biolabs | B7024S | |
| agarose B powder | Bio Basic | AB0014 | |
| BG-GLA-NHS | New England Biolabs | S9151S | |
| BL21 competent E. coli | Addgene | C2530H | |
| BLUeye prestained protein ladder | FroggaBio | PM007-0500 | |
| bromophenol blue | Bio Basic | BDB0001 | |
| coomassie blue (SimplyBlue SafeStain) | ThermoFisher | LC6060 | |
| cyanine dye (SYBR Gold nucleic acid gel stain) | Fisher Scientific | S11494 | |
| cyanine dye (SYBR Safe nucleic acid gel stain) | Fisher Scientific | S33102 | |
| dry dimethyl sulfoxide (DMSO) | Fisher Scientific | D12345 | |
| formamide | Sigma Aldrich | F9037-100ML | |
| glycerol | Bio Basic | GB0232 | |
| kanamycin sulfate | BioShop | KAN201.5 | |
| lysogeny broth | Sigma Aldrich | L2542-500ML | |
| malachite green oxalate | Sigma Aldrich | 2437-29-8 | |
| N,N,N'N'-Tetramethylethane-1,2-diamine (TEMED) | Sigma Aldrich | T9281-25ML | |
| NuPAGE MES SDS running buffer (20x) | Fisher Scientific | LSNP0002 | |
| NuPAGE Novex 4-12% Bis-Tris gel 1.0 mm 12-well | Life Technologies | NP0322BOX | |
| oligonucleotide (cage antisense) | IDT | N/A | TATAGTGAGTCGTATTAATTTG |
| oligonucleotide (cage sense) | IDT | N/A | TCAGTCACCTATCTGTTTCAAA TTAATACGACTCACTATA |
| oligonucleotide (malachite green aptamer antisense) | IDT | N/A | GGATCCATTCGTTACCTGGCT CTCGCCAGTCGGGATCCTATA GTGAGTCGTATTACAGTTCCAT TATCGCCGTAGTTGGTGTACT |
| oligonucleotide (malachite green aptamer sense) | IDT | N/A | TAATACGACTCACTATAGGATC CCGACTGGCGAGAGCCAGGT AACGAATGGATCC |
| oligonucleotide (Transcription Factor A) | IDT | N/A | AGTACACCAACTACGAGTGAG |
| oligonucleotide (Transcription Factor B) | IDT | N/A | TCAGTCACCTATCTGGCGATAA TGGAACTG |
| oligonucleotide with 3’ Amine modification (tether) | IDT | N/A | GCTACTCACTCAGATAGGTGAC TGA/3AmMO/ |
| Pierce strong ion exchange spin columns | Fisher Scientific | 90008 | |
| plasmid encoding SNAP T7 RNAP and kanamycin resistance genes | Genscript | N/A | custom gene insert |
| protein purification column (HisPur Ni-NTA spin column) | Fisher Scientific | 88226 | |
| rNTP mix | New England Biolabs | N0466S | |
| Roche mini quick DNA spin column | Sigma Aldrich | 11814419001 | |
| Triton X-100 | Sigma Aldrich | T8787-100ML | |
| Ultra Low Range DNA ladder | Fisher Scientific | 10597012 | |
| urea | BioShop | URE001.1 |
参考文献
- Cherry, K. M., Qian, L. Scaling up molecular pattern recognition with DNA-based winner-take-all neural networks. Nature. 559 (7714), 370-376 (2018).
- Qian, L., Winfree, E., Bruck, J. Neural network computation with DNA strand displacement cascades. Nature. 475 (7356), 368-372 (2011).
- Chen, Y. -. J., et al. Programmable chemical controllers made from DNA. Nature Nanotechnology. 8 (10), 755-762 (2013).
- di Bernardo, D., Marucci, L., Menolascina, F., Siciliano, V. Predicting synthetic gene networks. Synthetic Gene Networks: Methods and Protocols. 813, 57-81 (2012).
- Xiang, Y., Dalchau, N., Wang, B. Scaling up genetic circuit design for cellular computing: advances and prospects. Natural Computing. 17 (4), 833-853 (2018).
- Gould, N., Hendy, O., Papamichail, D. Computational tools and algorithms for designing customized synthetic genes. Frontiers in Bioengineering and Biotechnology. 2, (2014).
- MacDonald, J. T., Siciliano, V. Computational sequence design with R2oDNA Designer. Mammalian Synthetic Promoters. 1651, 249-262 (2017).
- Cervantes-Salido, V. M., Jaime, O., Brizuela, C. A., Martínez-Pérez, I. M. Improving the design of sequences for DNA computing: A multiobjective evolutionary approach. Applied Soft Computing. 13 (12), 4594-4607 (2013).
- Zadeh, J. N., et al. NUPACK: Analysis and design of nucleic acid systems. Journal of Computational Chemistry. 32 (1), 170-173 (2011).
- Fornace, M. E., Porubsky, N. J., Pierce, N. A. A unified dynamic programming framework for the analysis of interacting nucleic acid strands: enhanced models, scalability, and speed. ACS Synthetic Biology. 9 (10), 2665-2678 (2020).
- Wetterstrand, K. DNA sequencing costs: Data. Genome.gov. , (2020).
- Lopez, R., Wang, R., Seelig, G. A molecular multi-gene classifier for disease diagnostics. Nature Chemistry. 10 (7), 746-754 (2018).
- Pardee, K., et al. low-cost detection of Zika virus using programmable biomolecular components. Cell. 165 (5), 1255-1266 (2016).
- Yurke, B., Turberfield, A. J., Mills, A. P., Simmel, F. C., Neumann, J. L. A DNA-fuelled molecular machine made of DNA. Nature. 406 (6796), 605-608 (2000).
- Lin, K. N., Volkel, K., Tuck, J. M., Keung, A. J. Dynamic and scalable DNA-based information storage. Nature Communications. 11 (1), 2981 (2020).
- Yurke, B., Mills, A. P. Using DNA to power nanostructures. Genetic Programming and Evolvable Machines. 4 (2), 111-122 (2003).
- Zhang, D. Y., Turberfield, A. J., Yurke, B., Winfree, E. Engineering entropy-driven reactions and networks catalyzed by DNA. Science. 318 (5853), 1121-1125 (2007).
- Wang, B., Thachuk, C., Ellington, A. D., Winfree, E., Soloveichik, D. Effective design principles for leakless strand displacement systems. Proceedings of the National Academy of Sciences. 115 (52), 12182-12191 (2018).
- Machinek, R. R. F., Ouldridge, T. E., Haley, N. E. C., Bath, J., Turberfield, A. J. Programmable energy landscapes for kinetic control of DNA strand displacement. Nature Communications. 5 (1), 5324 (2014).
- Cabello-Garcia, J., Bae, W., Stan, G. -. B. V., Ouldridge, T. E. Handhold-mediated strand displacement: a nucleic acid-based mechanism for generating far-from-equilibrium assemblies through templated reactions. bioRxiv. , (2020).
- Brophy, J. A. N., Voigt, C. A. Principles of genetic circuit design. Nature Methods. 11 (5), 508-520 (2014).
- Khalil, A. S., et al. A synthetic biology framework for programming eukaryotic transcription functions. Cell. 150 (3), 647-658 (2012).
- Swank, Z., Laohakunakorn, N., Maerkl, S. J. Cell-free gene-regulatory network engineering with synthetic transcription factors. Proceedings of the National Academy of Sciences. 116 (13), 5892-5901 (2019).
- Howland, S. W., Tsuji, T., Gnjatic, S., Ritter, G., Old, L. J., Wittrup, K. D. Inducing efficient cross-priming using antigen-coated yeast particles. Journal of immunotherapy. 31 (7), 607 (2008).
- Abil, Z., Ellefson, J. W., Gollihar, J. D., Watkins, E., Ellington, A. D. Compartmentalized partnered replication for the directed evolution of genetic parts and circuits. Nature Protocols. 12 (12), 2493-2512 (2017).
- Baugh, C., Grate, D., Wilson, C., Doudna, J. A. 2.8 Å crystal structure of the malachite green aptamer11. Journal of Molecular Biology. 301 (1), 117-128 (2000).
- Chou, L. Y. T., Shih, W. M. In vitro transcriptional regulation via nucleic acid-based transcription factors. ACS Synthetic Biology. 8 (11), 2558-2565 (2019).
- Lykke-Andersen, J., Christiansen, J. The C-terminal carboxy group of T7 RNA polymerase ensures efficient magnesium ion-dependent catalysis. Nucleic Acids Research. 26 (24), 5630-5635 (1998).
- Pu, J., Disare, M., Dickinson, B. C. Evolution of C-terminal modification tolerance in full-length and split T7 RNA Polymerase biosensors. Chembiochem. 20 (12), 1547-1553 (2019).
- Gardner, L. P., Mookhtiar, K. A., Coleman, J. E. Initiation, elongation, and processivity of carboxyl-terminal mutants of T7 RNA polymerase. Biochemistry. 36 (10), 2908-2918 (1997).
- Yin, J., Lin, A. J., Golan, D. E., Walsh, C. T. Site-specific protein labeling by Sfp phosphopantetheinyl transferase. Nature Protocols. 1 (1), 280-285 (2006).
- Warden-Rothman, R., Caturegli, I., Popik, V., Tsourkas, A. Sortase-tag expressed protein ligation: combining protein purification and site-specific bioconjugation into a single step. Analytical Chemistry. 85 (22), 11090-11097 (2013).
- Zhang, W. -. B., Sun, F., Tirrell, D. A., Arnold, F. H. Controlling macromolecular topology with genetically encoded SpyTag-SpyCatcher chemistry. Journal of the American Chemical Society. 135 (37), 13988-13997 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。