
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
在体内成像完全活跃的脑组织在清醒斑马鱼幼虫和青少年由骷髅和皮肤去除
摘要
在这里,我们提出了一种方法,以图像斑马鱼胚胎大脑在体内到幼虫和幼年阶段。这种微创程序,根据电生理学方法改编,提供成熟神经元的细胞和亚细胞细节,并可与光遗传学和神经药理学研究相结合,以描述大脑功能和药物干预。
摘要
了解大脑发育和成熟过程中发生的短暂变化需要在空间和时间上以细胞和亚细胞分辨率进行详细的高分辨率成像。分子和成像技术的进步使我们能够对透明斑马鱼胚胎中大脑发育的细胞和分子机制获得许多详细的见解。最近,在受精几周后的幼虫阶段发生的神经元连接的细化过程,例如控制社会行为、决策或动机驱动行为,已经进入研究重点。在这些阶段,斑马鱼皮肤的色素沉着会干扰光线渗透到脑组织,胚胎阶段的解决方案,例如,药理抑制色素沉着,已经不可行了。
因此,提供微创手术解决方案,用于通过微观镜进入清醒斑马鱼的大脑,该解决方案源自电生理方法。在电传中,皮肤和柔软的头骨软骨可以通过微皮这些层,暴露潜在的神经元和轴向而不损坏仔细去除。这允许记录神经元形态,包括突触结构及其分子含量,以及观察生理变化,如Ca2+ 瞬态或细胞内传输事件。此外,通过药理抑制或光遗传学操作来审讯这些过程是可行的。这种大脑暴露方法提供有关神经元结构和生理变化的信息,以及这些事件在几分钟或几小时内在活脑组织中的相关性和相互依存性。该技术适用于斑马鱼幼虫在受精后长达30天的体内脑成像,这是迄今为止测试的最新发育阶段。因此,它提供了诸如突触细化和缩放、轴突和树突运输、细胞骨骼货物突触靶向或当地活动依赖表达等重要问题。因此,可以预见这种安装和成像方法的广泛应用。
引言
近几十年来,斑马鱼(Danio rerio)已发展成为胚胎和幼虫发育研究中最受欢迎的脊椎动物模型生物之一。斑马鱼雌性的巨大雌性,加上胚胎 的快速前子宫 发育及其在早期胚胎发育阶段的透明度,只是使斑马鱼成为解决发育问题的强大模型有机体的几个关键因素。分子遗传技术的进步与体内成像研究 的高 分辨率相结合,使得解决细胞生物学机制基础发育过程2。特别是在神经元分化、生理学、连通性和功能领域,斑马鱼以前所未有的细节揭示了分子动力学、大脑功能和有机体行为的相互作用。
然而,由于神经系统组织的透明度逐渐丧失,这些研究大多局限于发育第一周的胚胎和早期幼虫阶段。在这些阶段,脑组织被高分辨率显微镜方法阻止进入,从而被头骨分化和色素沉着3所屏蔽。
因此,神经元分化、成熟度和可塑性等关键问题,如神经元连接性或突触缩放的细化,都难以研究。这些细胞过程对于定义细胞机制驱动非常重要,例如,社会行为、决策或基于动机的行为,斑马鱼对数周幼虫的研究最近根据行为研究贡献了关键发现。
药理学方法抑制斑马鱼幼虫色素的色素消耗几个星期是几乎不可行,甚至可能造成有害影响5,6,7,8。具有特定色素沉着缺陷的双突变株(如casper9或晶体10)已成为极其有价值的工具,但在育种方面非常费力,提供很少的后代,并且由于近亲繁殖过度而造成遗传畸形的积累危险。
在这里,提供最小侵入性程序作为替代,适用于任何斑马鱼菌株。这个程序是从电生理学研究中改编而来,记录活的和清醒的斑马鱼幼虫的神经元活动。在电传中,皮肤和柔软的头骨软骨可以通过微皮仔细去除这些层,因为它们没有与大脑血管紧密交织。这允许暴露含有神经元和轴突的脑组织而不造成损伤,并记录神经元形态,包括突触结构及其分子含量,这反过来又包括观察生理变化,如Ca2+ 瞬态或细胞内传输事件长达数小时。此外,除了描述性特征外,直接接触脑组织还可以通过神经药物管理和光遗传学方法对成熟的神经元功能进行审讯。因此,使用这种大脑暴露策略,可以在幼斑马脑中揭示出真正的结构功能关系。
研究方案
此处描述的所有动物工作均符合法律规定(欧盟指令 2010/63)。养护和处理鱼类已得到地方当局和布劳恩施魏格技术大学动物福利代表的批准。
1. 准备人工脑脊液(ACSF)、低熔化藻和锋利的玻璃针
- 通过溶解蒸馏水中的以下浓度溶解所列化学品,准备 ACSF。134 m M NaCl (58.44 克/摩尔), 2.9 mM KCl (74.55 克/摩尔), 2.1 m CaCl2 (110.99 克/摩尔), 1.2 mM MgCl2 6x H2O (203.3 克/摩尔)、10 mM HEPES (238.31 克/摩尔) 和 10 mM d-葡萄糖(180.16 克/摩尔)。
注:对于MgCl2、CaCl2和 KCl,100 万库存溶液在脱盐无菌水中准备,并储存在 4 °C 下,以便随后准备新的 ACSF。葡萄糖、HEPES 和 NaCl 作为固体化合物溶解在新鲜的 ACSF 溶液中。对于溶解化学品,请按照制造商的说明操作。 - 将 ACSF 的 pH 值调整为 7.8,配备 10 M NaOH。ACSF的制备需要精确测量化学物质和pH值的微调,因为它取代了脑脊髓液,并维持神经元完全正常运作所需的生理条件,否则可能导致大脑功能失调和神经元死亡。
- 将新准备的 ACSF 存储在 4 °C 下,最长 4 周。对于工作条件,在 25-28 °C 下,将当天/实验和预温所需的 ACSF 体积(并可选地将其氧化,步骤 2.5)
注:新准备的ASCF可以 1 天。如果计划使用几天,ACSF 需要无菌过滤。 - 对于幼虫的麻醉,在蒸馏水中准备 50 mM 的 d-Tubocurarine 库存溶液,并将溶液存储在 -20 °C 作为 100 μL 的阿利奎特在冰箱中,直到需要为止。
- 要嵌入鱼,准备2.5%低熔化(LM)的玫瑰溶解1.25克LM-阿加罗斯(材料表)在50ml ACSF和煮沸,直到阿加罗斯完全溶解。
注意:或者,根据实验设置,可以使用更高或更低浓度的LM-agarose。然而,如果玫瑰太软,它将无法举行鱼的位置时,打开头骨。 - 将藻子储存在37°C的水浴中,以避免凝固,因为这种温度也不会在嵌入时伤害幼虫。在水浴中将煮沸的阿加罗斯冷却到 37 °C 后,在当天所需的藻类中加入必要的 d-Tubocurarine,以达到 10 μM 的工作浓度。供将来使用时,将剩余的藻子储存在 4 °C 处,以避免污染。
- 使用具有以下设置的微管拉拔器,从玻璃毛细血管中准备锋利而薄的玻璃针(补充图1)。
- 拉力I,毛细电类型1:热1:65.8:热 2: 55.1:2 步拉力
拉拔器 II, 毛细电类型 2: 热 + 700;菲尔 = 4;维尔 = 55;德尔 = 130;脉冲 = 55;1 步拉力。
注:这些装置分别用于此处使用的每个拉拔器和玻璃毛细圈(参见 材料表)。其他毛细血管和拉拔器也可用于准备玻璃针。但是玻璃针不应该太薄,因为它们在与头骨接触时可能会断裂。毛细电:长度:100毫米(4英寸):OD: 1.5 毫米;ID: 0.84 毫米:灯丝:是的
- 拉力I,毛细电类型1:热1:65.8:热 2: 55.1:2 步拉力
2. 幼虫麻醉和嵌入准备
- 当开始当天的实验时,将用塑料巴斯德移液器需要的动物转移到直径为 90 mm 的培养皿中,里面装满了 Danieau(对于仍然与 Danieau 一起保存在培养皿中的幼虫)或来自鱼类设施的水(对于年龄超过 7 dpf 且保存在鱼类设施中的幼虫)。
- 当管道鱼超过2周时,确保移液器的开口足够大,以避免在转移鱼时伤害它们。不要使用网,因为它会身体伤害,特别是年轻的幼虫。
- 加入适合培养皿中幼虫大小的 罗蒂费拉 或 艺术性纳 普利,以确保幼虫免费获得食物和最大健康状况,并减轻压力。
- 要嵌入,将选定的幼虫转移到一个直径为 35 mm 的培养皿中,该盘中充满了 ACSF。加入必要的量d-Tubocurarine,以达到工作浓度/有效剂量10μM,并等待几分钟,直到幼虫完全固定11。
注:当鱼变老或需要更快的全身麻醉(5分钟以下),有可能增加d-Tubocurarine的浓度(小鼠的LD50是0.13毫克/千克静脉注射12)。也可以使用不同的麻醉剂,如α-bungarotoxin(工作浓度:1毫克/毫升),它具有与库拉雷相同的效果,也保持大脑充分活跃13。如果一个完全活跃的大脑是没有必要的兴趣对象,特里卡因在非致命剂量(0.02%)也是一个选项,完全麻醉幼虫。然而,三卡因阻断钠通道,从而损害大脑活动14。 - 将直径为 35 mm 的 Petri 盘盖上盖子,倒置盖子,并将方形玻璃盖片(24 x 24 mm)放在盖子底部,从而准备安装室。有关这些步骤的示意图描述,请参阅 图 1( 上半部分)。玻璃表面更光滑,可防止从头骨开启过程中含有幼虫的玫瑰块中滑落。
- 在适当的小瓶(例如,50 mL 管、烧杯、肖特瓶等)中,将当天所需的 ACSF 量进行氧化,并将其与卡博根(5% CO 2、95% O2)进行氧合。如果成像只是形态学(例如荧光模式)ACSF仍然是必要的,以确保大脑的完整性,并确保细胞不受渗透效应的负面影响,但不需要ACSF的氧合。此步骤仅在成像所需的全脑活动时执行。
注:为了最佳的介质氧饱和度,在卡博根管的末端添加一块空气石。为了保证足够高的氧气水平,有必要每20-60分钟在成像室中用新鲜含氧的ACSF交换一次ACSF,这取决于嵌入在同一成像室中的幼虫的数量和年龄(例如,每小时交换一个嵌入式幼虫ACSF就足够了。对于平行嵌入的6只14分以上的幼虫,每20分钟交换一次ACSF是必要的),因此根据计划的实验计划必要的氧饱和ACSF量。
3. 幼虫的嵌入
- 将完全麻醉的幼虫用塑料巴斯德移液器转移到(第 2.4 步)准备好的安装室。然后,小心地去除多余的介质,以避免 LM-agarose 稀释。所有以下步骤应在具有足够放大的立体显微镜下执行。
注意:倾斜安装室有助于完全移除介质。 - 立即进入下一步,在幼虫(约1mL,取决于幼虫的大小)上添加足够大的LM-agarose下降,以保护动物不干涸,并减少不必要的压力。
- 在阿加罗斯凝固之前,将幼虫定向在位置。确保幼虫的正点部分向上定向。此外,请务必尽可能靠近幼虫表面嵌入幼虫。
注:根据计划同时嵌入的幼虫的大小和数量,可以调整藻糖浓度。例如,对于 1-3 幼虫是 30 dpf 老, 建议浓度 1.8%-2% LM-阿加罗斯.对于 1-4 幼虫是 7 dpf 老, 这是最可行的使用 2.5% LM - 阿加罗斯, 而, 对于 5-8 幼虫, 2% 更适合.如果需要一个完全活跃的大脑,建议只嵌入三条鱼在同一时间,以减少所需的时间操作幼虫。一般来说,建议使用浓度较低的(1.8%-2%),幼虫越老,或计划同时嵌入更多的幼虫。 - 如果图像将使用倒置显微镜记录,将含有幼虫的阿加罗斯块修剪成一个小的幼崽形状。这对于以后将幼虫转移到成像室非常重要。如果使用直立显微镜,则不需要进行这种修剪,因为安装室也可以用作成像室。在 图 1( 上半部分)中,可以找到这些步骤的示意图描述。
4. 暴露大脑
注意:应非常小心地执行以下所有步骤,不要不必要地伤害幼虫。如果实验需要一个完全活跃的大脑,请记住,每过一秒,鱼仍然完全安装在阿加罗斯,有一个开放的头骨没有氧化ACSF,大脑将遭受缺氧,也干涸。缺氧的影响将变得更加戏剧化,嵌入幼虫越老。因此,重要的是不仅在尽可能短的时间内进行手术,而且要以最大的精度,不要引起机械脑损伤与针头。训练时,步骤 4.2-4.4 每条鱼不应超过 30 秒。
- 一旦玫瑰凝固,就开始手术。首先,修剪掉大脑感兴趣的区域上方的所有多余的气喘,以获得头部和清晰的工作空间。如果头部的正点部分已经伸出了腹膜,请跳过此步骤。
- 根据感兴趣的区域,选择一个点开始手术。拿玻璃针,在皮肤上做一个小切口,但不会穿透组织太深。这将是剥去覆盖皮肤的起点。
注:为了获得最佳结果,切勿直接从感兴趣区域上方开始,以降低损坏重要结构的风险。如有必要,甚至可以开始后脑后脑,并从那里向前工作,直到皮肤的不需要的区域被剥去。 - 继续沿着皮肤的一部分非常小的切口,旨在通过在表面下方几乎移动针头来去除。大多数时候,没有必要完全围绕大脑移动,切出一块圆形的皮肤和头骨,而只需沿着头部做两个切口,然后把皮肤推到一边或另一边。 图2 显示了获得小脑自由访问的优化切割策略的示意图。
注:这种微手术是一个微妙的过程,它很可能需要一些培训,以完美地去除皮肤,而不会损害潜在的大脑。建议找出大脑感兴趣区域的最佳切割策略,并在实验期间坚持使用。 - 立即从所有嵌入的幼虫中去除皮肤后,立即将(含氧的)ACSF倒在玫瑰上,以冲走不需要的皮肤颗粒和血液,保持大脑充分活跃,并保护大脑不干涸。
注意:如果实验需要一个健康的大脑,建议每次最多吃三条鱼。 - 如果使用直立显微镜,则直接从成像开始。
- 使用倒置显微镜时,将一个小铲子滑到库博伊德玫瑰块(第 3.4 步)下面。
- 在成像室底部(例如玻璃底盘)上加入一小滴LM-agarose,然后立即用铲子翻转含有幼虫的阿加罗斯块180°,轻轻将其推到成像室底部,而液体气垫则充当胶水。
- 当气融凝固后,将成像室装满(含氧的)ACSF,然后开始成像。有关示意图描述,请参阅 图 1( 下半部分)。
- 当实验需要全脑活动时,始终确保成像室中的 ACSF 具有足够高的氧气水平。为了确保这一点,每20-60分钟(取决于鱼的数量和大小,成像室的大小和表面,以及成像持续时间),尽可能小心地与新鲜含氧的ACSF交换介质。

图1:为开放头骨斑马鱼的分步成像准备的示意图程序。不同步骤的工作说明可在图形本身中找到。图形由弗洛里安·赫奇设计,保罗·施拉姆改编。请单击此处查看此图的较大版本。
{kind=link}

图2:为去除大脑区域上方的皮肤和头骨碎片而进行的微手术的详细示意图。 红色圆圈标记了需要进行第一次切割的位置。红点线划出最佳路径,与针头一起切割,以获得免费进入小脑,而不会损坏它。绿色箭头标记了过度的皮肤和头骨碎片很容易被推开的方向。在整个过程中,确保不要渗透到脑组织中。成功剥去皮肤后,感兴趣的大脑区域(这里,小脑)将可自由访问任何类型的高分辨率 活体 成像。 请单击此处查看此图的较大版本。
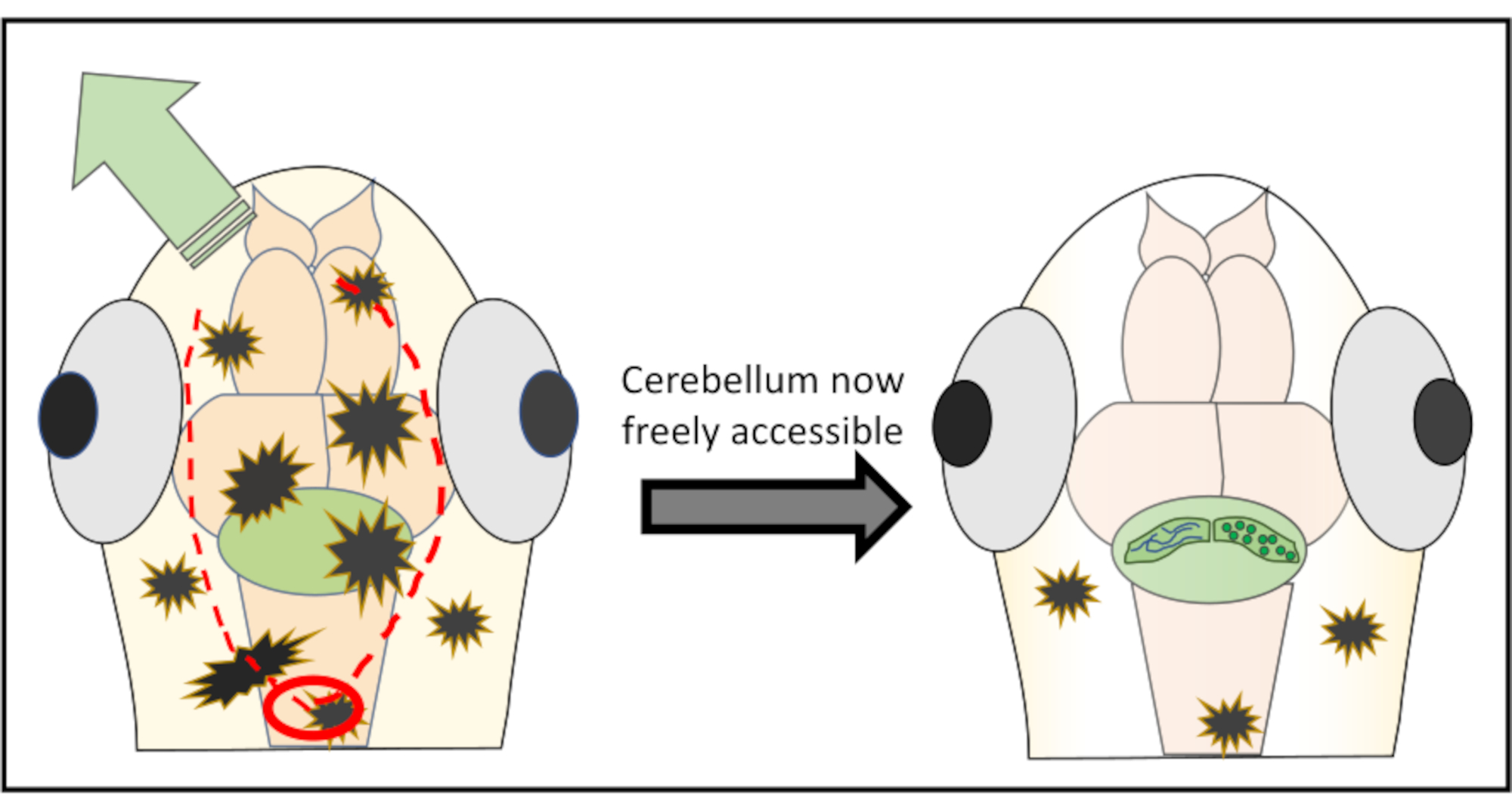{kind=link}
结果
图3A, C 显示转基因线 Tg [-7.5Ca8:GFP] 的 14 dpf 幼虫bz12]15] 头骨仍然完好无损覆盖皮肤中的色素细胞分布在头部,并干扰感兴趣的区域(这里,小脑)的荧光信号。在这种情况下,幼虫不可能获得大脑的高分辨率图像。 图 3B 在进行开放性头骨手术后,显示同一转基因线的幼虫。现在?...
讨论
提出的方法提供了一种替代方法,大脑隔离或斑马鱼幼虫的治疗与药物抑制色素干扰记录高分辨率图像的神经元在其 体内 环境。使用这种方法记录的图像质量可与从外生大脑中拍摄的图像相媲美,但在自然条件下。
此外,避免了荧光强度的损失,因为没有必要用固定剂18治疗。此外,这种方法不像隔离大脑那样具有侵入性,它只包括可能导致失败的?...
披露声明
作者没有什么可透露的。
致谢
我们特别感谢蒂莫·弗里奇出色的动物护理,赫尔曼·德林、穆罕默德·艾尔赛、索尔·波塞-门德斯、雅各布·冯·特罗塔、科马利·瓦利谢蒂和芭芭拉·温特给予我们帮助的支持。我们还感谢科斯特实验室的所有其他成员的反馈。该项目部分由德国研究基金会(DFG,KO1949/7-2)项目241961032(到RWK)和德国联邦委员会(BMBF)资助。时代网神经NII CIPRESS项目01EW1520到JCM)得到认可。
材料
| Name | Company | Catalog Number | Comments |
| Calcium chloride | Roth | A119.1 | |
| Confocal Laser scanning microscope | Leica | TCS SP8 | |
| d-Glucose | Sigma | G8270-1KG | |
| d-Tubocurare | Sigma-Aldrich | T2379-100MG | |
| Glass Capillary type 1 | WPI | 1B150F-4 | |
| Glass Capillary type 2 | Harvard Apparatus | GC100F-10 | |
| Glass Coverslip | deltalab | D102424 | |
| HEPES | Roth | 9105.4 | |
| Hoechst 33342 | Invitrogen (Thermo Fischer) | H3570 | |
| Imaging chamber | Ibidi | 81156 | |
| Potassium chloride | Normapur | 26764298 | |
| LM-Agarose | Condalab | 8050.55 | |
| Magnesium chloride (Hexahydrate) | Roth | A537.4 | |
| Microscope Camera | Leica | DFC9000 GTC | |
| Needle-Puller type 1 | NARISHIGE | Model PC-10 | |
| Needle-Puller type 2 | Sutter Instruments | Model P-2000 | |
| Pasteur-Pipettes 3ml | A.Hartenstein | 20170718 | |
| Sodium chloride | Roth | P029.2 | |
| Sodium hydroxide | Normapur | 28244262 | |
| Tricain | Sigma-Aldrich | E10521-50G | |
| Waterbath | Phoenix Instrument | WB-12 | |
| 35 mm petri dish | Sarstedt | 833900 | |
| 90 mm petri dish | Sarstedt | 821473001 |
参考文献
- Embryology. Zebrafish Development Available from: https://embryology.med.unsw.edu.au/embryology/index.php/Zebrafish_Development (2020)
- Sassen, W. A., Köster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. Dove Medical Press. 2015 (5), 151-163 (2015).
- Singh, A. P., Nüsslein-Volhard, C. Zebrafish stripes as a model for vertebrate colour pattern formation. Current Biology. 25 (2), 81-92 (2015).
- Kalueff, A. V., et al. Time to recognize zebrafish 'affective' behavior. Brill: Behaviour. 149 (10-12), 1019-1036 (2012).
- Karlsson, J., von Hofsten, J., Olsson, P. -. E. Generating transparent zebrafish: a refined method to improve detection of gene expression during embryonic development. Marine Biotechnology. 3, 522-527 (2001).
- Bohnsack, B. L., Gallina, D., Kahana, A. Phenothiourea sensitizes zebrafish cranial neural crest and extraocular muscle development to changes in retinoic acid and IGF signaling. PloS One. 6, 22991 (2011).
- Elsalini, O. A., Rohr, K. B. Phenylthiourea disrupts thyroid function in developing zebrafish. Development Genes and Evolution. 212, 593-598 (2003).
- Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., Segner, H. Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology. 172, 44-55 (2016).
- White, R., et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell. 2, 183-189 (2008).
- Antinucci, P., Hindges, R. A crystal-clear zebrafish for in vivo imaging. Scientific Reports. 6, 29490 (2016).
- Burr, S. A., Leung, Y. L. Curare (d-Tubocurarine). Encyclopedia of Toxicology (3rd Edition). , 1088-1089 (2014).
- Gesler, H. M., Hoppe, J. 3,6-bis(3-diethylaminopropoxy) pyridazine bismethiodide, a long-acting neuromuscular blocking agent. The Journal of Pharmacology and Experimental Therapeutics. 118 (4), 395-406 (1956).
- Furman, B. . Alpha Bungarotxin. Reference Module in Biomedical Sciences. , (2018).
- Attili, S., Hughes, S. M. Anaesthetic tricaine acts preferentially on neural voltage-gated sodium channels and fails to block directly evoked muscle contraction. PLoS One. 9 (8), 103751 (2014).
- Namikawa, K., et al. Modeling neurodegenerative spinocerebellar ataxia type 13 in zebrafish using a Purkinje neuron specific tunable coexpression system. Journal of Neuroscience. 39 (20), 3948-3969 (2019).
- Hennig, M. Theoretical models of synaptic short term plasticity. Frontiers in Computational Neuroscience. 7 (45), (2013).
- Wang, Y., et al. Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis. Development. 137, 3119-3128 (2010).
- Hobro, A., Smith, N. An evaluation of fixation methods: Spatial and compositional cellular changes observed by Raman imaging. Vibrational Spectroscopy. 91, 31-45 (2017).
- Knogler, L. D., Kist, A. M., Portugues, R. Motor context dominates output from purkinje cell functional regions during reflexive visuomotor behaviours. eLife. 8, 42138 (2019).
- Hsieh, J., Ulrich, B., Issa, F. A., Wan, J., Papazian, D. M. Rapid development of Purkinje cell excitability, functional cerebellar circuit, and afferent sensory input to cerebellum in zebrafish. Frontier in Neural Circuits. 8 (147), (2014).
- Scalise, K., Shimizu, T., Hibi, M., Sawtell, N. B. Responses of cerebellar Purkinje cells during fictive optomotor behavior in larval zebrafish. Journal of Neurophysiology. 116 (5), 2067-2080 (2016).
- Harmon, T. C., Magaram, U., McLean, D. L., Raman, I. M. Distinct responses of Purkinje neurons and roles of simple spikes during associative motor learning in larval zebrafish. eLife. 6, 22537 (2017).
- Zehendner, C. M., et al. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS One. 8 (12), 82823 (2013).
- Dhabhar, F. S. The short-term stress response - mother nature's mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Frontiers of Neuroendocrinology. 49, 175-192 (2018).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。