
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
从猪肾和小鼠肝脏中提取和纯化FAHD1蛋白
摘要
该协议描述了如何从猪肾和小鼠肝脏中提取富马酰乙酰乙酸水解酶结构域蛋白1(FAHD1)。所列的方法可以适应于其他感兴趣的蛋白质并针对其他组织进行修饰。
摘要
富马酰乙酰乙酸水解酶结构域含蛋白1(FAHD1)是真核生物中FAH超家族的第一个成员,在线粒体中充当草酰乙酸脱羧酶。本文介绍了一系列从猪肾和小鼠肝脏中提取和纯化FAHD1的方法。涵盖的方法包括使用快速蛋白质液相色谱 (FPLC) 进行离子交换色谱,使用 FPLC 进行制备和分析凝胶过滤,以及蛋白质组学方法。全蛋白提取后,探讨硫酸铵沉淀和离子交换色谱, 采用 离子交换和尺寸排阻色谱法提取FAHD1。这种具有代表性的方法可以适应其他感兴趣的蛋白质(在显着水平上表达)并针对其他组织进行修饰。从组织中纯化的蛋白质可以支持高质量抗体和/或有效和特异性药理抑制剂的开发。
引言
真核FAH结构域蛋白1(FAHD1)作为双功能草酰乙酸(OAA)脱羧酶(ODx)1和酰基丙酮酸水解酶(ApH)2。它定位于线粒体2中,属于酶1,2,3,4,5,6的广泛FAH超家族。虽然其ApH活性仅具有次要相关性,但FAHD1的ODx活性参与TCA循环通量1,7,8,9的调节。OAA不仅是三羧酸循环中柠檬酸盐合酶反应所必需的,而且还作为琥珀酸脱氢酶的竞争性抑制剂,作为电子传递系统的一部分和作为弹力代谢物。人脐静脉内皮细胞(HUVEC)中FAHD1基因表达的下调导致细胞增殖速率10的显着降低,线粒体膜电位的显着抑制,与同时切换到糖酵解有关。工作模型是指线粒体功能障碍相关衰老(MiDAS)11样表型8,其中线粒体OAA水平受到FAHD1活性1,8,9的严格调节。
重组蛋白更容易 通过 细菌12 的表达和纯化而不是从组织中获得。然而,在细菌中表达的蛋白质可能由于缺乏翻译后修饰而偏倚,或者可能只是有问题(即,由于质粒丢失,细菌应激反应,扭曲/未形成的二硫键,无或分泌不良,蛋白质聚集,蛋白水解裂解等)。对于某些应用,蛋白质需要从细胞裂解物或组织中获得,以便包括这种修饰和/或排除可能的伪影。从组织中纯化的蛋白质支持高质量抗体的开发,和/或针对选定酶的有效和特异性药理抑制剂,例如FAHD113。
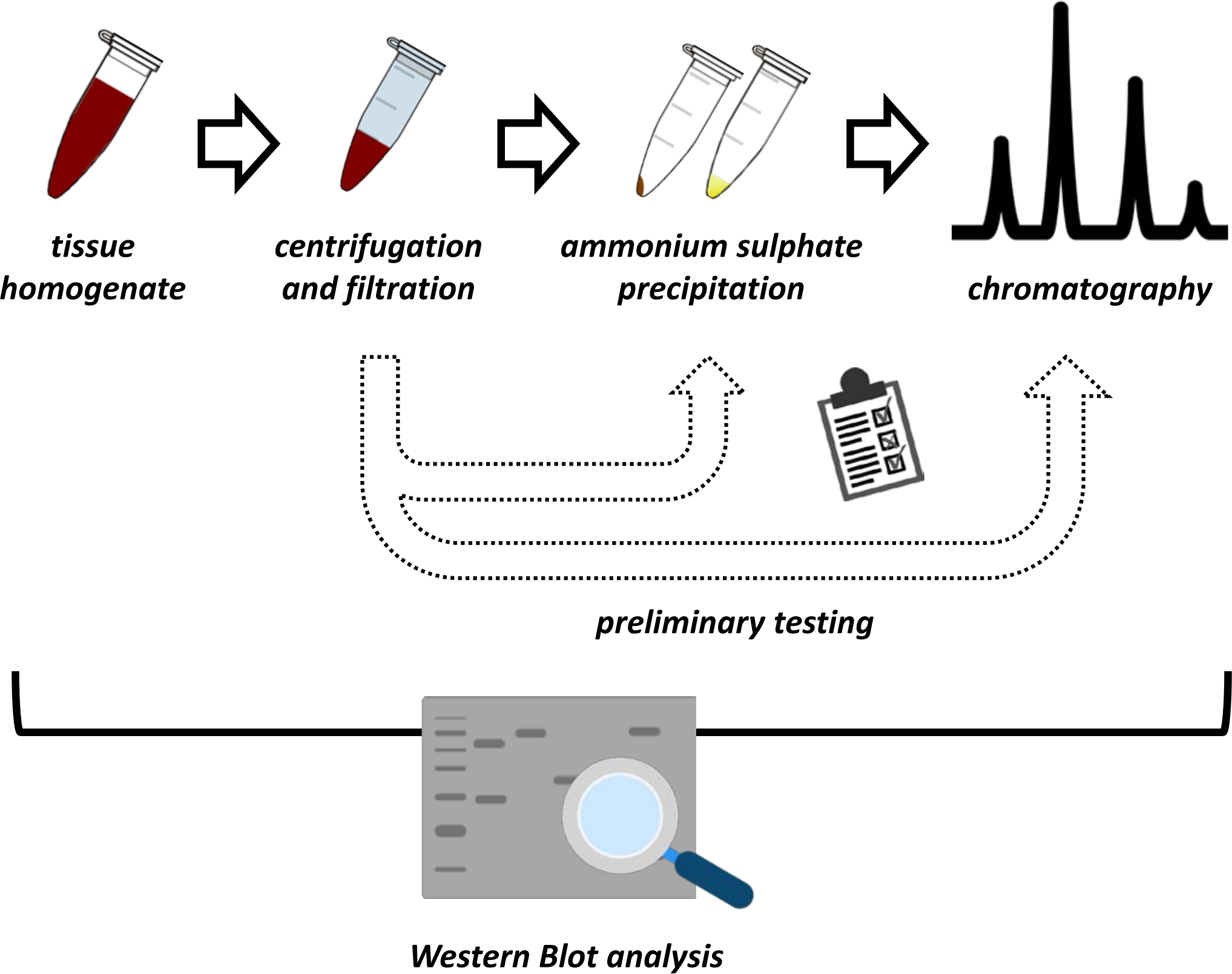
本手稿介绍了从猪肾和小鼠肝脏中提取和纯化FAHD1的一系列方法。所描述的方法需要快速蛋白质液相色谱(FPLC),但使用常见的实验室设备。替代方法可以在其他地方找到14,15,16,17。在全蛋白提取后,所提出的方案涉及一个测试阶段,其中讨论了硫酸铵沉淀和离子交换色谱的子方案(图1)。在定义了这些亚方案之后,通过使用离子交换和FPLC的尺寸排阻色谱法, 通过 顺序策略提取感兴趣的蛋白质。基于这些指南,最终方案可以单独适应其他感兴趣的蛋白质。

图 1:该协议的总体策略。从上到下:蛋白质是从组织中提取的。制备组织匀浆,离心和过滤。对于每对上清液和颗粒衍生样品,需要进行硫酸铵沉淀和离子交换色谱(FPLC)的测试以探针最佳条件。建立这些子方案后,可以通过硫酸铵沉淀,离子交换色谱和重复大小排阻色谱(FPLC)在不同pH和盐浓度下的顺序程序提取蛋白质。所有步骤都需要通过免疫印迹控制。请点击此处查看此图的大图。
{kind=link}
研究方案
所有实验均按照机构指南进行。猪肾是从当地超市新鲜获得的。从因斯布鲁克大学生物医学衰老研究所,Rennweg 10,6020奥地利因斯布鲁克大学生物医学衰老研究所保存的C57BL6野生型小鼠中收获肝脏组织,在Univ.-Doz的监督下。Pidder Jansen-Dürr 博士,2013 年获得项目负责人的道德许可(BMWF-66.008/0007-II/3b/2013)。该项目的小鼠的维护和使用由奥地利教育,科学和研究部(BMBWF)颁发的2020-0.242.978号道德许可No.2020-242.978涵盖。
1. 准备工作
注意:在方案开始之前,除了一般化学品和材料外,还需要准备几种东西,即蛋白质裂解缓冲液,粗组织样品和特定抗体。
- 每100g组织净重制备250mL蛋白质裂解缓冲液:250mL 1x PBS与50mM NaF,1mM PMSF,2μg/ mL抑肽酶和1mM活化的原钒酸盐(见 表1)。使用0.22μm注射器过滤单元过滤溶液。
注意:在使用前需要活化原钒酸盐,以将其转化为更有效的蛋白质酪氨酸磷酸酶18抑制剂。活化的原钒酸盐可以从商业供应商处获得,但也制备如下。- 在ddH 2O中制备200mM原钒酸钠(钠)储备溶液。为了制备10mL溶液,加入368mg Na3VO4 至9mL水,搅拌溶解。溶解后,用ddH 2 O将体积增至10mL。
注意:原钒酸钠溶液的起始pH值可能因材料来源而异,并且需要按如下重复方法将pH值调节至10。 - 根据溶液的初始pH值,用NaOH或HCl将pH值调节至10。在pH值>10时,溶液将具有黄色。将溶液煮沸直至无色,将其冷却至室温,并检查pH值。如果pH值>10,则加入少量HCl以将pH调节至10。此时,溶液可能会再次变黄。
- 重复煮沸和冷却,直到溶液保持无色,pH稳定在10(约5-7倍)。此时,加入HCl会导致溶液中淡淡的黄色外观。将活化的原钒酸盐储存在-20°C的1mL等分试样中。
- 在ddH 2O中制备200mM原钒酸钠(钠)储备溶液。为了制备10mL溶液,加入368mg Na3VO4 至9mL水,搅拌溶解。溶解后,用ddH 2 O将体积增至10mL。
- 每克组织用2 mL裂解缓冲液制备试管,并将其置于冰上。
注意:该方案使用八个50 mL管,每个管子总共填充30 mL裂解缓冲液,用于一个猪肾(约100-150g),两个管子每个管填充40mL裂解缓冲液,总共20个小鼠肝脏(每个1-2g)。 - 准备组织:在预先清洁的玻璃板上解剖组织,该玻璃板上放置在聚苯乙烯泡沫盒中的冰上。切割每个约100mg的组织片,以便于转移到各自的管中以进行随后的裂解。将组织片段转移到准备好的管中(步骤1.2)。
- 制备饱和硫酸铵溶液:将500mL ddH2O加热至70°C,搅拌的同时,逐渐加入硫酸铵粉末(见 材料表),直至不再溶解硫酸铵。将这种(过饱和)溶液冷却至室温,并将其储存在4°C过夜。
2. 总蛋白提取
注意:在冷蛋白质裂解缓冲液中制备样品后(参见步骤1.3), 通过 超声探针超声处理或使用电动均质器尽可能最好地匀浆组织,如下所示。
- 组织均质化
- 在猪肾的情况下,最好通过超声探头超声处理悬浮液,同时将样品保持在冰上(10个周期的15 s脉冲,脉冲之间的间隔为30 s,以在冰上冷却样品,中等幅度为50%占空比)。
- 在小鼠器官的情况下,使用电动均质机(从低力开始,缓慢加速到中等力)使悬浮液均质化,同时将样品保持在冰上。定期清洗PBS中的电均质机,以除去任何堵塞设备的有机物质。
- 从样品中取出20μL,并在显微镜下检查匀浆组织的细胞是否被适当破坏;否则,重复均质化。
- 在4°C下以10,000× g 的台式离心机中离心管30分钟。
注意:任选地,在4°C下以20,000× g 第二次离心上清液30分钟,以消除可能已转移的初始沉淀的一小部分。这将简化步骤2.3中的后续过滤。 - 将上清液收集在新鲜的管中并将其放在冰上。使用0.45μm和0.22μm注射器过滤单元按顺序过滤上清液。将上清液等分到10mL批次中,并在-20°C下冷冻以进行短期储存或在-80°C下冷冻以进行长期储存。
注意:在用0.22μm进行第二个过滤步骤去除更细小的颗粒之前,用0.45μm进行预过滤去除大部分颗粒。直接使用0.22 μm过滤器可能会导致过滤器堵塞的风险。 - 通过将10μL的5x SDS样品缓冲液(见 表1)加入40μL上清液,然后在95°C下沸腾10分钟,制备50μL样品用于SDS-PAGE/蛋白质印迹分析。
- 任选地,将步骤2.2中获得的约100μL沉淀重悬于900μLddH2O中,并如上所述制备用于SDS-PAGE/蛋白质印迹分析的样品。
注意:除了阳性对照外,在蛋白质印迹分析中纳入颗粒衍生样品将表明蛋白质的表达是否低,或者抗体有问题。
- 任选地,将步骤2.2中获得的约100μL沉淀重悬于900μLddH2O中,并如上所述制备用于SDS-PAGE/蛋白质印迹分析的样品。
3. SDS-PAGE和蛋白质印迹分析
注意:需要蛋白质印迹分析来检查蛋白质溶解度。下面描述了使用湿/罐印迹系统的电印迹方案(参见 材料表)。SDS-PAGE的替代协议可以在别处19找到。
- 根据制造商的说明制备不连续的12.5%聚丙烯酰胺SDS-PAGE凝胶(即,在分辨凝胶之上的堆叠凝胶;见 表1)。运行先前在步骤 2 中准备的样本(类似于步骤 4、5 和 6;见下文)。
- 将蛋白质标记分子量标准品架加载到第一个孔中(参见 材料表)。加载5ng的hFAHD1重组蛋白(从细菌12中获得;见 表1)作为阳性对照进入第二孔。
- 随后,加载20μL待分析样品,并用20μL制备的SDS-PAGE 1x样品缓冲液(即用ddh2O稀释的5x样品缓冲液)填充所有剩余的孔。使用SDS电泳缓冲液在125 V下电泳SDS-PAGE凝胶(见 表1)。
- SDS-PAGE完成后,进行蛋白质印迹分析,并使用针对FAHD1的可用抗体探测膜(见 表1)。
注意:由于样品是从粗组织匀浆中取出的,因此此时SDS-PAGE和蛋白质印迹分析的质量通常会受到影响;然而,重要的是要检查要提取的蛋白质是否可溶于上清液。以下方案针对猪肾和不同的小鼠器官(包括肝脏,心脏,大脑和肾脏)进行了测试。- 准备10x蛋白质印迹转印缓冲液(见 表1)。准备1x蛋白质印迹转印缓冲液(见 表1)并将其冷却至4°C。
- 在甲醇中激活PVDF膜2分钟。将膜在ddH2O中洗涤2分钟。在1x蛋白质印迹转印缓冲液中平衡膜15分钟。
- 用1x PBS洗涤SDS凝胶10分钟,同时摇动以除去SDS电泳缓冲液,然后将凝胶在1x蛋白质印迹转印缓冲液中孵育10分钟以进行平衡。根据制造商的说明组装电印迹盒(即,结合活化的PVDF膜和凝胶)。
- 在装满冰的聚苯乙烯泡沫盒中或在冷藏室(4°C)中 以 300 mA电印迹运行印迹1小时。将PVDF膜转移到50 mL管中,其暴露的一面朝向管的内侧。将膜在20mL蛋白质印迹封闭缓冲液(见 表1)中孵育4°C过夜,同时在管辊上滚动(参见 材料表)。
- 第二天,在滚动时用20mL蛋白质印迹洗涤缓冲液(PBS与0.1%(v / v)吐温20)在同一管中洗涤膜5分钟。将膜与一抗2(靶向FAHD1 ;见 表1)在同一管中孵育,在室温下在免疫印迹封闭缓冲液中稀释1:500,同时滚动。
- 滚动时,用20mL蛋白质印迹洗涤缓冲液在同一管中洗涤膜三次,每次10分钟。将膜在室温下用HRP偶联二抗(参见 材料表)在5mL蛋白质印迹封闭缓冲液中稀释1:3000孵育30分钟。
- 在同一管中洗涤膜三次,每次10分钟,每次用20mL蛋白质印迹洗涤缓冲液,每次两次5分钟,每次1x PBS。用镊子小心地握住膜,并用膜的相对(下部)边缘触摸一块纤维素或一张Whatman纸,干燥膜。将膜(暴露的一面朝上)放在清洁的玻璃板上。
- 使用移液器用1 mL制备的ECL蛋白质印迹底物小心地覆盖整个膜,注意不要产生任何气泡。让ECL溶液孵育3分钟,然后立即使用X射线胶片或使用成像系统显影膜。
注意:如果蛋白质没有在阳性对照中检测到,则可能表明蛋白质不溶,或者不存在足够的量来检测抗体。如果只加载了纳克的阳性对照,则第一种情况更有可能。如果根本没有检测到蛋白质,请检查抗体的质量,并可能改用多克隆抗体而不是单克隆抗体。在极少数情况下,即对于某些疏水性蛋白质,该蛋白质可以在离心后检测到,但在过滤后不能检测到。在这种情况下,建议对疏水性蛋白质使用特殊的过滤单元。
- 任选地,在蛋白质印迹后对PVDF膜进行染色,以控制蛋白质从SDS-PAGE凝胶成功转移到PVDF膜。
注意:建议使用考马斯染色进行故障排除、方法开发和记录,但请记住,在应用此方案后,膜会因进一步的蛋白质印迹分析而丢失。Ponceau S染色的染色较弱,但如果要重新探查膜,则可以使用。- 准备装有染色(考马斯或庞索S)和脱色溶液的小托盘。
- 使用镊子将膜放入染色溶液中并轻轻摇动,直到膜染色良好(5-10分钟)。
- 将膜转移到脱色溶液中并摇动,直到溶液饱和(5-10分钟)。重复脱色步骤,直到可以在膜上观察到蛋白质条带;如果根本没有观察到条带,则以较长的孵育时间重复染色。使用镊子将其放在玻璃板上,使膜干燥。
4.测试:硫酸铵沉淀
注意:硫酸铵沉淀是一种通过改变蛋白质溶解度来纯化蛋白质的方法。在初步实验中,硫酸铵浓度依次增加到沉淀最大量蛋白质污染物的值,同时将FAHD1留在溶液中。 通过 蛋白质印迹分析再次探测蛋白质的溶解度。
- 从步骤2.3开始:解冻样品的等分试样或在蛋白质提取后直接进行(即,不冻结样品)。使用0.22μm过滤单元过滤样品,以排除解冻后可能的沉淀物。在冰上制备六个1.5 mL管,并将250μL样品转移到每个管中。
- 在上面制备的试管中制备5%,10%,15%,20%,25%和30%硫酸铵的稀释系列,并用蛋白质裂解缓冲液组成最终体积至1000μL。将样品在4°C下在管旋转器上孵育过夜(参见 材料表)。
- 使用台式离心机,在4°C下以10,000× g 离心30分钟,并小心地将所有上清液转移到单独的管中。将得到的沉淀风干,并将其中的每一个重悬于1000μLddH2O中。
- 对于上一步中的每对重悬沉淀和上清液,将40μL与10μL5x SDS样品缓冲液混合,并在95°C下打开盖子沸腾,直到大部分液体蒸发。然后,将沉淀重悬于ddH2O中50%DMSO的混合物中。
- 执行SDS-PAGE(步骤3),但将凝胶在80 V下电泳3小时。对于每种浓度的硫酸铵,成对加载来自重悬沉淀和上清液的样品(步骤4.3)。执行蛋白质印迹分析(步骤 3)。
- 检查最高浓度的硫酸铵,此时待纯化的蛋白质(即FAHD1)保留在来自上清液的样品中。根据结果,为感兴趣的蛋白质定义硫酸铵沉淀方案,以用于未来的实验。
注意:众所周知,硫酸铵会扭曲SDS-PAGE和蛋白质印迹。随着硫酸铵浓度的增加,蛋白质印迹分析的质量将受到影响。然而,与前面的步骤3一样,该分析用于在给定浓度的硫酸铵下检查目标蛋白质的溶解度。该协议旨在沉淀其他蛋白质,而要纯化的蛋白质必须保持可溶性。
5. 测试:使用FPLC进行离子交换色谱
注意:具有带电官能团的分子与FPLC的二氧化硅颗粒柱结合,从而能够根据其表面电荷分化蛋白质。使用阳离子交换柱和阴离子交换柱执行此步骤两次(参见 材料表)。阳离子或阴离子交换色谱的方案步骤相同,但要使用的缓冲液不同(见 表1);均具有"低盐"15 mM NaCl和"高盐"1 M NaCl条件。对于使用的色谱柱,建议流速为 1 mL/min。
- 用阴离子或阳离子交换柱设置FPLC系统。用5个20%EtOH(在H 2O中)的柱体积(CV)洗涤色谱柱,然后用5个ddH2O的CVs洗涤柱。或者,用1 CV的低盐缓冲液,高盐缓冲液和再次低盐缓冲液洗涤柱,直到在色谱图中不再观察到峰,但至少洗涤一次。
- 在小规模上确定硫酸铵沉淀的最佳方案(步骤4)后,将沉淀方案应用于10 mL原始组织匀浆(步骤2)。任选地,用低盐缓冲液透析样品。
- 将样品涂在色谱柱上(例如,通过进样或使用样品泵)并收集流出物。用1 CV的低盐缓冲液洗涤色谱柱。
- 在3个CV内设置从100%低盐缓冲液/ 0%高盐缓冲液到0%低盐缓冲液/100%高盐缓冲液的线性梯度洗脱。连续收集1 mL级分。梯度完成后,继续使用高盐缓冲液运行,直到在色谱图中检测到1 CV范围内不再有蛋白质相关峰(280/255nm处的UV吸收)。
- 应用1 mL溶解在0.5 M NaOH(ddH 2 O中)的25%SDS以清洁色谱柱。连续用3个ddH 2 O的CVs和3个20%EtOH的CV(在ddH2O中)洗涤色谱柱。
- 收集所有峰级分和流通的SDS-PAGE样品, 并通过 蛋白质印迹探测它们是否存在感兴趣的蛋白质(步骤3)。将收集的馏分在液氮中速冻,并将其储存在-80°C。
- 蛋白质印迹分析完成后,解冻并池化含有目标蛋白质的馏分,并丢弃其他部分。用备用色谱柱(即阳离子或阴离子交换柱)重复步骤5.1-5.5。
- 在探测到两个色谱柱后,为感兴趣的蛋白质定义FPLC方案,以便在未来的实验中使用。使用超离心过滤装置(10 kDa,见 材料表)将蛋白质溶液的体积减小到2 mL。
注意:这一系列实验有两个预期结果。要么感兴趣的蛋白质已经附着在其中一个色谱柱上,并且蛋白质溶液在洗脱后已经非常纯净,要么在这两种情况下蛋白质都保持在流通中。在后一种情况下,尽管蛋白质处于流通状态,但此步骤的清洁效果可能仍然显着。在这种情况下,对于猪肾和小鼠肝脏中的FAHD1,仍将进行离子交换的这一步骤。如果阳离子或阴离子交换柱都不能提供适当的清洁效果,则可以尝试改变裂解液和缓冲液的pH值,并在施用于FPLC之前对样品进行电泳缓冲液透析。
6. 使用硫酸铵沉淀和FPLC的指定子协议进行蛋白质提取
注意:用于FPLC的硅胶柱中的多孔颗粒(参见 材料表)能够根据其流体动力学半径区分蛋白质。所描述的步骤将使用FPLC系统,使用尺寸排阻色谱(SEC)进行。对于使用的SEC色谱柱(参见 材料表),建议流速为0.3 mL/min。
- 准备所有必需的材料(见步骤1),并从组织中提取总蛋白质(见步骤2)。对未用于测试的所有组织匀浆进行硫酸铵沉淀(参见步骤4)。对于较大体积的裂解物,使用超离心过滤装置(10 kDa;参见 材料表)浓缩裂解物,体积较小,为50 mL或更小。
- 使用离子交换色谱法执行第一个纯化步骤(参见步骤5)。
- 准备用于蛋白质印迹的样品,如前面的步骤中所述。进行蛋白质印迹分析,并池化所有含有离子交换色谱的FAHD1级分。
- 使用超离心过滤装置 (10 kDa) 将蛋白质溶液的体积减小到 2 mL。用0.45μm和0.22μm注射器过滤单元按顺序过滤溶液,以除去任何微沉淀物。
- 用1 CV的SEC电泳缓冲液平衡SEC色谱柱(见 表1),其中包含1 mM DTT。将样品上样到色谱柱上并运行色谱法,直到洗脱所有蛋白质(1-2 CV)。
- 收集对应于色谱图中显着峰(280/255nm处的紫外线吸收)的1mL流出部分,并制备每个收集的馏分的50μL样品用于SDS-PAGE和蛋白质印迹分析,如前面的步骤中所述。使用液氮快速冷冻所有馏分,并将其储存在-80°C。
- 用1 CV的ddH 2 O和1 CV的20%EtOH(在ddH2O中)连续洗涤SEC色谱柱。进行蛋白质印迹分析,并池化所有含有FAHD1的馏分。使用超离心过滤装置将蛋白质溶液的体积减少到2 mL(10 kDa,参见 材料表)。
- 使用商业BCA检测试剂盒评估蛋白质浓度(参见 材料表)。
注意:流动相的pH和盐含量可能影响球状蛋白20的洗脱曲线。酸性或碱性条件可导致峰被较少定义和增加蛋白质 - 基质相互作用导致蛋白质部分保留在色谱柱20上。这种效应可用于进一步的蛋白质纯化。用不同的流速、pH和盐浓度重复步骤6可以提高蛋白质20的纯度。
7. 银染
注意:需要对SDS-PAGE凝胶进行银染色分析,以检查考马斯染色可能看不到的蛋白质污染。以下协议是可以在文献中找到的众多版本之一21.通过在干净的玻璃托盘中摇动来执行所有孵育步骤。将所有含银和含甲醛的液体收集在特殊的废物容器中,并妥善丢弃。
- 将SDS-PAGE凝胶在银染色固定溶液中孵育(见 表1)在冷室中过夜。将凝胶在室温下在银染色孵育溶液(见 表1)中孵育3小时。任选地,加入戊二醛(见 表1)以改善对微弱条带的检测。将凝胶在ddH2O中洗涤四次,每次10分钟。
- 将凝胶在银染银溶液中孵育1小时(见 表1)。
注意:请注意,从现在开始,所有液体和凝胶本身都含有有毒的银和甲醛。 - 将凝胶在银染色显影剂溶液(见 表1)中孵育,剧烈摇动,直到条带清晰可见。为了停止反应,丢弃显影剂溶液并立即将凝胶在银染色停止溶液中孵育(见 表1)至少10分钟。
注意:在步骤7.2和7.3中染色的条带将不断变得更加发达。如果染色较弱,则可能需要向溶液中添加比所述更多的甲醛。
结果
使用所提出的方案从猪肾和小鼠肝脏中提取FAHD1蛋白。对于小鼠组织,在最后的纯化步骤后需要多个器官才能获得几μg。出于这个原因,本文重点介绍从猪肾脏中提取FAHD1,这是一个更典型的实验。从小鼠肝脏中提取FAHD1是为了呈现该方案的困难和可能的陷阱。通常建议使用显示要纯化的蛋白质的高表达水平的器官。人蛋白图谱22 可能有助于估计模型系统中的表达,或者可以使用?...
讨论
协议中的关键步骤
遵循处理蛋白质的常见准则至关重要,例如在冰上以及在中等pH和盐条件下工作。使用蛋白酶抑制剂对该方法有益,而强烈建议使用蛋白酶体抑制剂。冷冻和解冻样品可能总是导致蛋白质沉淀(至少部分),因此任何解冻的初始蛋白质裂解物等分试样(步骤2)都应连续处理,不得中断。一般建议解冻后的离心和过滤,以除去微沉淀。
从组织?...
披露声明
作者没有相互竞争的经济利益。
致谢
作者非常感谢Ayse Öztürk和Eva Albertini的技术援助。用于生成肝组织的小鼠在Univ.-Doz的监督下维持。Pidder Jansen-Dürr博士(因斯布鲁克大学生物医学老龄化研究所,Rennweg 10,6020因斯布鲁克,奥地利)。
材料
| Name | Company | Catalog Number | Comments |
| 0.22 µm filter units | MERCK | SLGP033RS | Millex-HP, 0.22 µm, PES 33 mm, not steril |
| 0.45 µm filter units | MERCK | SLHP033NS | Millex-HP, 0.45 µm, PES 33 mm, not steril |
| 15 mL Falcon tubes | VWR | 734-0451 | centrifugal tubes |
| 50 mL Falcon tubes | VWR | 734-0448 | centrifugal tubes |
| 96-Well UV Microplate | Thermo-Fischer | 8404 | UV/VIS transparent flat-bottom 96 well plates |
| Acrylamide/Bis Solution (40%, 29:1 ratio) | BIO-RAD | #1610147 | 40% acrylamide/bis-acrylamide, 29:1 (3.3% crosslinker) solution for casting polyacrylamide gels |
| ÄKTA FPLC system | GE Healthcare Life Sciences / Cytiva | - | using the FPLC system by GE Healthcare; different custom versions exist; this work used the "ÄKTA pure" system |
| Amicon Ultra-15, PLGC Ultracel-PL Membran, 10 kDa | MERCK | UFC901024 | centrifigal filters for protein enrichment; 10 kDa molecular mass filter; 15 mL |
| Amicon Ultra-4, PLGC Ultracel-PL Membran, 10 kDa | MERCK | UFC801024 | centrifigal filters for protein enrichment; 10 kDa molecular mass filter; 4 mL |
| Ammonium sulfate powder | MERCK | A4418 | ammonium sulphate for molecular biology, ≥99.0% |
| Ammoniumpersulfat reagent grade, 98% | MERCK | 215589 | Catalyst for acrylamide gel polymerization. |
| Coomassie Brilliant blue R 250 | MERCK | 1125530025 | Coomassie Brilliant blue R 250 (C.I. 42660) for electrophoresis Trademark of Imperial Chemical Industries PLC. CAS 6104-59-2, pH 6.2 (10 g/l, H2O, 25 °C) |
| Dialysis tubing cellulose membrane | MERCK | D9277 | Cellulose membranes for the exchange of buffers via dialysis. |
| Eppendof tubes 1.5 mL | VWR | 525-1042 | microcentrifugal tubes; autoclaved |
| HiLoad 26/600 Superdex 75 pg | GE Healthcare Life Sciences / Cytiva | 28989334 | HiLoad Superdex 75 pg prepacked columns are for high-resolution size exclusion chromatography of recombinant proteins |
| Immun-Blot PVDF Membrane | BIO-RAD | #1620177 | PVDF membranes are protein blotting membranes optimized for fluorescent and multiplex fluorescent applications. |
| Mini Trans-Blot Electrophoretic Transfer Cell | BIO-RAD | #1703930 | Use the Mini Trans-Blot Cell for rapid blotting of Mini-PROTEAN precast and handcast gels. |
| Mini-PROTEAN Tetra Vertical Electrophoresis Cell for Mini Precast Gels | BIO-RAD | #1658004 | 4-gel vertical electrophoresis system, includes electrode assembly, companion running module, tank, lid with power cables, mini cell buffer dam. |
| Mono Q 10/100 GL | GE Healthcare Life Sciences / Cytiva | 17516701 | Mono Q columns are strong anion exchange chromatography columns for protein analysis or small scale, high resolution polishing of proteins. |
| Mono S 10/100 GL | GE Healthcare Life Sciences / Cytiva | 17516901 | Mono S columns are strong cation exchange chromatography columns for protein analysis or small scale high resolution polishing of proteins. |
| PageRuler Prestained Protein Ladder, 10 to 180 kDa | Thermo-Fischer | 26616 | A mixture of 10 blue-, orange-, and green-stained proteins (10 to 180 kDa) for use as size standards in protein electrophoresis (SDS-PAGE) and western blotting. |
| Pierce BCA Protein Assay Kit | Thermo-Fischer | 23225 | A two-component, high-precision, detergent-compatible protein assay for determination of protein concentration. |
| Sonifier 250; Ultrasonic Cell Disruptor w/ Converter | Branson | - | New models at https://www.emerson.com/documents/automation/brochure-sonifier-sfx250-sfx550-cell-disruptors-homogenizers-branson-en-us-168180.pdf |
| Swine Anti-Rabbit Immunoglobulins/HRP (affinity isolated) | Agilent Dako | P0399 | The antibody used for horseradish peroxidase conjugation reacts with rabbit immunoglobulins of all classes. |
| TEMED, 1,2-Bis(dimethylamino)ethane, TMEDA | MERCK | T9281 | TEMED (N,N,N′,N′-Tetramethylethylenediamine) is molecule which allows rapid polymerization of polyacrylamide gels. |
| Tube Roller | - | - | A general tube rotator roller; e.g. a new model at https://labstac.com/de/Mixer/Roller/c/71 |
| Tube Rotator | - | - | A general tube rotator wheel; e.g. a new model at https://labstac.com/de/Tube-Roller/p/MT123 |
| ULTRA-TURRAX; T 25 digital | IKA | 0003725000 | New models at https://www.ika.com/de/Produkte-Lab-Eq/Dispergierer-Dipergiergeraet-Homogenisierer-Homogenisator-csp-177/T-25-digital-ULTRA-TURRAX-cpdt-3725000/ |
参考文献
- Pircher, H., et al. Identification of FAH domain-containing protein 1 (FAHD1) as oxaloacetate decarboxylase. Journal of Biological Chemistry. 290 (11), 6755-6762 (2015).
- Pircher, H., et al. Identification of human Fumarylacetoacetate Hydrolase Domain-containing Protein 1 (FAHD1) as a novel mitochondrial acylpyruvase. Journal of Biological Chemistry. 286 (42), 36500-36508 (2011).
- Kang, T. -. W., et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 479 (7374), 547-551 (2011).
- Hong, H., Seo, H., Park, W., Kim, K. K. -. J. Sequence, structure and function-based classification of the broadly conserved FAH superfamily reveals two distinct fumarylpyruvate hydrolase subfamilies. Environmental Microbiology. 22 (1), 270-285 (2020).
- Timm, D. E., Mueller, H. A., Bhanumoorthy, P., Harp, J. M., Bunick, G. J. Crystal structure and mechanism of a carbon-carbon bond hydrolase. Structure. 7 (9), 1023-1033 (1999).
- Bateman, R. L., et al. Mechanistic inferences from the crystal structure of Fumarylacetoacetate Hydrolase with a bound phosphorus-based inhibitor. Journal of Biological Chemistry. 276 (18), 15284-15291 (2001).
- Weiss, A. K. H., et al. Structural basis for the bi-functionality of human oxaloacetate decarboxylase FAHD1. Biochemical Journal. 475 (22), 3561-3576 (2018).
- Etemad, S., et al. Oxaloacetate decarboxylase FAHD1 - a new regulator of mitochondrial function and senescence. Mechanisms of Ageing and Development. 177, 22-29 (2019).
- Weiss, A. K. H., et al. Regulation of cellular senescence by eukaryotic members of the FAH superfamily - A role in calcium homeostasis. Mechanisms of Ageing and Development. 190, 111284 (2020).
- Petit, M., Koziel, R., Etemad, S., Pircher, H., Jansen-Dürr, P. Depletion of oxaloacetate decarboxylase FAHD1 inhibits mitochondrial electron transport and induces cellular senescence in human endothelial cells. Experimental Gerontology. 92, 7-12 (2017).
- Wiley, C. D., et al. Mitochondrial dysfunction induces senescence with a distinct secretory phenotype. Cell Metabolism. 23 (2), 303-314 (2016).
- Weiss, A. K. H., et al. Expression, purification, crystallization, and enzyme assays of Fumarylacetoacetate Hydrolase Domain-containing proteins. Journal of Visualized Experiments: JoVE. (148), e59729 (2019).
- Weiss, A. K. H., et al. Inhibitors of Fumarylacetoacetate Hydrolase Domain Containing Protein 1 (FAHD1). Molcules. 26 (16), 5009 (2021).
- Mizutani, H., Kunishima, N. Purification, crystallization and preliminary X-ray analysis of the fumarylacetoacetase family member TTHA0809 from Thermus thermophilus HB8. Acta Crystallographica Section F Structural Biology and Crystallization Communications. 63 (9), 792-794 (2007).
- Lee, C. H. A simple outline of methods for protein isolation and purification. Endocrinology and Metabolism. 32 (1), 18-22 (2017).
- Amer, H. E. A. Purification of proteins: Between meaning and different methods). Proteomics Technologies and Applications. , (2019).
- Niu, L., Yuan, H., Gong, F., Wu, X., Wang, W. Protein extraction methods shape much of the extracted proteomes. Frontiers in Plant Science. 9, 802 (2018).
- Gordon, J. A. Use of vanadate as protein-phosphotyrosine phosphatase inhibitor. Methods in Enzymology. 201, 477-482 (1991).
- Gallagher, S. R. SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Current Protocols in Essential Laboratory Techniques. , (2012).
- . Effect of pH on Protein Size Exclusion Chromatography Available from: https://www.agilent.com/cs/library/applications/5990-8138EN.pdf (2011)
- Sørensen, B. K., et al. Silver staining of proteins on electroblotting membranes and intensification of silver staining of proteins separated by polyacrylamide gel electrophoresis. Analytical Biochemistry. 304 (1), 33-41 (2002).
- Fagerberg, L., et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Molecular & Cellular Proteomics. 13 (2), 397-406 (2014).
- . Cytiva Life Fundamentals of size exclusion chromatography Available from: https://www.cytivalifesciences.com/en/us/solutions/protein-research/knowledge-center/protein-purification-methods/size-exclusion-chromatography (2022)
- Rosano, G. L., Ceccarelli, E. A. Recombinant protein expression in Escherichia coli: advances and challenges. Frontiers in Microbiology. 5, 172 (2014).
- Rosano, G. L., Morales, E. S., Ceccarelli, E. A. New tools for recombinant protein production in Escherichia coli: A 5-year update. Protein Science: A Publication of the Protein Society. 28 (8), 1412-1422 (2019).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。