
Se requiere una suscripción a JoVE para ver este contenido. Inicie sesión o comience su prueba gratuita.
Method Article
Pinza de tensión Fluorometría en
En este artículo
Resumen
Este artículo describe una mejora de la Fluorometría de Voltaje-Clavija convencional (VCF) donde se usan Fluorescentes Aminoácidos No Naturales (fUAA) en lugar de tintes de maleimida, para sondar reordenamientos estructurales en canales iónicos. El procedimiento incluye inyección de ADN de ovocitos de Xenopus , coinjección de ARN / fUAA y mediciones de corriente y fluorescencia simultáneas.
Resumen
Fluorometría de tensión-abrazadera (VCF) ha sido la técnica de elección para investigar la estructura y función de las proteínas de membrana electrogenic donde las mediciones en tiempo real de la fluorescencia y las corrientes informan simultáneamente sobre los reordenamientos locales y la función global, respectivamente 1 . Mientras que las técnicas estructurales de alta resolución como la microscopía cryoelectrónica o la cristalografía de rayos X proporcionan imágenes estáticas de las proteínas de interés, el VCF proporciona datos estructurales dinámicos que nos permiten vincular los reordenamientos estructurales (fluorescencia) a datos funcionales dinámicos (electrofisiología). Hasta hace poco, la química tiol-reactiva utilizada para el marcado fluorescente dirigido a las plantas de las proteínas restringía el alcance del enfoque porque todas las cisteínas accesibles, incluidas las endógenas, estarían etiquetadas. De este modo se requirió la construcción de proteínas libres de cisteínas endógenas. El etiquetado también se restringió a los sitios accesibles desde el extracelularlado. Esto cambió con el uso de aminoácidos fluorescentes no naturales (fUAA) para incorporar específicamente una pequeña sonda fluorescente en respuesta a la supresión del codón de parada utilizando un par ortogonal tRNA y tRNA sintetasa par 2 . La mejora de VCF sólo requiere un procedimiento de inyección de dos etapas de inyección de ADN (par tRNA / sintetasa) seguido de co-inyección de ARN / fUAA. Ahora, el etiquetado tanto intracelular y sitios enterrados es posible, y el uso de VCF se ha expandido significativamente. La técnica VCF se convierte así en atractiva para estudiar una amplia gama de proteínas y, lo que es más importante, permite investigar numerosos mecanismos reguladores citosólicos.
Introducción
Más de 200 aminoácidos naturales de diversas propiedades químicas y físicas han sido incorporados genéticamente en proteínas en células de E. coli , levaduras y mamíferos [ 3] . El aminoácido no natural se incorpora en respuesta a un codón de parada específico a través de un par ortogonal de tRNA / sintetasa. El enfoque genético para modificar las proteínas ha proporcionado información valiosa sobre la estructura y función de la proteína. Aquí, presentamos un protocolo para el uso de Voltaje-Clamp Fluorometría (VCF) en combinación con un fluorescente SAU.
En VCF, la observación simultánea de datos funcionales y reordenamientos estructurales localizados alrededor de la sonda fluorescente (~ 5 Å) nos permite obtener información dinámica con una resolución de milisegundos 1 . Las sondas fluorescentes alteran su estado de extinción tras el movimiento localizado de la proteína. Un movimiento de sólo 1-2 Å es suficiente para conducir a cambios significativos en la fluorescenciaIntensidad 4 . Después de la identificación del sitio de interés en la proteína diana, el sitio es mutado por mutación puntual. Clásicamente, el residuo se había mutado a una cisteína, mientras que ahora, se introduce un codón de detención ámbar (TAG) para la incorporación genética de fUAA. La proteína se transcribe entonces in vitro .
Mientras que otros sistemas de expresión ( p. Ej., Células de mamíferos) pueden usarse 5 , 6 , 7 , los oocitos de Xenopus son preferibles para estudios de estructura-función debido a su mayor tamaño, lo que conduce a una manipulación más fácil ya una mayor intensidad de fluorescencia (más fluoróforos) Ruido. Además, los ovocitos de Xenopus tienen un bajo trasfondo de las proteínas endógenas 2 , 8 , y la pigmentación oscura en el polo animal se escuda contra la fluorescencia de fondo de tEl citosol. Los ovocitos de Xenopus se eliminan quirúrgicamente y el ADN que codifica el par ortogonal tRNA / tRNA-sintetasa específico para la fUAA se inyecta en el núcleo de los oocitos. Después de un tiempo de incubación de 6-24 h, la proteína ARN se co-inyecta con la fUAA en el citosol de los ovocitos, seguido por un período de incubación de 2-3 días. Con el fin de evitar cualquier daño a la fUAA (fotoblanqueo), los procedimientos que incluyen Anap tienen que llevarse a cabo bajo luz roja para evitar la excitación del fluoróforo.
Se estudian los ovocitos en una configuración de tensión-abrazadera de ovocitos abierta, que se monta en un microscopio de fluorescencia vertical, y se registran simultáneamente cambios de corriente eléctrica y fluorescencia 9 , 10 . Alternativamente, pueden usarse una abrazadera de tensión de dos electrodos 1 o configuraciones de abrazadera de parche 11 . La fluorescencia se excita mediante longitudes de onda apropiadas con bajo ruido RMS y Emisión registrada usando un fotodiodo ligado a un amplificador con alta amplificación.
Hay varias ventajas del uso de aminoácidos fluorescentes no naturales (fUAAs) en fluorometría de tensión-abrazadera. Una es el acceso al lado citosólico de las proteínas de la membrana; Aquí se encuentran muchos procesos reguladores ( por ejemplo, sitios de unión a Ca2 + o nucleótidos, inactivación rápida y cerrada de canales iónicos con voltaje, apertura de poros, acoplamiento de módulos). Todos estos procesos ahora son accesibles para el etiquetado fluorescente.
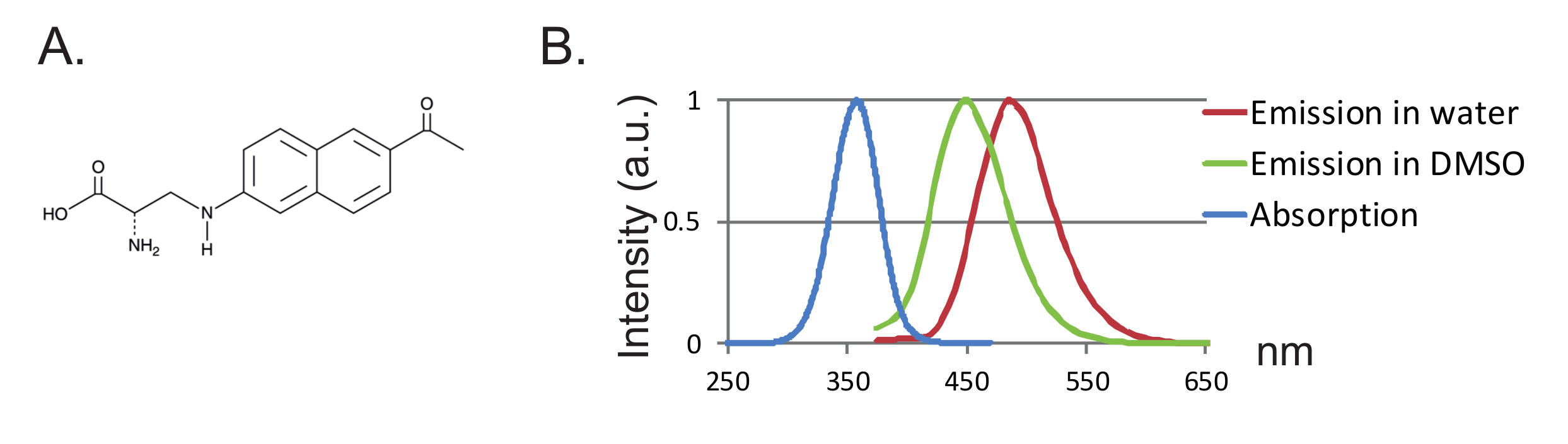
Otra ventaja es el tamaño pequeño de la sonda que conduce a menos alteración de la proteína. Hasta ahora, se han diseñado dos pares ortogonales tRNA / ARNt sintetasa para fUAA 12 , 13 , donde el ácido 3- (6-acetilnaftalen-2-ilamino) -2-aminopropanoico (Anap) es el único fUAA que se ha utilizado en ovocitos de Xenopus 2 ,"Xref"> 8. Anap es un fluoróforo ambientalmente sensible con un peso molecular de 272,3 g / mol y es sólo ligeramente mayor que el triptófano 12 ( Figuras 1A, 1B ). Debido a su pequeño tamaño, es probable que se introduzcan menos efectos estéricos por el fluoróforo en comparación con los fluoróforos convencionales unidos a través de un engarce (típicamente más de 500 g / mol). Por otra parte, en el caso de Anap, el fluoróforo se encuentra más cerca de la cadena principal de proteínas que los vinculados a las cisteínas, y, en consecuencia, Anap está probando más localizados reordenamientos. Por último, la eliminación de las cisteínas endógenas en el VCF convencional con el fin de asegurar el etiquetado específico del sitio ya no es un requisito en el UAA-VCF y por lo tanto (i) deja las proteínas en (casi) su estado nativo y (ii) permite aplicar VCF Para estudiar una gama más amplia de proteínas en las que la función puede ser alterada por la sustitución de cisteína.

Figura 1 : Anap y espectros de fluorescencia. ( A ) Estructura química de Anap. ( B ) Espectro de absorción normalizado y espectros de emisión para Anap 1 nM, demostrando la sensibilidad de la fluorescencia de Anap a la hidrofobicidad del disolvente. Los espectros de emisión se obtuvieron por excitación a 350 nm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Una desventaja del uso de UAA fluorescentes es que una población heterogénea de proteínas puede resultar de la lectura de codón de terminación, reiniciación traslacional, proteínas C-terminales truncadas o diafonía con aminoacilación endógena si la cantidad de ARNt aminoacilados es escasa. Tal expresión de fugas siempre debe comprobarse en ausencia de la fUAA y el tRNA / tRNA synthetase par. Abordamos la cuestión de la transReinicialización lacional y cómo evitarlo para los sitios de inserción N-terminal anteriormente 14 . Sin embargo, cuando la fUAA, tRNA y tRNA sintetasa están presentes en cantidades saturadas, sólo queda una baja probabilidad de expresión de fugas.
La principal diferencia de procedimiento entre fUAA-VCF y VCF convencional es la inyección y manipulación de los ovocitos; La inyección de ADN que codifica el tRNA y la ARNt sintetasa (pAnap) es seguida por la introducción de Anap, que se co-inyecta con el ARNm de la proteína o se añade alternativamente a la solución de incubación como un éster de acetoximetilo (AM).
Protocolo
Las manipulaciones de las ranas se realizaron de acuerdo con las directrices canadienses y han sido aprobadas por el comité de ética (CDEA, protocolo # 15-042) de la Universidad de Montreal.
1. Preparación de mRNA para la incorporación de fUAA
- Elija un sitio de interés en la proteína donde se esperan cambios conformacionales. Seleccione un aminoácido en esta región para sustituir a la fUAA.
NOTA: La elección de la posición se basa en los reordenamientos estructurales que se esperan. Si existe una estructura de alta resolución y una hipótesis de los movimientos esperados, el anap debe colocarse de tal manera que el ambiente químico se altere; Esto podría ser un cambio en la constante dieléctrica (ambiente hidrófobo frente a medio hidrófilo) o, más probablemente, extinción por otro aminoácido. Los mejores extinguidores son los triptófanos. Anap debe estar en contacto con el extintor en un estado (superposición de los radios van-der-Waals) y libre de él enel otro. Si no existen estructuras o modelos de alta resolución, habría que escanear la región de interés. En cualquier caso, es aconsejable seleccionar varias localizaciones cercanas para aumentar la probabilidad de obtener expresión y señal de fluorescencia. Con el fin de minimizar los efectos estéricos durante la maduración y / o función de la proteína, se puede optar por reemplazar los aminoácidos grandes y aromáticos (Phe, Trp, Tyr). Los autores, sin embargo, han experimentado que escanear una región de interés para la inserción de fUAA independientemente del aminoácido sustituido, es más productivo. - Insertar un codón de parada ámbar (TAG) en el sitio seleccionado utilizando mutagénesis dirigida al sitio 15 . Asegúrese de que la proteína de interés no termina en un codón de parada ámbar (TAG). Si es así, mutar a otro (codón de parada ocre o ópalo). Amplificar, aislar y secuenciar el ADN. Obtener ARNm de proteína con transcripción in vitro 16 y almacenar el mRNA a 20 ° C o 80 ° C.
- Obtención quirúrgica de ovocitos en estadio V o VI de ranas Xenopus laevis y defolliculate con colagenasa como se describió previamente 17 .
- Anestesiar las ranas con un anestésico apropiado según el protocolo animal aprobado (aquí: éster etílico del ácido 3-aminobenzoico). Cuando no responden a un pellizco suave a una punta del pie (pérdida del reflejo de la retirada), entonces se anestesian adecuadamente para la cirugía.
- Retire inmediatamente las ranas de la solución anestésica y enjuague bien su piel con agua fresca. Este enjuague impedirá que el animal caiga en niveles más profundos de anestesia al eliminar el producto químico no absorbido de la superficie de la piel.
- Quitar los ganglios ováricos de un lado quirúrgicamente y cuidadosamente abrir los nodos con dos pinzas. Incubar y agitar los ovocitos en "solución estándar de oocitos" (SOS) que contiene 1% (p / v) de colagenasa durante 20-30 minPara defolliculate. Lavar tres veces con solución SOS.
- Seleccionar ovocitos grandes y sanos individualmente e incubarlos en una solución de Barth suplementada con antibióticos (100 U / mL de penicilina, 100 μg / mL de estreptomicina, 10 mg / 100 mL de kanamicina) y 5% de suero de caballo a 18 ºC durante al menos 4 horas antes de la inyección .
NOTA: Después de 2 a 4 cirugías con un retraso de 4 meses, Xenopus laevis se eutanasia por incubación prolongada (> 1 h) con éster etílico del ácido 3-aminobenzoico.
- Para la inyección nuclear de ADN, prepare una punta de inyección larga y delgada para poder alcanzar el núcleo y evitar dañar el ovocito. Llene la punta de la inyección con aceite y monte en el dispositivo nanoinjetor.
- Instale el nano-inyector bajo un microscopio estéreo y utilice una pinza para romper el extremo de la punta. Expulse el aceite hasta que no haya burbujas de aire atrapadas dentro del extremo de la punta.
- Coloque 1 μl de 0,1 μg /# 181; L pAnap en agua libre de nucleasa que contenga NaOH (1% de NaOH 1 N) sobre un trozo de parafilm bajo un estereoscopio y llene la punta de inyección con el ADN.
- Transferir 40 oocitos a un plato de inyección revestido con malla que contenga solución de Barth suplementada con antibióticos.
NOTA: Para hacer el plato de inyección revestido con malla, corte una pieza de tamaño adecuado de malla de nylon de 800 μm para llenar una placa de Petri de poliestireno. Añadir cloroformo al centro y luego colocar la malla en la parte superior. Mantenga la malla plana hasta que el plástico se pone. - A medida que el núcleo del oocito se encuentra en el polo animal (oscuro), apunte la punta de la inyección en el centro del polo animal y empale de modo que la punta llegue cerca del centro del hemisferio animal (o 2-3x la profundidad comparada con la inyección de ARN ). Inyectar 9,2 nL de pAnap en el núcleo de cada ovocito. La punta fina y el volumen pequeño de inyección pueden resultar en una inyección irregular o una punta bloqueada. De vez en cuando, compruebe si la inyección funciona inyectándose en el aire.
NOTA: Si el ADN se inyecta correctamente en el núcleo es incierto. Espere por lo tanto 10 - 40% de los ovocitos a no expresar el par tRNA / sintetasa. Véase Discusión para mayor elaboración. - Incubar los ovocitos en 2 ml de solución de Barth suplementada con antibióticos y 5% de suero de caballo (HS) a 18 ° C durante 6-24 h para permitir la expresión robusta de Anap tRNAs específicos y tRNA-sintetasas.
NOTA: El tiempo de incubación del ADN puede durar varios días antes de la inyección de ARN, pero no aumenta la expresión. - Preparar el nano-inyector para la inyección de ARN (igual que el paso 2.2, pero la punta de inyección no necesita ser tan delgada como para la inyección de ADN). Trabajar sólo bajo luz roja desde este punto para evitar el fotoblanqueo del Anap.
- Mezclar 1 μl de Anap 1 mM con 1 μl de mRNA de 1-2 μg / μL directamente sobre un trozo de parafilm y llenar la punta de inyección con la solución mezclada. Impala justo debajo de la membrana en el(Brillante) e inyectar 46 nL en cada ovocito inyectado con pAnap.
NOTA: La concentración de mRNA requerida depende de la proteína de interés. - Incubar los ovocitos protegidos de la luz en una caja o envueltos en papel de aluminio, en antibióticos suplementados con solución de Barth y suero de caballo al 5% a 18 ° C durante 2-3 días. Intercambiar con la solución fresca de Barth todos los días y eliminar los ovocitos muertos para evitar la contaminación.
- Instale el equipo de sujeción de voltaje de oocitos abierto como se describe anteriormente 18 .
- Montar el sistema de registro electrofisiológico en un microscopio de fluorescencia vertical mediante la instalación de la cámara de grabación en un deslizador que permite moverlo entre el estereoscopio estándar para colocar el ovocito y el microscopio para realizar las mediciones de fluorescencia ( Figura 2c ).
NOTA: La geometría de la cámara para los ovocitos cortados no es adecuada para usar la tra normalNsmitted la luz para la iluminación durante la manipulación. Por lo tanto, una lámpara halógena "cuello de cisne" con filtro rojo se utiliza para iluminar lateralmente desde la parte superior. El condensador del microscopio puede ser removido hacer espacio para bajar la etapa de la cámara de electrofisiología. - Conecte un sistema de detección de fotodiodos al puerto de salida de montaje en C del microscopio de fluorescencia ( Figura 2a ). Conecte la lectura de la fotocorriente a un segundo canal de entrada en el procesador digital de señales (DSP, analógico / digital - digital / analógico).
- Utilice una lámpara halógena de 100 W, 12 V como fuente de luz para la excitación de fluorescencia.
NOTA: Alternativamente, pueden utilizarse quemadores de Hg, pero deben reducirse en intensidad para evitar un fotoblanqueo demasiado rápido durante las grabaciones. La iluminación LED sólo se recomienda si los respectivos LEDs muestran una intensidad significativa en el rango de excitación ( por ejemplo, ~ 350 nm para Anap). La mayoría de los LEDs blancos no llegan lejos en el espectro UV. - InsErt un obturador controlado eléctricamente entre la fuente de luz de excitación y el microscopio y conectar su control (típicamente TTL-pulso) a una salida digital del DSP. Tiempo de TTL-pulso en el software de grabación (véase la documentación del fabricante), de tal manera que el obturador se abre ~ 100 ms antes del comienzo de la grabación. De esta manera, cualquier vibración durante el proceso de apertura no interfiere con la grabación. El tiempo depende de la velocidad y vibración del obturador. Termine el pulso 5ms antes del final de la grabación como se muestra en la Figura 4 . De esta manera, también se registra el valor para la fluorescencia total.
- Inserte un cubo de filtro apropiado (filtro de excitación, espejo dicroico y filtro de emisión) en la torreta de cubo de filtro. Para Anap, utilice el paso de banda Ex: 377/50 nm, el paso largo dicroico 409 nm y el paso banda Em: 470/40 nm.
- Seguir los pasos de preparación para la abrazadera de voltaje de ovocitos abierta como se describió anteriormente y visualizar 18 (preparación del puente de agar, montaje del ovocito, permeabilización de la saponina). Sin embargo, trabaje bajo luz roja en todo momento para evitar blanquear el fluoróforo antes de las grabaciones. Al colocar el ovocito, asegúrese de que el polo animal esté orientado hacia arriba. La pigmentación bajo la membrana del polo animal protege contra la autofluorescencia procedente del citosol y por lo tanto reduce la fluorescencia de fondo.
- Deslice la cámara hacia el microscopio y enfoque con un objetivo de 4X.
- Empalar el ovocito con el electrodo V1 de detección de voltaje (3 M KCl), cambiar al objetivo 40X de inmersión en agua (NA 0.8 - 0.9). Concéntrese en el polo animal que está mirando hacia arriba.
- Apague la luz roja. Seleccione el cubo de filtro correcto girando la torreta de cubo de filtro y el puerto de salida óptica conectado al fotodiodo. Encienda el halEn la lámpara en la intensidad más alta y en breve cambiar el obturador abierto durante 2-5 s para leer la intensidad de fluorescencia de fondo procedente del ovocito. Con la configuración descrita el valor debe ser alrededor de 50-200 pA para Anap.
- Encienda la abrazadera, vuelva a activar el interruptor de baño / protector y ajuste el potencial de membrana (V1 - V2) al potencial de mando girando el mando en la posición I.
- Seleccione el potencial de retención, el protocolo de paso, el número y la longitud de los impulsos, etc. en el software de grabación. Registre las corrientes dependientes del voltaje y las intensidades de fluorescencia Anap.
- Para monitorizar simultáneamente dos localizaciones en la misma proteína, mute un aminoácido extracelular y accesible en cisteína, y elimine otras cisteínas para asegurar un etiquetado específico con química tiol.
- Realice el paso 2.1-2.5.
- Antes de las grabaciones de VCF, incube los oocitos en TMR-maleimida 5 μM en solución de marcado durante 15 min (u otro coloranteCon un espectro no superpuesto comparado con Anap).
- Lave los ovocitos con solución de etiquetado tres veces para eliminar el exceso de colorante.
- Realice el paso 4.1-4.6.
- Inserte un cubo de filtro apropiado para TMR (filtro de excitación, espejo dicroico y filtro de emisión) en la torreta de cubo de filtro. Cambie al cubo del filtro TMR girando la torreta del filtro.
- Lea la fluorescencia de fondo para TMR como se describe para Anap en el paso 4.4.
NOTA: El etiquetado con la química tiol produce una alta fluorescencia de fondo debido al etiquetado inespecífico en la membrana. Por lo tanto, la fluorescencia de fondo TMR puede saturar el amplificador (> 2000 pA). En ese caso, no disminuya la intensidad de la luz, sino que simplemente resta la fluorescencia de fondo añadiendo una corriente de desplazamiento al fotodiodo. En sistemas comercialmente disponibles, utilice la función de "muestreo y retención" en el sistema detector. Observe el valor de fluorescencia de fondo (utilizando un filtro de densidad neutra 10X) en un laboratorio jYa que este valor no será registrado (saturación). - Registre las corrientes dependientes del voltaje y las intensidades de fluorescencia TMR simultáneamente como en el paso 4.6.
- 2. Preparación y inyección de ovocitos

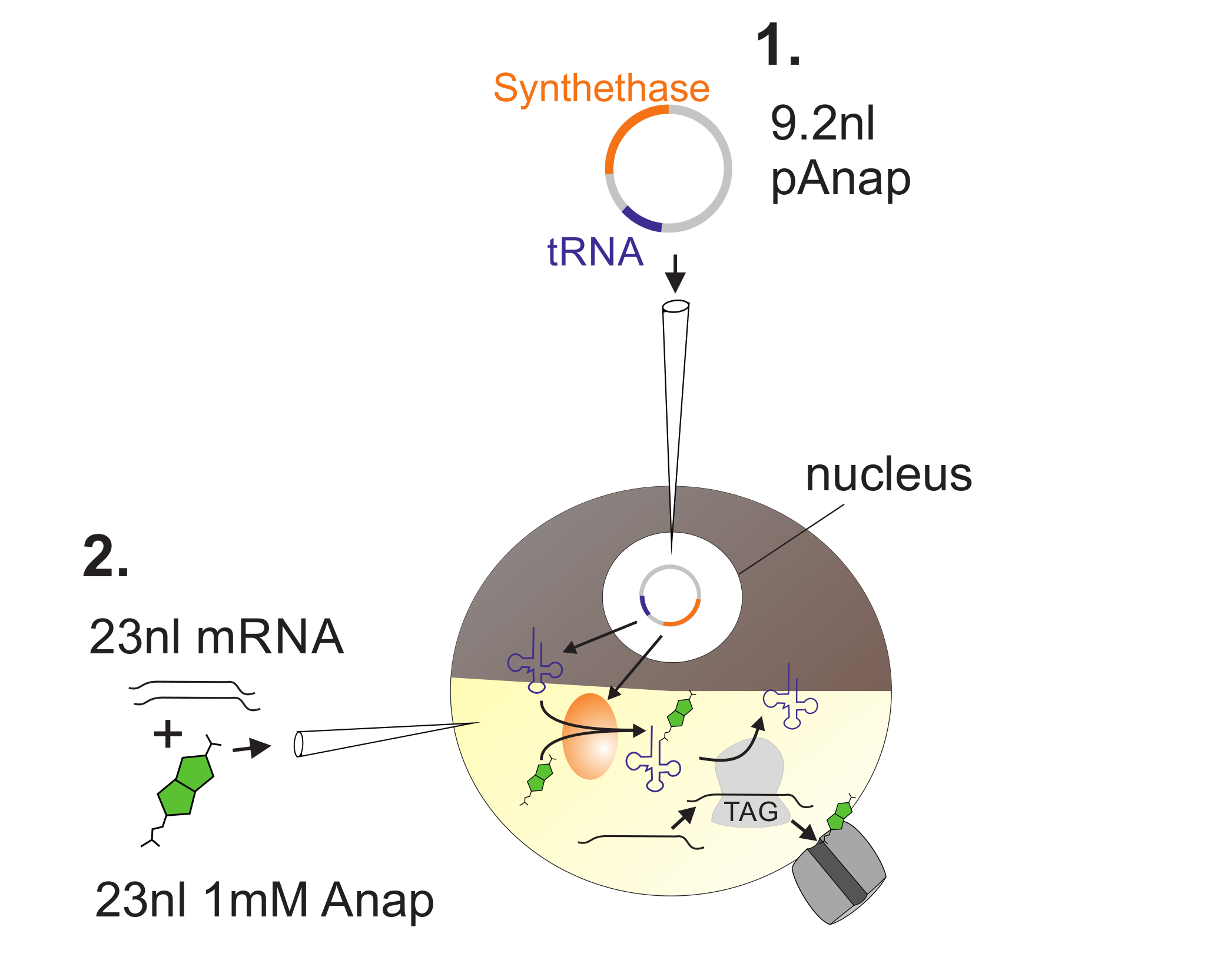
Figura 2 : Ilustración de la inyección de ADN y ARN en los ovocitos de Xenopus para la incorporación de Anap.
En primer lugar, pAnap se inyecta en el núcleo del ovocito de Xenopus ( 1 ). Después de 6-24 h, Anap y el canal ARN se coinjected en el polo vegetal ( 2 ). Anap será ortogonalmente aminoacilado con el tRNA que lleva un anti-codón de bloqueo ámbar, por la aminoacil-ARNt sintetasa que está codificada por pAnap. De esta manera, los Anap-tRNAs aminoacilados son reconocidos por el ribosoma en el codón de parada ámbar insertado en el canalL de ARN, dando como resultado la supresión del codón de parada y la inserción de Anap. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
3. Configuración VCF

Figura 3 < / Strong> : Configuración de VCF. ( A ) Vista lateral de la configuración VCF que muestra el trayecto de la luz dentro del microscopio. El cubo del filtro contiene un filtro de excitación, un espejo dicroico y un filtro de emisión. ( B ) Las dimensiones seleccionadas de la cámara de ovocitos son de 3,4 cm para el radio de la cámara superior (1), 5,5 cm para la cámara inferior (2), 1,4 cm para la anchura de la cámara inferior (3) y 1,7 cm para la anchura de la cámara intermedia (4). ( C ) Vista frontal de la configuración VCF. El primer ocular de la izquierda es para montar el ovocito en la cámara de abrazadera de tensión cortada y para la permeabilización. Entonces, la cámara se desliza bajo el microscopio en el segundo ocular a la derecha. Aquí, el electrodo V1 se inserta en el ovocito usando el objetivo 4X, y la fluorescencia se registra usando el objetivo 40X de inmersión en agua. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
5. VCF bicolor
Resultados
La Figura 4 muestra un ejemplo de grabaciones de VCF obtenidas a partir de un ovocito que expresa canales de Shaker con inactivación rápida eliminada (IR), L382stop-W434F en presencia de pAnap y Anap. La mutación W434F bloquea las corrientes de potasio iónico, lo que permite medir los desplazamientos transitorios de la carga de giro (corrientes de gating). Las grabaciones simultáneas de las corrientes de gating (traza superior) y Anap cambios de intensidad de fluore...
Discusión
La aminoacilación in vivo de tRNAs que se transcriben continuamente junto con la tRNA-sintetasa, hace posible obtener altos niveles de expresión para mediciones de fluorescencia. Para una eficaz incorporación de fUAA, es fundamental que pAnap se inyecte correctamente en el núcleo. Debido a la incertidumbre de la ubicación exacta del núcleo, 10-40% de las inyecciones de ADN se espera que fallen, dando lugar a no expresar (o expresar fugas) ovocitos. Por lo tanto, es importante verificar la expresión en au...
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
PAnap fue un regalo amable del Dr. Peter Schultz (Scripps Research Institute). Este trabajo fue financiado por el Instituto Canadiense de Investigaciones en Salud Subvenciones MOP-102689 y MOP-136894 (a RB) y la Fundación Canadiense para la Innovación Subvención 950-225005.
Materiales
| Name | Company | Catalog Number | Comments |
| Solutions | |||
| Barth's solution | |||
| NaCl | Sigma-Aldrich | S7653 | 90 mM |
| KCl | Fisher Scientific | BP366-500 | 3 mM |
| MgSO4 | Sigma-Aldrich | M-9397 | 0.82 mM |
| CaCl2 | Sigma-Aldrich | C-7902 | 0.41 mM |
| Ca(NO3)2 | Sigma-Aldrich | C-1396 | 0.33 mM |
| HEPES | Sigma-Aldrich | H4034 | 5 mM |
| NaOH hydrate | BDH | BDH7225-4 | pH 7.6 |
| Penicilin | Invitrogen | 15140122 | 100 U/mL |
| Streptomycin | Invitrogen | 15140122 | 100 µg/mL |
| Kanamycin | Invitrogen | 15160054 | 10 mg/100mL |
| Horse Serum (HS) | Invitrogen | 16050122 | 5% |
| SOS Standard Oocyte Solution | |||
| NaCl | Sigma-Aldrich | 746398 | 102 mM |
| KCl | Sigma-Aldrich | 746436 | 3 mM |
| MgCl2 | Sigma-Aldrich | M9272 | 1 mM |
| HEPES | Sigma-Aldrich | H4034 | 5 mM |
| External recording solution | |||
| N-methyl-D-glucamine (NMDG) | Alfa Aesar | L14282 | 115 mM |
| HEPES | Sigma-Aldrich | H4034 | 10 mM |
| Calcium hydroxide | Sigma-Aldrich | 239232 | 2 mM |
| MES hydrate | Sigma-Aldrich | 258105 | pH 7.2 |
| Internal recording solution | |||
| N-methyl-D-glucamine (NMDG) | Alfa Aesar | L14282 | 115 mM |
| HEPES | Sigma-Aldrich | H4034 | 10 mM |
| Ethylenediamine Tetraacetic Acid (EDTA) | Fisher Scientific | E478-500 | 2 mM |
| MES hydrate | Sigma-Aldrich | 258105 | pH 7.2 |
| Labeling solution | |||
| KOH | Fisher Scientific | P250-1 | 115 mM |
| HEPES | Sigma-Aldrich | H4034 | 10 mM |
| Calcium hydroxide | Sigma-Aldrich | 239232 | 2 mM |
| MES hydrate | Sigma-Aldrich | 258105 | pH 7.2 |
| TMR stock solution | |||
| Tetramethylrhodamine-5-maleimide (TMR) | Molcular Probes by Life Technologies | T6027 | 5 mM in DMSO |
| Anap stock solution | |||
| Anap | ABZENA (TCRS) | Custom synthesis TCRS-170 | 1 mM in nuclease-free water and 1% NaOH 1 N |
| Name | Company | Catalog Number | Comments |
| Material/Equipment | |||
| pAnap | Addgene | 48696 | |
| High Performance Oocyte Clamp | Dagan Corporation | CA-1B | |
| Gpatch Acquisition software | Department of Anesthesiology, University of California, Los Angeles | ||
| Analysis software | Department of Anesthesiology, University of California, Los Angeles | ||
| Recording Chamber | Custom machined | ||
| Photo diode detection system | Dagan Corporation | PhotoMax-200/PIN | |
| Electrical shutter driver | UNIBLITZ | VCM-D1 |
Referencias
- Mannuzzu, L. M., Moronne, M. M., Isacoff, E. Y. Direct physical measure of conformational rearrangement underlying potassium channel gating. Science. 271 (5246), 213-216 (1996).
- Kalstrup, T., Blunck, R. Dynamics of internal pore opening in K(V) channels probed by a fluorescent unnatural amino acid. Proc Natl Acad Sci U S A. 110 (20), 8272-8277 (2013).
- Xiao, H., Schultz, P. G. At the Interface of Chemical and Biological Synthesis: An Expanded Genetic Code. Cold Spring Harb Perspect Biol. 8 (9), (2016).
- Blunck, R. Chapter 9. Handbook of Ion Channels. , 113-133 (2015).
- Chatterjee, A., Guo, J., Lee, H. S., Schultz, P. G. A genetically encoded fluorescent probe in mammalian cells. J Am Chem Soc. 135 (34), 12540-12543 (2013).
- DeBerg, H. A., Brzovic, P. S., Flynn, G. E., Zagotta, W. N., Stoll, S. Structure and Energetics of Allosteric Regulation of HCN2 Ion Channels by Cyclic Nucleotides. J Biol Chem. 291 (1), 371-381 (2016).
- Shen, B., et al. Genetically encoding unnatural amino acids in neural stem cells and optically reporting voltage-sensitive domain changes in differentiated neurons. Stem Cells. 29 (8), 1231-1240 (2011).
- Aman, T. K., Gordon, S. E., Zagotta, W. N. Regulation of CNGA1 Channel Gating by Interactions with the Membrane. J Biol Chem. 291 (19), 9939-9947 (2016).
- Haddad, G. A., Blunck, R. Mode shift of the voltage sensors in Shaker K+ channels is caused by energetic coupling to the pore domain. J Gen Physiol. 137 (5), 455-472 (2011).
- Batulan, Z., Haddad, G. A., Blunck, R. An intersubunit interaction between S4-S5 linker and S6 is responsible for the slow off-gating component in Shaker K+ channels. J Biol Chem. 285 (18), 14005-14019 (2010).
- Kusch, J., et al. How subunits cooperate in cAMP-induced activation of homotetrameric HCN2 channels. Nat Chem Biol. 8 (2), 162-169 (2012).
- Lee, H. S., Guo, J., Lemke, E. A., Dimla, R. D., Schultz, P. G. Genetic incorporation of a small, environmentally sensitive, fluorescent probe into proteins in Saccharomyces cerevisiae. J Am Chem Soc. 131 (36), 12921-12923 (2009).
- Summerer, D., et al. A genetically encoded fluorescent amino acid. Proc Natl Acad Sci U S A. 103 (26), 9785-9789 (2006).
- Kalstrup, T., Blunck, R. Reinitiation at non-canonical start codons leads to leak expression when incorporating unnatural amino acids. Sci Rep. 5, 11866 (2015).
- Liu, H., Naismith, J. H. An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 8, 91 (2008).
- Beckert, B., Masquida, B. Synthesis of RNA by in vitro transcription. Methods Mol Biol. 703, 29-41 (2011).
- Goldin, A. L. Maintenance of Xenopus laevis and oocyte injection. Methods Enzymol. 207, 266-279 (1992).
- Rudokas, M. W., Varga, Z., Schubert, A. R., Asaro, A. B., Silva, J. R. The Xenopus oocyte cut-open vaseline gap voltage-clamp technique with fluorometry. J Vis Exp. (85), (2014).
- Zhao, J., Blunck, R. The isolated voltage sensing domain of the Shaker potassium channel forms a voltage-gated cation channel. Elife. 5, (2016).
- Posson, D. J., Ge, P., Miller, C., Bezanilla, F., Selvin, P. R. Small vertical movement of a K+ channel voltage sensor measured with luminescence energy transfer. Nature. 436 (7052), 848-851 (2005).
- Chanda, B., Asamoah, O. K., Blunck, R., Roux, B., Bezanilla, F. Gating charge displacement in voltage-gated ion channels involves limited transmembrane movement. Nature. 436 (7052), 852-856 (2005).
- Taraska, J. W., Puljung, M. C., Zagotta, W. N. Short-distance probes for protein backbone structure based on energy transfer between bimane and transition metal ions. Proc Natl Acad Sci U S A. 106 (38), 16227-16232 (2009).
- Baker, B. J., et al. Genetically encoded fluorescent sensors of membrane potential. Brain Cell Biol. 36 (1-4), 53-67 (2008).
- Miranda, P., et al. State-dependent FRET reports calcium- and voltage-dependent gating-ring motions in BK channels. Proc Natl Acad Sci U S A. , (2013).
- Sisido, M., Ninomiya, K., Ohtsuki, T., Hohsaka, T. Four-base codon/anticodon strategy and non-enzymatic aminoacylation for protein engineering with non-natural amino acids. Methods. 36 (3), 270-278 (2005).
- Hohsaka, T., Ashizuka, Y., Murakami, H., Sisido, M. Five-base codons for incorporation of nonnatural amino acids into proteins. Nucleic Acids Res. 29 (17), 3646-3651 (2001).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados