
Se requiere una suscripción a JoVE para ver este contenido. Inicie sesión o comience su prueba gratuita.
Method Article
Uso de la sola molécula fluorescente en Situ del hibridación (SM-pescado) para cuantificar y Localize mRNAs en oocitos murinos
En este artículo
Resumen
Reproducible, contar los números de los mRNAs de los ovocitos, in situ de la fluorescencia de una molécula de RNA hibridación (RNA-pescado) fue optimizado para las células no adherentes. Ovocitos se recolectaron, cruzado por hibridación con las sondas específicas de transcripción y cuantificaron utilizando un software de cuantificación de la imagen.
Resumen
Métodos actuales que utiliza habitualmente para cuantificar el mRNA en ovocitos y embriones incluyen la reacción en cadena de polimerasa de transcripción inversa digital (dPCR), cuantitativo, en tiempo real RT-PCR (RT-qPCR) y la secuencia de RNA. Cuando estas técnicas se realizan con un solo óvulo o embrión, copia baja mRNAs no se detectan confiablemente. Para superar este problema, ovocitos o embriones pueden se agruparon para el análisis; sin embargo, esto conduce a menudo a la alta variabilidad entre las muestras. En este protocolo, describimos el uso de hibridación fluorescente in situ (FISH) usando química de DNA ramificado. Esta técnica identifica el patrón espacial de mRNAs en células individuales. Cuando la técnica es encontrar lugar y seguimiento informático, también puede cuantificar la abundancia de mRNAs en la célula. Usando esta técnica, hay menor variabilidad dentro de un grupo experimental y se requieren menos ovocitos y embriones para detectar diferencias significativas entre los grupos experimentales. Comercialmente disponible DNA ramificado SM-kits de peces han sido optimizados para detectar mRNAs en tejidos seccionados o células adherentes en diapositivas. Sin embargo, ovocitos no eficazmente a diapositivas y algunos reactivos en el kit eran demasiado duros, resultando en lisis de ovocitos. Para evitar esta lisis, se hicieron varias modificaciones al kit de pescado. Específicamente, buffers de permeabilización y lavado de ovocitos diseñadas para la inmunofluorescencia de oocitos y embriones reemplazados los almacenadores intermediarios del propietarios. La permeabilización, lavados e incubaciones con amplificador y puntas de prueba fueron realizadas en placas de 6 pocillos y ovocitos fueron colocados en portaobjetos en el extremo del protocolo utilizando medios de montaje. Estas modificaciones fueron capaces de superar las limitaciones del kit disponible comercialmente, en particular, la lisis de ovocitos. Precisa y reproducible, contar el número de los mRNAs de los ovocitos, se utilizó el software de computadora. Juntos, este protocolo representa una alternativa a la PCR y secuenciación para comparar la expresión de las transcripciones específicas en las células.
Introducción
Reacción en cadena reversa-transcriptase de polimerasa (PCR) ha sido el estándar de oro para la cuantificación del mRNA. Actualmente se utilizan dos ensayos PCR (dPCR) digital1 y cuantitativa, real time PCR (qPCR)2 . De las dos técnicas PCR, dPCR tiene mayor sensibilidad que la qPCR sugiriendo que podría ser utilizado para medir la abundancia de ARNm en células individuales. Sin embargo, en nuestras manos, análisis de dPCR de mRNAs de baja abundancia en grupos de 5 a 10 ovocitos por cada muestra experimental ha producido datos con baja reproducibilidad y alta variación3. Esto es probablemente debido al error experimental asociado a la extracción de RNA y transcripción reversa eficiencia. La secuencia de RNA también se ha realizado utilizando un único ratón y ovocitos humanos4,5. Esta técnica requiere pasos de amplificación de cDNA para la generación de biblioteca que probablemente aumenta la variabilidad dentro de un grupo experimental. Además, las transcripciones de baja abundancia no pueden ser detectables. Aunque los precios de la secuencia han bajado en los últimos años, todavía puede ser prohibitivo debido al costo alto de Bioinformática análisis de costo. Por último, la localización de mRNA es un proceso dinámico con cambios espaciales que contribuyen a la función de la proteína6. Por lo tanto, nos propusimos adoptar una técnica que produciría medidas cuantitativas precisas y reproducibles y localización de los mRNAs individuales en ovocitos solo.
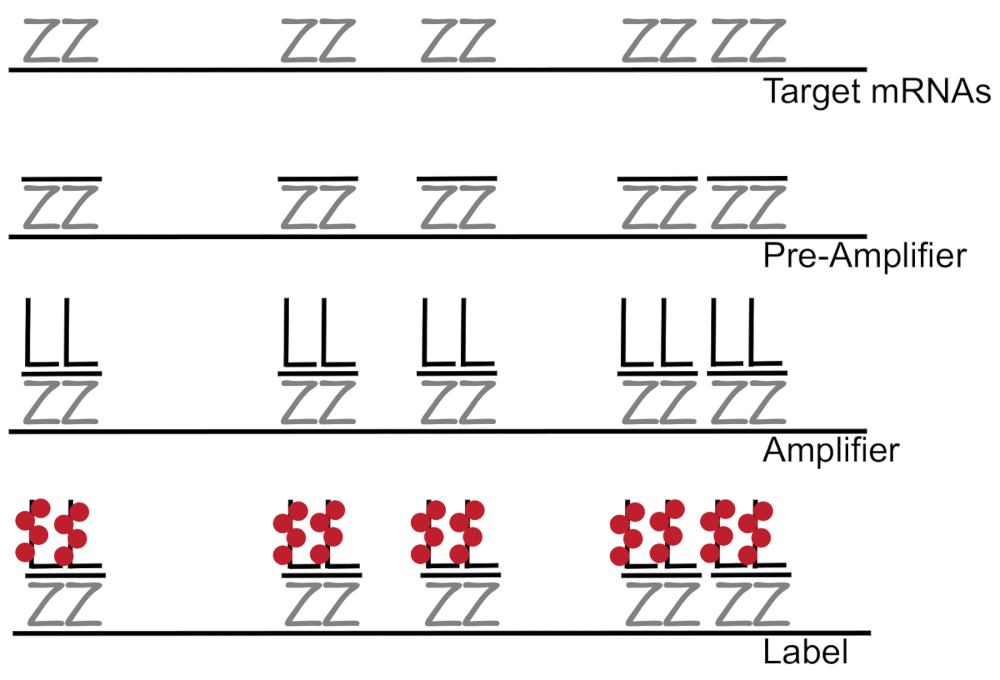
DNA ramificado juntada a hibridación fluorescente in situ amplifica la señal de fluorescencia en lugar de amplificar RNA/cDNA que permite detección de mRNAs individuales en células individuales 7,8,9. El ensayo se realiza a través de una serie de hibridación, amplificación (usando ADN ramificado) y fluorescencia de etiquetado pasos para amplificar la señal de la fluorescencia del7. La técnica comienza con el atascamiento de pares de sonda de 18 - 25 base de oligonucleótidos que son complementarias a un ARNm específico3,8,10. Quince a veinte pares de sonda están diseñados para cada especificidad asegurando de transcripción para la transcripción de destino. La hibridación específica del mRNA es seguida por sondas de preamplificador y amplificador que forman una configuración ramificada. Aproximadamente, 400 etiqueta fluoróforos se unen a cada amplificador, resultando en un 8000-fold incremento de fluorescencia que permite la detección de mRNAs individuales (figura 1)11.

Figura 1: esquema del protocolo SM-pescado. Hibridación secuencial de sonda específica de la transcripción, ramificado ADN amplificador y fluoróforo a un mRNA se muestra de destino. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Estudios previos usando in situ hibridación (SM-pescado) localizado β-actina mRNAs en neuronas individuales12 y papillomavirus humano DNA en cáncer de cuello uterino fluorescencia sola molécula célula líneas7. El software de computadora encontrar lugar y programa de seguimiento identifica la señal fluorescente punteada individual y se ha utilizado con éxito para cuantificar el número de mRNAs en cada célula3,13.
Basado en los resultados de la detección de mRNA en las neuronas12, presumimos que SM-pez sería una herramienta útil para cuantificar los niveles de transcripción en murinos ovocitos y embriones incluyendo mRNAs de baja abundancia. Sin embargo, la técnica está optimizada para el uso con las células fijas adherentes y formaldehído fijada parafina incorporado secciones de tejido (FFPE). Ovocitos no se adhieren a un portaobjetos, incluso cuando están recubiertos con poli-l-lisina. Además, son más frágiles que las células somáticas y las secciones de tejido que resulta en lisis celular cuando se someten a algunos de los buffers propietarios en kits disponibles en el mercado3. Para superar estos desafíos, ovocitos fueron fijos y transferidos manualmente entre las gotas de los amortiguadores. Además, buffers de permeabilización y lavado en los kits fueron substituidos para reducir la lisis celular. Sondas prediseñadas se compran junto con el kit de peces o pueden solicitar transcripciones específicas. Cada conjunto de sonda patentada está disponible en uno de los tres canales de fluorescencia (C1, C2 y C3) para permitir la multiplexación. En el experimento actual, oocitos murinos fueron cuantificados utilizando una sonda C2 Nanog y una sonda de C3 Pou5f1 y doble tinción. Estas sondas se seleccionaron con base en la expresión divulgada de Nanog y Pou5f1 en ovocitos y embriones. En la conclusión de los pasos de hibridación, ovocitos fueron colocados en gotas anti-fade del medio de montaje para aplicación a las diapositivas histológicas. Imágenes confocales fueron utilizados para cuantificar el número de señales fluorescentes punteadas que representan los mRNAs. Además de cuantificar los mRNAs, proyección de imagen demostró también la distribución espacial de los mRNA específicas en la célula, que otros métodos de cuantificación de RNA son incapaces de lograr. Esta técnica demostró para tener poca variabilidad dentro de un grupo experimental que permite el uso de un número menor de ovocitos en cada grupo experimental para identificar diferencias significativas entre grupos experimentales3.
Access restricted. Please log in or start a trial to view this content.
Protocolo
Procedimientos animales fueron revisados y aprobados por el cuidado institucional de Animal y uso de la Universidad de Nebraska-Lincoln y todos los métodos fueron realizados conforme a las normas y directrices pertinentes. Para este estudio, CD-1 marginado ratones tuvieron acceso ad libitum a chow roedor normal y agua; se mantuvieron en un 12:12 oscuras: ciclo de luz.
1. preparación de medios necesarios
- Para los medios de base (OMM), añadir 100 mM NaCl, KCl de 5 mM, 0,5 mM KH2PO4y 1,7 mM CaCl2-2 H2O a 100 mL de agua estéril.
Nota: Medio OMM puede almacenarse hasta por 1 mes. - Por completo los medios de comunicación (OMOPS), añadir 20 mM ácido 3-morpholinopropane-1-sulfónico (MOPS), 1,2 mM MgSO4-7 H2O, 0,5 mM de glucosa, L-lactato, de 6 mM 1 mM ala-gln, taurina, 0,1 mM 1 x aminoácidos no esenciales (AANE), (ácido etilendiaminotetracético) 0,01 mM EDTA), el ácido alfa lipoico de 10 μm, gentamicina de 10 μg/mL sin diluir, 21 mM 1 M NaOH, 5 mM NaHCO3, 0,2 mM piruvato, 0,5 mM citrato de FAF de 4 mg/mL BSA a un 1:10 dilución de OMM en agua estéril para un volumen total de 100 mL. Esterilizar el medio con un filtro de 0,22 μm.
Nota: OMOPS se pueden almacenar hasta 1 semana. - Para el medio de sujeción (HM) Añadir 5% de suero fetal bovino a OMOPS. Hacer 2 mL HM por ratón.
- Para hialuronidasa solución agregar 0,1 mg/mL de hialuronidasa derivada de testículos bovinos, 1 ml HM.
- Para el búfer de fijación combinan paraformaldehído al 4% en 10 mL de 1 x PBS y 0,1% embriones grado polivinilpirrolidona (PVP)14.
- Para preparar 50 mL de tampón de lavado (WB), agregar surfactante no iónico 0.1% y 0.1% PVP de 1 x PBS14.
- Para preparar 10 mL de tampón de permeabilización, añadir 1% de tensioactivo no - iónico a 1 x PBS14.
Nota: Los buffers lavado y permeabilización descritos anteriormente cambie los propiedad búferes en los kits disponibles en el mercado.
2. colección de ovuladas ovocitos de ratones femeninos
- Preparación:
- Estimular ratones hembra en 5 a 8 semanas de edad por inyección intraperitoneal (IP) de 5 UI gonadotropina coriónica equina (eCG) seguida de 5 UI de gonadotropina coriónica (hCG) 44-48 h después de15,16.
- Mantener platos de Petri de 35 mm que contiene 2 mL de HM en una placa calentadora de 37 ° C. Tomar con pipeta una gota de 100 μl de HM que contiene hialuronidasa diluida seguida por tres gotas de 50 μl de HM sin hialuronidasa en caja Petri de 60 mm. Coloque las placas de la placa de gotas que contienen en el calentamiento de 37 ° C antes de usar.
Nota: Las gotas de hialuronidasa deben hacerse justo antes de la disección de cada par de oviducto para prevenir la evaporación y concentración de los componentes de HM con o sin hialuronidasa.
- Eutanasia a ratones, 16 h después de la inyección del IP de hCG, uso de sobredosis de isoflurano seguido por dislocación cervical.
- Limpiar el ratón utilizando etanol al 70%. Exponer la cavidad abdominal y visualizar el tracto reproductor femenino. Sostenga el ovario con pinzas y retire los ligamentos uterinos y exceso de tejido adiposo de alrededor del ovario. Corte el oviducto del útero y el par de ovario oviducto en el HM caliente en el plato de 35 mm.
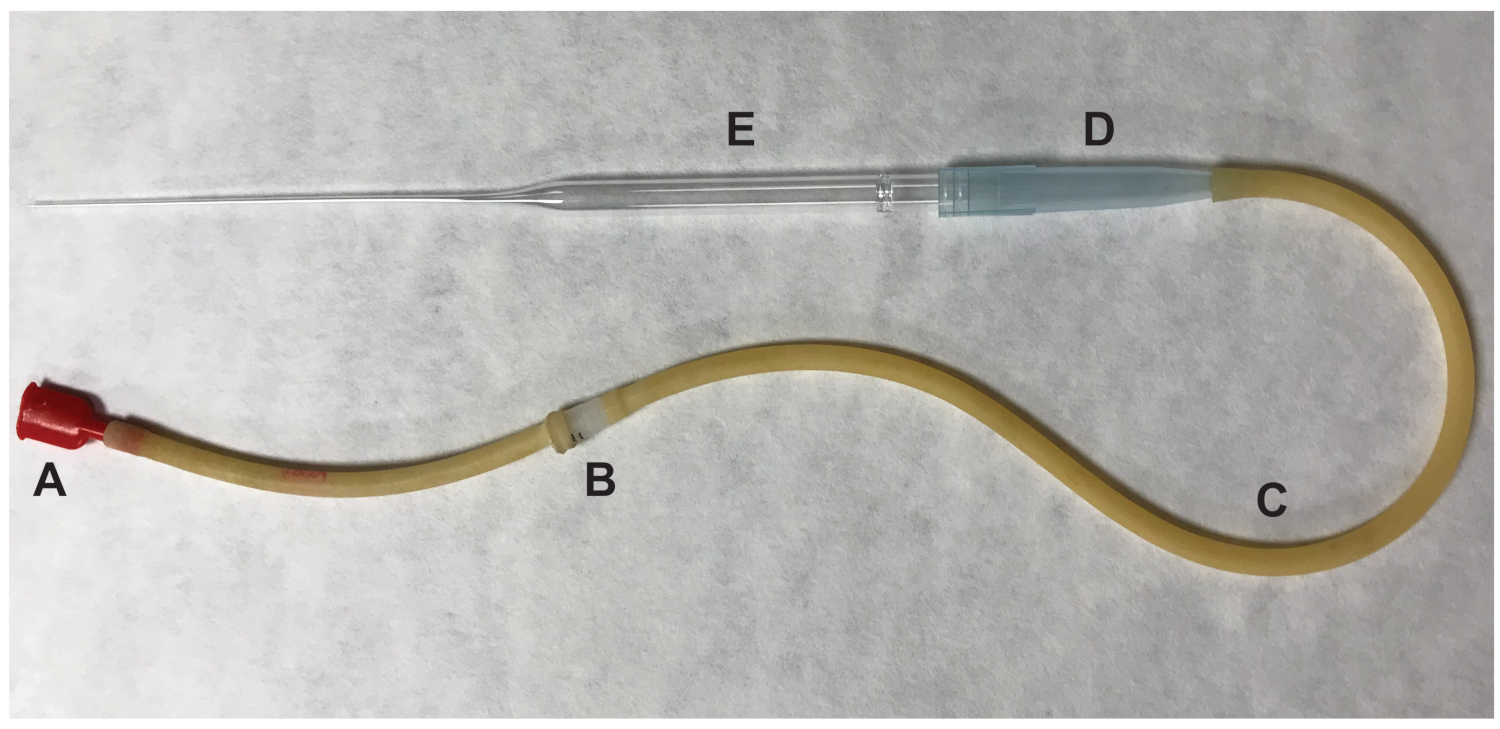
- Quitar el ovario y cualquier tejido adiposo circundante. Romper la ampolla inflamación del oviducto utilizando una aguja de calibre 27 1/2 pulgada. Empuje el oviducto en el sitio de la lágrima y los complejos de la célula ovocito cumulus (AOC) serán expulsados. Transfiera los ovocitos ovulados, que se presumen que en metafase II (MII) de la meiosis, a la caída de 100 μL que contiene medios HM con hialuronidasa con una pipeta de boca (figura 2).

Figura 2 : Partes de la pipeta de boca utilizado para transferencia de ovocitos. (A) boca pieza (B) 0.22 um, 4 mm filtro (C) aspirador tubería (D) 1000 μL Punta de la pipeta (E) 9" pipeta Pasteur. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
- Pipetear los complejos de la célula ovocito-cumulus MII hacia arriba y hacia abajo en la hialuronidasa con HM con la pipeta de boca para desalojar células del cúmulo. Transferir cada ovocito, una vez que están desprovistas de células del cúmulo a un lavado gota HM contiene solamente usando la pipeta de boca. Repita esto para cada gotita de lavado. No transferencia de ovocitos fragmentadas o transparente15.
Nota: Es importante transferir los ovocitos de cada gota en el plato de 35 mm con como poco HM como sea posible. Esto es cierto para cada transferencia en el protocolo. Los ovocitos MII no deben permanecer en la hialuronidasa que contiene medio de HM para más de un minuto.
3. SM-pescado coloración de ovocitos
- Fijar los ovocitos en un pozo individual de una placa de 6 pozos conteniendo 500 μl de tampón de fijación. Sumergir 20 ovocitos o menos en el pozo. Incubar por 20 min a temperatura ambiente.
Nota: Cada paso de tinción SM-pescado ocurre dentro de un pozo individual en una placa de 6 pozos cónico. Asegúrese de que los ovocitos son totalmente sumergidas en tampones y no flotante en la parte superior del búfer. Cada paso debe ser realizado con 20 oocitos o menos en cada pozo. - Transferencia de ovocitos fijadas a 500 μL de tampón de lavado (descrito en el paso 1.6 WB) durante 10 minutos. Repetir 2 veces más.
- Incubar ovocitos en buffer de permeabilización durante 30 min a temperatura ambiente.
Nota: El buffer de permeabilización se describe en el paso 1.7 reemplaza el buffer de permeabilización de la propiedad.- Se reúnen conjuntos de sonda y girar rápidamente hacia abajo en una microcentrífuga. Caliente cada sonda de 10 min en un baño de agua de 40 ° C o incubadora. Enfríe a la temperatura ambiente.
Nota: Este paso debe realizarse durante la incubación de permeabilización
- Se reúnen conjuntos de sonda y girar rápidamente hacia abajo en una microcentrífuga. Caliente cada sonda de 10 min en un baño de agua de 40 ° C o incubadora. Enfríe a la temperatura ambiente.
- Lavado de ovocitos en 500 μl de WB por 10 min a temperatura ambiente.
- Transfiera los ovocitos a 80 μL de proteasa III tampón (disponible en el kit), que es diluida 1:8 en 1 PBS X durante 30 min a temperatura ambiente.
Nota: El volumen 80 μl cubre adecuadamente el fondo de un pozo individual en una placa de 6 pozos. - Lavado de ovocitos en 500 μl de WB por 10 min a temperatura ambiente.
- Diluir los sonda calentada conjuntos de Nanog, Pou5f1 y DapB (un gen de control negativo), 1:50 en diluyente de sonda. Incubar ovocitos en 80 μL de la sonda de transcripción específicos durante 2 horas a 40° C.
Nota: Cada conjunto de sonda patentada está disponible en uno de los tres canales de fluorescencia (C1, C2 y C3). Las sondas de Nanog y Pou5f1 fueron marcadas con C2 y C3, respectivamente. - Caliente el propietario, 1 amplificador (1 AMP), amplificador 2 (AMP2), amplificador 3 (AMP3) y amplificador 4-fluorescencia (FL 4 AMP) a temperatura ambiente.
Nota: Este paso debe realizarse durante la incubación de 2 horas transcripción-específico sonda. - Transfiera los ovocitos a 500 μL de WB e incubar 10 min a temperatura ambiente.
- Incubar ovocitos secuencialmente en buffers de amplificación.
- Incubar ovocitos en 80 μl de AMP1 durante 30 min a ovocitos de transferencia de 40° C. a 500 μl de WB por 10 min a temperatura ambiente.
- Incubar ovocitos en 80 μl de AMP2 por 15 min a 40 ° C. Transfiera los ovocitos a 500 μl de WB por 10 min a temperatura ambiente.
- Incubar los ovocitos en 80 μl de AMP3 durante 30 min a 40 ° C. Transfiera los ovocitos a 500 μl de WB por 10 min a temperatura ambiente.
Nota: El resto del Protocolo se realiza en la oscuridad porque AMP-FL contiene el fluoróforo. Cuando se trabaja con el microscopio de disección, reduce la luz tanto como sea posible. - Añadir ovocitos a 80 μL de AMP4-FL durante 15 min a 40° C.
Nota: AMP4-FL se ofrece como alternativa buffer-A Alt-B (Alt-A), o Alt-C. Seleccione AMP4-FL búfer dependiente sobre qué emisión de longitud de onda es deseado.
- Lavado de ovocitos en 500 μl de WB por 10 min a temperatura ambiente. Incubar ovocitos en 80 μl de DAPI por 20 min a temperatura ambiente. Lavado de ovocitos en 500 μl de WB por 5 min a temperatura ambiente.
- Pipeta 12 μl antidecoloración del medio de montaje en el centro de un portaobjetos sin añadir burbujas al reactivo. Transferencia de ovocitos con como poco WB como sea posible en los medios de montaje y se aplica un cubreobjetos.
- El cubreobjetos en un ángulo de inclinación y lenta y suavemente sobre el líquido en la diapositiva. Evite presionar demasiado evitar la distorsión de los ovocitos y la introducción de burbujas el cubreobjetos.
- Almacenar las diapositivas en una oscura caja seca durante la noche a temperatura ambiente. Cubra los bordes de las diapositivas en pulimento de clavo claro para sellar el cubreobjetos.
- Utilizar un microscopio estándar para encontrar ovocitos en la diapositiva y el círculo con un marcador permanente.
Nota: Este paso no es necesario pero mejora localizar ovocitos en la diapositiva. Para mejores resultados, diapositivas de imagen dentro de 1 a 5 días como la señal fluorescente comenzará a desvanecerse.
4. tratamiento de la imagen
- Imagen de los ovocitos 3 dimensiones, usando microscopia confocal de paso z.
Nota: Para analizar con precisión las imágenes, cada paso de z debe ser 1,0 μm/rebanada. - Guardar imágenes confocales como un nd2 comprimido o individuales. Archivos TIFF para cada ovocito. Ambos tipos de imagen son compatibles con el programa de procesamiento de imágenes de código abierto, Fiji.
- Descargar e instalar el software de Fiji de acceso abierto (https://imagej.net/Fiji/Downloads).
- Arrastre archivos de nd2 en Fiji y elija hyperstack. Si imágenes confocales fueron guardados como. Archivos TIFF salte al paso 4.4.
Nota: Cuando se cae el archivo nd2 en Fiji el desplegable hyperstack aparecerá automáticamente. - Haga clic en la ficha imagen , seleccione Colory haga clic en Dividir canales para separar los canales fluorescentes del archivo nd2.
- Generar individuales. Archivos TIFF para cada rodaja z de los ovocitos en cada canal fluorescente. Haga clic en la ficha imagen , seleccione pilasy pila de imágenes, haga clic en. Click en la imagen , seleccione tipoy haga clic en Color RGB para convertir cada rodaja z en una imagen de color RGB individual.
Nota: El color RGB es artificial y puede ser elegido como deseado para cada longitud de onda de emisión. - Guarda cada imagen se puede convertir como. Archivo TIFF. Colocar imágenes de un ovocito único para cada canal fluorescente en una nueva carpeta para evitar confusiones durante la costura (paso 4.3).
- Arrastre archivos de nd2 en Fiji y elija hyperstack. Si imágenes confocales fueron guardados como. Archivos TIFF salte al paso 4.4.
- Normalizar cada uno. Imagen TIFF de Pou5f1 y Nanog utilizando imágenes de control negativo (DapB).
Nota: La normalización se realiza mediante un programa de retoque fotográfico. Asegúrese de eliminar los mismos niveles de fluorescencia del fondo de cada imagen de control. - Abierto cada uno normalizado. Archivo TIFF en Fiji para unir todos los z-sectores para cada ovocito en cada longitud de onda.
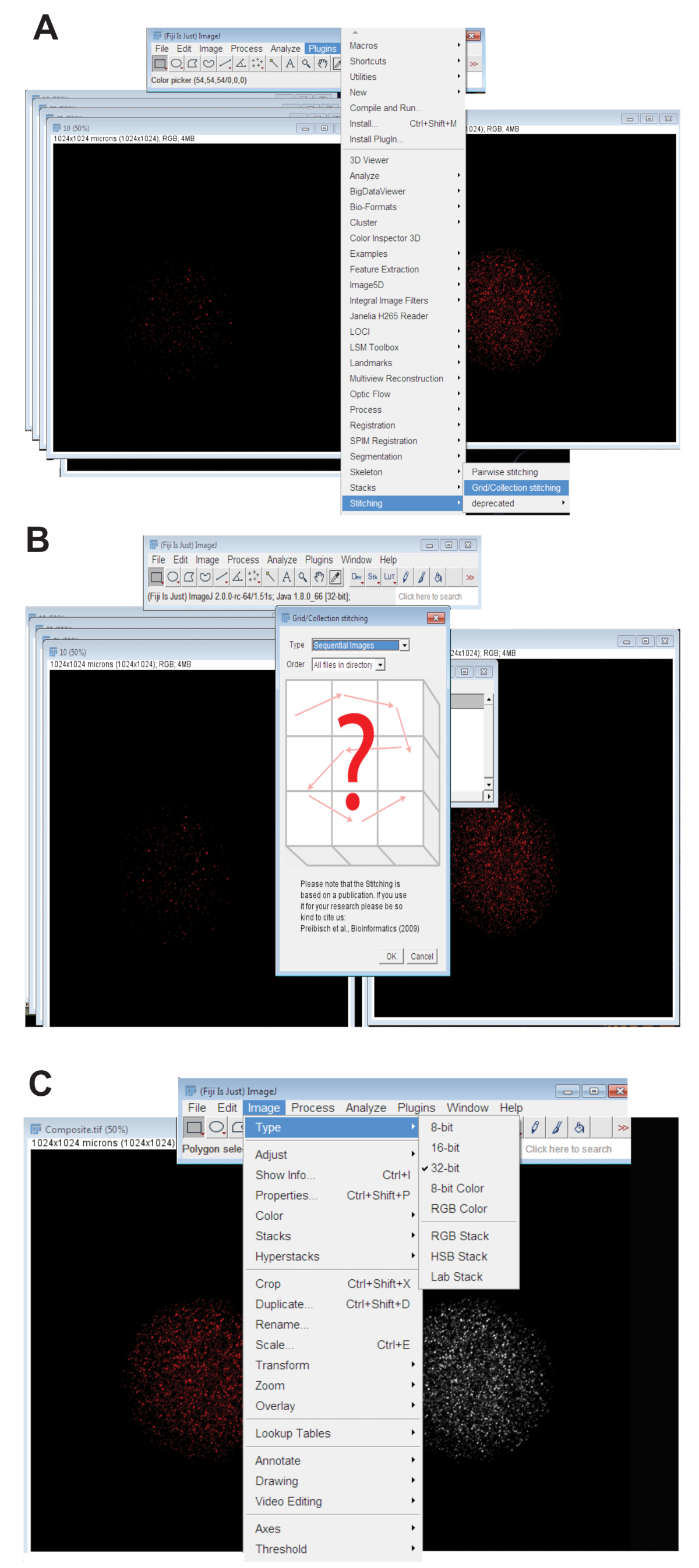
- Haga clic en la pestaña de Plugins , seleccione costuray haga clic en Colección de rejilla (Figura 3A). Seleccione Imágenes secuenciales en el menú desplegable y haga clic en aceptar (figura 3B).
- Navegar por el directorio y seleccione la carpeta que contiene todas las imágenes de rodaja z para un ovocito individual en una longitud de onda (ver paso 4.3.4). Haga clic en Aceptar.
- Mueva el control deslizante en la parte inferior de la imagen cosida en el canal de color apropiado para la longitud de onda utilizada y crear la imagen final de cosido de RGB haciendo clic en la imagen, seleccionando el tipo, haga clic en Color RGB.
Nota: Esta imagen se utilizará para la cuantificación de la fluorescencia que se describe en el paso 4.6 abajo.
- Convertir la imagen cosida a la imagen proyectada máxima de 32-bit. Haga clic en la imagen, seleccione el tipoy haga clic en 32 bits (figura 3). Guardar esta imagen como una nueva. Archivo TIFF.

Figura 3 : Sutura de imágenes confocales de serie z de los ovocitos. (A) captura de pantalla que muestra la herramienta de plug-in red/colección de Fiji que se usó para producir las imágenes compuestas de los ovocitos. (B) imágenes secuenciales utiliza fluorescencia se solapan alternativamente. Archivos TIFF para generar una imagen compuesta. (C) la imagen compuesta fue guarda como un 32-bit. Archivo TIFF. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
- Descargar e instalar el programa seguimiento y encontrar punto13, que está disponible en el sitio web D.R. Larson, un investigador en el nacional institutos de Salud Instituto Nacional del cáncer (https://ccr.cancer.gov/Laboratory-of-Receptor-Biology-and-Gene-Expression/daniel-r-larson). Descargar e instalar la máquina virtual de libre acceso para el sistema de operativo de lenguaje (IDL) de datos interactivo que es necesaria para ejecutar el encontrar lugar y programa de seguimiento (http://www.spacewx.com/pdf/idlvm.pdf).
- Abre la imagen de 32 bits, cosida, que se generó en el paso 4.6 (Figura 4A), en el programa de seguimiento y encontrar lugar. Seleccione el desplegable Localize y Localize (Figura 4B), que a calcular el número puntos encontrados la imagen, haga clic en.
Nota: Cada punto contado representa un mRNA individuales. Ajustes de banda paso y fotón umbral se muestran en la pantalla (Figura 4B). De este protocolo, fue utilizado por defecto para cada ajuste de umbral. Representante de positivo y los puntos de fondo se muestran (figura 4).

Figura 4 : Cuantificación de los mRNAs con buscador de Spot y seguimiento. (A) serie z Individual imágenes suturan como se describe en la figura 3 y guardados como un máximo de 32-bit proyectado. Archivo TIFF. (B) imagen compuesta fue inaugurada en buscador Spot y seguimiento. Localizar fue utilizado para contar las manchas fluorescentes (caja roja). Band pass y fotón umbral están indicadas por la caja azul. (C) la flecha azul señala una señal positiva (por encima del umbral). La flecha blanca muestra un fluorescente punto por debajo del umbral y, por lo tanto, no cuenta. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Resultados
Al finalizar el protocolo, el resultado es imágenes individuales de la serie z confocal (Figura 4A y figura 5), cosido de imágenes (figura 4), y mRNA cuenta (Figura 4B). Cuando se realiza la multiplexación, también habrá combinados imágenes que muestran la etiqueta de dos mRNAs diferentes (
Access restricted. Please log in or start a trial to view this content.
Discusión
Una serie de pasos pequeños durante el protocolo asegurará de fluorescencia exitosa y precisa de mRNAs. En primer lugar, el protocolo debe realizarse inmediatamente después de recogida y fijación de los ovocitos. Tenga en cuenta que PVP se ha agregado el buffer de fijación de paraformaldehído al 4% para evitar que los ovocitos se pegue uno al otro. Encontramos que es necesario realizar el experimento inmediatamente después de la recogida y fijación de los ovocitos. Cualquier retraso se traduce en una mucho menor ...
Access restricted. Please log in or start a trial to view this content.
Divulgaciones
Los autores no tienen nada que declarar
Agradecimientos
Agradecemos a Dr. Daniel R. Larson por su generosa ayuda con la instalación y uso del programa de seguimiento y encontrar lugar 13 y el apoyo técnico de la Universidad de Nebraska Lincoln microscopia base para la proyección de imagen de microscopía confocal. Este estudio representa una contribución de la Universidad de la división de investigación agrícola de Nebraska, Lincoln, Nebraska y fue apoyado por fondos de la portilla de UNL (NEB-26-206/número de-232435 y NEB-26-231/número de-1013511).
Access restricted. Please log in or start a trial to view this content.
Materiales
| Name | Company | Catalog Number | Comments |
| (±)-α-Lipoic acid | Sigma-Aldrich | T1395 | Alpha Lipoic Acid |
| Albumin, Bovine Serum, Low Fatty Acid | MP Biomedicals, LLC | 199899 | FAF BSA |
| BD 10mL TB Syringe | Becton, Dickinson and Company | 309659 | 10 mL syringe |
| BD PrecisionGlide Needle | Becton, Dickinson and Company | 305109 | 27 1/2 gauge needle |
| Calcium chloride dihydrate | Sigma-Aldrich | C7902 | CaCl2-2H2O |
| Citric acid | Sigma-Aldrich | C2404 | Citrate |
| D-(+)-Glucose | Sigma-Aldrich | G6152 | Glucose |
| Disodium phosphate | Na2HPO4 | ||
| Easy Grip Petri Dish | Falcon Corning | 351008 | 35 mm dish |
| Edetate Disodium | Avantor | 8994-01 | EDTA |
| Extra Fine Bonn Scissors | Fine Science Tools | 14084-08 | Straight, Sharp/Sharp, non-serrated, 13mm cutting edge scissors |
| Fetal Bovine Serum | Atlanta biologicals | S10250 | FBS |
| Gentamicin Reagent Solution | gibco | 15710-064 | Gentamicin |
| GlutaMAX-I (100X) | gibco | 35050-061 | Glutamax |
| Gold Seal Micro Slides | Gold Seal | 3039 | 25 x 75mm slides |
| Gonadotropin, From Pregnant Mares' Serum | Sigma | G4877 | eCG |
| hCG recombinant | NHPP | AFP8456A | hCG |
| Hyaluronidase, Type IV-S: From Bovine Testes | Sigma-Aldrich | H3884 | Hyaluronidase |
| Jewelers Style Forceps | Integra | 17-305X | Forceps 4-3/8", Style 5F, Straight, Micro Fine Jaw |
| L-(+)-Lactic Acid, free acid | MP Biomedicals, LLC | 190228 | L-Lactate |
| Magnesium sulfate heptahydrate | Sigma-Aldrich | M2773 | MgSO4-7H2O |
| MEM Nonessential Amino Acids | Corning | 25-025-Cl | NEAA |
| Microscope Cover Glass | Fisher Scientific | 12-542-C | 25 x 25x 0.15 mm cover slips |
| Mm-Nanog-O2-C2 RNAscope Probe | Advanced Cell Diagnostics | 501891-C2 | Nanog Probe |
| Mm-Pou5f1-O1-C3 RNAscope Probe | Advanced Cell Diagnostics | 501611-C3 | Pou5f1 Probe |
| MOPS | Sigma-Aldrich | M3183 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | Paraformaldehyde |
| PES 0.22 um Membrane -sterile | Millex-GP | SLGP033RS | 0.22 um filters |
| Polyvinylpyrrolidone | Sigma-Aldrich | P0930 | PVP |
| Potassium chloride | Sigma-Aldrich | 60128 | KCl |
| Potassium phosphate monobasic | Sigma-Aldrich | 60218 | KH2PO4 |
| Prolong Gold antifade reagent | invitrogen | P36934 | Antifade reagent without DAPI |
| RNAscope DAPI | Advanced Cell Diagnostics | 320858 | DAPI |
| RNAscope FL AMP 1 | Advanced Cell Diagnostics | 320852 | Amplifier 1 |
| RNAscope FL AMP 2 | Advanced Cell Diagnostics | 320853 | Amplifier 2 |
| RNAscope FL AMP 3 | Advanced Cell Diagnostics | 320854 | Amplifier 3 |
| RNAscope FL AMP 4 ALT A | Advanced Cell Diagnostics | 320855 | Amplifier 4 ALT A |
| RNAscope FL AMP 4 ALT B | Advanced Cell Diagnostics | 320856 | Amplifier 4 ALT B |
| RNAscope FL AMP 4 ALT C | Advanced Cell Diagnostics | 320857 | Amplifier 4 ALT C |
| RNAscope Fluorescent Multiplex Detection Reagents Kit | Advanced Cell Diagnostics | 320851 | FISH Reagent Kit |
| RNAscope Probe 3-plex Negative Control Probe | Advanced Cell Diagnostics | 320871 | Negative Control |
| RNAscope Probe 3-plex Positive Control | Advanced Cell Diagnostics | 320881 | Positive Control |
| RNAscope Probe Diluent | Advanced Cell Diagnostics | 300041 | Probe Diluent |
| RNAscope Protease III | Advanced Cell Diagnositics | 322337 | Protease III |
| RNAscope Protease III & IV Reagent Kit | Advanced Cell Diagnostics | 322340 | FISH Protease Kit |
| RNAscope Protease IV | Advanced Cell Diagnostics | 322336 | Protease IV |
| S/S Needle with Luer Hub 30G | Component Supply Co. | NE-301PL-50 | blunt 30 gauge needle |
| Sodium bicarbonate | Sigma-Aldrich | S6297 | NaHCO3 |
| Sodium chloride | Sigma-Aldrich | S6191 | NaCl |
| Sodium hydroxide | Sigma-Aldrich | 306576 | NaOH |
| Sodium pyruvate, >= 99% | Sigma-Aldrich | P5280 | Pyruvate |
| Solution 6 Well Dish | Agtechinc | D18 | 6 well dish |
| Taurine | Sigma-Aldrich | T8691 | Taurine |
| Tissue Culture Dish | Falcon Corning | 353002 | 60 mm dish |
| Triton X-100 | Sigma-Aldrich | X100 | Triton X-100 |
Referencias
- Vogelstein, B., Kinzler, K. W. Digital PCR. Proceedings of the National Academy of Sciences. 96 (16), 9236-9241 (1999).
- MacK, E. M., Smith, J. E., Kurz, S. G., Wood, J. R. CAMP-dependent regulation of ovulatory response genes is amplified by IGF1 due to synergistic effects on Akt phosphorylation and NF-kB transcription factors. Reproduction. 144 (5), 595-602 (2012).
- Xie, F., Timme, K. A., Wood, J. R. Using Single Molecule mRNA Fluorescent in Situ Hybridization (RNA-FISH) to Quantify mRNAs in Individual Murine Oocytes and Embryos. Scientific Reports. 8 (1), 7930(2018).
- Ruebel, M. L., et al. Obesity modulates inflammation and lipidmetabolism oocyte gene expression: A single-cell transcriptome perspective. Journal of Clinical Endocrinology and Metabolism. 102 (6), 2029-2038 (2017).
- Borensztein, M., Syx, L., Servant, N., Heard, E. Mouse Oocyte Development. 1818, 51-65 (2018).
- Jansova, D., Tetkova, A., Koncicka, M., Kubelka, M., Susor, A. Localization of RNA and translation in the mammalian oocyte and embryo. PLoS ONE. 13 (3), 1-25 (2018).
- Player, A. N., Shen, L. P., Kenny, D., Antao, V. P., Kolberg, J. A. Single-copy gene detection using branched DNA (bDNA) in situ hybridization. Journal of Histochemistry and Cytochemistry. 49 (5), 603-611 (2001).
- Wang, F., et al. RNAscope: A novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. Journal of Molecular Diagnostics. 14 (1), 22-29 (2012).
- Itzkovitz, S., van Oudenaarden, A. Validating transcripts with probes and imaging technology. Nature Methods. 8 (4), S12-S19 (2011).
- Derti, A., et al. ProbeDesigner: for the design of probesets for branched DNA (bDNA) signal amplification assays. Bioinformatics. 15 (5), 348-355 (1999).
- Larson, B., Malayter, D., Shure, M. Multiplexed Detection of Cytokine Cancer Biomarkers using Fluorescence RNA In Situ Hybridization and Cellular Imaging. BioTek Application Notes. , https://www.biotek.com/resources/application-notes/multiplexed-detection-of-cytokine-cancer-biomarkers-using-fluorescence-rna-in-situ-hybridization-and-cellular-imaging/ 1-5 (2016).
- Buxbaum, A. R., Wu, B., Singer, R. H. Single β-Actin mRNA Detection in Neurons Reveals a Mechanism for Regulating Its Translatability. Science. 343 (6169), 419-422 (2014).
- Thompson, R. E., Larson, D. R., Webb, W. W. Precise nanometer localization analysis for individual fluorescent probes. Biophysical Journal. 82 (5), 2775-2783 (2002).
- Herrick, J. R., Paik, T., Strauss, K. J., Schoolcraft, W. B., Krisher, R. L. Building a better mouse embryo assay: effects of mouse strain and in vitro maturation on sensitivity to contaminants of the culture environment. Journal of Assisted Reproduction and Genetics. 33 (2), 237-245 (2016).
- Pohlmeier, W. E., Xie, F., Kurz, S. G., Lu, N., Wood, J. R. Progressive obesity alters the steroidogenic response to ovulatory stimulation and increases the abundance of mRNAs stored in the ovulated oocyte. Molecular Reproduction and Development. 81 (8), 735-747 (2014).
- Xie, F., Anderson, C. L., Timme, K. R., Kurz, S. G., Fernando, S. C., Wood, J. R. Obesity-dependent increases in oocyte mRNAs are associated with increases in proinflammatory signaling and gut microbial abundance of lachnospiraceae in female mice. Endocrinology. 157 (4), 1630-1643 (2016).
- Hirao, Y., Yanagimachi, R. Detrimental effect of visible light on meiosis of mammalian eggs in vitro. Journal of Experimental Zoology. , (1978).
- Takenaka, M., Horiuchi, T., Yanagimachi, R. Effects of light on development of mammalian zygotes. Proceedings of the National Academy of Sciences. 104 (36), 14293-14293 (2007).
- Komminoth, P., Werner, M. Target and signal amplification: Approaches to increase the sensitivity of in situ hybridization. Histochemistry and Cell Biology. 108 (4-5), 325-333 (1997).
- Hornick, J. E., Duncan, F. E., Shea, L. D., Woodruff, T. K. Multiple follicle culture supports primary follicle growth through paracrine-acting signals. Reproduction. 145 (1), 19-32 (2013).
- Vlasova-St. Louis, I., Bohjanen, P. Feedback Regulation of Kinase Signaling Pathways by AREs and GREs. Cells. 5 (1), 4(2016).
- Gilbert, C., Svejstrup, J. Q. RNA Immunoprecipitation for Determining RNA-Protein Associations In Vivo. Current Protocols in Molecular Biology. 75 (1), 27.4.1-27.4.11 (2006).
- Kwon, S., Chin, K., Nederlof, M., Gray, J. W. Quantitative, in situ analysis of mRNAs and proteins with subcellular resolution. Scientific Reports. 7 (1), 16459(2017).
- Voigt, F., et al. Single-Molecule Quantification of Translation-Dependent Association of mRNAs with the Endoplasmic Reticulum. Cell Reports. 21 (13), 3740-3753 (2017).
- Halstead, J. M., Wilbertz, J. H., Wippich, F., Lionnet, T., Ephrussi, A., Chao, J. A. TRICK: A Single-Molecule Method for Imaging the First Round of Translation in Living Cells and Animals. Methods in Enzymology. 572, Elsevier Inc. (2016).
- Cookson, W., Liang, L., Abecasis, G., Moffatt, M., Lathrop, M. Mapping complex disease traits with global gene expression. Nature Reviews Genetics. 10 (3), 184-194 (2009).
- Houle, D., Govindaraju, D. R., Omholt, S. Phenomics: the next challenge. Nature Reviews Genetics. 11, 855(2010).
- Freimer, N., Sabatti, C. The Human Phenome Project. Nature Genetics. 34, 15(2003).
Access restricted. Please log in or start a trial to view this content.
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados