
Method Article
Programa integral de autopsias para individuos con esclerosis múltiple
* Estos autores han contribuido por igual
En este artículo
Resumen
La esclerosis múltiple es una enfermedad desmielinante inflamatoria sin cura. El análisis del tejido cerebral proporciona pistas importantes para entender la patogénesis de la enfermedad. Aquí discutimos la metodología y el análisis posterior del tejido cerebral de la ESCLEROSIs recogido a través de un programa único de autopsia rápida en funcionamiento en la Clínica Cleveland.
Resumen
Describimos un programa de donación rápida de tejidos para personas con esclerosis múltiple (ESCLEROSIs múltiple) que requiere que los científicos y técnicos estén de guardia las 24 horas del día, los 7 días de la semana, los 365 días del año. Los participantes consienten en donar su cerebro y la médula espinal. La mayoría de los pacientes fueron seguidos por neurólogos en el Cleveland Clinic Mellen Center for MS Treatment and Research. Sus cursos clínicos y discapacidades neurológicas están bien caracterizados. Poco después de la muerte, el cuerpo es transportado al Centro de Imágenes de MS, donde el cerebro es escaneado in situ por imágenes de resonancia magnética de 3 T (RM). El cuerpo se transfiere a la sala de autopsias, donde se extraen el cerebro y la médula espinal. El cerebro se divide en dos hemisferios. Un hemisferio se coloca inmediatamente en una caja de corte y las rodajas alternas de 1 cm de espesor se fijan en 4% de paraformaldehído durante dos días o se congelan rápidamente en hielo seco y 2-metilbutano. Las rodajas de cerebro de corte corto fijo se almacenan en una solución criopreservación y se utilizan para análisis histológicos y detección inmunocitoquímica de antígenos sensibles. Las rodajas congeladas se almacenan a -80 oC y se utilizan para estudios moleculares, inmunocitoquímicos e in situ de hibridación/ámbito de ARN. El otro hemisferio se coloca en 4% de paraformaldehído durante varios meses, se coloca en la caja de corte, se vuelve a escanear en el escáner de resonancia magnética de 3 T (MR) y se corta en rodajas de centímetro de espesor. Las imágenes de RM in situ (RM) postmortem se registran en conjunto con rodajas cerebrales de 1 cm de espesor para facilitar las correlaciones RMN-patología. Todas las rebanadas cerebrales son fotografiadas y se identifican lesiones cerebrales de materia blanca. La médula espinal se corta en segmentos de 2 cm. Los segmentos alternativos se fijan en un 4% de paraformaldehyde o se congelan rápidamente. La rápida adquisición de los tejidos de EM postmortem permite análisis patológicos y moleculares de los cerebros y las médulas espinales de la EM y correlaciones patológicas de las anomalías de la RMN cerebral. La calidad de estos tejidos postmortem procesados rápidamente (generalmente dentro de 6 h de la muerte) es de gran valor para la investigación de la EM y ha dado lugar a muchos descubrimientos de alto impacto.
Introducción
Una de las mejores maneras de estudiar una enfermedad es examinar el propio tejido enfermo. Esto presenta desafíos para aquellos que estudian enfermedades del sistema nervioso central (SNC). Las biopsias del cerebro y la médula espinal enfermos son extremadamente raras y generalmente implican casos atípicos. Las tasas de autopsia para individuos con enfermedades del SNC han disminuido drásticamente en los últimos años, y cuando se realizan, a menudo no proporcionan una rápida adquisición de tejidos. Estos desafíos han dado lugar al establecimiento de bancos cerebrales centrados en la enfermedad, incluyendo varios centrados en la recolección de tejidos de individuos con esclerosis múltiple (EP). La EM es una enfermedad mediada por inflamación del SNC que destruye la mielina, los oligodendrocitos (células formadoras de mielina), las neuronas y los axones. La mayoría de los pacientes con EM tienen un curso de enfermedad bifásica que comienza con brotes de discapacidad neurológica con recuperación variable que eventualmente evoluciona hacia una enfermedad gradualmente progresiva que es probablemente neurodegenerativa en la naturaleza1. Para la mayoría de los cerebros donados de EM, los intervalos de postmortem (PMI) entre la muerte y el procesamiento de tejidos superan las 24 h. Si bien estos tejidos han proporcionado información valiosa sobre los cambios patológicos en los cerebros de la EM, no son adecuados para estudios moleculares más avanzados que pueden proporcionar información poderosa sobre la fisiopatología de la enfermedad. Este es particularmente el caso de los estudios de perfilado de genes, que requieren ARN intacto.
Para superar las limitaciones discutidas anteriormente, hemos desarrollado un programa de donación rápida de tejidos que permite la RMN/correlaciones patológicas. Este protocolo proporciona tejidos bien conservados adecuados para estudios moleculares modernos y permite la comparación directa de la patología cerebral y las anomalías de la RMN en los cerebros de la EM. El Programa de Donación de Tejido de Esclerosis Múltiple de Cleveland Clinic ha existido por más de 20 años. Este programa de donación rápida de tejidos adquiere cerebros y médula espinal de individuos con EM y otras afecciones neurológicas autoinmunes asociadas. El programa tiene como objetivo obtener resonancias magnéticas in situ dentro de las 6 horas de la muerte, seguido de la eliminación del cerebro y la médula espinal para el procesamiento de tejido.
Reclutamiento
Las donaciones se obtienen a través del consentimiento ante mortem obtenido directamente de pacientes (pre-consentidos) o de parientes cercanos después de la muerte. Los pacientes pre-consentidos son típicamente identificados de la población clínica en el Centro Mellen para el Tratamiento e Investigación de la Esclerosis Múltiple en Cleveland, Ohio. Aunque la preferencia en el reclutamiento en el programa de donación rápida de tejidos se da a los pacientes que han sido seguidos en estudios de investigación longitudinal, está abierto a todos los pacientes vistos en el centro. Los participantes que se inscriban antes de la muerte reciben instrucciones para que los miembros de la familia o los proveedores de atención médica se comuniquen con el equipo de investigación, ya sea en el momento de la muerte o cuando se cree que la muerte es inminente. El segundo método para que las personas ingresen al programa de donación de tejidoes es en el momento de la muerte a través del consentimiento de sus parientes. El estado de Ohio requiere que las muertes sean referidas a una organización de adquisición de órganos de mandato federal llamada LifeBanc, que opera en 20 condados del noreste de Ohio. LifeBanc analiza todas las muertes para un diagnóstico de EM, que es una exclusión para la donación de órganos. Se hicieron arreglos para que LifeBanc notificara a los investigadores del programa de donación de tejido de la MS para todas las muertes con un diagnóstico asociado de LA EM que ocurren dentro de un radio de 75 millas de la Clínica Cleveland. El siguiente personal de los parientes y del hospital es contactado por el personal del Programa de Donación de Tejidos y se obtiene el consentimiento para la donación de cerebro y tejidos de la médula espinal. Estos dos métodos de reclutamiento ante-mortem y post-mortem a través de LifeBanc resultan en aproximadamente 10-12 donaciones de cerebro sin año. Los ajustes se realizan al límite de edad superior de la muerte para gestionar el número de referencias derivadas de LifeBanc.
Adquisición de donaciones
El programa requiere cobertura de 24 horas, 365 días al año por los miembros del programa de donación de tejidos para la adquisición de tejidos. El equipo clínico que cubre las donaciones de tejidos utiliza un sistema centralizado de notificación de donación de tejidos por correo electrónico/localizador/dispositivo móvil. LifeBanc recibe números para contactar al personal de guardia para el programa de donación de tejidos. Los miembros son notificados de la muerte por parte de los proveedores de hospitales/próximos de parientes (pre-consentidos) o por LifeBanc y otras fuentes de referencia. En primer lugar, se hace una determinación del tiempo de muerte y la viabilidad de la donación de tejidos. Las muertes se examinan en busca de condiciones que potencialmente resulten en tejido de mala calidad, incluyendo hipoxia premortem prolongada, tejido cerebral destructivo masivo (por ejemplo, hemorragia intracraneal grande, accidentes cerebrovasculares bihemisféricos extensos, tumor extenso apoyo prolongado del respirador (>3 días) y uso prolongado de agentes vasoactivos (>3 días) antes de la muerte. Cuando un médico forense está involucrado en una muerte, el neurólogo del estudio puede hablar con el médico forense para explorar una manera de recibir tejidos oportunos sin comprometer la responsabilidad del médico forense. Si se considera que el tejido viable está presente, se obtiene el consentimiento por escrito (si no se obtiene antes de la autopsia) y se hacen preparaciones para el transporte corporal. A continuación, se contacta con un servicio de transporte de difuntos previamente contratado para su transporte a las instalaciones de resonancia magnética de Cleveland Clinic. Se tiene cuidado de asegurarse de que el cuerpo permanece a temperatura ambiente y no se coloca en la refrigeración, ya que las temperaturas corporales más bajas se asocian con alteraciones en las características de la señal de RMN.
Historia clínica
La historia clínica incluye detalles sobre el diagnóstico de EM, la aparición de síntomas, tratamientos utilizados, resultados de pruebas clínicas y paraclínicas (potenciales evocados, resultados del líquido cefalorraquídeo, tomografía de coherencia óptica), esclerosis múltiple compuesto funcional , y la escala ampliada del estado de discapacidad (EDSS; real o estimada), que se recogen del expediente médico (cuando esté disponible), y la entrevista directa de parientes cercanos. También se recogen resonancias magnéticas premortem.
Protocolo
Este protocolo ha sido aprobado por la Junta de Revisión Institucional de Cleveland Clinic y sigue las directrices del comité de ética de investigación humana de Cleveland Clinic.
1. Rmn in situ
- Lleve el cuerpo del donante a la sala de resonancia magnética y lleve a cabo un protocolo de imágenes por RMN de 2 horas en MS Imaging Facility. Realice una resonancia magnética en un imager de 3 T o 7 T.
NOTA: La prioridad se da a 3T ya que la mayoría de los datos heredados se han realizado en 3 T, pero cuando no están disponibles, la creación de imágenes se lleva a cabo con 7T. Se realizan secuencias básicas designadas para todos los casos (Tabla1) y se realizan secuencias adicionales que dependen de los intereses actuales de investigación si el tiempo lo permite (limitado por el logro de la fijación tisular menos de 12 h después de la muerte). La Tabla 1 describe las secuencias principales.
2. Autopsia
NOTA: Después de la resonancia magnética in situ, el cuerpo es transportado a la morgue para la extracción de cerebro y médula espinal por un troquel y procesamiento de tejido por los miembros del laboratorio.
- Realice los siguientes pasos antes de la llegada del cuerpo a la morgue. Dos horas antes, preparar 3 L de 4% de paraformaldehida (PFA) y etiquetar envases y bolsas para el almacenamiento de tejidos. Preparar 3 L de 8% PFA y diluir 1.5 L de 8% PFA a 4% PFA. Colocar el 8% restante de PFA en 4 oC para el día 2.
- Antes de viajar a la morgue, llene 2 refrigeradores de viaje a 50% de capacidad con hielo seco (bloques grandes rotos para caber y pequeños pellets).
- En la morgue, llene un recipiente de acero inoxidable a la mitad con 2-metilbutano y hielo seco y cubra con una tapa en preparación para congelar el tejido.
- Pesar y fotografiar el cerebro una vez eliminado por el diener.
- Coloque cualquier dura adjunta en un recipiente lleno de PFA.
- Separe el cerebelo y el tronco encefálico del cerebro y fotografíe el cerebro.
- Identifique los nervios ópticos, el quiasmo y las vías y separe con una sonda y pinzas. Reseque la estructura con un bisturí.
NOTA: El segmento distal de un lado del nervio óptico se marca utilizando tinta Higgins para su identificación. - Separar los hemisferios cerebrales longitudinalmente y fotografiar cada hemisferio individualmente.
- Tintar la corteza motora primaria (PMC) para el hemisferio izquierdo, volver a fotografiarla, y colocarla en un contenedor de 3.3 L para una fijación prolongada. Documente la hora de inicio para la fijación cerebral.
- El PMC para el hemisferio derecho puede ser entintado o extirpado.
- Si se extirpa, primero retire las meninges que cubren.
- Volver a fotografiar PMC con tinta o extirpado.
- Si se extirpa PMC, se corta en 6 secciones de igual tamaño.
- Tinta el aspecto rostral de cada sección.
- Coloque secciones impares en contenedores llenos de PFA para una fijación corta.
- Congele las secciones con números uniformes y colóquelas en bolsas de congelador selladas en #1 más frías.
- Cortar el hemisferio derecho anterior a posterior en secciones coronales de 1 cm de espesor.
- Documentar anomalías graves (p. ej., artefacto de corte, hemorragia y lesiones).
- Coloque secciones impares en contenedores llenos de PFA para una fijación corta.
- Cierre las secciones con números uniformes y colóquelas en bolsas de congelador selladas.
- Documento fin del tiempo de fijación cerebral.
- Separe el tronco encefálico del cerebelo y colóquelo en un recipiente lleno de PFA para una fijación corta.
- Separar los hemisferios cerebelosos longitudinalmente.
- Cortar cada hemisferio en 4 secciones sagital igualmente gruesas.
- Fotografía vistas mediales y laterales.
- Coloque las rodajas hemisféricas cerebelosas izquierdas en un recipiente lleno de PFA para una fijación corta.
- Congele las rodajas hemisféricas cemisféricas derechas y colóquelas en bolsas de congelador selladas en un #1 más frío.
- Obtener la médula espinal con raíces nerviosas del diener.
- Retire la médula espinal dura mater y almacene la dura en un recipiente con PFA.
- Separar las raíces nerviosas izquierda y derecha anterior y posterior. Cortar las raíces nerviosas izquierdas anteriores y posteriores cortadas de la médula espinal y colocar en un recipiente lleno de PFA para la fijación corta.
- Cortar las raíces nerviosas derechas anteriores y posteriores de la médula espinal, congelar a presión, colocar en bolsas de congelador selladas, y luego colocar en #2 más frío.
- Fotografiar los 20 cm más caudales de la médula espinal. Documente la ubicación de la agrandación lumbar.
- Corte secciones transversales de 2 cm del cordón que proceden de caudal a rostral.
- Tintar el aspecto rostral de cada sección cortada.
- Coloque secciones impares en contenedores llenos de PFA para una fijación corta.
- Congele las secciones con números uniformes, colóquelas en bolsas de congelador selladas y luego colóquelas en #2 más frías.
- Documente la hora de inicio de la fijación de la médula espinal y cualquier anomalía grave.
- Fotografíe la porción rostral restante de la médula espinal.
- Documentar la posición del agrandamiento cervical. Siga los pasos 2.13.5–2.13.8 para la médula espinal restante.
- Después de la morgue coloque el tejido congelado en cajas etiquetadas en congeladores de -80 oC. Conservar el tejido fijo a 4oC.
- A 24 h después de la autopsia (día 2) diluir el 8% restante PFA a 4%.
- Reemplace el 4% de PFA en recipientes de fijación por un 4% PFA recién diluido.
- A 60 h post-autopsia preparar soluciones de 2.5% glutaraldehído en 4% PFA de glutaraldehído, PFA, dH2O y tampón de Sorenson (preparado por mezcla en secuencia: 0.2 M tampón de fosfato pH 7.4, polivinilpiridona 1% w/v, sacarosa 30% w/v, y etilenglicol 30% v/v).
- Retire el 4% de PFA usado de los contenedores de fijación corta.
- Enjuague el tejido en el tampón de Sorenson y colóquelo en una solución de crioprotección (glicerol 20%, tampón de 0,4 M Sorenson 20% y 0,02% de azida sódica en dH2O).
- Fotografíe las rebanadas de cerebro de corto fijo, el cerebelo, el tronco encefálico y la corteza motora (si corresponde).
- Con una hoja de bisturí cortada secciones transversales de 2 mm de espesor de cada sección de cuerda espinal corta fija de 2 cm.
- Colocar secciones en viales de centelleo de 2 ml y llenar con una solución de 2,5% de glutaraldehyde en 4% de PFA.
- Vuelva a colocar la sección restante en el vial de centelleo original de 20 ml. Enjuague la sección con el tampón de Sorenson y sustitúyala con solución de crioprotección.
3. Patología
NOTA: Las rodajas fijas cortas del hemisferio derecho, así como el hemisferio izquierdo fijo durante mucho tiempo (colocados en 4% de PFA durante varios meses) se cortan en secciones de 30 m (denominadas de flotación libre) o se incrustan en la parafina y se cortan como secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m (denominadas secciones de 12-14 m parafina incrustada). Estas secciones se procesan generalmente con proteína proteolípido (PLP) para detectar lesiones desmielinizantes y complejo de histocompatibilidad principal II (MHC-II) para la actividad inmune utilizando el método de diaminobenzidina (DAB). Estos protocolos han sido estandarizadosy utilizados en varias publicaciones 2,3,4,5,6,7,8,9 , 10 , 11 , 12.
-
Tinción de tejido del complejo DAB-Avidin-Biotin (ABC) de flotación libre (30 m)
- Retire las secciones de la solución de crioalmacenamiento, transfiera las secciones a una placa de seis pocillos y lávelas 3 veces durante 5 min cada una en 2 ml de 1x de fosfato salino pH 7.0 (PBS). Cuando se transfiere al siguiente pozo en la placa de seis pocillos, tenga cuidado de no romper el tejido. Durante cada paso de lavado e incubación, coloque la placa de seis pocillos en una coctelera y deje que el tejido se agite suavemente.
NOTA: Las secciones de tejido incubadas en volúmenes más pequeños y en placas más grandes (es decir, placas de 12 y 24 pocillos) tienden a exhibir desgarro de superficies y bordes. - Realizar la recuperación de antígenos mediante secciones de microagitamiento en un vaso de precipitados de vidrio que contenga aproximadamente 30 ml de tampón de citrato de 10 mM (pH 6.0). Asegúrese de que los tejidos no se doblan manipulando con un pincel y secciones de microondas durante 2-3 minutos o hasta que el tampón de citrato comience a hervir. Deje que las secciones se enfríen a temperatura ambiente (20 min).
- Transfiera las secciones de vuelta a una placa de seis pocillos y lave las secciones 3x durante 5 min cada una en 2 ml de PBS/0.3% Triton X-100. Bloquear las peroxidasas endógenas incubando secciones en 2 ml de 3% H2O2/0.3% Triton X-100/PBS durante 30 min a temperatura ambiente (RT).
- Lave las secciones 3x durante 5 min cada una en 2 mL de PBS/0.3% Triton X-100. Secciones de bloque en 2 mL de 3% de suero normal de cabra/0,3% Tritón X-100/1x PBS durante 1 h a RT.
- Incubar secciones durante la noche a 5 días (dependiendo del anticuerpo) en anticuerpos primarios dirigidos contra la microglia y los epítopos de mielina para detectar inflamación (MHCII) y desmielinización (PLP) (ver la Tabla de Materiales)a 4oC.
NOTA: Asegúrese de que las secciones no estén plegadas al incubar en este paso o en los pasos posteriores, ya que esto dará lugar a que las áreas dentro de las secciones estén vacías de mancha. - Lavar las secciones 3x durante 5 min cada una en 2 mL de 1x PBS. A continuación, incubar secciones en anticuerpos biotinilados secundarios (ver la Tabla de Materiales)durante 1 h en RT. Preparar la solución del Complejo Avidin-Biotina (ABC) durante la incubación aproximadamente 45 minutos antes del siguiente paso de lavado para permitir la formación de complejos ABC.
- Lavar las secciones 3x durante 5 min cada una en 2 mL de 1x PBS. A continuación, incubar secciones en ABC durante 1 h a RT.
- Lave las secciones en 2 ml de 1x PBS tres veces durante 5 min cada una. Incubar secciones en DAB filtrado (2 ml/pozo/sección) que contengan H2O2 (1:500 dilución de 30% H2O2 en DAB) hasta que el color se desarrolle adecuadamente (3-8 min).
- Lavar las secciones 3x durante 5 min cada una en 2 mL de 1x PBS. Para mejorar la señal (opcional), el osmicato con un 0,04% de OsO4 (30 s).
- Lave las secciones tres veces durante 5 min cada una en 1x PBS. Individualmente, transfiera cada sección de la placa de seis pocillos a un recipiente pequeño lleno de 1PBS y coloque la sección de tejido en un portaobjetos de vidrio lo más plano posible.
- Levante suavemente el portaobjetos fuera del PBS mientras se asegura de que la sección del tejido sea lo más plana posible. Usando dos pinceles, aplanar y estirar suavemente el tejido en el portaobjetos y quitar el exceso de agua con una toalla de papel. Monte las secciones de tejido con glicerol (o un medio de montaje equivalente) y selle el cubreobjetos con esmalte de uñas transparente.
- Retire las secciones de la solución de crioalmacenamiento, transfiera las secciones a una placa de seis pocillos y lávelas 3 veces durante 5 min cada una en 2 ml de 1x de fosfato salino pH 7.0 (PBS). Cuando se transfiere al siguiente pozo en la placa de seis pocillos, tenga cuidado de no romper el tejido. Durante cada paso de lavado e incubación, coloque la placa de seis pocillos en una coctelera y deje que el tejido se agite suavemente.
4. Secciones Hemisféricas incrustadas en Parafina: Tinción DAB
- Derretir la parafina de los portaobjetos en un horno de 60 oC durante 5-10 min.
- Desparafina las secciones incubando en xileno 3x durante 5 min cada una.
- Rehidratar el tejido en etanol clasificado al 100% (2x para 5 min cada uno), 95% (2x para 5 min), 70% (2x para 5 min cada uno), y 50% (1x para 5 min cada uno). Cubra las diapositivas en PBS.
- Recuperación de antígenos mediante correderas microwaving en un vaso de precipitados de 10 mM de tampón de citrato (pH 6.0).
- Lave los portaobjetos en 1PBS (3x para 5 min cada uno) una vez enfriados a temperatura ambiente. Bloquear las peroxidasas endógenas incubando tejido en 3% H2O2/1% Triton-X 100/PBS durante 30 min.
- Lavar el tejido en 1pbS tres veces durante 5 minutos cada uno. Bloquee el tejido con un 6% de suero normal de cabra en PBS durante 1 h.
- Incubar secciones en anticuerpos primarios (ver la Tabla de Materiales)en PBS a temperatura ambiente (RT) durante la noche (máx. 20 h).
- Lave las secciones 3x durante 5 min cada una en 1x PBS. A continuación, incubar secciones en el anticuerpo secundario correspondiente (ver la Tabla de Materiales)en PBS durante 1 h en RT.
- Preparar la solución ABC aproximadamente 45 minutos antes del siguiente paso de lavado durante la incubación.
- Lavar las secciones 3x durante 5 min cada una en 1x PBS y luego incubar en ABC durante 1 h a RT.
- Lavar la sección 3x durante 5 min cada una en 1x PBS.
- Incubar secciones en poros filtrados (0,45 m de poro de filtro) DAB que contiene H2O2 (1:500 dilución de 30% H2O2 en DAB) hasta que el color se desarrolle adecuadamente (3-8 min).
- Secciones de lavado (3x 5 min) en 1x PBS. Para mejorar la señal, el osmicato con 0,04% detetróxido de osmio (OsO 4; aproximadamente 30 s).
- Secciones de lavado (3x 5min) en 1x PBS.
- Deshidratar tejido en una serie calificada de etanol 50% (1x 5min), 70% (1x 5min), 95% (2x 5min), 100% (2x 5 min), y 100% xileno (1x 5min). Deje que los xilenos se evaporense.
- Secciones de montaje con medios de montaje rápido en seco a base de tolueno. Se recomiendan formulaciones que contienen antioxidantes para prevenir el blanqueo de manchas. Retire el exceso de soporte de montaje de los bordes de la corredera con una maquinilla de afeitar para su posterior almacenamiento.
5. Correlaciones de RMN/Patología
NOTA: Para correlacionar la RMN con la patología, primero realizamos una resonancia magnética ex vivo del hemisferio cerebral intacto fijado durante mucho tiempo (paso 2.9 anterior) en una caja ajustable con marcadores visibles para RMN que indican ranuras de corte. Luego cortamos el cerebro y fotografiamos las rebanadas de 1 cm para permitir el registro automático de resonancias magnéticas in situ a las rebanadas individuales del cerebro. A continuación, podemos realizar análisis guiados por RMN, donde las regiones de interés (ROI) se identifican en la RMN para dirigir el análisis de tejidos. También podemos realizar análisis guiados por histopatología, donde se identifican los IRO en el tejido (por ejemplo, lesiones de materia blanca, materia blanca sin desmielinización, etc.) y luego se caracterizan por las medidas de RMN colocalizadas (Tabla1).
-
Identificación de LOS ROI basados en RMN
NOTA: En estudios anteriores, hemos identificado LOS IRO en materia blanca11,12,13,14,15 y materia gris16,17, 18. El ejemplo siguiente es para el análisis de materia blanca.- Segmento T2 lesiones hiperintensas en resonancias magnéticas in situ (a partir del paso 1.1), inicialmente procesado por un algoritmo automático, y luego corregido manualmente por usuarios experimentados.
- Segmento T1 lesiones hipointensas dentro de lesiones T2 como vóxeles con una intensidad de señal menor o igual al 80% de la intensidad de la señal del tejido cerebral de aparición normal circundante.
- Segmente las áreas hipointensas en los mapas de relación de transferencia de magnetización (MTR) con un umbral del 80%.
- Cree tres clasificaciones utilizando las segmentaciones anteriores: (a) lesiones sólo T2 que son anormales en las exploraciones Ponderadas en T2/FLAIR pero normales en las exploraciones Ponderadas En T1 o MTR, (b) lesiones T2T1MTR, que son anormales en todas las exploraciones Ponderadas en T2/FLAIR, T1 y MTR; y (c) materia blanca de apariencia normal (NAWM), que son normales en todos los escaneos ponderados en T2/FLAIR, T1 y MTR.
NOTA: Nuestra selección de sectores se basa en la existencia de los tres tipos de regiones de interés (solo T2, T2T1MTR y NAWM) en los mismos sectores. - Calcular intensidades normalizadas para cada ROI para minimizar la variabilidad derivada de diferentes cerebros y diferentes ubicaciones cerebrales.
-
Registro co-registro de resonancia magnética in situ en rebanadas cerebrales
- Escanee el hemisferio cerebral fijo en una caja de corte personalizada con cuatro filas de marcadores sensibles a la RMN que localizan las ranuras donde se puede insertar un cuchillo para cortar el cerebro. Realice una adquisición 3D MPRAGE ponderada en T1 con resolución isotrópica de 1 mm que cubra el cerebro fijo y todos los marcadores.
NOTA: Esto se denomina RMN posterior a la fijación y solo se utiliza como paso intermedio para el registro automático de las resonancias magnéticas in situ en las rebanadas cerebrales. - Inmediatamente después de escanear, cortar el hemisferio cerebral en ranuras situadas a 1 cm de distancia, lo que resulta en aproximadamente 15 rebanadas.
- Fotografiar las rebanadas cerebrales en los lados anterior y posterior.
- Co-registre la resonancia magnética in situ y fotografías de las rebanadas cerebrales con los siguientes pasos.
- Para la segmentación de la posfijación y el MPRAGE in situ, preprocesar tanto los MRI de post-fijación como los MMI MPRAGE in situ para la no uniformidad de intensidad19.
- Segmente el cerebro y cuatro filas de marcadores de MPRAGE preprocesados después de la fijación.
- Segmentar el hemisferio correspondiente a la resonancia magnética20posterior a la fijación,21 del MPRAGE in situ preprocesado.
- Co-registre los MRI in situ y posteriores a la fijación extraídos en el cerebro a través de una serie de procesos de registro lineal de hasta 12 grados de libertad (registro afín) utilizando FSL FLIRT22. Los componentes de escalado y cizallamiento representan el efecto de la contracción de la fijación.
- Encuentre la dirección normal del plano de corte minimizando la suma de las intensidades máximas proyectadas utilizando los marcadores segmentados. Estos ángulos de reorientación se incorporan en la matriz de transformación obtenida del paso anterior.
- Coincide visualmente con las imágenes de RMN con fotografías de cortes fijos del cerebro posterior utilizando un visor de imágenes interno que permite cambiar la profundidad y la orientación de los vectores normales. Se necesitan pequeñas modificaciones porque el corte cerebral es imperfecto.
- Aplique la transformación de imagen AFFINE13 para cada sector para transformar las resonancias magnéticas in situ en las mismas ubicaciones de corte que las fotografías de las rebanadas cerebrales.
- Para la segmentación de la posfijación y el MPRAGE in situ, preprocesar tanto los MRI de post-fijación como los MMI MPRAGE in situ para la no uniformidad de intensidad19.
- Escanee el hemisferio cerebral fijo en una caja de corte personalizada con cuatro filas de marcadores sensibles a la RMN que localizan las ranuras donde se puede insertar un cuchillo para cortar el cerebro. Realice una adquisición 3D MPRAGE ponderada en T1 con resolución isotrópica de 1 mm que cubra el cerebro fijo y todos los marcadores.
Resultados
Como se mencionó anteriormente, casi la mitad del hemisferio cerebral está congelado y disponible para estudios moleculares utilizando ADN, ARN, o proteína. Históricamente, se ha demostrado que los estudios con tejidos cerebrales postmortem se ven afectados por afecciones premortem, edad, sexo, pH tisular, integridad del ARNm (RIN), intervalo postmortem (PMI), certeza diagnóstica, uso de comorbidsustancias y tratamiento previo con medicamentos estado23. Según estudios que utilizan tejidos cerebrales, el ADN y la proteína parecen verse afectados en menor medida en comparación con el ARN. Sin embargo, según nuestra experiencia, se ha encontrado que el aislamiento de ARN y el análisis aguas abajo son los más afectados por las condiciones premortem y el intervalo postmortem del tejido cerebral. Por lo tanto, discutimos algunas de las condiciones a seguir para realizar análisis basados en ARN utilizando tejidos emordezcos postmortem.
Para todos nuestros estudios, después de que el cerebro se recoge en la autopsia, se corta en rodajas (1 cm de espesor) y luego se fija en 4% paraformaldehído para estudios morfológicos o se congela rápidamente para el análisis bioquímico. Todos los bloques de tejido se caracterizan por la desmielinización mediante inmunostainización utilizando PLP como se describió anteriormente. En la Figura 1se muestra un esquema de análisis representativo. Las secciones de tejido se examinan para detectar la presencia de lesiones de materia blanca (Figura1A). Las regiones seleccionadas se tiñen para la actividad inmunitaria (Figura1B)y la desmielinización (Figura 1C). El tejido congelado se monta en el criostato (Figura1D)y se cortan secciones congeladas de 30 m. Esto es seguido por la recolección de 3-4 secciones posteriores, la separación de los tejidos adyacentes, y el almacenamiento para el ADN, ARN, o aislamiento de proteínas. Utilizando este protocolo, hemos aislado con éxito ADN24,25,ARN5,6,7,8,9 así como proteínas26. Mientras que se discuten los principales hallazgos de algunos de los estudios que analizan el ARN de los cerebros de la EM, aquí están algunos de los temas relacionados con el análisis de los cerebros de la EM postmortem del ARN.

Figura 1: Colección de muestras para el análisis de ARNm. (A) El tejido de la autopsia se selecciona para el análisis. Se seleccionan áreas de tejido y se extirpada una porción de tejido. Todas las secciones están teñidas con (B) MHC-II (Major histocompatibility complex (MHC) clase II HLA-DR CR3/43) anticuerpo para detectar la actividad inflamatoria y con (C) proteína proteolídica (PLP) para determinar el estado de la mielina utilizando protocolos publicados. Según el estado de la mielina, el bloque se puntúa mediante un bisturí (D). Las secciones (60 m) se cortan (E) y las áreas que se han puntuado previamente se eliminan, se separan en tubos y se etiquetan (F). Las manchas PLP y MHC-II se repiten después de cada 5 secciones para garantizar una correcta recolección de tejido. Se observa materia blanca que aparece normal (NAWM) y las lesiones de materia blanca (WML) se describen en rojo. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
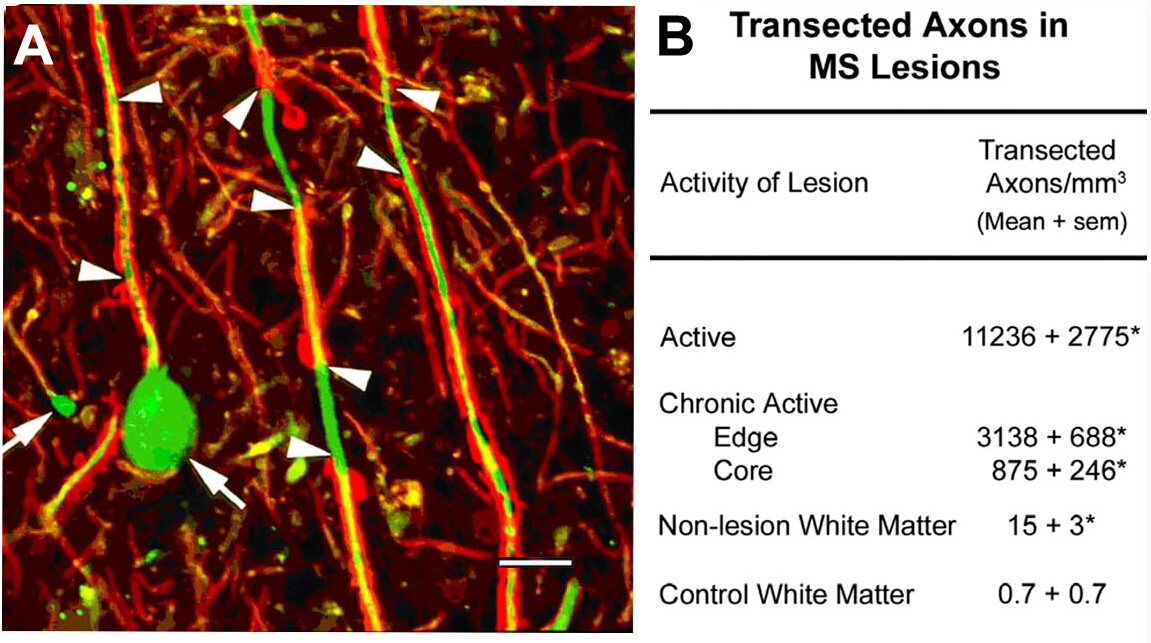
Transección axonal en las lesiones de la SM10. Un enfoque científico inicial de este programa fue la caracterización de los componentes celulares de las lesiones de sormielinizadas de la materia blanca. Entre los antígenos localizados había neurofilamentos no fosforilados (NF). La mayoría de los NF son fosforilados en axones mielinados. Tras la desmielinización, los axones se defosforilan. Detectamos la expresión esperada de NFs no fosforilados en axones desmielinizados. En las lesiones agudas de la EP, muchos de estos axones desmielinizados terminaron como bombillas de retracción axonal (Figura2A),que reflejan los extremos proximales de los axones transectos. Los axons transecicétos superan los 11.000 mm3 en las lesiones agudas en comparación con las regiones normales adyacentes10. Estas observaciones ayudaron a catalizar un cambio de paradigma en la investigación de la EP que movió el campo hacia la caracterización de la neurodegeneración como la principal causa de discapacidad neurológica permanente en individuos con EM.

Figura 2: Transección axonal durante la desmielinización inflamatoria. La transección axonal ocurre durante ladesmielinización inflamatoria (A , puntas de flecha) e induce la formación de ovoides axonales terminales (A , flechas). Cuando se cuantifican (B), los axónicos transectos son abundantes en las lesiones de la EP y parecen correlacionarse con la actividad inflamatoria de la lesión. Panel A reproducido de Trapp et al.10 con permiso. Rojo: proteína proteólídica, verde: Neurofilamento anti no fosforilado. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
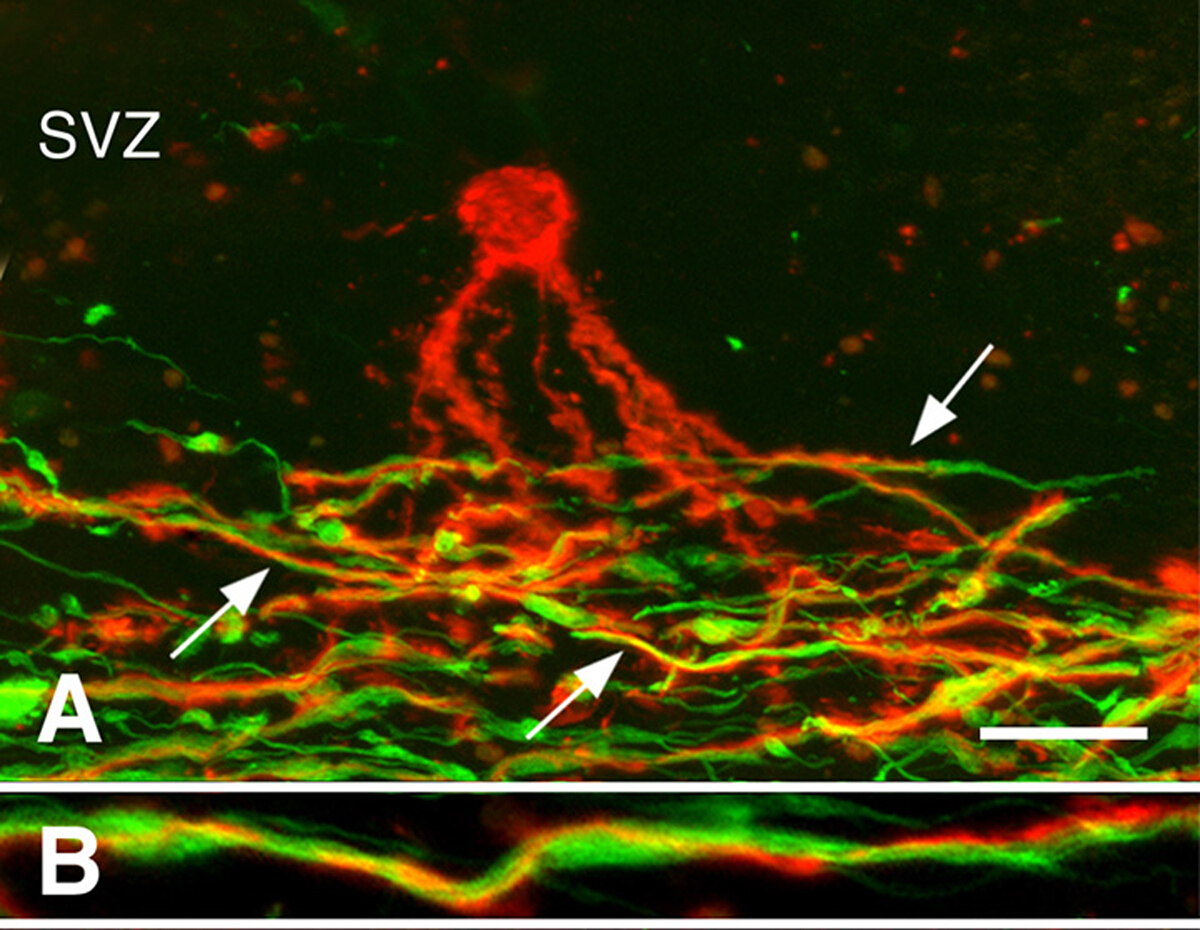
Remielinización en cerebroscrónicos de LAMS 3 . La remielinización puede ser robusta durante las primeras etapas de la MUS. Sin embargo, muchas lesiones crónicas de la EM no se remielinan. Investigamos si la presencia de células progenitoras de oligodendrocitos (OPO) o la generación de nuevos oligodendrocitos limita la remielinización de lesiones de materia blanca desmielinizadas crónicas. Mientras que la densidad de la OPC a menudo disminuyó, estaban presentes en todas las lesiones desmielinizadas crónicamente3. Los oligodendrocitos recién generados también estuvieron presentes en muchas lesiones crónicas de la EM. Procesos de oligodendrocitos asociados con, pero no mielinato axones desmielinizados (Figura 3). Estos estudios indican que los OPO y su capacidad para producir nuevos oligodendrocitos no están limitando la remielinización de lesiones crónicas de materia blanca. Hemos planteado la hipótesis de que los axones crónicos desmielinizados, que a menudo parecían distróficos, no eran receptivos a la remielinización por los oligodendrocitos recién producidos.

Figura 3: Procesos de pre-mielinización de oligodendrocitos asociados con axones. Se muestran micrografías confocales de lesiones de LA MS manchadas con anticuerpos PLP (rojo en los paneles A, B)y anticuerpos de neurofilamento (verdes en los paneles A, B). Un oligodendrocitos premielinizantes (rojo en el panel A) en la zona subventricular (SVZ) extendió los procesos a la región de los axones desmielinizados (verdes en el panel A) en una lesión crónica de EM. Muchos de estos procesos (flechas en el Panel A) se enespiralron alrededor de los axons, como se muestra en un aumento más alto (Panel B). Las barras de escala representan 20 m (A) y 5 m (B). Reproducido de Chang et al.3 con permiso. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Disfunción mitocondrial en MS6. Realizamos una búsqueda imparcial de los cambios genéticos neuronales en la corteza motora rápidamente congelada obtenida de pacientes con EM crónica (Figura4A). Una búsqueda imparcial de este conjunto de datos identificó reducciones significativas en 23 ARNM mitocondriales codificados nuclearmente en MS (Figura4B). Los estudios de acreditación utilizando inmunocitoquímica e hibridación in situ indicaron que estos genes estaban altamente enriquecidos en neuronas de proyección cortical (Figura4C)y que las mitocondrias aisladas de los axones de proyección muestran glucólisis reducida ( Figura 4D). Este documento catalizó un enfoque en la disfunción mitocondrial y redujo la producción de ATP como un importante contribuyente a la degeneración axonal en la EM.

Figura 4: Datos de microarray y técnicas de validación aguas abajo realizadas en la corteza motora de La Sra. (A) Agrupación jerárquica de transcripciones significativamente alteradas de muestras de corteza motora de control (C1-C6) y SPMS (MS1-MS6), apoyando por separado patrones de expresión génica relacionadas con enfermedades. Entre la disminución de las transcripciones en la corteza motora de la SM, veintiséis pertenecían a la cadena de transporte de electrones (B). El ARNm del complejo mitocondrial I (NDUFA6) se redujo en las neuronas (n a 55-130) en la corteza motora de la SM(CII)en comparación con el control (CI), mientras que las densidades del ARNm PLP fueron similares entre el control (CIII) y la corteza cerebral de la EM (CIV). La actividad de los complejos de transporte de electrones I y III se redujo en fracciones mitocondriales enriquecidas a partir de la corteza motora de los pacientes con SM (n.o 3) (D). 6 con permiso. Las barras de error representan SEM; La barra de escala en CI-IV es de 25 m. * p < 0.05 Prueba t-test de los estudiantes. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
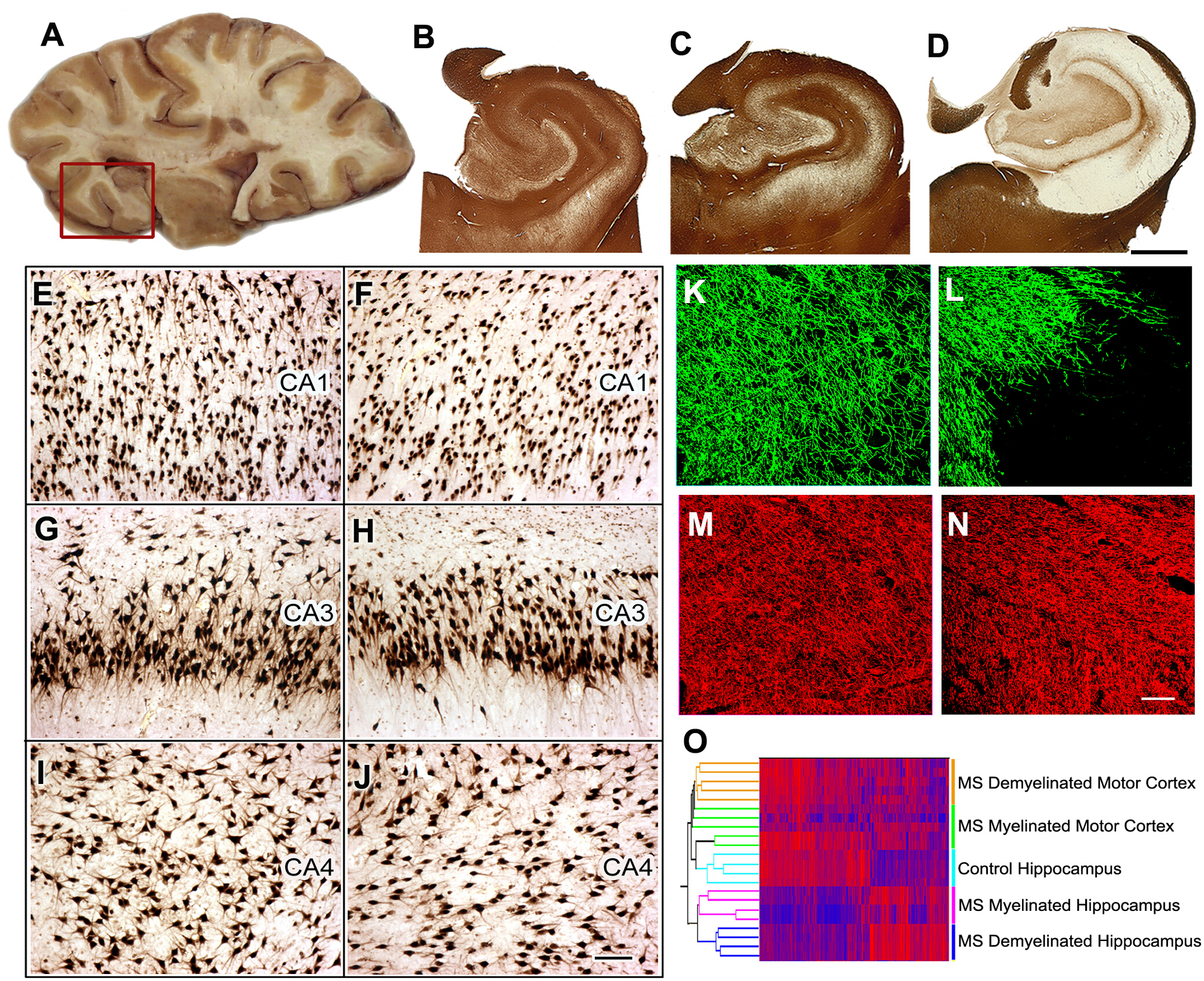
Patogénesis de la disfunción cognitiva en la SM8. Cuarenta y 60% de los pacientes con EM tienen deterioro cognitivo y reducción de la función ejecutiva. Identificamos el hipocampo, que es un sitio funcional de memoria/aprendizaje, como un sitio común para la desmielinización en la MI. A continuación comparamos la expresión génica neuronal en hipocampo mielinizado y desmielinizado y encontramos reducciones significativas en las ARN neuronales que codifican proteínas implicadas en la memoria/aprendizaje. Ampliamos estos datos demostrando que los microARN selectos se incrementan en el hipocampo desmielinizado y que estos microARN pueden disminuir la expresión de receptores de glutamato. Hemos reproducido y ampliado estas observaciones en modelos de roedores. A continuación comparamos la expresión génica neuronal en el hipocampo mielinizado y desmielinizado y encontramos reducciones significativas en las proteínas de codificación de ARNm neuronal implicadas en la memoria/aprendizaje.

Figura 5: Recopilación de tejidos, análisis histológico y estudios de expresión génica en el hipocampo de EM. Las rebanadas cerebrales que contienen hipocampo se seleccionan durante la autopsia (A) y se elimina el hipocampo y la región contigua (caja roja) para su análisis posterior. La inmunopreservación para PLP mostró la preservación de la mielina en todos los controles (B) y el 40% de la MAYOR hippocampi (C). Se detectó una desmielinización extensiva enel 60 % de la Mayor Hippocampi (D ). En comparación con el control del hipocampo (E, G, I), no se detectó una pérdida neuronal significativa en las regiones CA1, CA3 o CA4 de hipocampo desmielinizado (F, H, J) como se muestra en la inmunohistoquímica HuR. La inmunofluorescencia de doble etiqueta para mielina (proteína básica de mielina (MBP), verde) y axones (SMI32, rojo) mostró pérdida de mielina (L) con conservación relativa de axones (N) en el hipocampo desmielinizado de MS en comparación con el hipocampo de control ( MBP, K; SMI32, M). La agrupación dual de los niveles de expresión de ARNm organizó muestras en grupos discretos basados en el estado de mielina (mielinado y desmielinizado) y la ubicación (hippocampus vs corteza motora) (O). Los altos niveles de ARNm se indican con los niveles de expresión bajos de los niveles de expresión de color rojo y azul. 8 con permiso. B-D: 2 mm, E-J: 100 m, K-N: 50 m. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
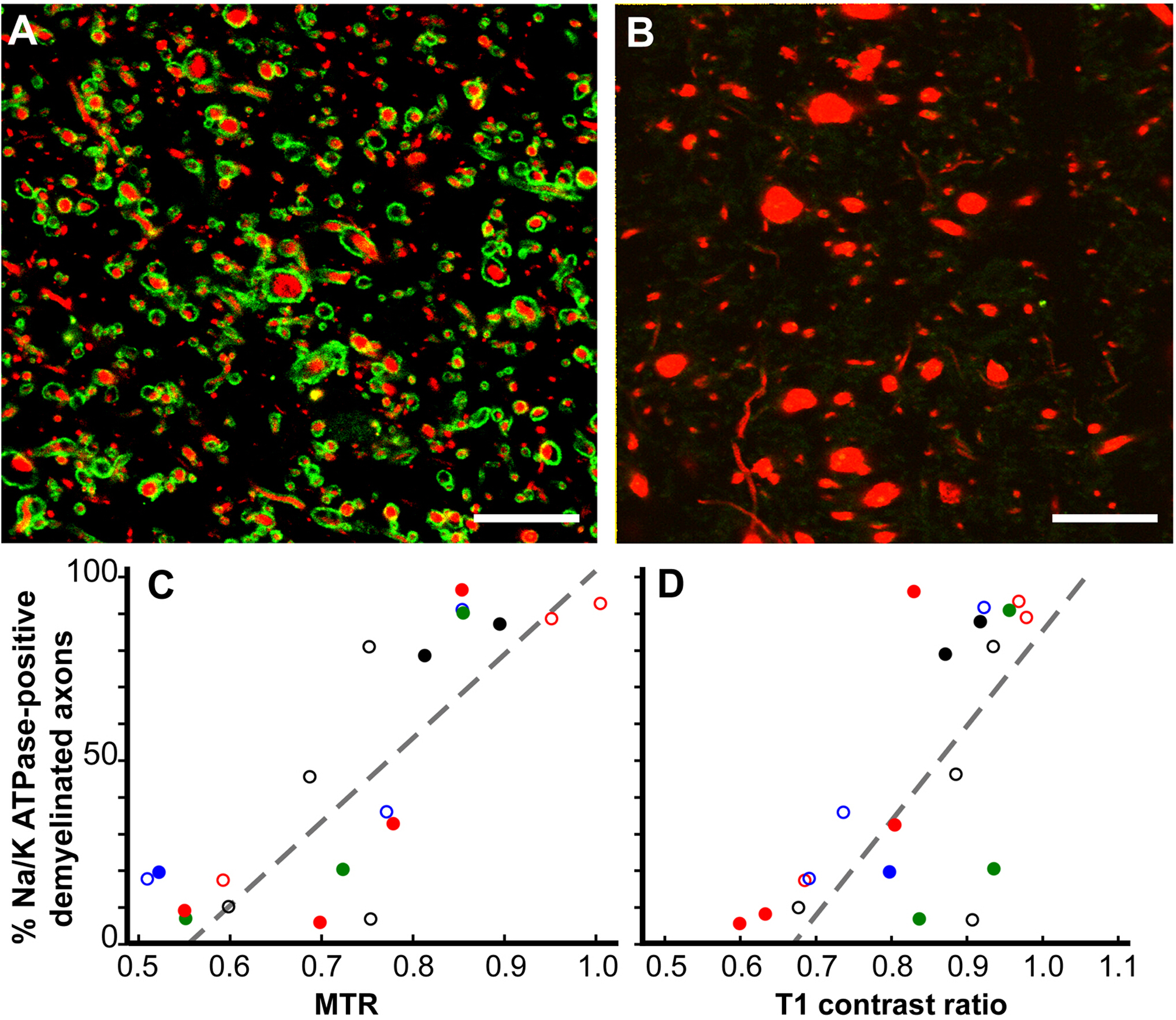
Correlaciones patológicas de cambios de RMN12. Si bien la RMN es un indicador valioso del diagnóstico de la EM y la respuesta al tratamiento, y también es un predictor de la progresión de la enfermedad de EM, los correlatos patológicos de los cambios en la RMN no se entienden bien. Nuestros estudios de resonancia magnética postmortem se han centrado en dos RIS de RMN. LOS ROI cerebrales de materia blanca que solo eran hiperintensos t2 (solo T2) y ROIs que tenían una combinación de hipointensidad T1, hiperintensidad T2 y relación de transferencia de magnetización reducida (MTR) (T2T1MTR). Aproximadamente el 45% de los ROI de materia blanca cerebral T2 fueron mielinizados, lo que confirma su naturaleza no específica. Por el contrario, el 83% de los ROC T2T1MTR fueron desmielinizados crónicamente y aparecieron como agujeros negros. Los valores T1 y MTR son semicuantitativos y sus valores variaron ampliamente en los ROI T2T1MTR. Si la pérdida de mielina es el único contribuyente a estos cambios de RMN, los valores deben ser constantes. Axones desmielinizados hinchados correlacionados con valores T1 y MTR.

Figura 6: Las relaciones de transferencia de magnetización (MTR) y las relaciones de contraste T1 se correlacionan linealmente con el porcentaje de axónicos Na+/K+ ATPase positivos en lesiones crónicas de LAM. Las lesiones desmielinizadas crónicamente manchadas para Na+/K+ ATPase (verde) variaron de casi 100% (A) a cero (B) en neurofilamento (rojo). Muchos axons sin Na+/K+ ATPase habían aumentado los diámetros (B). Comparación del porcentaje de axones Positivos Na+/K+ ATPase en lesiones de EM crónicamente desmielinizadas correlacionadas con MTR postmortem cuantitativo (p < 0.0001, C) y relaciones de contraste T1 (p < 0.0006, D). Cada punto de datos es de una sola lesión y cada combinación única color-símbolo denota uno de los cerebros estudiados. Barras de escalas de 5 m. Reproducidas de Young et al.12 con permiso. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Neurodegeneración independiente de la desmielinización11. Históricamente, se ha pensado que la neurodegeneración en la MUS es el resultado de la desmielinización. Sin embargo, los estudios por imágenes cerebrales han planteado la posibilidad de que la neurodegeneración y la desmielinización puedan ser eventos independientes. Recientemente identificamos una subpoblación de pacientes con EP que tienen desmielinización de la médula espinal y la corteza cerebral, pero no de la materia blanca cerebral. Acuñamos este subtipo de MS como MS mielocortical (MCMS). Los casos de MCMS proporcionaron una plataforma para investigar la relación entre la desmielinización de la materia blanca cerebral y la pérdida neuronal cortical. En comparación con el control de cortices, la pérdida neuronal cortical fue significativamente mayor en los cortices MCMS que en los cortices típicos de la SM. El tejido cerebral de control se obtuvo del Departamento de Patología de la Clínica Cleveland. Este estudio proporciona la primera evidencia patológica de neurodegeneración en ausencia de desmielinización.

Figura 7: Pérdida neuronal en ausencia de desmielinización cerebral de la materia blanca. Una sección hemisférica coronal teñida de cresilo violeta de un individuo clasificado como que tiene una EM típica (A). Las densidades neuronales se compararon en las capas corticales III, V y VI en cada una de las cinco áreas etiquetadas. Las neuronas con un área superior a 60 m2 (amarillo) se muestran en una imagen representativa de la corteza temporal superior (B ). Etiquetado para PLP y distribución de lesiones desmielinizadas (la desmielinización de materia blanca se resalta en azul; la desmielinización subpial se resalta en rosa) en secciones hemisféricas de individuos con SM típica (C) y MS mielocortical (D ) se muestran. Se encontró una correlación significativa entre la reducción de la densidad neuronal cortical y el aumento del volumen de lesiones de materia blanca cerebral en la SM típica, pero no en la SM mielocortical (E); líneas discontinuas indican un intervalo de confianza (CI) del 95%. IFG - gyrus frontal inferior. STG - giso temporal superior. INi inferior insula. Insula superior. CG - gyrus cingulado. Reproducido de Trapp et al.11 con permiso. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Duración de la secuencia | Descripción de la secuencia | Uso de secuencia |
| 0:09 | Localizador | Localización de secuencias posteriores |
| 9:14 | Magnetización 3D preparado eco de gradiente rápido (MPRAGE) | Estimación volumétrica por imágenes estructurales de las estructuras cerebrales |
| 5:14 | Recuperación de inversión atenuada por fluido 3D (FLAIR) | Identificación de lesiones Segmentación de lesiones Evaluación de lesiones volumétricas |
| 2:35 | 2D T2 ponderado | Identificación de lesiones Segmentación de lesiones Evaluación de lesiones volumétricas |
| 5:12 | Eco 3D recuperado por gradiente con pre-pulso de transferencia de magnetización , (MT-ON) | Medida supuesta del contenido de mielina en el tejido normal que aparece y la lesión |
| 5:12 | Eco 3D recuperado por gradiente sin prepulso de transferencia de magnetización , (MT-OFF) | |
| 0:27 | Asignación de campos de Diffusion Tensor Imaging (DTI) | Medición de la difusión del agua en el tejido cerebral que se cree que refleja la integridad del tejido cerebral. |
| 10:27 | Diffusion Tensor Imaging (DTI) multi-shell | |
| 1:18 | Diffusion Tensor Imaging (DTI) multi-shell | |
| 39:48:00 | SUBTOTAL: CORE |
Tabla 1: Protocolo de imágenes postmortem.
Discusión
Describimos un protocolo que se ha utilizado para adquirir y procesar rápidamente tejido de más de 150 individuos con EM. Una característica importante de este protocolo es que los científicos que utilizan el tejido también están a cargo de establecer el protocolo y realizar la recolección de tejido. Esto proporciona flexibilidad para satisfacer las necesidades científicas de los proyectos de investigación individuales. Varios aspectos de este protocolo mejoran su utilidad. Los pacientes suelen estar bien caracterizados antes de la muerte, ya que muchos de ellos han sido seguidos por neurólogos en nuestro centro. Un paso crítico es el procesamiento de donaciones de tejido poco después de la muerte, lo que aumenta la calidad del tejido congelado en comparación con algunos otros bancos cerebrales. Esto permite estudios moleculares que son de gran valor en la descripción de los cambios en los productos genéticos transcripcionales y traslacionales, que son esenciales para la fundamentación de observaciones histológicas e inmunocitoquímicas. El aprovechamiento de los datos morfológicos/inmunocitoquímicos y moleculares en múltiples casos mejora la fiabilidad de las conclusiones. Esto se ilustra mejor con nuestra descripción de los cambios del gen mitocondrial en la corteza cerebral y los cambios del gen neuronal en el hipocampo desmielinizado. Se están desarrollando nuevos protocolos de generación de perfiles genéticos a un ritmo rápido y el tejido congelado de nuestro banco debería proporcionar ARN de alta calidad para el análisis de tejidos y células monocelulares.
Otro aspecto valioso de nuestro protocolo son las rebanadas de cerebro cortas fijas. Estos tejidos se cortan en secciones de 30 m de espesor y flotación libre. Estas secciones son ideales para el uso de microscopía confocal para analizar dos o más antígenos en tres dimensiones. Algunos buenos ejemplos incluyen la interacción de procesos de oligodendrocitos premielinantes con axones distróficos en lesiones crónicas de EM, así como la identificación de conexiones axonales individuales a bombillas de retracción axonal transectadas. Esto contrasta con el uso rutinario de secciones de parafina de 7 m de espesor, donde las imágenes 3D no son factibles. Los tejidos incrustados en parafina tienen un gran valor para algunas preguntas, especialmente la cuantificación de las densidades neuronales en secciones hemisféricas de 7 m de espesor. Nuestros protocolos de procesamiento de tejidos, por lo tanto, son diversos y proporcionan flexibilidad para asegurar tejidos fijos y rápidamente congelados.
Otra característica única de nuestro protocolo es la resonancia magnética cerebral in situ postmortem. Las resonancias magnéticas cerebrales son un biomarcador insustituible de la enfermedad de la MI. Por lo tanto, es esencial establecer los correlatos patológicos de las señales de RMN anormales. Nuestros estudios establecieron que los ROI T2 y T2T1MTR a menudo son mielinados. Este hallazgo apoya la necesidad de modalidades de diagnóstico por imágenes más específicas que distingan de forma fiable entre la materia blanca cerebral mielinizada y desmielinizada. La RMN parece ser sensible para la detección de mielina, pero nuestros estudios demuestran que incluso una combinación de T1/T2/MTR no es específica para identificar la mielinización. Nuestro protocolo postmortem proporciona una plataforma ideal para probar la capacidad de nuevas modalidades de imagen para distinguir entre materia blanca cerebral mielinizada y desmielinizada. La RMN también proporciona el vehículo ideal para la traducción de los resultados científicos básicos a la práctica clínica, dado el uso de la RMN en nuestra investigación traslacional y su uso clínico generalizado en pacientes vivos.
Mientras que el corte corto y largo fijo, así como las rodajas congeladas ofrece una ventaja para el procesamiento de tejido en múltiples modos para diferentes estudios, hay algunas limitaciones con este método. La evaluación de la totalidad de una estructura puede limitarse, ya que las partes de ella pueden procesarse de manera diferente en sectores adyacentes. El gran volumen del banco de tejidos, sin embargo, ofrece la capacidad de investigar una estructura de interés en múltiples sujetos para mejorar el muestreo. Otra limitación general a los estudios que utilizan tejidos postmortem es que son transversales. Las conclusiones relativas al calendario y la progresión de los cambios deben interpretarse en este contexto. Puede haber un sesgo de selección para los pacientes que donan sus tejidos, lo que puede limitar la generalización de los datos a todos los pacientes con EM. Dado que la mayoría de los donantes mueren por complicaciones de la EM avanzada, puede que no sea apropiado extrapolar los hallazgos de estos pacientes a los que se encuentran en etapas tempranas de la EM. No obstante, hemos recibido tejidos de pacientes más jóvenes que murieron de afecciones no relacionadas con la EMM (es decir, infarto agudo de miocardio, sobredosis de drogas, suicidio). El alcance de nuestro protocolo no incluye el muestreo de otros órganos (por ejemplo, gastrointestinales y médula ósea) que han estado implicados en la EM. Creemos que las fortalezas del programa superan en gran medida sus limitaciones.
Divulgaciones
Los autores no declaran conflictos de intereses.
Agradecimientos
Los autores también quieren agradecer al Dr. Christopher Nelson por su asistencia editorial. El programa de autopsia es apoyado en parte por la concesión R35 NS097303 a BDT. El trabajo en el laboratorio de RD está respaldado por subvenciones de NINDS (NS096148) y la Sociedad Nacional de Esclerosis Múltiple, EE.UU. (RG 5298).
Materiales
| Name | Company | Catalog Number | Comments |
| Antibodies | |||
| Biotinylated goat anti-mouse IgG | Vector Laboratories | BA-9200 | 1:500 dilution for hemispheric; 1:1,000 for 30µm free-floating. RRID: AB_2336171 |
| Biotinylated goat anti-rabbit IgG | Vector Laboratories | BA-1000 | 1:500 dilution. |
| Biotinylated goat anti-rat IgG | Vector Laboratories | BA-9400 | 1:500 dilution for hemispheric; 1:1,000 for 30µm free-floating. RRID: AB_2336208 |
| Glial fibrillary acid protein (GFAP) | Dako | Z0334 | 1:700 dilution for hemispheric. RRID: AB_10013382 |
| HuR, mouse IgG, 3A2 clone | Santa Cruz | SC-5261 | 1:500 for 30µm free floating. RRID: AB_627770 |
| Major histocompatibility complex (MHC) class II HLA-DR CR3/43 | Dako | Mo746 | 1:250 dilution for hemispheric; 1:500 for 30 µm free floating. RRID: AB_2313661 |
| Non-phosphorylated neurofilament (SMI32) | Biolegend | 801701 | 1:5,000 dilution for hemispheric; 1:2,500 for 30 µm free-floating. RRID: AB_2564642 |
| Phosphorylated neurofilament (SMI31) | Biolegend | 801601 | 1:5,000 dilution for hemispheric; 1:2,500 for 30 µm free-floating. RRID: AB_2564641 |
| Proteolipid protein (PLP) | Gift from Wendy Macklin | 1:250 dilution for IHC; alternative anti-PLP antibodies commercially available. | |
| Reagents | |||
| 125 mm filter paper | Whatman | 1452-125 | For filtering PFA. |
| 50% Glutaraldehyde | Electron Microscopy Sciences | 16320 | Electron microscopy grade. |
| Cytoseal | ThermoScientific | 8310-16 | |
| Ethylene glycol | Fisher Chemical | BP230-4 | |
| Glycerol | Sigma-Aldrich | G7893 | 400 mL/2 L Cryoprotection solution. |
| Millex-HV Syringe Filter Unit, 0.45 µm, PVDF, 33 mm, gamma sterilized | Millipore-Sigma | SLHV033RB | |
| Paraformaldehyde | Electron Microscopy Sciences | 19200 | Prills form. |
| Polyvinylpyrolidone (PVP-40) | Fisher Chemical | BP220-212 | |
| Sodium azide | Fisher Chemical | S227I | 2 g/2 L Sorenson's buffer. |
| Sodium phosphate dibasic | Sigma-Aldrich | S0876 | 98.8 g/2 L Sorenson's buffer. |
| Sodium phosphate mono basic monohydrate | Sigma-Aldrich | S9638 | 14.352 g/2 L Sorenson's buffer. |
| Sucrose | Sigma-Aldrich | PVP40-500G | |
| VectaStain ABC Kit | Vector Laboratories | PK-6100 | 1:1,000 dilution of A and B. RRID: AB_2336819 |
| Waterproof drawing black ink | Higgins | 44201 | |
| Xylene | Fisher Chemical | X3S | Histological grade. |
| Equipment | |||
| 3T MRI Magnetom Prisma | Siemens Healthineers | ||
| 7T MRI Agilent 830AS | Siemens Healthineers |
Referencias
- Trapp, B. D., Nave, K. A. Multiple sclerosis: an immune or neurodegenerative disorder?. Annual Review of Neuroscience. 31, 247-269 (2008).
- Chang, A., Nishiyama, A., Peterson, J., Prineas, J., Trapp, B. D. NG2-positive oligodendrocyte progenitor cells in adult human brain and multiple sclerosis lesions. Journal of Neuroscience. 20, 6404-6412 (2000).
- Chang, A., Tourtellotte, W. W., Rudick, R., Trapp, B. D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. New England Journal of Medicine. 346, 165-173 (2002).
- Chang, A., et al. Neurogenesis in the chronic lesions of multiple sclerosis. Brain. 131, 2366-2375 (2008).
- Chang, A., et al. Cortical remyelination: A new target for repair therapies in multiple sclerosis. Annals of Neurology. 72, 918-926 (2012).
- Dutta, R., et al. Mitochondrial dysfunction as a cause of axonal degeneration in multiple sclerosis patients. Annals of Neurology. 59, 478-489 (2006).
- Dutta, R., et al. Activation of the ciliary neurotrophic factor (CNTF) signalling pathway in cortical neurons of multiple sclerosis patients. Brain. 130, 2566-2576 (2007).
- Dutta, R., et al. Demyelination causes synaptic alterations in hippocampi from multiple sclerosis patients. Annals of Neurology. 69, 445-454 (2011).
- Dutta, R., et al. Hippocampal demyelination and memory dysfunction are associated with increased levels of the neuronal microRNA miR-124 and reduced AMPA receptors. Annals of Neurology. 73, 637-645 (2013).
- Trapp, B. D., et al. Axonal transection in the lesions of multiple sclerosis. New England Journal of Medicine. 338, 278-285 (1998).
- Trapp, B. D., et al. Cortical neuronal densities and cerebral white matter demyelination in multiple sclerosis: a retrospective study. Lancet Neurology. 17, 870-884 (2018).
- Young, E. A., et al. Imaging correlates of decreased axonal Na+/K+ ATPase in chronic multiple sclerosis lesions. Annals of Neurology. 63, 428-435 (2008).
- Fisher, E., et al. Imaging correlates of axonal swelling in chronic multiple sclerosis brains. Annals of Neurology. 62, 219-228 (2007).
- Moll, N. M., et al. Imaging correlates of leukocyte accumulation and CXCR4/CXCL12 in multiple sclerosis. Archieves of Neurology. 66, 44-53 (2009).
- Moll, N. M., et al. Multiple sclerosis normal-appearing white matter: pathology-imaging correlations. Annals of Neurology. 70, 764-773 (2011).
- Nakamura, K., Chen, J. T., Ontaneda, D., Fox, R. J., Trapp, B. D. T1-/T2-weighted ratio differs in demyelinated cortex in multiple sclerosis. Annals of Neurology. 82, 635-639 (2017).
- Chen, J. T., et al. Clinically feasible MTR is sensitive to cortical demyelination in MS. Neurology. 80, 246-252 (2013).
- Nakamura, K., Fox, R., Fisher, E. CLADA: cortical longitudinal atrophy detection algorithm. Neuroimage. 54, 278-289 (2011).
- Sled, J. G., Zijdenbos, A. P., Evans, A. C. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Transactions of Medical imaging. 17, 87-97 (1998).
- Fisher, E., Cothren, J. R. M., Tkach, J. A., Masaryk, T. J., Cornhill, J. F. Knowledge-based 3D segmentation of the brain in MR images for quantitative multiple sclerosis lesion tracking. Proc. SPIE 3034, Medical Imaging. , 19-25 (1997).
- Avants, B. B., Epstein, C. L., Grossman, M., Gee, J. C. Symmetric diffeomorphic image registration with cross-correlation: evaluating automated labeling of elderly and neurodegenerative brain. Medical Image Analysis. 12, 26-41 (2008).
- Jenkinson, M., Bannister, P., Brady, M., Smith, S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage. 17, 825-841 (2002).
- Lewis, D. A. The human brain revisited: opportunities and challenges in postmortem studies of psychiatric disorders. Neuropsychopharmacology. 26, 143-154 (2002).
- Chomyk, A. M., et al. DNA methylation in demyelinated multiple sclerosis hippocampus. Scientific Reports. 7, 8696 (2017).
- Huynh, J. L., et al. Epigenome-wide differences in pathology-free regions of multiple sclerosis brains. Nature Neuroscience. , (2014).
- Ishii, A., et al. Human myelin proteome and comparative analysis with mouse myelin. Proceedings of the National Academy of Sciences. U. S. A. 106, 14605-14610 (2009).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados