
Method Article
Evaluación morfológica y funcional de los axónyis y sus sinapsis durante la muerte de Axon en Drosophila melanogaster
* Estos autores han contribuido por igual
En este artículo
Resumen
Aquí, proporcionamos protocolos para realizar tres ensayos simples de degeneración de axón inducida por lesiones (muerte por axón) en Drosophila melanogaster para evaluar la preservación morfológica y funcional de los axones cortados y sus sinapsis.
Resumen
La degeneración de Axón es una característica compartida en la enfermedad neurodegenerativa y cuando los sistemas nerviosos son desafiados por las fuerzas mecánicas o químicas. Sin embargo, nuestra comprensión de los mecanismos moleculares subyacentes a la degeneración del axón sigue siendo limitada. La degeneración de axón inducida por lesiones sirve como un modelo simple para estudiar cómo los axones cortados ejecutan su propio desmontaje (muerte de axón). En los últimos años, se ha identificado una cascada de señalización de muerte de axón evolutivamente conservada de moscas a mamíferos, que es necesaria para que el axón separado degenere después de una lesión. Por el contrario, la señalización de la muerte de axón atenuada da como resultado la preservación morfológica y funcional de los axones cortados y sus sinapsis. Aquí, presentamos tres protocolos simples y recientemente desarrollados que permiten la observación de la morfología axonal, o la función axonal y sináptica de los axones cortados que han sido cortados del cuerpo de las células neuronales, en la mosca de la fruta Drosophila. La morfología se puede observar en el ala, donde una lesión parcial resulta en muerte de axón lado a lado de axón de axón control de axones de control no lesionados dentro del mismo haz nervioso. Alternativamente, la morfología axonal también se puede observar en el cerebro, donde todo el haz nervioso sufre la muerte por axón desencadenada por la ablación antenal. La preservación funcional de los axones cortados y sus sinapsis puede evaluarse mediante un enfoque optogenético simple junto con un comportamiento de aseo postsináptico. Presentamos ejemplos utilizando una mutación de pérdida de función de hilos altos y por expresar excesivamente dnmnat, ambos capaces de retrasar la muerte de axón durante semanas o meses. Es importante destacar que estos protocolos se pueden utilizar más allá de las lesiones; facilitan la caracterización de los factores de mantenimiento neuronal, el transporte axonal y las mitocondrias axonales.
Introducción
La integridad morfológica de las neuronas es esencial para la función sostenida del sistema nervioso a lo largo de la vida. La gran mayoría del volumen neuronal es tomado por axons1,2; por lo tanto, el mantenimiento de por vida de axones particularmente largos es un gran desafío biológico y bioenergético para el sistema nervioso. Se han identificado múltiples mecanismos de apoyo axonal-intrínseco y glial-extrínseco, asegurando la supervivencia axonal de por vida. Su deterioro se traduce en degeneración de axón3, que es una característica común de los sistemas nerviosos que se desafian en la enfermedad, y por las fuerzas mecánicas o químicas4,5. Sin embargo, los mecanismos moleculares subyacentes de la degeneración del axón siguen siendo poco comprendidos en cualquier contexto, lo que dificulta el desarrollo de tratamientos eficaces para bloquear la pérdida de axón. El desarrollo de terapias eficaces contra estas condiciones neurológicas es importante, ya que crean una enorme carga en nuestra sociedad6.
La degeneración de axón inducida por lesiones sirve como un modelo simple para estudiar cómo los axones cortados ejecutan su propio desmontaje. Descubierto por Augustus Waller en 1850, la degeneración de Walleriano (WD) es un término paraguas que comprende dos procesos distintos, molecularmente separables7. En primer lugar, después de una lesión axonal, los axones separados de sus cuerpos celulares ejecutan activamente su propia autodestrucción (muerte de axón) a través de una cascada de señalización de muerte de axón evolutivamente conservada dentro de un día después de la lesión8. En segundo lugar, la glia circundante y los fagocitos especializados enganchan y limpian los desechos axonales resultantes en un plazo de tres a cinco días. La atenuación de la señalización de la muerte de axón da lugar a axón cortado que permanecen preservados durante las semanas9,,10,11,12, mientras que la atenuación del engullir glial culmina en escombros axonales que persiste durante semanas en vivo13,14,15.
La investigación en moscas, ratones, ratas y peces cebra reveló varios mediadores evolutivamente conservados y esenciales de la muerte de axón que indica8. Los mutantes de la muerte de Axon contienen axónes y sinapsis cortados que no pueden sufrir la muerte por axón; permanecen morfológicamente y funcionalmente preservados durante semanas, en ausencia de soporte corporal celular9,10,12,13,16,17,18,19,20,21,22,23. El descubrimiento y la caracterización de estos mediadores llevaron a la definición de una vía molecular que ejecutó la muerte de axón. Es importante destacar que la señalización de muerte axón se activa no sólo cuando el axón se corta, aplasta o estira24,25; también parece ser un contribuyente en distintos modelos animales de condiciones neurológicas (por ejemplo, cuando los axones degeneran de manera independiente de las lesiones4, pero con una gama de resultados beneficiosos4,8). Por lo tanto, entender cómo la muerte de axón ejecuta la degeneración de axón después de una lesión podría ofrecer información más allá de un modelo de lesión simple; también podría proporcionar dianas para la intervención terapéutica.
La mosca de la fruta Drosophila melanogaster (Drosophila) ha demostrado ser un sistema invaluable para la señalización de la muerte de axón. La investigación en la mosca reveló cuatro genes esenciales de la muerte del axón evolutivamente conservados: hilo alto (hiw)11,14, dnmnat12,26, dsarm10 y axundead (axed)12. La modificación de estos mediadores —mutaciones de pérdida de función de hiw, dsarm y axed,y sobreexpresión de dnmnat— bloquea poderosamente la muerte por axón durante toda la vida útil de la mosca. Mientras que los axons de tipo salvaje cortados sufren la muerte de axón dentro de 1 día, los axones cortados y sus sinapsis carentes de hiw, dsarm o hacha permanecen no sólo morfológicamente, sino también funcionalmente preservados durante semanas. Queda por determinar si la preservación funcional también puede lograrse a través de altos niveles de dnmnat.
Aquí, presentaremos tres protocolos simples y recientemente desarrollados para estudiar la muerte por axón (por ejemplo, la morfología y la función de los axones cortados y sus sinapsis a lo largo del tiempo) en ausencia de soporte corporal celular. Demostramos cómo la muerte de axón atenuada resulta en axónes cortados que se conservan morfológicamente con una mutación de pérdida de función(hiw-N)∆Ny cómo la muerte de axón atenuada resulta en axónes y sinapsis cortados que permanecen preservados funcionalmente durante al menos 7 días con la sobreexpresión de dnmnat (dnmnatOE). hiw Estos protocolos permiten la observación de la morfología axonal y sináptica individual, ya sea en el sistema nervioso central, o periférico (SNC y PNS, respectivamente)13,14, mientras que la preservación funcional de los axones cortados y sus sinapsis en el SNC se puede visualizar mediante el uso de una configuración optogenética simple combinada con el aseo como una lectura conductual12.
Protocolo
1. Observación de la Morfología de Axón durante la muerte de Axon en el PNS
- Lesión en las alas: lesión parcial de los paquetes de axón
- Utilice 5 hembras vírgenes y 5 machos del genotipo derecho(Figura 4A,generación P0) para realizar cruces a temperatura ambiente (RT). Pasar P0 en viales nuevos cada 3-4 días. Recoger la progenie adulta recién cerrada (generación F1) diariamente y envejecer durante 7-14 días.
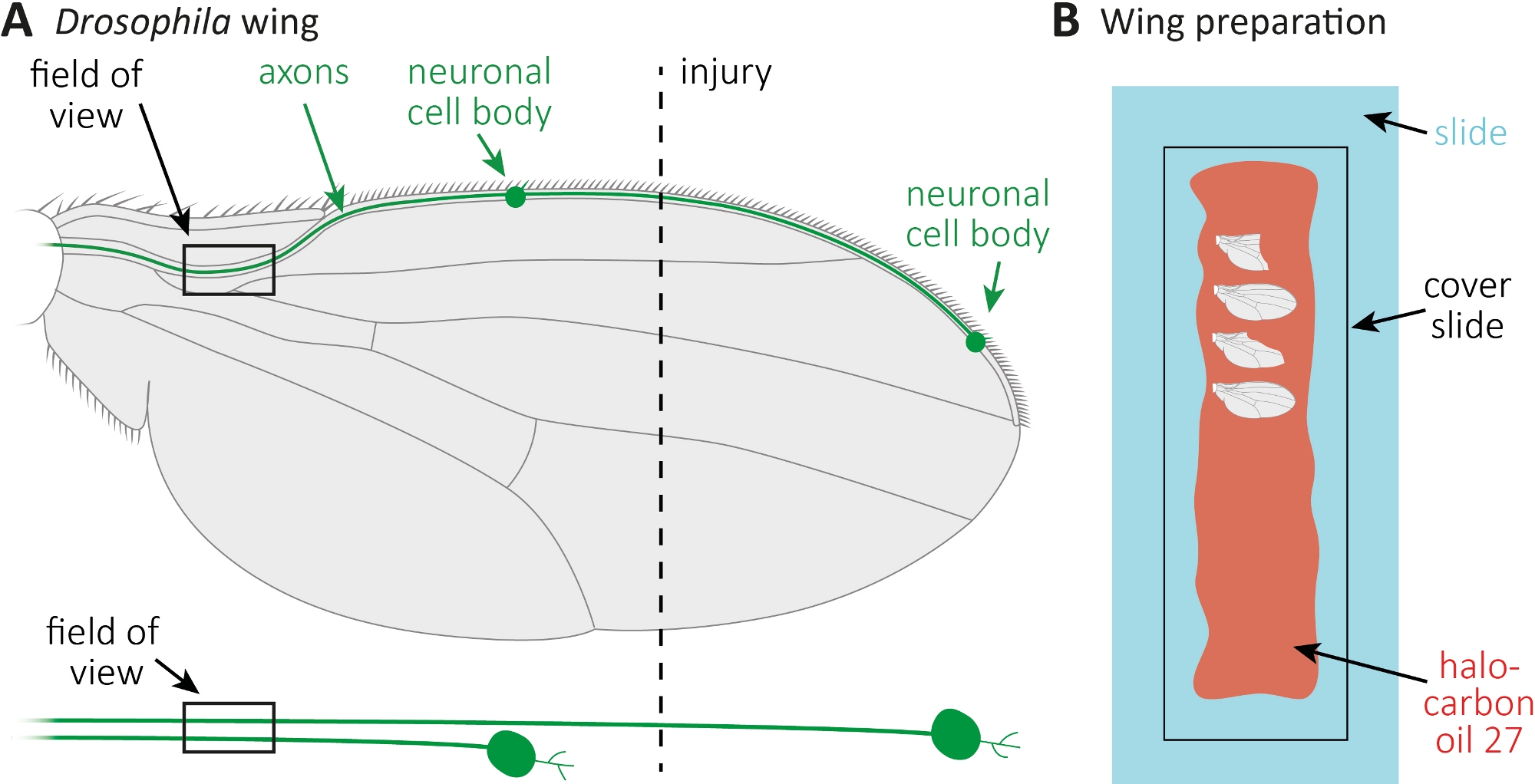
- Anestetizar moscas en almohadillas CO2. Utilice microtijeras para cortar la vena del ala anterior aproximadamente en el centro del ala(Figura 1A). Utilice un ala para la lesión y el otro ala como un control no lesionado partido por edad. Aplicar una lesión por ala, y asegúrese de obtener suficientes alas lesionadas (aproximadamente 15 alas).
NOTA: Todo el ala se puede cortar a través, pero es suficiente para cortar sólo la vena del ala anterior. Esta es la parte más fuerte del ala. - Recuperar las moscas en viales que contienen alimentos.
- Disección de alas y visualización de axónes
- Esparza 10 ml de aceite de halocarbono 27 con una pipeta a lo largo de un portaobjetos de vidrio entero(Figura 1B).
- Cortar el lesionado, así como, el ala de control no lesionada en los puntos de tiempo deseados (por ejemplo, 1 o 7 días después de la lesión). Usa microtijeras para cortar, y pinzas para agarrar el ala. Monte el máximo de 4 alas en aceite de halocarbono 27(Figura 1B)y cúbralas con un portaobjetos de cubierta.
- Imagen del ala inmediatamente usando un microscopio de disco giratorio. Adquiera una serie de secciones ópticas a lo largo del eje z con un tamaño de paso de 0,33 m y comprima las pilas z en un único archivo para análisis posteriores.
NOTA: No agarre la vena del ala anterior donde se alojan cuerpos celulares y axones. Coge el ala en el centro. El tejido en las alas no es fijo; mantener el tiempo de montaje de las alas a la toma de imágenes de estos en menos de 8 minutos.

Figura 1: Observación de la morfología del axón durante la muerte del axón en el ala. (A) Ala de mosca esquemática con dos neuronas sensoriales escasamente etiquetadas por GFP, que también se indican por separado a continuación. Se indican el lugar de la lesión y el campo de observación. (B) Configuración esquemática para imágenes de alas. Las alas de control lesionadas y no lesionadas (gris) están montadas en aceite de halocarbono 27 (rojo) sobre un portaobjetos de vidrio (azul claro) y cubiertas con un portaobjetos (negro). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
2. Observación de la Morfología de Axón y Sinapsis durante la muerte de Axon en el SNC
- Ablación antenal: lesión de paquetes enteros de axón
- Utilice 5 hembras vírgenes y 5 machos del genotipo derecho(figura 5A,generación P0) para realizar cruces en RT. Pase P0 a viales nuevos cada 3-4 días. Recoger la progenie adulta recién cerrada (generación F1) diariamente y dejar que fensite durante 7 hasta 14 días.
- Anestetizar moscas en almohadillas CO2. Utilice pinzas para ablar el segmentoantenal derecho 3 rd para la ablación unilateral; o ambos segmentos antenales 3rd izquierdo y derecho para la ablación bilateral(Figura 2A-C). Esto eliminará los cuerpos celulares neuronales etiquetados por gFP, mientras que sus proyecciones axonales permanecen en el SNC.
NOTA: La ablación de antenas separa todo el haz de axón. Si se realiza la ablación unilateral, el haz de axón en el lado contralateral (la antena no blada) sirve como control interno. Asegúrese de realizar suficientes ablaciones antenales (aproximadamente 15 animales). - Recuperar las moscas en viales que contienen alimentos.
- Disección cerebral y visualización de axónes
- Mezclar la base de elastómero de silicona (9 ml) y el agente de curado (1 ml) en una relación de volumen de 10:1. Transfiera cada mezcla de 5 ml en una placa de cultivo de tejido de 35 mm y reduzca el aire introducido mezclando con agitación suave en la campana de humos durante la noche. La mezcla se solidifica en un plazo de 24 h.
NOTA: Las placas de disección deben prepararse una sola vez y se pueden utilizar varias veces. - Anestetizar moscas en almohadillas de CO2 y decapitar cabezas adultas usando dos pinzas en los puntos de tiempo deseados (por ejemplo, 1 o 7 días después de la ablación de antenas). Usa una pinza para agarrar el cuello, y la otra pinza para fijar el tórax. Tire suavemente del cuello y la cabeza del tórax.
NOTA: Deje las cabezas decapitadas en el cojín CO2 hasta que se alcance el número deseado, pero asegúrese de continuar con el siguiente paso en un plazo de 30 minutos. - Transfiera todos los cabezales a un tubo de microcentrífuga de 1,5 ml que contenga 1 ml de solución de fijación que contenga un 4% de paraformaldehído (PFA) y un 0,1% de Tritón X-100 en solución salina tamponada (PBS) con pinzas que se hayan sumergido en la solución de fijación.
NOTA: Las cabezas de mosca se pegan bien en las pinzas mojadas. Hace que sea factible transferir todas las cabezas fácilmente al tubo de microcentrífuga. - Fijar cabezas durante 20 minutos con agitación suave en RT. Coloque el tubo de microcentrífuga en el hielo, las cabezas gravitarán en la parte inferior del tubo de microcentrífuga. Retire el sobrenadante con una pipeta y repita este procedimiento con cinco lavados de 2 min con 1 ml de tampón de lavado que contenga 0,1% De Tritón X-100 en PBS con agitación suave en RT, para eliminar la solución de fijación residual.
NOTA: Videos sobre cómo diseccionar cerebros Drosophila adultos están disponibles27. - Transfiera los cabezales con una pipeta de vidrio a una placa de disección llena de tampón de lavado. Use una pinza para agarrar y sacar la probóscsí de la cabeza, mientras sostiene la cabeza con la otra pinza. Esto dejará un agujero si la probóscsí estuviera unida al exoesqueleto.
- Utilice dos pinzas para eliminar el exoesqueleto entre el agujero y cada ojo compuesto. Esto hará que sea factible abrir la estructura de la cabeza con ambas pinzas, y rascar suavemente el cerebro dentro.
- Limpie cada cerebro eliminando la tráquea o la grasa pegada a él(Figura 2D,parte superior). Una vez que el cerebro esté limpio, colóquelo en un nuevo tubo de microcentrífuga que contenga 1 ml de tampón de lavado sobre hielo.
NOTA: Los lóbulos ópticos dañados o perdidos no afectarán al lóbulo olfativo en el centro del cerebro(Figura 2D,parte superior). - Reemplace el tampón de lavado por 1 ml de solución de fijación una vez que todos los cerebros se recojan y acumulen en la parte inferior del tubo de microcentrífuga. Fijar cerebros durante 10 minutos con balanceo en RT, seguido de cinco lavados de 2 minutos en 1 ml de tampón de lavado con balanceo en RT.
- Aplicar anticuerpos primarios (1:500) en el tampón de lavado durante la noche con balanceo a 4oC, seguido de 10 lavados de más de 2 h utilizando 1 ml de tampón de lavado con balanceo a RT.
- Aplicar anticuerpos secundarios (1:500) en el tampón de lavado 2 h con balanceo en RT y envolver el tubo microcentrífugo en papel de aluminio para bloquear la luz. Mantenga el tubo de microcentrífuga cubierto con papel de aluminio durante el resto del procedimiento. Aplicar diez lavados con 1 ml de tampón de lavado durante 2 h con balanceo a RT.
- Retire el sobrenadante y utilice una sola gota de reactivo antidescolorpara cubrir los cerebros en el tubo de microcentrífuga. Incubar cerebros durante al menos 30 minutos a 4 oC antes de prepararlos para el montaje y la toma de imágenes.
- Prepare una diapositiva de cubierta, pegue cinta de laboratorio en ella y corte una forma similar a "T" de la cinta(Figura 2D,parte inferior). El espacio resultante sirve como área donde el reactivo antifade que contiene el cerebro28 será pipeteado en, preferiblemente en ambas cámaras.
NOTA: Utilice una punta de pipeta de 20-200 ml en la que se hayan cortado 3 mm de la punta para ensanchar la abertura de la pipeta. Esto hará factible pipetear el reactivo antidesor que contiene el cerebro. Cubra cuidadosamente el cerebro con un portaobjetos. - Usa arcilla para preparar dos rollos pequeños y uniformes. Asegúrese de que los rollos de arcilla no sean superiores a la altura de un portaobjetos de vidrio. Pegue los rollos de arcilla en el portaobjetos de vidrio(Figura 2D,parte inferior). Coloque el sándwich de diapositivades de cubierta que contiene cerebro en los rollos de arcilla.
NOTA: Los axons etiquetados con GFP y sus sinapsis se encuentran en la parte frontal del cerebro. Por lo tanto, es más fácil crear imágenes desde el frente. Sin embargo, los cerebros se enfrentarán hacia arriba o boca abajo en el sándwich de diapositivas de la cubierta. Los rollos de arcilla sirven como soportes de sándwich, y durante la toma de imágenes, el sándwich se puede voltear al revés. Esto hará que sea factible adquirir imágenes del frente de cada cerebro. - Adquiera una serie de secciones ópticas a lo largo del eje z con un tamaño de paso de 1,0 m utilizando un microscopio confocal, y comprima las pilas z en un solo archivo para análisis posteriores, para evaluar el número de proyecciones axonales que permanecen intactas.
- Mezclar la base de elastómero de silicona (9 ml) y el agente de curado (1 ml) en una relación de volumen de 10:1. Transfiera cada mezcla de 5 ml en una placa de cultivo de tejido de 35 mm y reduzca el aire introducido mezclando con agitación suave en la campana de humos durante la noche. La mezcla se solidifica en un plazo de 24 h.

Figura 2: Observación de la morfología del axón y la sinapsis durante la muerte del axón en el cerebro. (A) Vista lateral de una cabeza de mosca esquemática con cuerpos de celda, axónes y sinapsis etiquetados por GFP. (B) Vista frontal de alta aumento de las neuronas receptoras olfativas etiquetadas por GPF y sus axónas y sinapsis. Los cuerpos celulares se alojan en elsegmento antenal 3, y sus axones se proyectan en el SNC. Los axons forman sinapsis en un glomérulo en el lóbulo olfativo izquierdo, cruzan la línea media y forman sinapsis en el glomérulo en el lóbulo olfativo contralateral. (C) Ejemplos de cabezas de mosca con ablación antena unilateral. Parte superior: Control incontrolado. Medio: Ablación delsegmento antenal 3. Parte inferior: Ablación del segmento de antena2nd (y por lo tanto también 3rd). (D) Preparación del cerebro. Arriba: Cerebro de mosca diseccionado esquemático con lóbulos olfativos indicados y proyecciones axonales en el campo de visión. Parte inferior: Configuración esquemática para imágenes cerebrales. Dos rollos de arcilla (verde) se montan en un tobogán de vidrio (azul claro), llevan un sándwich de diapositiva de cubierta, que contiene cerebros mosca (gris). Los cerebros están montados en reactivo antifade (púrpura), rodeados por una cinta de laboratorio (naranja), y cubiertos por dos diapositivas de cubierta (negro). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
3. Grooming Induced by Optogenetics as a Readout for Axon and Synapse Function
- Configuración optogenética
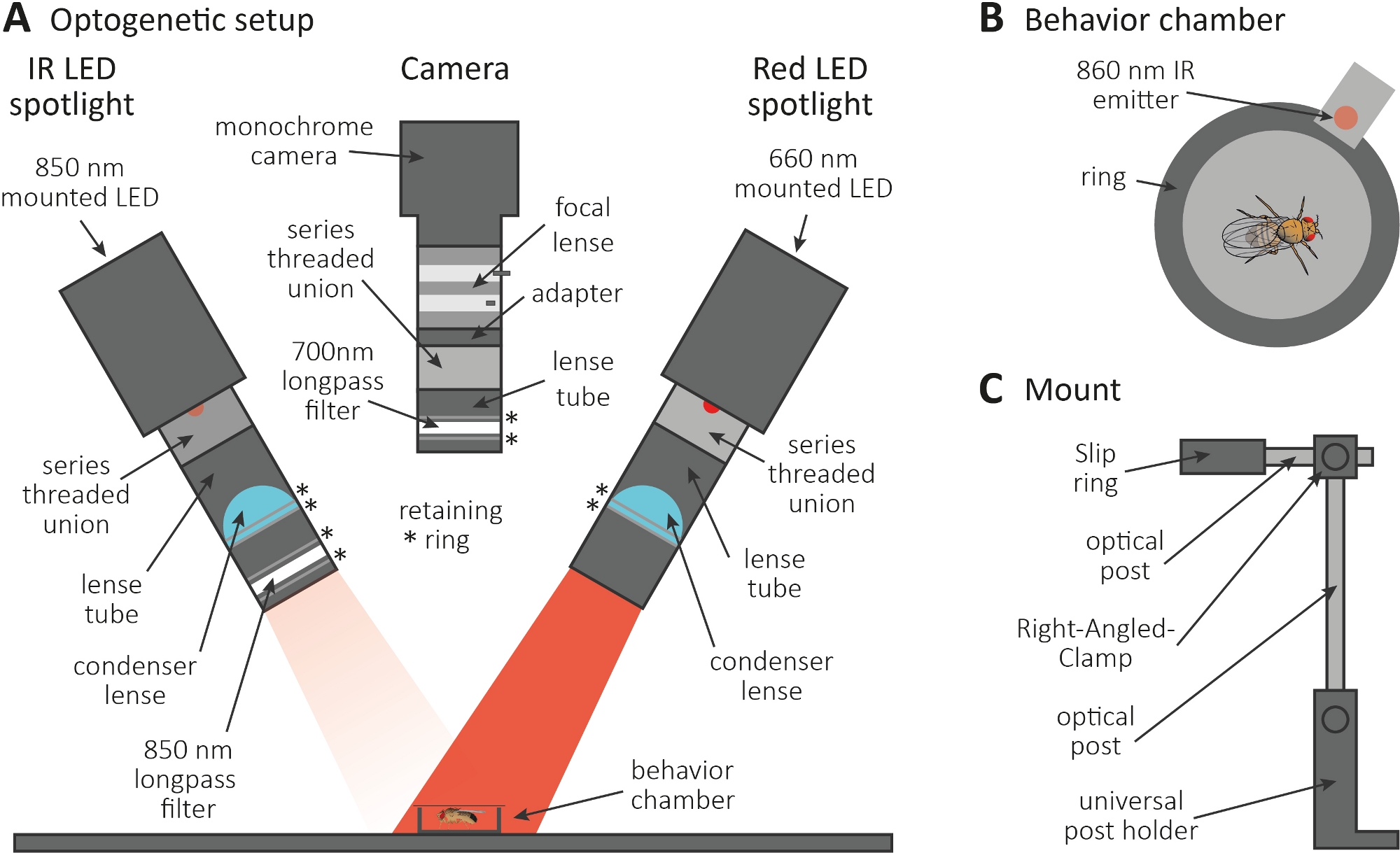
- Realizar el experimento optogenético en una habitación oscura. Asegúrese de que la configuración consiste en un proyector LED infrarrojo (IR) de 850 nm para iluminar moscas en la oscuridad(Figura 3A),un intermitente proyector LED rojo de 660 nm para activar las neuronas que expresan CsChrimson, y una cámara monocroma con un filtro de paso largo de 700 nm, que evita la grabación de destellos de luz roja.
- Utilice una impresora 3D para generar una pequeña cámara de comportamiento circular con un diámetro de 1 cm, cúbrala con una diapositiva de cubierta y coloque un emisor de 860 nm acoplado al proyector LED rojo junto a la cámara(Figura 3B).
NOTA: El emisor indica cuando el proyector LED rojo está encendido, activando así las neuronas. - Monte los proyectores LED y la cámara en la parte superior de la cámara(Figura 3A, C).
- Activar las neuronas por 10 Hz parpadea durante 10 s. La duración de la activación se puede ajustar de acuerdo con el diseño experimental.
- Preparación de moscas para optogenética
- Derretir los alimentos mosca en un microondas. Después de que el alimento se enfríe, antes de la solidificación, agregue 1:100 de 20 mM todo trans-retiniano en etanol (EtOH) a una concentración final de 200 M. Mezclar bien, y verter el alimento inmediatamente en viales vacíos.
NOTA: Evite agregar toda la trans-retinal a los alimentos calientes, esto podría resultar en una optogenética menos eficiente. - Cubra los viales que contienen alimentos solidificados con tapones o bolas de algodón. Envuelva los viales con papel de aluminio. A continuación, guarde los viales que contienen alimentos en una habitación oscura y fría.
- Utilice 5 hembras vírgenes y 5 machos(Figura 6A,generación P0) del genotipo correcto para realizar cruces en RT. Pase P0 a viales nuevos cada 3-4 días. Recoger la progenie adulta recién cerrada (generación F1) en una base diaria y dejar que fírban durante 7 hasta 14 días en viales recubiertos de aluminio que contienen 200 m de retiniano trans en comida volante.
- Recoger moscas tocándolas de los viales que contienen alimentos en un vial vacío sin alimentos. Enfríe el vial en agua que contenga hielo durante aproximadamente 30 s. Las moscas se quedarán dormidas. Coloque moscas individuales rápidamente en pequeñas cámaras cubiertas con una diapositiva de cubierta(Figura 3B).
NOTA: Tan pronto como las moscas se calientan, se despiertan. Es crucial esparcir rápidamente moscas individuales en cámaras individuales cada una. Evitar almohadillas de CO2 para anestesiar moscas, esto afectará su comportamiento. - Realizar optogenética para lograr el aseo de antenas. Aquí, el protocolo consta de los siguientes intervalos: 30 s donde la luz roja está ausente, seguido de 10 s de exposición a luz roja a 10 Hz. Repita este procedimiento tres veces en total, seguido de un intervalo adicional de 30 s donde la luz roja está ausente12,,29,30.
NOTA: Este protocolo se puede ajustar según la preferencia experimental. - Recoger moscas individuales de cada cámara en almohadillas CO2. Someterlos a una lesión en la antena. Ablate los segmentos de antena 2a la izquierda y a la derecha 2(Figura 2C). Esto eliminará los cuerpos celulares de las neuronas del órgano de Johnston (JO), mientras que las proyecciones axonales permanecen en el SNC. Recuperar las moscas en viales revestidos de aluminio que contienen 200 m de retina trans.
NOTA: Para el aseo de antenas inducido por la optogenética, los cuerpos celulares de las neuronas sensoriales se alojan en el2o segmento de antena(Figura 2C). - En los puntos de tiempo correspondientes (por ejemplo, 7 días después de la ablación de antena), el sujeto vuela a otro ensayo de aseo (volver al paso 3.2.4).
- Derretir los alimentos mosca en un microondas. Después de que el alimento se enfríe, antes de la solidificación, agregue 1:100 de 20 mM todo trans-retiniano en etanol (EtOH) a una concentración final de 200 M. Mezclar bien, y verter el alimento inmediatamente en viales vacíos.

Figura 3: Configuración optogenética para inducir el aseo como una lectura para la función de axón y sinapsis. (A) Ilustración de los componentes ensamblados necesarios para la optogenética. Foco LED infrarrojo (IR), cámara y proyector LED rojo (de izquierda a derecha, respectivamente). Los componentes que incluyen una descripción detallada se enumeran en la Tabla de materiales. (B) Ilustración de vista superior de una cámara de comportamiento que incluye un emisor IR para indicar la activación del foco LED rojo. (C) Ilustración de una sola configuración de montaje. Se requieren un total de tres configuraciones de montaje para los dos proyectores LED y la cámara, respectivamente. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Resultados
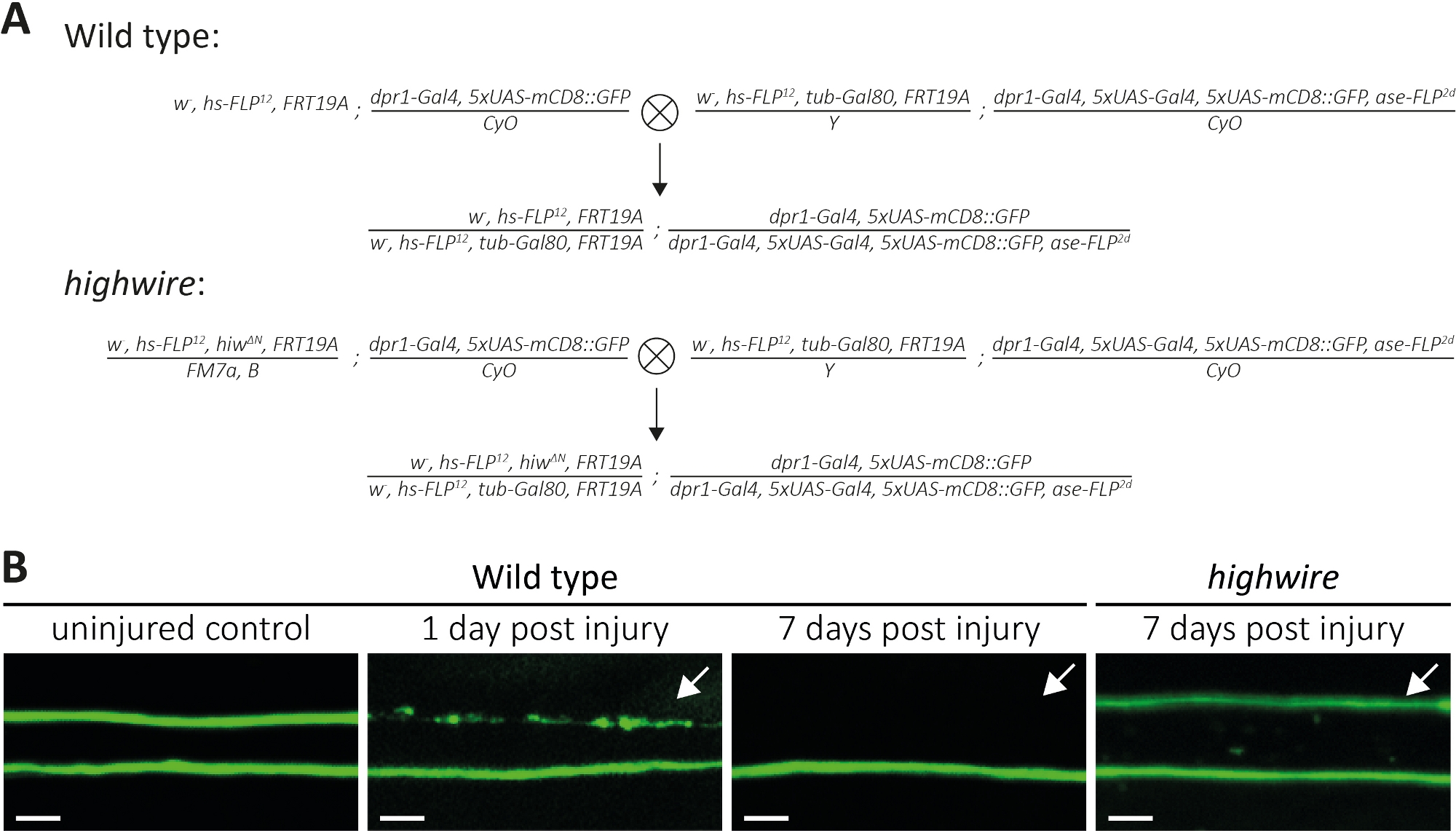
Arriba, describimos tres métodos para estudiar la morfología y la función de los axons cortados y sus sinapsis. El primer método permite la observación de alta resolución de axónes individuales en el PNS. Requiere clones generados por la técnica MARCM14,31. Aquí, realizamos cruces para generar clones MARCM mutantes de tipo salvaje y de alto cableado (Figura 4A). Un simple corte en el centro del ala induce lesión de axón de las neuronas alojados distal (por ejemplo, en el lado exterior del ala), mientras que las neuronas proximales (por ejemplo, entre el sitio de corte y el tórax) permanecen sin lesiones. Este enfoque hace factible observar la muerte de axón lado a lado de los axónicos de control no lesionados en el mismo haz nervioso(Figura 1A, Figura 4B). Aquí, utilizamos un fondo genético que resulta en un bajo número de clones etiquetados por GFP (por ejemplo, dos en cada experimento14). Presentamos ejemplos de 1 y 7 días después de la lesión de axónes de tipo salvaje, para proporcionar ejemplos de axónes de control, axones sometidos a muerte por axón y fragmentos axonales que están siendo despejados por la glia circundante, respectivamente. Además, repetimos lesiones axonales en mutantes de alto alambre donde analizamos el resultado 7 días después de la lesión.
Las alas de control lesionadas albergan dos clones de tipo salvaje, por lo tanto dos axons de tipo salvaje etiquetados por la GFP(Figura 4B,tipo salvaje, control sin lesionarse). Un día después de cortar la mitad del ala mediante el uso de microtijeras, la muerte por axón se induce en axones etiquetados por GFP donde los cuerpos celulares son distales al sitio de corte, mientras que los axones de cuerpos celulares alojados proximalmente sirven como un control interno dentro del mismo haz nervioso(Figura 4B,tipo salvaje, 1 día después de la lesión). Observe el rastro de escombros axonales en la parte superior indicada por la flecha. 7 días después de una lesión axonal, los desechos axonales etiquetados por la GFP son despejados por la glia circundante, mientras que los axónicos de control no lesionados etiquetados por GFP permanecen en el haz nervioso(Figura 4B,tipo salvaje, 7 días después de la lesión, flecha). Por el contrario, los axónicos mutantes de hilos altos que han sido cortados durante 7 días permanecen morfológicamente preservados, de acuerdo con los hallazgos anteriores11,14 (Figura 4B, highwire, 7 días después de la lesión, flecha). Estos resultados demuestran la poderosa resolución visual del ala Drosophila. La muerte por axón se puede observar lado a lado de los controles no lesionados en el mismo haz nervioso. Mientras que los axons de tipo salvaje sufren muerte de axón dentro de 1 día después de la lesión y los desechos resultantes se limpian dentro de 7 días, los mutantes de alambre alto deficientes de axón permanecen morfológicamente preservados durante 7 días.

Figura 4: Enfoque para estudiar la muerte por axón de los axónicos de neuronas sensoriales etiquetados por GFP en el ala. (A) Cruces esquemáticas para generar clones de tipo salvaje y highwire en el ala (generación P0 y F1, respectivamente). Las hembras vírgenes están a la izquierda, los machos a la derecha. Consulte la Tabla de materiales para obtener detalles del genotipo. (B) Ejemplos de control y axons con etiqueta GFP lesionados. El campo de visión se indica en (Figura 1A). De izquierda a derecha: axons de control de tipo salvaje no lesionados, axons de tipo salvaje 1 día después de la lesión, axón de tipo salvaje 7 días después de la lesión, axón esclosos mutantes de alambre alto 7 días después de la lesión, respectivamente. Las flechas indican axónes cortados, Barra de escala de 5 m. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
El segundo método describe cómo visualizar los haces enteros de axón que se proyectan en el SNC donde forman sinapsis, que pertenecen a las neuronas alojados en antenas izquierda y derecha(Figura 2A-C). Aquí, realizamos cruces para generar clones MARCM mutantes de tipo salvaje y de alto cableado (Figura 5A). Los axons etiquetados por gFP y sus sinapsis se pueden visualizar en el transcurso de días a semanas, en ausencia de lesión(Figura 5B,Tipo salvaje, control lesionado). Alternativamente, los animales pueden ser sometidos ala ablación del segmento antenal 3, y los axónicos cortados etiquetados con GFP y sus sinapsis se pueden observar durante un curso de tiempo durante horas a días. Nos centramos en 7 días después de la ablación antenal, porque en este momento, los axón y sus sinapsis han sufrido la muerte de axón, y los desechos resultantes son limpiados por la glia circundante. Si se realiza la ablación unilateral de la antena derecha, entonces el haz de axón derecho se corta y se desmontará y los desechos resultantes se limpian completamente 7 días después de la lesión(Figura 5B,tipo salvaje, ablación unilateral, 7 días después de la lesión, flechas), de acuerdo con los hallazgos anteriores13. Alternativamente, tanto la antena derecha como la izquierda pueden ser abladas, que cortarán ambos haces de axón, y 7 días después de la lesión, los axones y sus sinapsis desaparecieron(Figura 5B,tipo salvaje, ablación bilateral, 7 días después de la lesión, flecha). Por el contrario, la ablación unilateral de las antenas derechas en mutantes de hilos altos da como resultado axónes cortados que permanecen preservados 7 días después de la lesión, de conformidad con los hallazgos anteriores11,,14 (Figura 5B, alta,ablación unilateral, 7 días después de la lesión, flecha). Estos resultados demuestran que los axons de tipo salvaje cortados sufren muerte de axón y los desechos resultantes se limpian en 7 días, mientras que los mutantes de alambre alto deficientes en muerte por axón no pueden sufrir la muerte por axón y permanecen morfológicamente preservados durante 7 días.

Figura 5: Enfoque para estudiar la muerte por axón de los axónicos de neuronas sensoriales etiquetadas con GFP en el cerebro. (A) Cruces esquemáticas para generar clones de tipo salvaje y highwire en el cerebro (generación P0 y F1, respectivamente). Las hembras vírgenes están a la izquierda, los machos a la derecha. Consulte la Tabla de materiales para obtener detalles del genotipo. (B) Ejemplos de control y axons con etiqueta GFP lesionados. De izquierda a derecha: controles de tipo salvaje no lesionados, tipo salvaje 7 días después de la ablación antena unilateral, tipo salvaje 7 días después de la ablación antenal bilateral, y mutantes de hilos altos 7 días después de la ablación antena unilateral, respectivamente. Las flechas indican paquetes de axón cortados, Barra de escala a 10 m. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
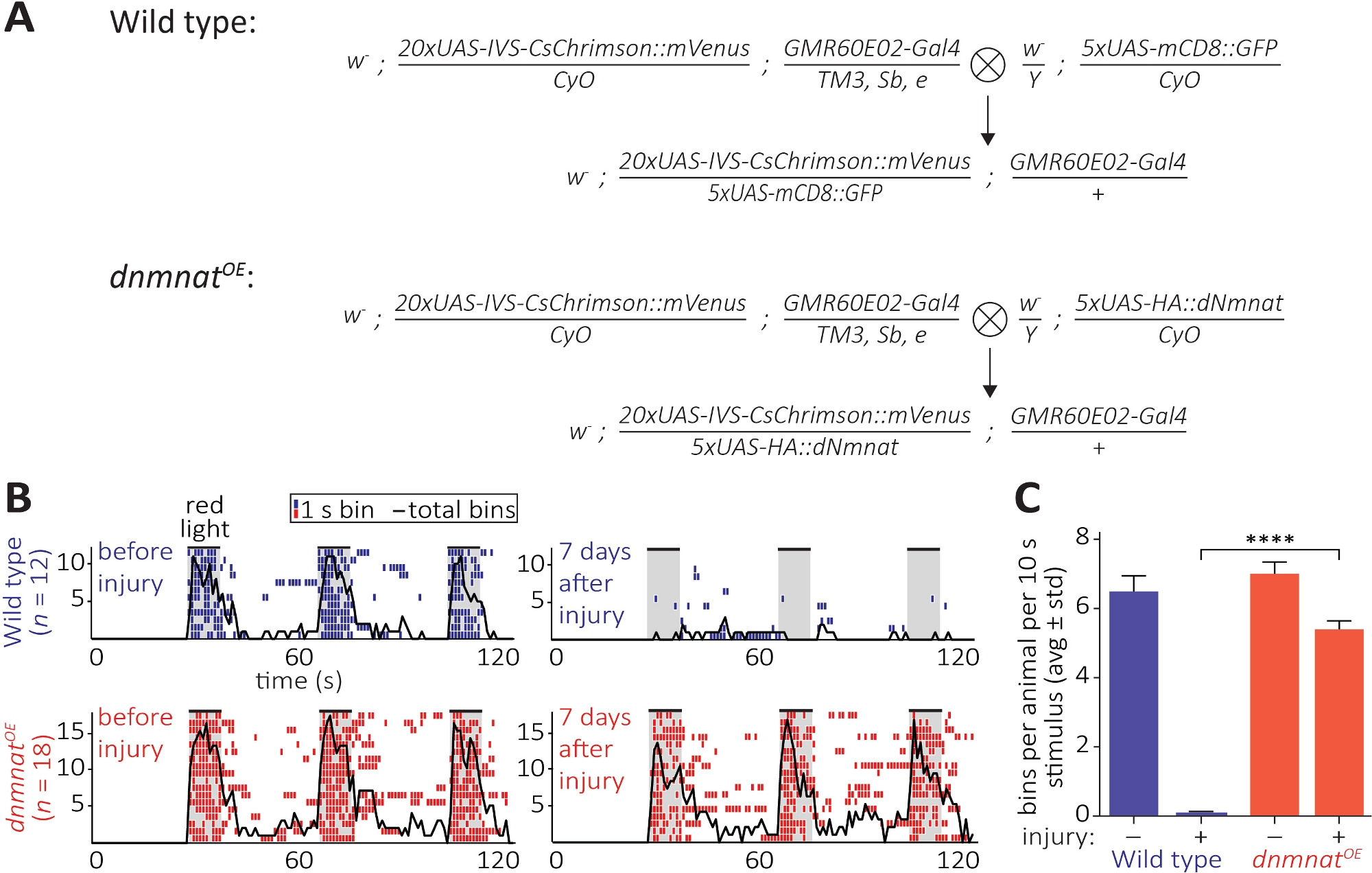
El tercer método permite la observación de la preservación funcional de los axons cortados y sus sinapsis en el SNC. Se basa en la manipulación de un subconjunto de neuronas JO alojados en elsegmento de antena 2 que son suficientes para inducir el aseo de la antena. La expresión de un canal rhodopsin de cambio rojo (CsChrimson) en las neuronas JO, combinado con la suplementación dietética de toda la trans-retiniana, es suficiente para lograr un simple comportamiento de aseo postsináptico sobre la exposición a la luz roja12,30. Aquí, realizamos cruces para generar neuronas JO de tipo salvaje, y las neuronas JO sobreexpresan dnmnat (dnmnatOE) (Figura 6A). Las moscas de tipo salvaje o las moscas que contienen neuronas JO con muerte de axón atenuada(dnmnatOE),ambos albergan un potente comportamiento de aseo antes de la lesión. Sin embargo, 7 días después de una lesión (por ejemplo, ablación bilateral del2o segmento de antena), el aseo no puede ser provocado por la optogenética en moscas de tipo salvaje debido a la degeneración inducida por axón y sinapsis inducida por lesiones, mientras que los animales con muerte por axón atenuado continúan acicalándose(Figura 6B, Película 1,2). Por lo tanto, la muerte de axón atenuado es capaz de preservar funcionalmente los axones cortados y sus sinapsis durante 7 días.

Figura 6: Enfoque para visualizar la función axonal y sináptica después de la axotomía. (A) Cruces esquemáticas para generar tipo salvaje y dnmnat sobreexpresar las neuronas sensoriales JO (generación P0 y F1, respectivamente). Las hembras vírgenes están a la izquierda, los machos a la derecha. Consulte la Tabla de materiales para obtener detalles del genotipo. (B) Etogramas individuales del comportamiento de aseo inducidos por la optogenética. Parte superior: eomitas individuales de tipo salvaje vuela antes y 7 días después de la lesión (azul). Parte inferior: eomitas individuales de moscas sobre-expresando dnmnat (dnmnatOE) en las neuronas JO antes y 7 días después de la lesión (rojo). Cada bin indica al menos 1 comportamiento de aseo dentro de 1 s. La línea negra indica la suma de todas las ubicaciones. (C) Cuantificación del comportamiento de aseo. Los datos se muestran como promedio de desviación estándar, p > 0.001 (ANOVA unidireccional, comparación múltiple con la prueba post hoc de Tukey). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Película 1: Comportamiento representativo de acicalamiento de tipo salvaje provocado por la optogenética antes y 7 días después de la ablación antenal. Haga clic aquí para descargar este video.
Película 2: Comportamiento representativo del aseo provocado por la optogenética en moscas que expresan excesivamente dnmnat en las neuronas JO antes y 7 días después de la ablación antenal. Haga clic aquí para descargar este video.
Discusión
Los protocolos descritos aquí permiten la observación robusta y reproducible de la morfología, así como la función de los axónicos y sus sinapsis separadas de sus cuerpos celulares en Drosophila. El ensayo del ala facilita la observación de axón de muerte lado a lado de axón de axón de control no lesionado en el PNS14,mientras que el ensayo antenal facilita la observación de bultos nerviosos enteros de axónicos etiquetados con GFP y sus sinapsis, para evaluar tanto la morfología como la función en el cerebro (CNS)12. Hay pasos críticos y ciertas ventajas para cada enfoque para estudiar la morfología que deben tenerse en cuenta al diseñar experimentos.
Para observar la morfología del axón en el PNS en el ala, los experimentos se pueden realizar fácilmente, debido a la transparencia del ala: permite eludir la disección y la inmunohistoquímica. Sin embargo, debido a la falta de fijación, las alas tienen que ser imágenes inmediatamente después del montaje14. Actualmente, se utilizan con frecuencia dos controladores Gal4 distintos, ok371Gal4 o dpr1Gal4,y ambas referencias ofrecen enfoques distintos para cuantificar la degeneración14,26. Se recomienda el etiquetado escaso de algunas neuronas, mediante el uso de "Análisis de mosaico con un marcador celular reprimible (MARCM)"14,31, ya que la resolución de morfología axonal no tiene precedentes. Por el contrario, la observación de las sinapsis no es posible en las alas, se encuentran en el cordón del nervio ventral dentro del tórax de las moscas. Además, la inmunohistoquímica no puede visualizar marcadores axonales adicionales: la cutícula cerosa hace imposible la difusión de fijadores y anticuerpos en el tejido subyacente.
Para observar la morfología del axón y la sinapsis en el SNC, se deben realizar disecciones cerebrales. Ofrecen la ventaja de visualizar marcadores axonales y sinápticos adicionales mediante el uso de inmunohistoquímica, y se pueden observar sinapsis junto con axones en el mismo campo de visión10,13. Una gran colección de factores caracterizados de la neurona receptora olfativa (ORN) Gal4 es fácilmente disponible32,y con frecuencia, OR22aGal4 es el conductor de elección. Para la ablación antenal, los cuerpos celulares de las neuronas OR22a se alojan en el3er segmento(Figura 2B). Se utiliza una cuantificación basada en la intensidad de fluorescencia para cuantificar la degeneración de axónes o sinapsis13. Por el contrario, los experimentos consumen mucho tiempo debido a la disección cerebral y la tinción de anticuerpos.
Para visualizar la función axonal y sináptica después de la axotomía, la optogenética se utiliza para activar el aseo de antenas: sirve como una lectura para la preservación funcional de los axones cortados y sus sinapsis12. El circuito de aseo y los controladores Gal4 sensoriales, inter y motorneuron correspondientes se han descrito a fondo29,,30. GMR60E02Gal4 etiqueta un subconjunto de neuronas sensoriales de órgano (JO) de Johnston, que son necesarias y suficientes para el aseo29,30. Para la ablación antenal, los cuerpos celulares de las neuronas JO se alojan en el2o segmento de antena(Figura 2B). Una configuración optogenética se puede construir fácilmente desde cero, o una configuración existente ajustada. Es importante destacar que los experimentos tienen que realizarse en una habitación oscura, y las moscas se visualizan así con un foco LED infrarrojo (IR). Cuando se utiliza CsChrimson como canal, es crucial suministrar el alimento con toda la retina trans y un foco LED rojo para activar las neuronas JO29. Alternativamente, los canales sensibles a la luz azul y un proyector LED azul, o el canal TrpA1 y la temperatura se pueden utilizar para la activación neuronal29,33. La cuantificación del comportamiento de aseo ya se ha descrito12,29.
Cuando estos ensayos se utilizan para estudiar específicamente la muerte por axón, es importante tener en cuenta que el fenotipo de la preservación morfológica o funcional debe ser robusto con el tiempo. Hay casos en los que la muerte por axón conduce a un fenotipo consistente pero menos pronunciado en la preservación morfológica34,35,y si tal fenotipo se traduce en preservación funcional aún está por determinar.
También se han observado fenotipos de muerte por axón en neuronas durante el desarrollo de larvas de Drosophila, donde los nervios fueron aplastados en lugar de heridos11,23. Aquí, nos centramos específicamente en las neuronas Drosophila adultas que completaron el desarrollo. En este contexto, se puede implementar fácilmente el uso de la interferencia de ARN36o CRISPR/Cas937 específica del tejido. Es importante destacar que las técnicas anteriores se pueden utilizar en un contexto independiente de la muerte por axón: facilitan la caracterización de los factores de mantenimiento neuronal38,transporte axonal39,cambios en las mitocondrias axonales dependientes de la edad40,y morfología de las mitocondrias axonales41.
Divulgaciones
Los autores declaran que no tienen nada que revelar.
Agradecimientos
Nos gustaría dar las gracias a todo el laboratorio de Neukomm por sus contribuciones. Este trabajo fue apoyado por un premio de Profesor Asistente de la Fundación Nacional Suiza de Ciencias (SNSF) (concesión 176855), la Fundación Internacional para la Investigación en Paraplegia (IRP, subvención P180), SNSF Spark (concesión 190919) y por el apoyo de la Universidad de Lausana y el Departamento de Neurociencias Fundamentales ('tat de Vaud') a LJN.
Materiales
| Name | Company | Catalog Number | Comments |
| Tweezers (high precision, ultra fine) | EMS | 78520-5 | Antennal ablation |
| MicroPoint Scissors (5-mm cutting edge) | EMS | 72933-04 | Wing injury |
| 1.5 mL microcentrifuge tube | Eppendorf | 30120086.0000 | |
| 35mm tissue culture dish | Sarstedt | 83.3900 | |
| Cover Slips, Thickness 1 | Thermo Scientific™ | BB02400600A113MNT0 | |
| Superfrost Microscope Slides | Thermo Scientific™ | AA00008032E00MNT10 | |
| High-Sensitivity USB 2.0 CMOS Camera, 1280 x 1024, Global Shutter | Thorlabs | DCC1240M | Camera setup |
| SM1 Retaining Ring for Ø1" Lens Tubes and Mounts | Thorlabs | SM1RR | |
| 25mm 1/1.2" C mount Lens | Tamron | M112FM25 | |
| Adapter with External M27 x 0.5 Threads and Internal SM1 Threads | Thorlabs | SM1A36 | |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| Ø25.0 mm Premium Longpass Filter, Cut-On Wavelength: 700 nm | Thorlabs | FELH0700 | |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 1" Long, Two Retaining Rings Included | Thorlabs | SM1M10 | |
| 850 nm, 900 mW (Min) Mounted LED, 1200 mA | Thorlabs | M850L3 | IR LED spotlight |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 2" Long, Two Retaining Rings Included | Thorlabs | SM1M20 | |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| Ø25.0 mm Premium Longpass Filter, Cut-On Wavelength: 850 nm | Thorlabs | FELH0850 | |
| SM1 Retaining Ring for Ø1" Lens Tubes and Mounts | Thorlabs | SM1RR | |
| 660 nm, 940 mW (Min) Mounted LED, 1200 mA | Thorlabs | M660L4 | Red LED spotlight |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 2" Long, Two Retaining Rings Included | Thorlabs | SM1M20 | |
| 15 V, 2.4 A Power Supply Unit with 3.5 mm Jack Connector for One K- or T-Cube | Thorlabs | KPS101 | LED control |
| T-Cube LED Driver, 1200 mA Max Drive Current | Thorlabs | LEDD1B | |
| 150 mm x 300 mm x 12.7 mm Aluminum Breadboard, M6 Double-Density Taps | Thorlabs | MB1530/M | Mount base |
| Ø12.7 mm Universal Post Holder, Spring-Loaded Locking Thumbscrew, L = 75 mm | Thorlabs | UPH75/M | Mount, 3x (IR LED, red LED, cam) |
| Ø1.20" Slip Ring for SM1 Lens Tubes and C-Mount Extension Tubes, M4 Tap | Thorlabs | SM1RC/M | |
| Ø12.7 mm Optical Post, SS, M4 Setscrew, M6 Tap, L = 150 mm | Thorlabs | TR150/M | |
| Ø12.7 mm Optical Post, SS, M4 Setscrew, M6 Tap, L = 40 mm | Thorlabs | TR40/M | |
| Right-Angle Clamp for Ø1/2" Posts, 5 mm Hex | Thorlabs | RA90/M | |
| M6 x 1.0 Stainless Steel Cap Screw, 16 mm Long, Pack of 25 | Thorlabs | SH6MS16 | screws for mount onto base |
| USB-6001 14-Bit 20 kS/s Multifunction I/O and NI-DAQmx | National Instruments | 782604-01 | Red LED spotlight controller |
| 20k Ohm 1 Gang Linear Panel Mount Potentiometer | TT Electronics/BI | P230-2EC22BR20K | fintuner for indicator |
| IR (860nm) emitter, 100 mA radial | Osram | 475-1365-ND | Red light indicator |
| cable | - | - | Misc |
| All-trans retinal | Sigma | R2625 | |
| Ethanol absolute | Vwr | 20821.296 | |
| Halocarbon Oil 27 | Sigma | H8773 | |
| Mowiol | Merk | 81381 | |
| Paraformaldehyde | Sigma | F8775 | |
| Phosphate buffered saline (PBS) | Sigma | P5493 | |
| Sylgard 184 silicone elastomer base | Dow Corning Corp | 4019862 | |
| Sylgard 184 silicone elastomer curing agent | Dow Corning Corp | 4019862 | |
| Triton X-100 | Sigma | T8787 | |
| Chicken anti-GFP antibodies | Rockland | 600-901-215 | Antibodies |
| Goat Dylight anti-Chicken | Abcam | ab96947 | |
| FM7a, B | BDSC | RRID:BDSC_785 | X chromosome |
| FRT19A[hs-neo] | BDSC | RRID:BDSC_1709 | |
| hiw[ΔN] | BDSC | RRID:BDSC_51637 | |
| hs-FLP[12] | BDSC | RRID:BDSC_1929 | |
| tub-Gal80[LL1] | BDSC | RRID:BDSC_5132 | |
| w[1118] | BDSC | RRID:BDSC_3605 | |
| 20xUAS-IVS-CsChrimson::mVenus | BDSC | RRID:BDSC_55135 | 2nd chromosome |
| 5xUAS-Gal4[12B] | Kyoto | RRID:Kyoto_108492 | |
| 5xUAS-HA::dnmnat | BDSC | RRID:BDSC_39702 | |
| 5xUAS-mCD8::GFP[LL5] | BDSC | RRID:BDSC_5134 | |
| ase-FLP[2d] | Freeman laboratory | Neukomm et al., 2014 (PNAS) | |
| CyO | BDSC | RRID:BDSC_2555 | |
| dpr1-Gal4 | BDSC | RRID:BDSC_25083 | |
| OR22a-Gal4 | BDSC | RRID:BDSC_9952 | |
| ey-FLP[6] | BDSC | RRID:BDSC_5577 | 3rd chromosome |
| GMR60E02-Gal4 | BDSC | RRID:BDSC_39250 | |
| TM3,Sb,e | BDSC | RRID:BDSC_3644 |
Referencias
- Matsuda, W., et al. Single Nigrostriatal Dopaminergic Neurons Form Widely Spread and Highly Dense Axonal Arborizations in the Neostriatum. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 29 (2), 444-453 (2009).
- Wedel, M. J. A Monument of Inefficiency: The Presumed Course of the Recurrent Laryngeal Nerve in Sauropod Dinosaurs. Acta Palaeontologica Polonica. 57 (2), 251-256 (2012).
- Mariano, V., Domínguez-Iturza, N., Neukomm, L. J., Bagni, C. Maintenance mechanisms of circuit-integrated axons. Current Opinion in Neurobiology. 53, 162-173 (2018).
- Conforti, L., Gilley, J., Coleman, M. P. Wallerian degeneration: an emerging axon death pathway linking injury and disease. Nature reviews Neuroscience. 15 (6), 394-409 (2014).
- Neukomm, L. J., Freeman, M. R. Diverse cellular and molecular modes of axon degeneration. Trends in Cell Biology. 24 (9), 515-523 (2014).
- Gustavsson, A., et al. Cost of disorders of the brain in Europe 2010. European Neuropsychopharmacology: The Journal of the European College of Neuropsychopharmacology. 21 (10), 718-779 (2011).
- Waller, A. Experiments on the Section of the Glossopharyngeal and Hypoglossal Nerves of the Frog, and Observations of the Alterations Produced Thereby in the Structure of Their Primitive Fibres. Philosophical Transactions of the Royal Society of London. 140, 423-429 (1850).
- Rosell, A. L., Neukomm, L. J. Axon death signalling in Wallerian degeneration among species and in disease. Open Biology. 9 (8), 190118(2019).
- Mack, T. G., et al. Wallerian degeneration of injured axons and synapses is delayed by a Ube4b/Nmnat chimeric gene. Nature Neuroscience. 4 (12), 1199-1206 (2001).
- Osterloh, J. M., et al. dSarm/Sarm1 is required for activation of an injury-induced axon death pathway. Science. 337 (6093), New York, NY. 481-484 (2012).
- Xiong, X., et al. The highwire ubiquitin ligase promotes axonal degeneration by tuning levels of nmnat protein. PLoS Biology. 10 (12), 1001440(2012).
- Neukomm, L. J., et al. Axon Death Pathways Converge on Axundead to Promote Functional and Structural Axon Disassembly. Neuron. 95 (1), 78-91 (2017).
- MacDonald, J. M., et al. The Drosophila cell corpse engulfment receptor Draper mediates glial clearance of severed axons. Neuron. 50 (6), 869-881 (2006).
- Neukomm, L. J., Burdett, T. C., Gonzalez, M. A., Zuchner, S., Freeman, M. R. Rapid in vivo forward genetic approach for identifying axon death genes in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 111 (27), 9965-9970 (2014).
- Lu, T. Y., et al. Axon degeneration induces glial responses through Draper-TRAF4-JNK signalling. Nature Communications. 8, 14355(2017).
- Lunn, E. R., Perry, V. H., Brown, M. C., Rosen, H., Gordon, S. Absence of Wallerian Degeneration does not Hinder Regeneration in Peripheral Nerve. The European Journal of Neuroscience. 1 (1), 27-33 (1989).
- Adalbert, R., et al. A rat model of slow Wallerian degeneration (Wld(S)) with improved preservation of neuromuscular synapses. The European Journal of Neuroscience. 21 (1), 271-277 (2005).
- Martin, S. M., O'Brien, G. S., Portera-Cailliau, C., Sagasti, A. Wallerian degeneration of zebrafish trigeminal axons in the skin is required for regeneration and developmental pruning. Development. 137 (23), Cambridge, England. 3985-3994 (2010).
- Feng, Y., et al. Overexpression of Wld(S) or Nmnat2 in Mauthner Cells by Single-Cell Electroporation Delays Axon Degeneration in Live Zebrafish. Journal of Neuroscience Research. 88 (15), 3319-3327 (2010).
- Gilley, J., Coleman, M. P. Endogenous Nmnat2 is an essential survival factor for maintenance of healthy axons. PLoS Biology. 8 (1), 1000300(2010).
- Babetto, E., Beirowski, B., Russler, E. V., Milbrandt, J., DiAntonio, A. The Phr1 ubiquitin ligase promotes injury-induced axon self-destruction. Cell Reports. 3 (5), 1422-1429 (2013).
- Gerdts, J., Summers, D. W., Sasaki, Y., DiAntonio, A., Milbrandt, J. Sarm1-mediated axon degeneration requires both SAM and TIR interactions. The Journal of Neuroscience. 33 (33), 13569-13580 (2013).
- Gerdts, J., Brace, E. J., Sasaki, Y., DiAntonio, A., Milbrandt, J. SARM1 activation triggers axon degeneration locally via NAD+ destruction. Science. 348 (6233), New York, NY. 453-457 (2015).
- Bridge, P. M., et al. Nerve crush injuries--a model for axonotmesis. Experimental Neurology. 127 (2), 284-290 (1994).
- Maxwell, W. L., Bartlett, E., Morgan, H. Wallerian Degeneration in the Optic Nerve Stretch-Injury Model of Traumatic Brain Injury: A Stereological Analysis. Journal of Neurotrauma. 32 (11), 780-790 (2015).
- Fang, Y., Soares, L., Teng, X., Geary, M., Bonini, N. M. A novel Drosophila model of nerve injury reveals an essential role of Nmnat in maintaining axonal integrity. Current Biology. 22 (7), 590-595 (2012).
- Janelia Farm Adult Drosophila Brain Dissection. , Available from: https://www.janelia.org/project-team/flylight/protocols (2015).
- Cold Spring Harbor. Mowiol mounting medium. Cold Spring Harbor Protocols. 2006 (1), (2006).
- Seeds, A. M., et al. A suppression hierarchy among competing motor programs drives sequential grooming in Drosophila. eLife. 3, 02951(2014).
- Hampel, S., Franconville, R., Simpson, J. H., Seeds, A. M. A neural command circuit for grooming movement control. eLife. 4, 08758(2015).
- Lee, T., Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron. 22 (3), 451-461 (1999).
- Vosshall, L. B., Wong, A. M., Axel, R. An olfactory sensory map in the fly brain. Cell. 102 (2), 147-159 (2000).
- Hampel, S., McKellar, C. E., Simpson, J. H., Seeds, A. M. Simultaneous activation of parallel sensory pathways promotes a grooming sequence in Drosophila. eLife. 6, (2017).
- Farley, J. E., et al. Transcription factor Pebbled/RREB1 regulates injury-induced axon degeneration. Proceedings of the National Academy of Sciences of the United States of America. 23 (6), (2018).
- Wang, H., et al. Rapid depletion of ESCRT protein Vps4 underlies injury-induced autophagic impediment and Wallerian degeneration. Science Advances. 5 (2), 4971(2019).
- Dietzl, G., et al. A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature. 448 (7150), 151-156 (2007).
- Port, F., et al. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. bioRxiv. 102, 636076(2019).
- Vagnoni, A., Hoffmann, P. C., Bullock, S. L. Reducing Lissencephaly-1 levels augments mitochondrial transport and has a protective effect in adult Drosophila neurons. Journal of Cell Science. 129 (1), 178-190 (2016).
- Vagnoni, A., Bullock, S. L. A cAMP/PKA/Kinesin-1 Axis Promotes the Axonal Transport of Mitochondria in Aging Drosophila Neurons. Current Biology. 28 (8), 1265-1272 (2018).
- Cao, X., et al. In vivo imaging reveals mitophagy independence in the maintenance of axonal mitochondria during normal aging. Aging Cell. 16 (5), 1180-1190 (2017).
- Smith, G. A., et al. Glutathione S-Transferase Regulates Mitochondrial Populations in Axons through Increased Glutathione Oxidation. Neuron. 103 (1), 52-65 (2019).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados