
Method Article
Un ensayo de actividad acoplada a enzimas de alto rendimiento para sondear la interacción de moléculas pequeñas con la dNTPasa SAMHD1
En este artículo
Resumen
SAMHD1 es una trifosfofosfato trifosfohidrolasa desoxinucleósido con funciones críticas en la salud y la enfermedad humanas. Aquí presentamos un ensayo versátil de actividad de SAMHD1 acoplado a enzimas, desplegado en un formato de microplaca de 384 pocillos, que permite la evaluación de moléculas pequeñas y análogos de nucleótidos como sustratos, activadores e inhibidores de SAMHD1.
Resumen
El motivo alfa estéril y la proteína 1 que contiene el dominio de la EH (SAMHD1) es un regulador fundamental de los grupos intracelulares de trifosfato de desoxinucleósidos (dNTP), ya que esta enzima puede hidrolizar los dNTP en sus correspondientes nucleósidos y trifosfatos inorgánicos. Debido a su papel crítico en el metabolismo de los nucleótidos, su asociación a diversas patologías y su papel en la resistencia a la terapia, actualmente se están llevando a cabo intensas investigaciones para comprender mejor tanto la regulación como la función celular de esta enzima. Por esta razón, es vital el desarrollo de métodos sencillos y económicos de alto rendimiento para sondear la interacción de moléculas pequeñas con SAMHD1, como reguladores alostéricos, sustratos o inhibidores. Para ello, el ensayo verde de malaquita acoplado a enzimas es un ensayo colorimétrico simple y robusto que se puede implementar en un formato de placa de 384 micropocillos que permite la medición indirecta de la actividad de SAMHD1. Como SAMHD1 libera el grupo trifosfato de los sustratos de nucleótidos, podemos acoplar una actividad pirofosfatasa a esta reacción, produciendo así fosfato inorgánico, que puede ser cuantificado por el reactivo verde de malaquita a través de la formación de un complejo verde de malaquita de fosfomolibdato. En este trabajo se muestra la aplicación de esta metodología para caracterizar inhibidores conocidos de SAMHD1 y descifrar los mecanismos implicados en la catálisis de sustratos no canónicos por SAMHD1 y su regulación por activadores alostéricos, ejemplificados por fármacos anticancerígenos basados en nucleósidos. Por lo tanto, el ensayo verde de malaquita acoplado a enzimas es una herramienta poderosa para estudiar SAMHD1 y, además, también podría utilizarse en el estudio de varias enzimas que liberan especies de fosfato.
Introducción
El motivo alfa estéril y la proteína 1 que contiene el dominio histidina-aspartato (SAMHD1) es un regulador central de la homeostasis de nucleótidos en células de mamíferos1 con muchas funciones en la salud y la enfermedadhumanas 2. Esta enzima es capaz de hidrolizar trifosfatos desoxinucleósidos (dNTPs) en sus moléculas afines de desoxinucleósido y trifosfato inorgánico 3,4, siendo esta actividad regulada alostéricamente por la abundancia de (d)NTP (revisado en la referencia5). Cada monómero SAMHD1 contiene dos sitios alostéricos (AS1 y AS2) y un sitio catalítico, y la formación de la enzima activa requiere el ensamblaje ordenado de un homotetrámero tras la unión de (d)NTP. La dimerización de los monómeros SAMHD1 se desencadena primero a través de la unión de un trifosfato de guanina (GTP o dGTP) a AS1, y la tetramerización posterior se logra cuando una molécula dNTP adicional se une a AS2, lo que permite el acceso del sustrato al sitio catalítico y la posterior hidrólisis.
Los sustratos de SAMHD1 incluyen los cuatro dNTPs canónicos 3,4 junto con algunos nucleótidos modificados con base y azúcar, incluyendo los metabolitos trifosfato de varios fármacos basados en nucleósidos utilizados en el tratamiento de infecciones virales y cáncer, varios de los cuales también pueden servir como activadores alostéricos 6,7,8,9,10,11. En consecuencia, SAMHD1 modula la eficacia de muchos de estos compuestos en modelos de enfermedad 7,8,9,10,11,12,13,14,15 y, además, en el caso del análogo de la desoxicitidina citarabina (ara-C), que se ha mantenido como tratamiento estándar para la leucemia mieloide aguda (LMA) durante décadas, en realidad dicta eficacia del tratamiento en esta enfermedad 7,8,16. Por lo tanto, SAMHD1 es un biomarcador potencial y una diana terapéutica para mejorar la eficacia de las terapias basadas en nucleósidos17 y, en consecuencia, nosotros y otros hemos tratado de identificar estrategias para inactivar SAMHD1 en las células. Propusimos el uso de la proteína viral X (Vpx) como inhibidor biológico para dirigirse a SAMHD1 para su degradación dentro de las células cancerosas7, sin embargo, este enfoque tiene una serie de limitaciones (discutidas en la referencia12), y también informamos recientemente un enfoque indirecto para suprimir la actividad de SAMHD1 a través de la inhibición de la ribonucleótido reductasa que demostramos en varios modelos de LMA18. Varios estudios han buscado identificar moléculas pequeñas capaces de inhibir directamente la SAMHD1 y, hasta la fecha, se han reportado varias moléculas de este tipo, sin embargo, solo se ha documentado la inhibición in vitro 6,9,19,20,21,22. En consecuencia, la falta de moléculas pequeñas que inhiban de forma potente la actividad de SAMHD1 en las células, junto con los complejos mecanismos de catálisis de SAMHD1 de las terapias basadas en nucleósidos, subraya la necesidad de seguir investigando. Por lo tanto, los métodos robustos e idealmente de alto rendimiento para sondear la interacción de moléculas pequeñas con SAMHD1 son ideales para identificar sustratos, reguladores alostéricos e inhibidores de esta enzima clínicamente relevante.
Existen varias metodologías que miden directamente la actividad dNTPasa de SAMHD1, como la cromatografía en capa fina (TLC)9,20,23 y la cromatografía líquida de alta resolución (HPLC)9,21, pero no son fácilmente susceptibles de configuraciones de alto rendimiento. Una excepción es el ensayo reportado por Mauney et al., que explota la capacidad de SAMHD1 para hidrolizar bis (4-nitrofenil) fosfato (b4NPP) a p-nitrofenol y p-nitrofenil fosfato cuando se usa Mn2+ como catión activador, lo que resulta en un cambio colorimétrico que se puede medir fácilmente en una placa de micropocillos21. Este ensayo se ha utilizado con éxito para la identificación y caracterización de inhibidores de SAMHD1, pero debe tenerse en cuenta que la hidrólisis se produce en ausencia de activadores de (d)NTP y en presencia de un probable catión activador no fisiológico, siendo ambas advertencias importantes a tener en cuenta. Esto también hace que este ensayo sea menos aplicable al estudio e identificación de reguladores alostéricos de SAMHD1.
En este contexto, un enfoque acoplado a enzimas combinado con el reactivo verde de malaquita, como se detalla en este informe, puede ser un método versátil para medir indirectamente la actividad de la dNTPasa de SAMHD1 y, además, interrogar el impacto de varias moléculas pequeñas sobre ella. El ensayo de verde de malaquita es una técnica colorimétrica robusta y fiable para la detección de fosfato inorgánico libre (Pi), basada en la formación de un complejo de ácido molibdofosfórico que conduce a un cambio colorimétrico medido a 620 nm24. Como la hidrólisis de SAMHD1 libera el grupo trifosfato de los sustratos de nucleótidos, es necesario acoplar esta reacción con una actividad de (piro)fosfatasa, que generará fosfato inorgánico libre, antes de la adición del reactivo verde de malaquita. El ensayo de verde de malaquita es sensible y rentable y se ha utilizado ampliamente para la identificación y caracterización de inhibidores y sustratos para enzimas que liberan grupos fosfato inorgánicos, ya sea en sus reacciones o en presencia de una enzima de acoplamiento. Se ha aplicado ampliamente en la caracterización de las actividades ATPasa de las helicasas 25,26,27, o en el estudio de la actividad enzimática CD73, que media la degradación del AMP a adenosina y fosfato inorgánico28. Además, cuando se acopla, se ha empleado en el descubrimiento de antibióticos dirigidos a la UDP-2,3-diacilglucosamina pirofosfatasa LpxH, una enzima esencial en la mayoría de los patógenos gramnegativos29. Con respecto a la investigación del cáncer, el enfoque acoplado a enzimas se ha desplegado ampliamente contra las hidrolasas NUDIX, una familia de enzimas metabolizadoras de nucleótidos, tanto en la caracterización de sustratos 30,31,32 como en la identificación y desarrollo de fármacos y sondas químicas 33,34,35,36.
Con respecto a la dNTPasa SAMHD1, este enfoque se ha utilizado en varios informes. Utilizando la exopolifosfatasa Ppx1 de Saccharomyces cerevisiae como enzima de acoplamiento, este ensayo se utilizó para probar varios análogos de nucleótidos como sustratos, activadores o inhibidores de SAMHD1, y dio como resultado la identificación del metabolito trifosfato del fármaco antileucémico clofarabina como activador y sustrato6. Además, con pirofosfatasa inorgánica de Escherichia coli como enzima de acoplamiento, se ha empleado en el cribado de una biblioteca de compuestos clínicamente aprobados contra SAMHD1 para identificar inhibidores20. En nuestra investigación, utilizamos este enfoque para demostrar que ara-CTP, el metabolito activo de ara-C, es un sustrato de SAMHD1 pero no un activador alostérico7 y, posteriormente, utilizamos este ensayo para demostrar que varias moléculas pequeñas que podían sensibilizar modelos de LMA a ara-C de una manera dependiente de SAMHD1, en realidad no inhibían directamente SAMHD118. En este informe, detallaremos este método versátil y demostraremos su aplicabilidad, en una configuración de alto rendimiento, para la identificación de inhibidores, activadores y sustratos de SAMHD1.
Protocolo
En la Figura 1 se muestra una descripción general esquemática de los métodos a continuación y una lista detallada de materiales y reactivos está disponible en la Tabla de materiales.
1. Preparación de tampones de ensayo.
- Preparación de buffers de stock.
NOTA: Como el ensayo es sensible a la detección de fosfatos, que puede ser común, enjuague la cristalería tres veces con agua ultrapura o bidestilada para evitar la contaminación. Todos los tampones se pueden almacenar a temperatura ambiente (RT).- Prepare 1 L de solución madre tampón de reacción (RB) SAMHD1 (25 mM de Tris-Acetato pH 8, 40 mM NaCl, 1 mM MgCl2) disolviendo 4,5 g de acetato de Tris, 2,3 g de NaCl y 0,2 g de MgCl2, en aproximadamente 800 ml de agua antes de ajustar al pH 8 y al volumen final.
- Prepare 5 ml de solución madre de TCEP 0,1 M diluyendo 1 ml de TCEP 0,5 M en 4 ml de agua.
- Prepare 50 ml de solución madre de Tween-20 al 11 % diluyendo 5 ml de Tween-20 al 100 % en 44,5 ml de agua.
NOTA: Tween-20 es sensible a la luz. - Prepare 50 ml de solución de parada de EDTA 0,5 M disolviendo 9,3 g de EDTA en aproximadamente 40 ml de agua antes de ajustar a pH 8 y volumen final.
- Prepare la solución madre de verde de malaquita (MG) (3,2 mM de verde de malaquita enH2, SO,4) agregando lentamente 60 ml de ácido sulfúrico concentrado a 300 ml de agua en una botella de vidrio marrón. Enfriar la solución a RT y disolver 0,44 g de verde de malaquita.
CAUTELA: La reacción del ácido sulfúrico con el agua es exotérmica, por lo que la botella puede calentarse y provocar una acumulación de presión; Asegúrese de que esta presión se libere con frecuencia.
NOTA: La solución naranja resultante es sensible a la luz (por lo tanto, botella marrón) y estable durante al menos 1 año en RT. El precipitado puede formarse con el tiempo, asegúrese de que solo se use el sobrenadante. - Preparar 50 ml de solución madre de molibdato amónico al 7% disolviendo 3,75 g de molibdato amónico en 50 ml de agua.
NOTA: El precipitado puede formarse con el tiempo, asegúrese de usar solo el sobrenadante.
- Preparación de tampones de ensayo completos
NOTA: Esto debe hacerse el día del experimento- Prepare SAMHD1 RB completo (25 mM de Tris-Acetato pH 8, 40 mM de NaCl, 1 mM de MgCl2, 0,3 mM de TCEP, 0,005% de Tween-20). Utilice existencias de TCEP de 11% para Tween-20 y 0,1 M previamente preparadas para añadir estos componentes a una concentración final de 0,005% para Tween-20 y 0,3 mM para TCEP a la acción de SAMHD1 RB.
- Prepare la solución de parada de EDTA (25 mM de Tris-Acetato pH 8, 40 mM de NaCl, 1 mM de MgCl2, 0,3 mM de TCEP, 0,005% de Tween-20, 7,9 mM de EDTA). Para completar SAMHD1 RB, utilice una solución madre de EDTA 0,5 M para añadir EDTA a una concentración final de 7,9 mM.
- Prepare la solución de trabajo de MG (2,5 mM de verde de malaquita, 1,4% de molibdato de amonio, 0,18% de Tween-20) mezclando 10 partes de solución madre de MG con 2,5 partes de molibdato de amonio al 7% y 0,2 partes de Tween-20 al 11%.
2. Ensayo de inhibición de SAMHD1 y determinación del compuesto IC50
NOTA: Las condiciones finales del ensayo se muestran en la Tabla 1.
- Preparación de compuestos en placa de ensayo
NOTA: Los compuestos de peso molecular pequeño generalmente se disuelven en DMSO al 100% y análogos de nucleótidos en agua. La concentración de stock oscila entre 10 y 100 mM y está influenciada por la potencia y la solubilidad de los compuestos, junto con la tolerancia al DMSO del ensayo. Compruebe que la concentración final de DMSO en la reacción no supere el 1% para asegurarse de que las actividades enzimáticas no se vean afectadas por este disolvente. Es una buena práctica probar la tolerancia del ensayo al disolvente antes del experimento.- Prepare compuestos de prueba diluidos en serie a una concentración final de 100 veces en el disolvente correspondiente (por ejemplo, 100 % de DMSO para moléculas pequeñas o agua para análogos de nucleótidos) en una placa transparente de polipropileno de 96 pocillos de fondo redondo utilizando una pipeta multicanal o un equipo automatizado de manipulación de líquidos.
NOTA: Dependiendo de la estabilidad del compuesto, las placas de dilución pueden prepararse con antelación, sellarse y almacenarse a -20 °C. Deje que las placas se equilibren a RT antes de continuar con el protocolo. - Utilizando SAMHD1 RB completo, diluya los compuestos hasta una concentración final de 25 veces (para mantener la concentración final de disolvente por debajo del 1 %) y transfiera 5 μl a los pocillos apropiados de una placa de ensayo transparente de fondo plano de 384 pocillos. Repita el procedimiento con muestras de control solo con disolvente.
- Prepare compuestos de prueba diluidos en serie a una concentración final de 100 veces en el disolvente correspondiente (por ejemplo, 100 % de DMSO para moléculas pequeñas o agua para análogos de nucleótidos) en una placa transparente de polipropileno de 96 pocillos de fondo redondo utilizando una pipeta multicanal o un equipo automatizado de manipulación de líquidos.
- Preparación de los componentes de la reacción
NOTA: Esto debe hacerse el día del ensayo. Las alícuotas recombinantes humanas de SAMHD1 y E. coli pirofosfatasa (PPasa) se almacenan a largo plazo a -80 °C y se diluyen a 9,1 mg/ml y 23,0 mg/ml, respectivamente, en tampón de almacenamiento (20 mM HEPES pH 7,5, 300 mM NaCl, 10% glicerol, 2 mM TCEP). Una vez descongeladas, las alícuotas se almacenan a corto plazo a -20 °C.- Prepare la mezcla maestra de la enzima (SAMHD1/PPasa) diluyendo la proteína SAMHD1 humana recombinante y la PPasa recombinante en SAMHD1 RB completa hasta 4 veces la concentración final deseada, es decir, 1,4 μM de SAMHD1 y 50 U/mL de PPasa.
- Prepare el dGTP activador/sustrato diluyendo el material de dGTP (normalmente 10 o 100 mM en agua) en SAMHD1 RB completo a una concentración final de 2x, es decir, 50 μM de dGTP.
- Realizar el ensayo
NOTA: Todos los componentes del ensayo deben equilibrarse a RT. Las adiciones de líquido se pueden realizar con una pipeta multicanal o un dispensador de líquido reactivo a granel.- En una placa de ensayo de 384 pocillos que contenga diluciones de compuestos y controles de solo disolvente, dispense 5 μL de mezcla maestra SAMHD1/PPasa. A los pocillos de control no enzimáticos, dispense 5 μL de SAMHD1 RB completo. Preincubar la enzima y los compuestos durante 10 min en RT.
- A todos los pocillos, dispense 10 μL de solución 2x dGTP para iniciar la reacción.
- Incubar la reacción durante 20 min a RT.
- Detenga la reacción dispensando 20 μL de solución de tope de EDTA a todos los pocillos.
NOTA: El experimento se puede pausar aquí si se desea. - Agregue 10 μL de solución de trabajo MG a todos los pocillos.
CAUTELA: La solución de trabajo MG contiene ácido sulfúrico. - Asegure la mezcla del contenido del pocillo utilizando un agitador orbital de placas de micropocillos y la centrifugación a 1.000 x g durante 1 min.
- Incubar la placa durante 20 min a RT.
- Lea la absorción a una longitud de onda de 630 nm en un lector de placas de micropocillos.
- Visualización y análisis de datos
- Calcular la media y la desviación estándar de los pocillos de control positivo y negativo (positiva, reacción completa con disolvente; negativa, dGTP solo con disolvente). Calcule el factorZ 37 como indicador de la calidad del ensayo.
- Normalice cada valor de absorbancia a los valores medios de los controles positivos y negativos, estableciendo el control positivo como 100% de actividad SAMHD1 y el control negativo como 0% de actividad SAMHD1.
- Grafique la actividad de SAMHD1 (%) en función de la concentración del compuesto y ajuste una curva dosis-respuesta de pendiente variable de cuatro parámetros, lo que permite la determinación del compuesto IC50.
3. Activador SAMHD1 y pantalla de sustrato
NOTA: Las condiciones finales del ensayo se muestran en la Tabla 2. Las alícuotas recombinantes de SAMHD1 y PPasa se almacenan a largo plazo a -80 °C diluidas a 9,1 mg/ml y 23,0 mg/ml, respectivamente, en tampón de almacenamiento (20 mM HEPES pH 7,5, 300 mM NaCl, 10% glicerol, 2 mM TCEP) a -80 °C. Una vez descongeladas, las alícuotas se almacenan a corto plazo a -20 °C.
- Preparación de análogos de nucleótidos en placa de ensayo
- Diluir las existencias de análogos de nucleótidos (normalmente 10 o 100 mM en agua) a una concentración final de 4 veces en SAMHD1 RB completo, en nuestro caso un análogo de nucleótido de 800 μM, y transferir 5 μL a los pocillos apropiados de una placa de ensayo de 384 pocillos.
- Preparación de los componentes de la reacción
NOTA: Esto debe hacerse el día del ensayo- Prepare la mezcla maestra de la enzima (SAMHD1/PPasa) diluyendo la proteína SAMHD1 humana recombinante y la PPasa recombinante de E. coli en SAMHD1 RB completa hasta 2 veces la concentración final deseada, es decir, 0,7 μM de SAMHD1 y 25 U/mL de PPasa.
- Prepare la solución de PPasa sola diluyendo la PPasa recombinante de E. coli en SAMHD1 RB completa a 2 veces la concentración final deseada, por lo tanto, 25 U/mL de PPasa.
- Prepare activadores GTP (AS1) y dGTPαS (AS1 y AS2) diluyendo material (normalmente 10 o 100 mM en agua) en SAMHD1 RB completo a una concentración final de 4x, por lo tanto, 50 μM GTP o dGTPαS.
- Realizar el ensayo
NOTA: Todos los componentes del ensayo deben equilibrarse a RT. Las adiciones de líquido se pueden realizar con una pipeta multicanal o un dispensador de líquido reactivo a granel.- En una placa de ensayo de 384 pocillos que contenga análogos de nucleótidos, dispense 5 μL del activador (ya sea GTP o dGTPαS) o complete SAMHD1 RB a los pocillos apropiados.
- Inicie la reacción dispensando 10 μL de mezcla maestra SAMHD1/PPasa, PPasa sola o SAMHD1 RB completa a los pocillos apropiados.
- Incubar la reacción durante 20 min a RT.
- Detenga la reacción dispensando 20 μL de solución de parada de EDTA a todos los pocillos.
NOTA: El experimento se puede pausar aquí si se desea. - Agregue 10 μL de MG de solución de trabajo a todos los pocillos.
CAUTELA: La solución de trabajo MG contiene ácido sulfúrico. - Asegure la mezcla del contenido del pocillo utilizando un agitador orbital de placas de micropocillos y la centrifugación a 1.000 x g durante 1 min.
- Incubar la placa durante 20 min a RT.
- Lea la absorción a una longitud de onda de 630 nm en un lector de placas de micropocillos.
- Visualización y análisis de datos
- Calcule los valores medios de absorbancia para los pocillos de reacción de PPase (control negativo o señal de fondo).
NOTA: Como control positivo de un activador alostérico SAMHD1 y sustrato, se puede incluir dGTP en la placa. En este caso, puede utilizar esta condición para calcular el factor Z como indicador de la calidad del ensayo. - Reste el valor de fondo de los pocillos correspondientes en las reacciones SAMHD1/PPase.
- Grafique los valores de absorbancia corregidos para cada análogo de nucleótido con condiciones de tampón, GTP y dGTPαS.
- Calcule los valores medios de absorbancia para los pocillos de reacción de PPase (control negativo o señal de fondo).
Resultados
El protocolo descrito en la Figura 1 describe el flujo de trabajo básico para utilizar el ensayo de verde de malaquita acoplado a enzimas para sondear la interacción de moléculas pequeñas con la dNTPasa SAMHD1 y se puede adaptar de varias maneras para interrogar diferentes cuestiones bioquímicas. En los resultados representativos discutidos en los siguientes párrafos, ilustramos ejemplos del uso de este ensayo para determinar las propiedades inhibitorias de moléculas pequeñas hacia SAMHD1 y para probar si diferentes análogos de nucleótidos son sustratos y/o activadores de esta enzima.
Los resultados que se muestran en la Figura 2 ilustran varios principios básicos de este ensayo. El reactivo verde de malaquita permite la detección colorimétrica de fosfato inorgánico a través de la formación de un complejo verde de malaquita fosfomolibdato y, en consecuencia, este enfoque se puede aplicar al estudio de reacciones enzimáticas cuyo producto es fosfato. Para demostrar la sensibilidad de este método para detectar fosfato inorgánico libre, en la Figura 2A se muestran los valores de absorbancia obtenidos con concentraciones crecientes de Na3PO4 después de una incubación de 20 min con el reactivo verde de malaquita. Mientras que la señal alcanza la saturación a 0,25 mM Na3PO4, el rango de detección lineal del fosfato es visible de 0,004 a 0,03 mM (Figura 2A, panel derecho), de acuerdo con otros estudios que reportaron un rango lineal de fosfato de hasta 10-20 μM utilizando el ensayo de verde de malaquita38.
SAMHD1 es una dNTPasa que libera trifosfato inorgánico al hidrolizar una molécula de dNTP y, por lo tanto, para generar fosfato inorgánico libre para su detección por verde de malaquita, se requiere una enzima de acoplamiento. La pirofosfatasa inorgánica (PPasa) de E. coli ha demostrado ser útil para este propósito, tanto con respecto a SAMHD1 7,20, como a otras enzimas metabolizadoras de nucleótidos 30,33,35. Además, SAMHD1 es una dNTPasa activa cuando es homotetrámero, y esto requiere la activación alostérica por (d)NTPs, específicamente un trifosfato de guanina (GTP o dGTP) en AS1 y cualquier dNTP en AS2. Posteriormente, el sitio catalítico se vuelve accesible para la unión del sustrato y se produce la reacción enzimática. Dado que dGTP cumple los requisitos para unirse a AS1 y AS2, y es un sustrato, el uso de este nucleótido en el ensayo de inhibición simplifica enormemente el flujo de trabajo. La Figura 2B ilustra el requisito de los diferentes componentes del ensayo para lograr una actividad medible de SAMHD1 indicada por un aumento en la absorbancia a 630 nm. Ni SAMHD1 ni PPasa por sí solas son capaces de generar fosfato inorgánico en presencia de dGTP, lo que es consistente con las actividades documentadas de estas enzimas. Sin embargo, en la condición en la que todos los componentes del ensayo están presentes (SAMHD1, PPasa y el activador/sustrato dGTP) observamos un aumento de la señal. El factor Z37 del ejemplo que se muestra aquí (sin tomar enzimas + dGTP como control negativo y SAMHD1/PPasa + dGTP como control positivo) fue de 0,74, lo que indica un ensayo robusto.
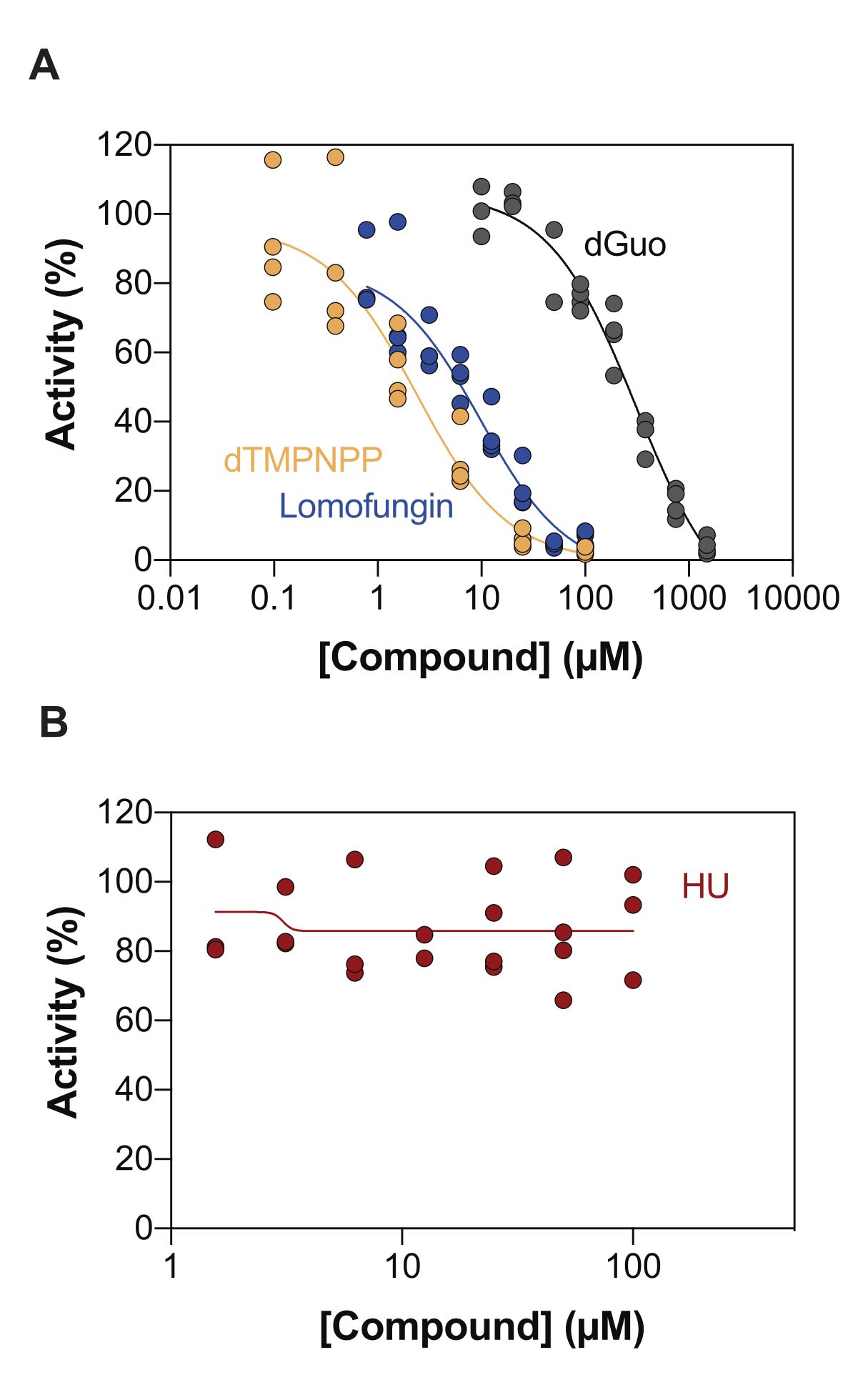
Una de las aplicaciones potenciales del ensayo de actividad SAMHD1 acoplado a enzimas es la identificación de inhibidores a través del cribado de alto rendimiento (HTS). Por lo tanto, en este informe, validamos la detección de la inhibición de SAMHD1 en este ensayo utilizando un conjunto diverso de compuestos ya descritos en la literatura. Seamon et al. evaluaron la inhibición dependiente de la dosis de nucleósidos canónicos hacia SAMHD1 utilizando un ensayo similar al que se muestra aquí, y encontraron que la desoxiguanosina (dGuo) era el único nucleósido canónico capaz de inhibir significativamente SAMHD1, con un valor de IC50 de 488 μM20. Un HTS de fármacos aprobados por la FDA realizado con el ensayo directo b4NPP reveló varios resultados que inhibieron la actividad de SAMHD1 a concentraciones micromolares, de los cuales la lomofungina fue la molécula que inhibió más potentemente la actividad de la dNTPasa de SAMHD1 in vitro, exhibiendo un IC50 de 20,1 μM cuando se determinó en presencia de dGTP como sustrato21. Además, los cuatro análogos de α,β-imido-dNTP también se han identificado como inhibidores competitivos de SAMHD1 utilizando el sensor MDCC-PBP y SAMHD1 acoplado a la actividad de Ppx, lo que mostró que las constantes inhibitorias de los análogos de dNMPNPP estaban en el rango micromolar bajo / nanomolar alto 6,22. Por lo tanto, para demostrar que el ensayo de actividad de SAMHD1 acoplado a enzimas se puede utilizar para identificar inhibidores de SAMHD1, se utilizaron dGuo, lomofungina y 2'-desoxitimidina-5'-[(α,β)-imido]trifosfato (dTMPNPP), para validar la técnica. La Figura 3A ilustra las curvas dosis-respuesta obtenidas para estos compuestos, mostrando que el aumento de las concentraciones inhibe eficazmente la actividad de SAMHD1. Los valores medios de IC50 obtenidos para estas moléculas en tres experimentos independientes (± desviación estándar) fueron los siguientes: dGuo = 361,9 ± 72,8 μM, lomofungina 6,78 ± 3,9 μM y dTMPNPP = 2,10 ± 0,9 μM. Como ejemplo de resultado negativo, también se determinó el impacto de la hidroxiurea (HU) en la actividad de SAMHD1. El HU es un inhibidor de la ribonucleótido reductasa y, aunque limita la actividad de la ara-CTPasa de SAMHD1 en varios modelos de LMA, se demostró que los efectos del HU sobre SAMHD1 son indirectos y dependen de la perturbación de la regulación alostérica de SAMHD118. La curva dosis-respuesta de HU se muestra en la Figura 3B, y no se observaron cambios en la actividad de SAMHD1 con el aumento de las dosis de HU, lo que demuestra que HU no inhibe la actividad de SAMHD1 in vitro.
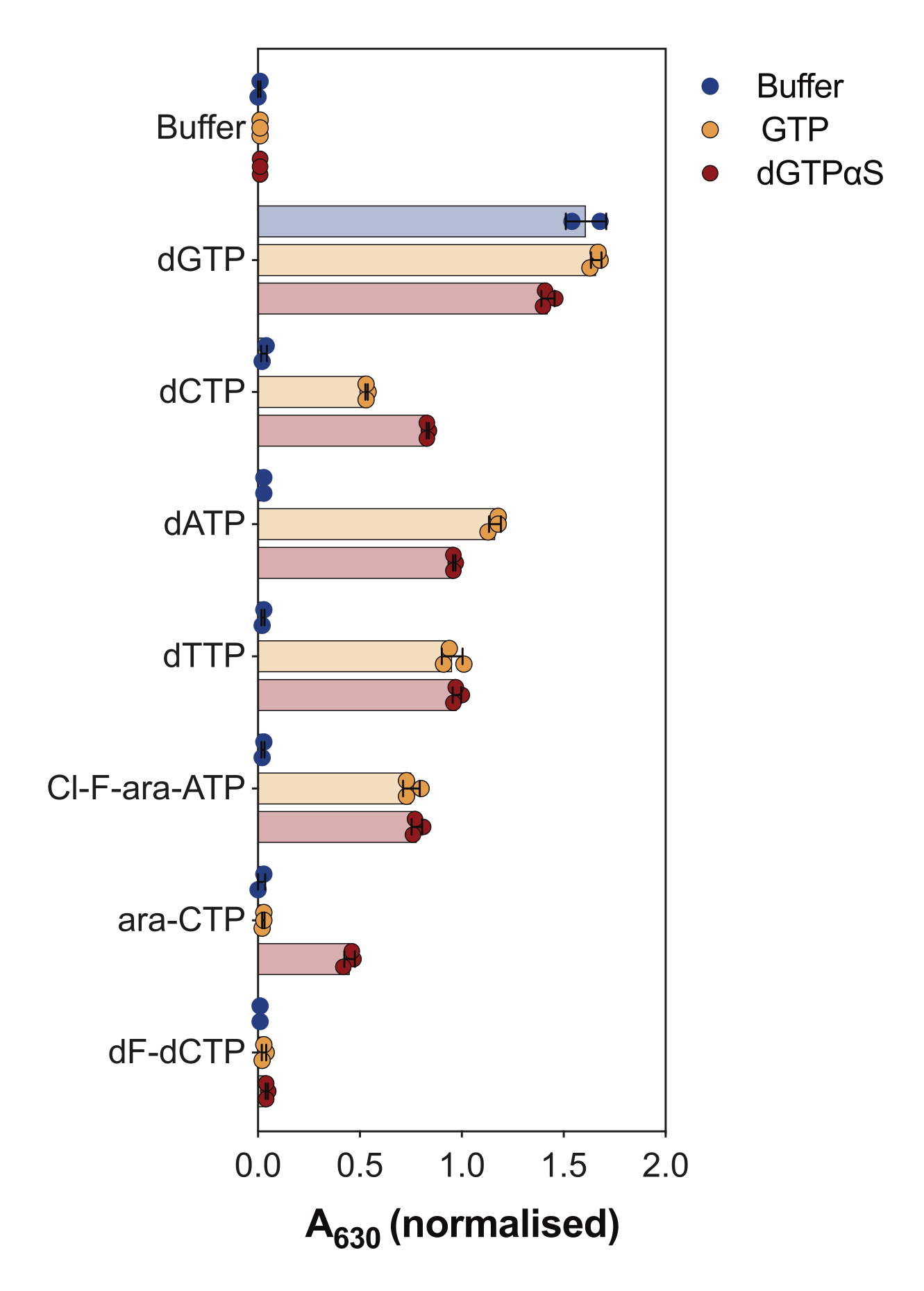
Otro uso del ensayo de actividad SAMHD1 acoplado a enzima es interrogar si los nucleótidos y sus análogos son sustratos y/o activadores alostéricos de esta enzima, lo que se ilustra en la Figura 4. En este experimento, se probaron nucleótidos canónicos, así como los metabolitos activos de varios análogos de nucleósidos anticancerígenos, como la citarabina (ara-CTP), la clofarabina (Cl-F-ara-ATP) y la gemcitabina (dF-dCTP), como sustratos y activadores de SAMHD1. Debido a la compleja regulación alostérica de SAMHD1, la reacción se realiza en presencia de GTP como activador de AS1 o del análogo no hidrolizable de dGTP 2'-desoxiguanosina-5'-(α-tio)-trifosfato (dGTPαS), que puede ocupar AS1 y AS2. La actividad de SAMHD1 en presencia del análogo de nucleótido probado y GTP indica que el nucleótido es capaz de unirse al sitio alostérico secundario y al sitio catalítico (es decir, el activador AS2 y el sustrato), mientras que la actividad de SAMHD1 con el análogo de nucleótido y dGTPαS indica que el nucleótido solo puede ocupar el sitio catalítico (es decir, solo un sustrato). Si el nucleótido es capaz de unirse a los sitios alostéricos AS1 y AS2 y al sitio catalítico, SAMHD1 estará activo en presencia del nucleótido solo, como se muestra en el caso de dGTP. Los resultados muestran que todos los dNTP canónicos son capaces de unirse al sitio AS2 y al sitio catalítico. En el caso de los análogos de nucleótidos, el trifosfato de clofarabina es un activador de AS2 y un sustrato, mientras que el trifosfato de citarabina solo es capaz de ocupar el sitio catalítico. Por otro lado, no se observó actividad con el trifosfato de gemcitabina, lo que sugiere que en las condiciones probadas el trifosfato de gemcitabina no es capaz de actuar como efector alostérico ni como sustrato. Aunque este resultado es consistente con predicciones previas9, estudios posteriores de cristalización y cinética10 revelaron que el trifosfato de gemcitabina es capaz de unirse a la bolsa catalítica SAMHD1, y que es de hecho un sustrato de la enzima. Sin embargo, en este último estudio10, los autores muestran que la tasa de hidrólisis es considerablemente menor en comparación con otros sustratos reportados, como el trifosfato de citarabina, lo que explica por qué no pudimos observar esto con esta configuración de cribado.
En conjunto, estos resultados representativos validan el uso del ensayo de actividad SAMHD1 acoplado a enzimas como una técnica robusta para la identificación y caracterización de inhibidores, reguladores alostéricos y sustratos de SAMHD1. Sin embargo, al igual que todos los enfoques experimentales, este método tiene sus advertencias, por lo que se deben utilizar ensayos ortogonales (por ejemplo, utilizando una tecnología de ensayo diferente) para validar aún más los hallazgos.

Figura 1: Descripción general esquemática del protocolo descrito en este artículo. Haga clic aquí para ver una versión más grande de esta figura.
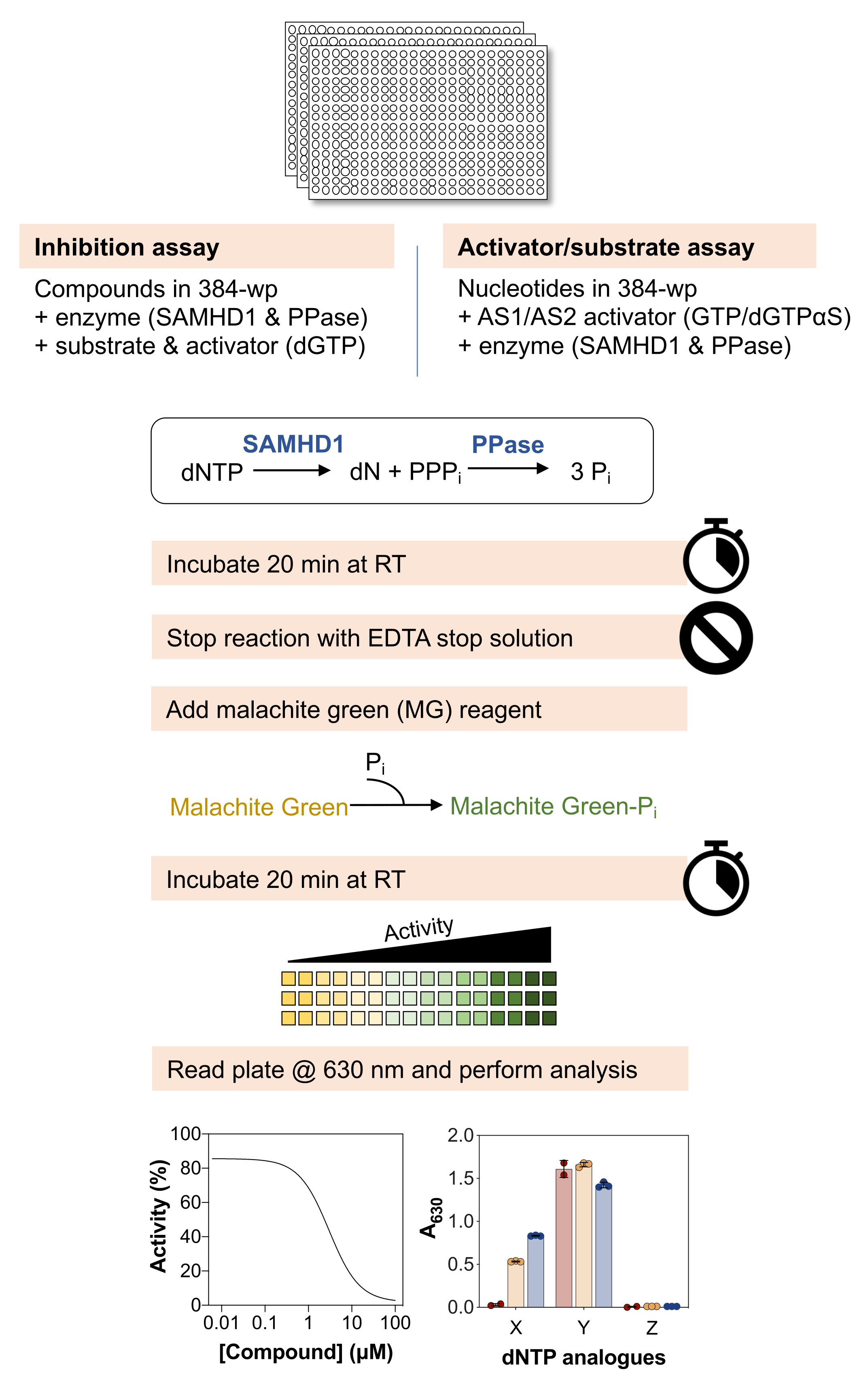{kind=link}

Figura 2: Ensayo de actividad SAMHD1 acoplado a enzimas. (A) Curva estándar Na3PO4 en el ensayo de verde de malaquita. Se preparó una dilución seriada de Na3PO4 (2 veces) de 1 mM a 0,004 mM por triplicado y se incubó con reactivo verde de malaquita durante 20 min. Los valores de absorbancia bruta en todo el rango de concentraciones probadas se muestran en el panel izquierdo y el rango lineal en el panel derecho. Representativo de dos experimentos independientes mostrados. (B) Validación del ensayo de actividad acoplada a enzimas. SAMHD1 (0,35 μM) y/o PPasa (12,5 U/mL) en presencia o ausencia de activador/sustrato dGTP (25 μM) se incubaron durante 20 min en el ensayo de actividad acoplada a enzimas. Los cuatrillizos de un representante de dos experimentos independientes se muestran con valores de absorbancia sin procesar trazados, barras y barras de error que indican la media y la desviación estándar. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Evaluación de compuestos para la inhibición de SAMHD1 en el ensayo de actividad acoplada a enzimas. Respuesta a la dosis de lomofungina (0,78-100 μM), 2'-desoxitimidina-5'-[(α,β)-imido]trifosfato (dTMPNPP, 0,01-100 μM) y desoxiguanosina (dGuo, 10-1.500 μM) (A) o hidroxiurea (HU) (0,78-100 μM) (B) en el ensayo de actividad SAMHD1 acoplado a enzimas con dGTP (25 μM) como activador/sustrato. Porcentaje de actividad relativa a los controles de reacción de réplicas individuales graficadas (DMSO + SAMHD1/PPase + dGTP = 100% de actividad, DMSO + dGTP = 0% de actividad) con un representante de tres experimentos mostrados. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Evaluación de análogos de nucleótidos como activadores alostéricos y sustratos de SAMHD1 en el ensayo de actividad acoplada a enzimas. Los nucleótidos canónicos y los metabolitos trifosfato seleccionados de los fármacos anticancerígenos citarabina (ara-CTP), clofarabina (Cl-F-ara-ATP) y gemcitabina (dF-dCTP) se probaron a 200 μM en el ensayo de actividad SAMHD1 acoplado a enzimas en presencia o ausencia de GTP o dGTP análogo de dGTP no hidrolizable dGTPαS (12,5 μM). Se indican los valores de absorbancia normalizados de las réplicas experimentales individuales graficadas, la media y la DE. Representativo de dos experimentos independientes mostrados, adaptados de nuestro estudio anterior7. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Paso | Reactivo | Volumen dispensado (μL) | Volumen de reacción final (μL) | Concentración dispensada | Dilución de pliegues en reacción | Concentración final en reacción |
| 1 | Inhibidor | 5 | 20 | 0,4 mM | 4 | 0,1 mM |
| 2 | Mezcla SAMHD1+PPasa | 5 | 1,4 μM SAMHD1, 50 U/mL PPasa | 4 | 0,35 μM SAMHD1, 12,5 U/mL de Ppasa | |
| 3 | dGTP | 10 | 50 μM | 2 | 25 μM | |
| 4 | Incubación durante 20 minutos | |||||
| 5 | Solución EDTA | 20 | 40 | 7,9 mM | 2 | 3,95 mM |
| 6 | Reactivo MG | 10 | 50 | 2,5 mM de verde de malaquita, 64,4 mM de molibdato de amonio, 0,18% Tween-20 | 5 | 0,5 mM verde de malaquita, 12,9 mM de molibdato de amonio, 0,036% Tween-20 |
| 7 | Incubación durante 20 minutos | |||||
| 8 | Lectura @ 630 nm | |||||
Tabla 1: Resumen de las condiciones finales en el ensayo acoplado a enzimas para el cribado de inhibidores.
| Paso | Reactivo | Volumen dispensado (μL) | Volumen de reacción final (μL) | Concentración dispensada | Dilución de pliegues en reacción | Concentración final en reacción |
| 1 | Regulador alostérico | 5 | 20 | 800 μM | 4 | 200 μM |
| 2 | GTP o dGTPαS | 5 | 50 μM | 4 | 12,5 μM | |
| 3 | SAMHD1 y/o PPase | 10 | 0,7 μM SAMHD1, 25 U/mL PPasa | 2 | 0,35 μM SAMHD1, 12,5 U/mL PPasa | |
| 4 | Incubación durante 20 minutos | |||||
| 5 | Solución EDTA | 20 | 40 | 7,9 mM | 2 | 3,95 mM |
| 6 | Reactivo MG | 10 | 50 | 2,5 mM de verde de malaquita, 64,4 mM de molibdato de amonio, 0,18% Tween-20 | 5 | 0,5 mM verde de malaquita, 12,9 mM de molibdato de amonio, 0,036% Tween-20 |
| 7 | Incubación durante 20 minutos | |||||
| 8 | Lectura @ 630 nm | |||||
Tabla 2: Resumen de las condiciones finales en el ensayo acoplado a enzimas para el cribado de reguladores alostéricos
Discusión
El ensayo de actividad acoplada a enzimas que se detalla aquí es un ensayo colorimétrico de alto rendimiento que permite la medición indirecta de la hidrólisis de dNTP mediante SAMHD1. Este método aprovecha la capacidad de la PPasa inorgánica de E. coli, que cuando se incluye en exceso en la mezcla de reacción, convierte cada trifosfato inorgánico generado por SAMHD1 en tres fosfatos libres individuales que se pueden cuantificar utilizando el reactivo verde de malaquita simple y económico. Proporcionamos este ensayo en un formato de placa de 384 micropocillos, que es ideal para el cribado de bibliotecas de compuestos, y demostramos la aplicabilidad y versatilidad de esta técnica en la identificación y caracterización de inhibidores, activadores y sustratos de SAMHD1.
Al igual que con todos los ensayos de cribado bioquímico in vitro , hay una serie de pasos críticos y consideraciones importantes, y muchos de ellos se analizan en profundidad en el Manual de orientación de ensayos39, disponible gratuitamente. La integridad de las enzimas recombinantes purificadas, tanto SAMHD1 como la enzima acoplada PPasa inorgánica, es extremadamente importante y debe confirmarse antes de establecer el ensayo. Y, en consecuencia, cada nueva purificación de estas enzimas debe estar sujeta a cierto nivel de pruebas por lotes, ya que las variabilidades de un lote a otro podrían introducir inconsistencias en los resultados. Idealmente, se debe utilizar un ensayo directo ortogonal, como la HPLC, que permite la detección tanto del sustrato como del producto de reacción, para verificar la actividad de la trifosfohidrolasa dNTP de la SAMHD1 recombinante purificada que se está utilizando.
En cuanto a las limitaciones de este ensayo, la principal es que mide la actividad dNTPasa de SAMHD1 de manera indirecta, explotando la actividad de la PPasa inorgánica, lo que tiene una serie de implicaciones. Es importante confirmar que la PPasa posee poca o ninguna actividad hacia los nucleótidos utilizados en el ensayo y, del mismo modo, que las pequeñas moléculas inhibidoras identificadas no poseen actividad hacia la PPasa. Por lo tanto, con respecto a la detección, una contradetección contra PPase puede ser una consideración importante. La presencia de PPasa en la reacción también hace que sea fundamental utilizar un ensayo ortogonal para confirmar los hallazgos. Con respecto a los ensayos de actividad directa, hasta la fecha se han descrito varios de ellos, incluidos TLC 9,20,23 y HPLC 9,21, que detectan con precisión el agotamiento del sustrato y la formación de productos. Además, el ensayo b4NPP21, que también es de alto rendimiento, podría utilizarse para probar posibles inhibidores; Sin embargo, no es ideal para probar sustratos o activadores alostéricos. Los ensayos biofísicos, como la fluorimetría diferencial de barrido (DSF), de la que hemos informado previamente con SAMHD118, también pueden ser especialmente potentes para identificar y caracterizar ligandos. Otra limitación del ensayo, específicamente como se muestra en la configuración aquí para identificar sustratos y activadores, es el uso del análogo dGTP no hidrolizable dGTPαS como activador de AS1 y AS2. Si bien esto permite la activación de SAMHD1 sin actividad observable en el ensayo, dGTPαS es un inhibidor competitivo de SAMHD1 y, por lo tanto, el uso de altas concentraciones inactivará la enzima. A medida que avanza nuestra comprensión de SAMHD1, los estudios futuros podrían utilizar moléculas que ocupen exclusivamente cada sitio de SAMHD1, negando así este problema potencial.
Como hemos mostrado aquí, este método es versátil y se puede utilizar para abordar una serie de cuestiones bioquímicas. Hemos descrito dos variaciones de este ensayo, una para la identificación de reguladores alostéricos y sustratos de SAMHD1, y otra para la caracterización de inhibidores, pero se pueden hacer más adaptaciones. Con respecto a los inhibidores potenciales, este ensayo, al estar basado en placas de micropocillos, lo hace muy adecuado para estudios posteriores de mecanismos de acción39,40. Del mismo modo, para una mayor caracterización de sustratos y reguladores alostéricos, esta técnica puede ser utilizada para determinar parámetros cinéticos de catálisis, como hicimos para el metabolito activo de citarabina y clofarabina7. Sin embargo, un inconveniente es que el ensayo acoplado a enzimas que se informa aquí es un ensayo de criterio de valoración y, por lo tanto, aunque es adecuado para el cribado, un ensayo continuo sería más adecuado para algunos estudios mecanicistas. Arnold et al. informaron de un ensayo continuo acoplado a enzimas que utiliza el biosensor MDCC-PBP6, que se basa en el uso de la proteína de unión al fosfato periplásmico (PBP) marcada con fluoróforo de maleimida de cumarina (MDCC) que puede unirse a un grupo fosfato libre. MDCC-PBP es muy sensible y permite la cuantificación de concentraciones de fosfato muy bajas, con un tiempo de respuesta del sensor de milisegundos a segundos.
SAMHD1 desempeña una serie de funciones importantes en la salud y la enfermedadhumanas 2 , muchas de las cuales podrían estar relacionadas con su papel central en el mantenimiento de los niveles intracelulares de dNTP1. Por lo tanto, la identificación de una sonda química de alta calidad hacia la actividad dNTPasa de SAMHD1 sería una herramienta poderosa para definir estos enlaces, y el ensayo acoplado a enzimas reportado aquí podría emplearse fácilmente para identificar tales sondas. Además, dado que los fármacos basados en nucleósidos, muchos de los cuales están modulados por SAMHD1, constituyen un grupo diverso e importante de fármacos terapéuticos41; Las sondas químicas podrían convertirse en fármacos dirigidos a SAMHD1 en el entorno clínico, con el objetivo de mejorar la eficacia de estas terapias. También es fundamental comprender el alcance total de la interacción de estos compuestos basados en nucleósidos con SAMHD1, una cuestión que también puede abordarse utilizando este ensayo acoplado a enzimas. En conjunto, el ensayo de actividad SAMHD1 acoplado a enzimas, como se informa aquí, es un ensayo de bajo costo, versátil y de alto rendimiento que se puede utilizar para ampliar nuestra comprensión de esta importante enzima.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Agradecemos a Thomas Lundbäck y a los miembros del laboratorio de Thomas Helliday por su asesoramiento y apoyo. Parte de este trabajo fue facilitado por el Centro de Ciencias de las Proteínas del Instituto Karolinska/SciLifeLab (http://ki.se/psf), y agradecemos al Instituto Nacional del Cáncer (NCI), a la División de Tratamiento y Diagnóstico del Cáncer (DCTD) y al Programa de Terapias del Desarrollo (DTP) (http://dtp.cancer.gov) por proporcionar un compuesto. La financiación provino de subvenciones otorgadas a S.G.R. por el Consejo Sueco de Investigación (2018-02114), la Sociedad Sueca del Cáncer (19-0056-JIA, 20-0879-PJ), el Fondo Sueco de Cáncer Infantil (PR2019-0014) y el Instituto Karolinska.
Materiales
| Name | Company | Catalog Number | Comments |
| 2'-deoxyadenosine-5'-triphosphate (dATP) | Jena bioscience | NU-1001 | Compound tested in SAMHD1 activator/substrate assay |
| 2'-deoxycytidine-5'-triphosphate (dCTP) | Jena bioscience | NU-1002 | Compound tested in SAMHD1 activator/substrate assay |
| 2'-Deoxyguanosine-5'-(α-thio)-triphosphate (dGTPαS) | Jena bioscience | NU-424 | Non-hydrolyzable dGTP analogue for SAMHD1 activator/substrate assay |
| 2'-deoxyguanosine-5'-triphosphate (dGTP) | GE Healthcare | 27-1870-04 | SAMHD1 allosteric activator and substrate used in inhibition assay and activator/substrate assay |
| 2'-Deoxythymidine-5'-[(α,β)-imido]triphosphate (dTMPNPP) | Jena bioscience | NU-907-1 | Compound tested in SAMHD1 inhibition assay |
| 2'-deoxythymidine-5'-triphosphate (dTTP) | Jena bioscience | NU-1004 | Compound tested in SAMHD1 activator/substrate assay |
| 2'Deoxyguanosine mohohydrate (dGuo) | Sigma-Aldrich | D0901 | Compound tested in SAMHD1 inhibition assay |
| 384 well clear flat-bottom microplate | Thermo Fisher Scientific | 262160 | Assay plate |
| 96 well clear U-bottom polypropylene microplate | Thermo Fisher Scientific | 267245 | Compound dilution plate |
| Ammonium heptamolybdate tetrahydrate | Sigma-Aldrich | A1343 | Reagent required for malachite green working reagent |
| ara-Cytidine-5'-triphosphate (ara-CTP) | Jena bioscience | NU-1170 | Compound tested in SAMHD1 activator/substrate assay |
| Clofarabine-5'-triphosphate (Cl-F-ara-ATP) | Jena bioscience | NU-874 | Compound tested in SAMHD1 activator/substrate assay |
| Dimethyl sulphoxide (DMSO) | VWR | 23486.297 | Solvent |
| Ethylenediaminetetraacetic acid disodium salt dihydrate (EDTA) | Sigma-Aldrich | E5134 | EDTA stop solution component |
| Gemcitabine-5'-triphosphate (dF-dCTP) | Jena bioscience | NU-1607 | Compound tested in SAMHD1 activator/substrate assay |
| GraphPad Prism | GraphPad Software | Prism 8 | Data analysis and visualisation |
| Guanosine 5′-triphosphate (GTP) sodium salt hydrate | Sigma-Aldrich | G8877 | Allosteric activator for SAMHD1 activator/substrate assay |
| His-tagged E. coli inorganic pyrophosphatase (PPase) | Generated in house using Protein Science Facility, Karolinska Institutet | - | Recombinant PPase protein, hydrolises inorganic triphosphate and pyrophosphate to orthophosphate so it can form complex with malachite green |
| His-tagged human SAMHD1 | Generated in house using Protein Science Facility, Karolinska Institutet | - | Recombinant SAMHD1 protein, hydrolizes dNTPs into its corresponding nucleoside and inorganic triphosphate |
| Hydroxyurea | Sigma-Aldrich | H8627 | Compound tested in SAMHD1 inhibition assay |
| Lomofungin | National Cancer Institute (NCI)/Division of Cancer Treatment and Diagnosis (DCTD)/Developmental Therapeutics Program (DTP) | NSC106995 | Compound tested in SAMHD1 inhibition assay |
| Magnesium Chloride hexahydrate (MgCl2) | Sigma-Aldrich | M2670 | SAMHD1 reaction buffer component |
| Malachite green Carbinol hydrochloride | Sigma-Aldrich | 213020 | Malachite green stock component |
| Microplate Reader | Hidex | Hidex Sense Microplate reader | Data acquisition, absorption read at 630 nm wavelength |
| Sodium Chloride (NaCl) | Sigma-Aldrich | 31434 | SAMHD1 reaction buffer component |
| Sodium hydroxide (NaOH) | Sigma-Aldrich | 567530 | SAMHD1 reaction buffer component |
| Sodium phosphate (Na3PO4) | Sigma-Aldrich | 342483 | Required for phosphate standard curve |
| Sulphuric acid 95-97% | Sigma-Aldrich | 84720 | Malachite green stock component |
| Tris-Acetate salt | Sigma-Aldrich | T1258 | SAMHD1 reaction buffer component |
| Tris(2-carboxyethyl)phosphine hydrochloride (TCEP) | Sigma-Aldrich | C4706 | SAMHD1 reaction buffer component |
| Tween-20 | Sigma-Aldrich | P1379 | SAMHD1 reaction buffer and malachite green working reagent component |
Referencias
- Franzolin, E., et al. The deoxynucleotide triphosphohydrolase SAMHD1 is a major regulator of DNA precursor pools in mammalian cells. Proceedings of the National Academy of Sciences of the United States of America. 110 (35), 14272-14277 (2013).
- Coggins, S. A., Mahboubi, B., Schinazi, R. F., Kim, B. SAMHD1 functions and human diseases. Viruses. 12 (4), 382(2020).
- Goldstone, D. C., et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature. 480 (7377), 379-382 (2011).
- Powell, R. D., Holland, P. J., Hollis, T., Perrino, F. W. Aicardi-Goutieres syndrome gene and HIV-1 restriction factor SAMHD1 is a dGTP-regulated deoxynucleotide triphosphohydrolase. The Journal of Biological Chemistry. 286 (51), 43596-43600 (2011).
- Morris, E. R., Taylor, I. A. The missing link: Allostery and catalysis in the anti-viral protein SAMHD1. Biochemical Society Transactions. 47 (4), 1013-1027 (2019).
- Arnold, L. H., Kunzelmann, S., Webb, M. R., Taylor, I. A. A continuous enzyme-coupled assay for triphosphohydrolase activity of HIV-1 restriction factor SAMHD1. Antimicrobial Agents and Chemotherapy. 59 (1), 186-192 (2015).
- Herold, N., et al. Targeting SAMHD1 with the Vpx protein to improve cytarabine therapy for hematological malignancies. Nature Medicine. 23 (2), 256-263 (2017).
- Schneider, C., et al. SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia. Nature Medicine. 23 (2), 250-255 (2017).
- Hollenbaugh, J. A., et al. Substrates and inhibitors of SAMHD1. PloS One. 12 (1), 0169052(2017).
- Knecht, K. M., et al. The structural basis for cancer drug interactions with the catalytic and allosteric sites of SAMHD1. Proceedings of the National Academy of Sciences of the United States of America. 115 (43), 10022-10031 (2018).
- Oellerich, T., et al. Selective inactivation of hypomethylating agents by SAMHD1 provides a rationale for therapeutic stratification in AML. Nature Communications. 10 (1), 3475(2019).
- Herold, N., et al. SAMHD1 protects cancer cells from various nucleoside-based antimetabolites. Cell Cycle. 16 (11), 1029-1038 (2017).
- Rothenburger, T., et al. SAMHD1 is a key regulator of the lineage-specific response of acute lymphoblastic leukaemias to nelarabine. Communications Biology. 3 (1), 324(2020).
- Ordonez, P., et al. SAMHD1 enhances nucleoside- analogue efficacy against HIV-1 in myeloid cells. Scientific Reports. 7, 42824(2017).
- Castellví, M., et al. Pharmacological modulation of SAMHD1 activity by CDK4/6 inhibitors improves anticancer therapy. Cancers. 12 (3), 713-719 (2020).
- Rassidakis, G. Z., et al. Low-level expression of SAMHD1 in acute myeloid leukemia (AML) blasts correlates with improved outcome upon consolidation chemotherapy with high-dose cytarabine-based regimens. Blood Cancer Journal. 8 (11), 98(2018).
- Rudd, S. G., Schaller, T., Herold, N. SAMHD1 is a barrier to antimetabolite-based cancer therapies. Molecular & Cellular Oncology. 4 (2), 1287554(2017).
- Rudd, S. G., et al. Ribonucleotide reductase inhibitors suppress SAMHD1 ara-CTPase activity enhancing cytarabine efficacy. EMBO Molecular Medicine. 41, 10419(2020).
- Seamon, K. J., et al. Small molecule inhibition of SAMHD1 dNTPase by tetramer destabilization. Journal of the American Chemical Society. 136 (28), 9822-9825 (2014).
- Seamon, K. J., Stivers, J. T. A high-throughput enzyme-coupled assay for SAMHD1 dNTPase. Journal of Biomolecular Screening. 20 (6), 801-809 (2015).
- Mauney, C. H., Perrino, F. W., Hollis, T. Identification of inhibitors of the dNTP triphosphohydrolase SAMHD1 using a novel and direct high-throughput assay. Biochemistry. 57 (47), 6624-6636 (2018).
- Morris, E. R., et al. Crystal structures of SAMHD1 inhibitor complexes reveal the mechanism of water-mediated dNTP hydrolysis. Nature Communications. 11 (1), 3165(2020).
- Hansen, E. C., Seamon, K. J., Cravens, S. L., Stivers, J. T. GTP activator and dNTP substrates of HIV-1 restriction factor SAMHD1 generate a long-lived activated state. Proceedings of the National Academy of Sciences of the United States of America. 111 (18), 1843-1851 (2014).
- Baykov, A. A., Evtushenko, O. A., Avaeva, S. M. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Analytical Biochemistry. 171 (2), 266-270 (1988).
- Hyun, M., Bohr, V. A., Ahn, B. Biochemical characterization of the WRN-1 RecQ helicase of Caenorhabditis elegans. Biochemistry. 47 (28), 7583-7593 (2008).
- Lin, H. -H., Huang, C. -Y. Characterization of flavonol inhibition of DnaB helicase: real-time monitoring, structural modeling, and proposed mechanism. Journal of Biomedicine & Biotechnology. 2012 (4), 735368(2012).
- Yang, M., Wang, G. ATPase activity measurement of DNA replicative helicase from Bacillus stearothermophilus by malachite green method. Analytical Biochemistry. 509, 46-49 (2016).
- Allard, B., Cousineau, I., Spring, K., Stagg, J. Measurement of CD73 enzymatic activity using luminescence-based and colorimetric assays. Methods in Enzymology. 629, 269-289 (2019).
- Lee, M., et al. Structure-activity relationship of sulfonyl piperazine LpxH inhibitors analyzed by an LpxE-coupled malachite green assay. ACS Infectious Diseases. 5 (4), 641-651 (2019).
- Carreras-Puigvert, J., et al. A comprehensive structural, biochemical and biological profiling of the human NUDIX hydrolase family. Nature Communications. 8 (1), 1541(2017).
- Valerie, N. C. K., et al. NUDT15 hydrolyzes 6-thio-deoxyGTP to mediate the anticancer efficacy of 6-thioguanine. Cancer Research. 76 (18), 5501-5511 (2016).
- Carter, M., et al. Human NUDT22 Is a UDP-glucose/galactose hydrolase exhibiting a unique structural fold. Structure. 26 (2), 295-303 (2018).
- Gad, H., et al. MTH1 inhibition eradicates cancer by preventing sanitation of the dNTP pool. Nature. 508 (7495), 215-221 (2014).
- Page, B. D. G., et al. Targeted NUDT5 inhibitors block hormone signaling in breast cancer cells. Nature Communications. 9 (1), 250(2018).
- Zhang, S. M., et al. Development of a chemical probe against NUDT15. Nature Chemical Biology. 16 (10), 1120-1128 (2020).
- Michel, M., et al. In silico druggability assessment of the NUDIX hydrolase protein family as a workflow for target prioritization. Frontiers in Chemistry. 8, 443(2020).
- Zhang, J., Chung, T., Oldenburg, K. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. Journal of Biomolecular Screening. 4 (2), 67(1999).
- Baykov, A. A., Evtushenko, O. A., Avaeva, S. M. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Analytical Biochemistry. 171 (2), 266-270 (1988).
- Markossian, S., et al. Assay guidance manual. Eli Lilly & Company and the National Center for Advancing Translational Sciences. , Bethesda (MD). (2004).
- Holdgate, G. A., Meek, T. D., Grimley, R. L. Mechanistic enzymology in drug discovery: a fresh perspective. Nature Reviews Drug Discovery. 17 (2), 115-132 (2018).
- Tsesmetzis, N., Paulin, C. B. J., Rudd, S. G., Herold, N. Nucleobase and nucleoside analogues: resistance and re-sensitisation at the level of pharmacokinetics, pharmacodynamics and metabolism. Cancers. 10 (7), 240(2018).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoExplorar más artículos
This article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados