
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
הערכת Vivo לשעבר של התכווצות, ועיפנות Alternans בשרירי שלד מבודדים
In This Article
Summary
אנו מתארים שיטה ישירה למדידת כוח שרירים, כוח שרירים, התכווצות וקינטיקה עיפנות של שרירי שלד מבודדים ב במבחנה מערכת באמצעות גירוי תחום. מידע רב ערך על Ca 2 + מאפייני טיפול ומכונה התכווצות של השריר ניתן להשיג באמצעות פרוטוקולים שונים המגרים את הדמיון.
Abstract
מתואר כאן היא שיטה למדידת התכווצות של שרירי שלד מבודדים. פרמטרים כגון כוח שריר, כוח השריר, קינטיקה התכווצות, עיפנות, והתאוששות לאחר עייפות ניתן להשיג להעריך היבטים ספציפיים של צימוד עירור הכיווץ (ECC) תהליך כגון רגישות, מכונה התכווצות וCa 2 + יכולת טיפול. שיטה זו מסירה את אספקת דם והעצבים ומתמקדת בשרירי השלד המבודדים עצמו. אנו משתמשים באופן שגרתי בשיטה זו כדי לזהות מרכיבים גנטיים המשנים את נכס ההתכווצות של שריר השלד למרות ויסות Ca 2 + מסלולי איתות. כאן, אנו מתארים פנוטיפ זיהה לאחרונה שלד שריר, כלומר, alternans מכונאי, כדוגמה למידע המגוון ועשיר שניתן להשיג באמצעות בדיקה בהתכווצות שרירי מבחנה. שילוב של assay זה עם מבחנים בודדים סלולריים, גישות גנטיות וbiochemiמבחני stry יכולים לספק תובנה חשובות למנגנונים של ECC בשרירי שלד.
Introduction
שרירי שלד לצרף לעצמות השלד וליצור כוחות התכווצות תחת השליטה של מערכת העצבים המרכזית. צימוד עירור-התכווצות (ECC) מתייחס לתהליך של המרת גירוי חשמלי לתגובה מכאנית. Ca 2 + איתות היא מרכיב חיוני בתפקוד כויץ בשרירי שלד. יעיל Ca 2 + גיוס מreticulum sarcoplasmic (SR) הוא מרכיב חשוב לECC בתאי שריר 1, 2, ושינויים בCa 2 + איתות התאי בבסיס תפקוד כויץ המקביל במספר מחל שרירים 3-5. הערכה נכונה של התכווצות שרירים היא חיונית ומחמיא לCa 2 + הדמיה ומבחנים אחרים כדי להשיג תובנה לתפקוד שרירי שלד, ולא רק ברמת כויץ, אלא גם ברמה קינטית. כוח ומהירות גם ניתן להשיג על מנת ליידע את התכונה החשובה שלכוח שרירים ומצב תהליך ECC בתנאים פיסיולוגיים וpathophysiological שונים.
תחום זה של מחקר פורה יש היסטוריה עשירה מאוד ותאוריות רבות של התכווצות שרירים, הופיעו מעל אלפיים שנה 6. מחקר השרירים מודרני כנראה מתחיל ב1674-1682 באבחנה המיקרוסקופית של-קווקווים צולבים וmyofibrils בסיבי שריר על ידי 6 יוונהוק. כמעט מאה שנה מאוחר יותר, לואיג'י גלוואני הבחין כי תוקף חוזי שרירי צפרדע כשהעצב שלה הוא נגע עם אזמל במהלך פריקת ניצוץ ממכונה חשמלית רחוקה 7-9. התכווצות יכולה גם להיות מיוצרת על ידי חיבור העצב לשריר הרגל דרך מוליך מתכתי. את הפרטים של מנגנון האיתות החשמלי המורכב הטיפו גלווני סופו של דבר גובשו על ידי הודג'קין, הקסלי וכץ במשוואה המפורסמת שלהם 10, 11, שהפכה לבסיס של אלקטרו. התצפיות המדהימות של ריןגר על ההשפעות של תאי Ca 2 + על ההתכווצות של לב צפרדע ושרירי שלד 12-15 מייצג את הצעד הראשון החשוב בהכרה של Ca 2 + כרגולטור מפתח של התכווצות שרירים 16, 17. משינה 1980 ועד ימינו פרץ של תגליות בתחום התכווצות השרירים מומש עקב כניסתה של התכווצות שרירים ופרוטוקולי עיפנות בשרירי שלד Murine 18. ג'ונס ואדוארדס היו הראשון שטענה כי עייפות לסירוגין בתדירות נמוכה (הפחתת פעילות גופנית הנגרמת בכוח) 19 היו קשורים לשינויים במכונות ECC ולא מנגנון ההתכווצות. בשנת 1980 מאוחר ו1990 המוקדמים, Kolkeck et 20 אל, Kolbeck וNosek 21, ו 22 ריד היו באמצעות שריר סרעפת ממכרסמים כדי לחקור את ההשפעות של theophyllines, cortiosterone, ורדיקלים חופשיים בהתכווצות שרירי שלד, ואילו ברוקס וFaulknאה היו ראשון לדווח על מדידות של כוח חזר ומדידות כוח ובפה מהר שרירים איטיים מעכברים 22. בנוסף, Lannegren, Westerblad, כבש, וWesterblad היו הראשון ישירות לקשר contractility vivo לשעבר עם Ca 2 + תאי רגולציה והתחיל לחקור את תפקידו של חמצת בעייפות שרירים 23, 24.
המעבדות שלנו תרמו באופן משמעותי מאז 2000 מוקדמים לקראת הבנה של גני רומן עם עצב ותפקידים הרגולטוריים על השריר ECC עם תפקידים קריטיים בהתכווצות שרירים, עיפנות, והזדקנות באמצעות שילוב של מחקרי contractility שרירי עכבר שלמים, תאיים Ca 2 + ניטור ב סיבי שריר שלם ועור ומניפולציות גנטיות מולקולרית 3-5, 25-29.
כאן אנו מפורטים פרוטוקול הניסוי, למדידת התכווצות של soleus המבודד Murine וlongus digitorum פושטים (EDL) שרירים, אשר תואמים את רוב איטי חמצונים (אני סוג וסיבי שריר IIa) ובעיקר שרירים מהירים glyocolytic (הסוג IIb וסיבי שריר IIx) עם תכונות התכווצות מובחנות. בפרוטוקול זה, קומפלקסים שלמים שריר גיד היינו מבודדים ורחצו במערכת ADI PowerLab Radnotti קאמרית מסופקת גם עם חמצן טהור או בתערובת של חמצן (95%) ו-CO 2 (5%). כוחות התכווצות נוצרו על ידי גירויים חשמליים מממריץ גראס וזוהה באמצעות מתמר כוח שהיה משולב עם מערכת PowerLab/400 ADI, המאפשר התאמה אישית של שגרת מאקרו לשלוט רכישה, האיסוף, דיגיטציה, והאחסון של נתונים. התקנה זו יכולה למדוד כוח שרירים, כוח שרירים, כמו גם את יחסי כוח מול תדר, עייפות שרירים, התאוששות מעייפות שרירים, מהירות ותכונות הקינטית כוללות של התכווצות שרירים. בנוסף, השפעות של תרופות על התכווצות שרירים ניתן לנטר באמצעות ניסויים אלה.
יתרונות של שיטה זו טמונות בהסרת רכיבים העצביים וכלי דם משריר השלד, המאפשרים הערכה ישירה של המאפיינים הפנימיים של הידבקות שרירים. בנוסף, מבחני contractility vivo לשעבר לאפשר מניפולציה של הסביבה התאית המקיפה את השרירים המבודדים, המאפשרת שימוש במניפולציות פרמקולוגיים של ערוצים שונים יון חלחול ומובילים במטרה להגדיר את תפקידיהם הפיסיולוגיים לתפקוד שרירי שלד.
מערכת vivo לשעבר זה אפשרה לנו לאחרונה מגלה התנהגות מובהקת alternan בהכנות שרירי מוטציה מסוימות, שהיו קשורים לCa שינה התאי 2 + טיפול נכסים 4. Alternans מוגדרים כפרקי פרץ תנודות של כוח התכווצות בשלב הירידה של הפרופיל עייף אותו. במהלך אירועים אלה כוחות התכווצות רגע להגדיל מעל רמתו הקודמת בכוח דuring גירוי מתיש, אולי כי גם יותר Ca 2 + מתפרסם או מכונות כויץ הפכו רגישות יותר לCa 2 + 30. טיפול בחומצת cyclopiazonic (רו"ח), חוסם הפיך של סידן reticulum ATPase sarcoplasmic-endoplasmic (SERCA), קפאין, אגוניסט של ryanodine ערוץ (RyR) וחזרתי לעייף גירויים יכול לגרום לכל alternans המכאני 4, מה שמראה כי alternans קשור ישירות אפנון של תהליך צימוד EC. הדגמה של השיטה כדי לגרום ולהקליט alternans מכונאי בהתקנת contractility מבחנה משמשת כדוגמה כדי להראות את הפרמטרים הניסיוניים המגוונים שיכול להיות שהושגו עם מערכת זו או דומים לאלה, המבוסס על תחומי מחקר בודדים.
שיטה זו עשויה להיות עניין לחוקרים הלומדים פיזיולוגיה של שרירים. התקנה דומה יכולה לשמש גם למתחמי muscle-tendon/ligament שלד מבודדים משנימקומות אנטומיים, כמו גם לסיבים בודדים ורצועות שרירים.
Protocol
הרכב פתרון:
2.5 מ"מ Ca 2 + פתרון Tyrode: 140 המ"מ NaCl, 5 המ"מ KCl, 10 HEPES מ"מ, 2.5 מ"מ CaCl 2, 2 מ"מ MgCl 2 וגלוקוז 10 מ"מימ
0 מ"מ Ca 2 + פתרון Tyrode: 140 המ"מ NaCl, 5 המ"מ KCl, 10 HEPES מ"מ, 2 המ"מ MgCl 2, חומצת אתילן גליקול 0.1 מ"מ tetraacetic (EGTA) וגלוקוז 10 מ"מימ
הערה: פתרון רחצה צריך להיות רווי עם 100% O 2 אם משתמש בפתרון לעיל, אך עם 95% O 2 עם 5% CO 2, אם באמצעות חוצצי יקרבונט מבוססים כדי לשמור על pH קבוע. 2.5 מ"מ Ca 2 + מתווסף למאגר הים כדי לסכם את הרמה של Ca 2 + מצא בחלל החוץ התאי וגלוקוז 10 מ"מימ הוא חשוב שכן המיטוכונדריה עדיין מתפקדות בשרירים האלה ברציפות כדי לייצר ATP בנוכחות של גלוקוז.
1. הגדרה למעלה ניסוי contractility Vivo Ex שימושמערכת ADI PowerLab
- ציור סכמטי של מערכת contractility ערוץ 4-vivo לשעבר מוצג באיור 1. מחשב שולט ממריץ לייצר פולסים כיכר גל, שמסוננים על ידי יחידת הבידוד להגיע זוג חוטי פלטינה סביב כל מתחם מבודד שריר גיד. זה נקרא גם גירוי תחום. התכווצות של השריר להגיב לגירוי שחשה ידי מתמר כוח, אות שנוצרה ממנו היה מוגברת, מסוננות והועברה בחזרה למחשב על ידי / D ממיר (מיזוג אות). אז אות דיגיטציה וניתן לשמור לניתוחים מאוחרים יותר.
- כדי להתכונן לניסוי vivo לשעבר התכווצות, הפנייה הראשונה בתחום הממריץ, A / D הממיר (במקרה זה עדי חשמל מעבדה) או תוכנה דומה (למשל, LABVIEW), ואחריו את המחשב.
- פתח Chart4 תוכנה (או גרסת תוכנה אחרת שתואמים למערכת) וinitiatEA משימת 4-ערוץ; תקשורת התקינה של התוכנה וחומרה מצוינת באמצעות מתחיל תצורה של החומרה, ועל הכפתור המתאים בתוכנה הופך מבצעי (חי); הפונקציה 4-הערוץ מאפשרת מדידה בו זמנית של זוג מהשרירים סוג בר, ועכברים שעברו מוטציה, או 2 edl ו2 שרירים soleus מאותה העכבר, וניתן להשתמש בו לשרירים אחרים כגון סרעפת (סרעפת שלמה, חמה, סרעפת, שריר סרעפת או רצועות) וtibialis הקדמי; זה יכול לשמש גם לשריר לב, והכנות לצרורות של שרירים, במיוחד אם אחד משתמש בשרירי חולדה. במקרה זה הגודל של השרירים עלול להגביל דיפוזיה חמצן כך צרורות שריר ובכך בחירה טובה יותר. עם זאת, טכניקות נתיחת מומחים נדרשות להכנות צרור שרירים.
- כיול של מתמר הכח: זו היא להבטיח את ההשוואה של בסיס נתונים שנוצר בערוצים שונים ובזמן אחר. התחלת ההקלטה ראשונה, sequentially לתלות 1-g, 2-g ומשקולות 5-g במטע המדגם של מתמר הכח, לעצור את ההקלטה, לחשב שינויים מקבילים בmV שמוצגים בכל ערוץ של Chart4 תוכנה, עלילת ΔmV ומשקל כדי לבדוק וליניאריות לקבוע את גורם ההמרה בין mV וגרם של כוח. אנו ממליצים לבצע כיולים אלה או בתחילתו או בסופו של כל ניסוי כדי להבטיח כי כיול ספציפי מתקבל עבור כל ניסוי ספציפי.
- בדקו את החיבור נכון של צינור הטפלון, לנקז כל נוכחי נוזל בצינור אמבטית הרקמות, לשטוף את תאי שלוש פעמים או יותר עם DDH 2 O ולמלא עם 20 ~ 25 מ"ל 2.5 מ"מ Ca 2 + פתרון Tyrode. Ca 2 + וגלוקוז מסופקים בפתרון הרחצה כדי לעזור לשמור על שלמות קרום, אופטימלית Ca 2 + טעינה של SR, ומקור זמין של אנרגיה לייצור של ה-ATP על ידי השריר עצמו, שכן המיטוכונדריה להישאר פונקציונלית מלא בתכשירים אלה.
- בדקו את החיבור נכון של אספקת החמצן, לפתוח את מכל החמצן, ולהתאים את זרימת חמצן כדי לספק מבעבע diffusive והומוגנית של קאמרי. רמות חמצן בקלות ניתן לקבוע כפונקציה של כוח לעומת זמן התכווצות. מד זרימה יכול גם להיות מותקן, במיוחד אם חוקרים מתעניייינים בחקר ההשפעות של היפוקסיה או hyperoxia. אם שרירים להיות היפוקסי, הכח באופן טבעי יורד. בעוד hyperoxia יכול גם להזיק שרירים אך השפעותיו עשויות להיות קשות יותר כדי להתגלות, כי רוב הניסויים מבוצעים בתנאי hyperoxic מלאכותיים כדי לפצות על היעדר אספקת דם תקינה לשרירים. ברוב המערכות ניתן בועת התאים השונים עם אותה הכמות של חמצן פשוט על ידי שליטה על זרימת חמצן לכל תא קבוע.
- התאם את הרגישות של כל הערוצים על מנת להבטיח את הרזולוציה המקסימלי הכח בלי להרוות את הערוץ. רגישות ערוץ יכול להשתנות במידה ניכרת כלתפקדגיל ד של בעל חיים, טמפרטורה ניסיונית, מניפולציות גנטיות, גודל שריר, נקעי עכבר, ואת היכולת של הנסיינית לנתח שרירים ללא ניזקים כמו שצריך. מניסיוננו, כאשר מודדים את כוח התכווצות soleus, הרגישות משתנית 0.5-10 mV / סנטימטר, ואילו במקרה של אדי, הרגישות יכולה להשתנות 1-20 mV / סנטימטר.
- פרוטוקול לגירוי לעייף צריך להיות הוקם. בChart4 תוכנת מאקרו ניתן להשתמש. כדי לתכנת מאקרו חדש: בChart4 התוכנה, לחץ להתחיל במדידה, ולאחר מכן לחץ על מאקרו, פיקוד מאקרו, להתחיל להקליט, מתחיל לחזור, לבחור תדירות גירוי רצוי, ולאחר מכן חזור על סיום. לפרוטוקול איזון, גירוי חוזר על עצמו בכל דקה למשך 30 דקות ומייגעת לפרוטוקול, גירוי חוזר על עצמו כל 2 שניות למשך 5 דקות. שינויים אלו במחזוריות של הגירוי, משפיעים ישירות על המחזור, שהוא גם פונקציה של משך הגירוי. פרמטרים אלו יכולים להיות מגוונים כדי לבחון אספה שונהCTS של התכווצות ועיפנות.
2. הכנת חבילות שרירי אינטקט
- נתיחת אדי: העכבר הקריב בעקבות הנחיות NIH ופרוטוקולי בעלי חיים מוסדיים IACUC. העכבר הקריב על ידי נקע בצוואר רחם. אז מאוס מסודר במיקום הצדדי. אדי שריר הוא שריר מהיר glycolytic עם צבע ורוד בהיר, לבן. אורכה כ 10-13 מ"מ ומשקל 8-11 מ"ג בסוג פראי C57BL / 6 עכבר. אדי יש את הפונקציות של הארכת בוהן 2-5, וdorso כיפוף-הרגל בקרסול. הוא מעוצבבים על ידי עצב peroneal. כדי לנתח את האד, לבצע חתך בעור שטחי וחתך את fascia בין tibialis הקדמיים וקבוצת השרירים האחורית, ולמקם את המקור (פרוקסימלי) שבו רצועה מחוברת לcondyle הרוחבי של שוקה ו3 / 4 מעולות של קדמי פני השטח של שוקית (מרווח interosseous). חותך את הרצועות עם מספריים לרפואת עיניים עדינים כdistally ככל האפשר מהשרירים; זה procedurדואר משחרר מוצא הפרוקסימלי של שריר האד. החזק את הרצועה עם מלקחיים בוטים ולמשוך לאט כדי לשחרר את האד. ייתכן שיהיה צורך לחתוך חלק מperimysium מקיף את שרירי edl ואחרים ברחבי אדי, צעד זה הוא קריטי שכן ניזק יכול להתרחש לשריר האד במהלך השלב העדין הזה. בשלב הבא, לעבור לאזור ההכנסה במקום הארבעה גידי דיסטלי להכניס את הפלנגות האמצעיות והדיסטלי של 2-5 ספרות. חותך את הגידים ככל האפשר מהשרירים.
- העברת שריר אדי המבודד לתוך צלחת נתיחה המכילה פתרון Tyrode איזוטוני. כמה מודלים של בעלי חיים מוטנטים, מהונדסים, ויש נוקאאוט שרירים מאוד שבירים וניצול של Ca 2 +-פתרון Tyrode חופשי הוא הכרחי כדי למנוע ניזק לשריר Ca 2 +-מושרה לפני מדידות contractility. לאחר מכן, השתמש קשר כירורגית לקשור בחוזקה בשני הקצוות של שריר האד כdistally משרירים ככל האפשר. מידה טובה היא לקשור מעט מעל אמצע עמoint אורך של הרצועה או הגיד. השתמש תפר גודל 6-0 להליך זה, ולהעביר את שריר האד לחדר אמבטית רקמות 2-הרווי O, לעגן את השריר בגרוב המדגם של מתמר הכח ווו נייר בתחתית תא האמבטיה, חזור תהליך לרגל השנייה. אנחנו גם מתחילים לנצל את שיטה חדשה שמחזיקה את השרירים עם מהדק במקום תפרים. אם המטרה העיקרית היא ללמוד תכונות קינטיים ו / או להשיג כוח שרירים, מומלץ שתפר יהיה קצר ככל האפשר, או התפר יוחלף במוט מתכת.
- נתיחת Soleus: soleus הוא שריר איטי חמצונים בעיקר בצבע אדום עשיר. זה ~ 1 מ"מ קצר יותר מאשר האד אבל שוקל מעט יותר מהאד. הוא מעוצבבים על ידי העצב הטיביאלי והיא מבצעת את הפעולה של כיפוף הפלנטרי של כף הרגל. כדי לנתח את soleus, לגשת לצד לרוחב האחורי של הרגל, לדחוף הצידה את שריר הגסטרוקנמיוס שבדרך כלל מכסה soleus, ולזהות את השריר עם צבע אדום כהה. במקור (פרוקסימלי), לחתוך את רצועות חיבור למחצית הפרוקסימלי של עצם שוק אחורי לאורך קו soleal והפרוקסימלי 1/3 מהשוקית האחורית; הבא, בהכנסה (דיסטלי), לחתוך את גיד calcaneal שמחדיר לתוך אחורי calcaneus. זהירות חופשיה את soleus ולעגן את שריר soleus כראוי בחדר האמבטיה כלאד.
3. מדידת התכווצות של שרירי השלד המבודדים
- ברגע שהשרירים הם רכובים בתאי רקמות אמבטיה נפרדים, להתחיל בהקלטה. מערכות דומות ביותר מאפשרות איפוס של נקודת ההתחלה של הקלטת כוח. פונקציה זו מזוהית בדרך כלל עם המגבר, במקרה הספציפי של מערכת PowerLab, כפונקציה של מגבר הגשר. הוא מאפשר התבוננות בשינויים בסיסיים ומספק דרך נוחה לכל התכווצויות השרירים להיות מנותח מאפס. שרירים מבודדים אז מומרצים עם פולסים ריבוע גלשיכול לנוע באופן נרחב כפונקציה של גודל התא, עובי חוטי פלטינה, מרחק בין החוטים, ואפילו בהרכב הפתרון הניסיוני. יש לנו עובדים נוכחיים של 60 אמפר (שכבר ציינו לעיל, כי זרמי mA 350 נראים להזיק להכנות שריר) ורכבות גירוי של 350, 500, 1000 וMS, בהתאם למטרותיו של פרוטוקול ספציפי. קטניות הגל המרובעות הבודדות צריכות משך נעות 0.3-1 ms.
- הצעד הבא הוא לבחור תדר הגירויים מסוגלים לייצר גירוי tetanic התמזג (לשעבר: ~ 100 הרץ כדי לאפשר ייצור של כוח מקסימאלי בשריר ואד ~ 60 רץ בשריר soleus) ואילו באיטיות ומותח את השרירים בזהירות כדי לזהות את אורך אופטימלי של שרירים אלה. כאשר שהרירים נמתחים בזהירות, להמתין 30 שניות ולעורר ב100 הרץ; לחכות 30 שניות ולמתוח שוב, ולאחר מכן לחזור על הגירוי, עד לנקודה שבי הכח לא להגדיל יותר.
- שרירים אלה יש undergone שינויים משמעותיים בסביבה. מומלץ לאפשר לשרירים להסתגל לסביבה החדשה, צעד בפרוטוקולים שלנו מכונה "איזון". לאזן את השרירים האלה באותה התדירות של גירוי המשמש בשלב המתיחה, 100 הרץ ל20-30 דקות, עד שלפחות 5 התכווצויות tetanic רצופות הן לגמרי יציבות (לא צמצום, לא גדל, בסיס יציב). מחזוריות של רכבות הגירוי (100 הרץ, 500 מילי משך, 1 MS פרט הדופק) במהלך האיזון היא דקות 1, אשר שווה למחזור של 1.66%, גירוי שאינו מייגע. בהקשר זה, מחזור פעולה של 1.66% המשמעות הוא שעל סך של 100%, שרירים עובדים 1.66% מהזמן, על ידי הגדלת שרירי מחזור סופו של דבר יכולים להיגרם לעייפות. אם אחרת שרירים של עכבר סוג בריא, פראי להראות פרופיל מתיש בתקופת האיזון הזה, ייתכן שהם נפגעו במהלך נתיחה, היפוקסיה מתרחשת בתאים / שרירים, או ele המוגזםctrolysis דרך אלקטרודות הגירוי הוא יצירת רדיקלים חופשיים. Westerblad הציע בעבר כי זרמים גדולים מ 400 אמפר לגרום היווצרות של רדיקלים חופשיים 23. ברור, הפלטינה באיכות הגבוהה ביותר שמוצעת, מאז מתכות אחרות בהחלט להוביל להיווצרות של רדיקלים חופשיים ורעילות שריר.
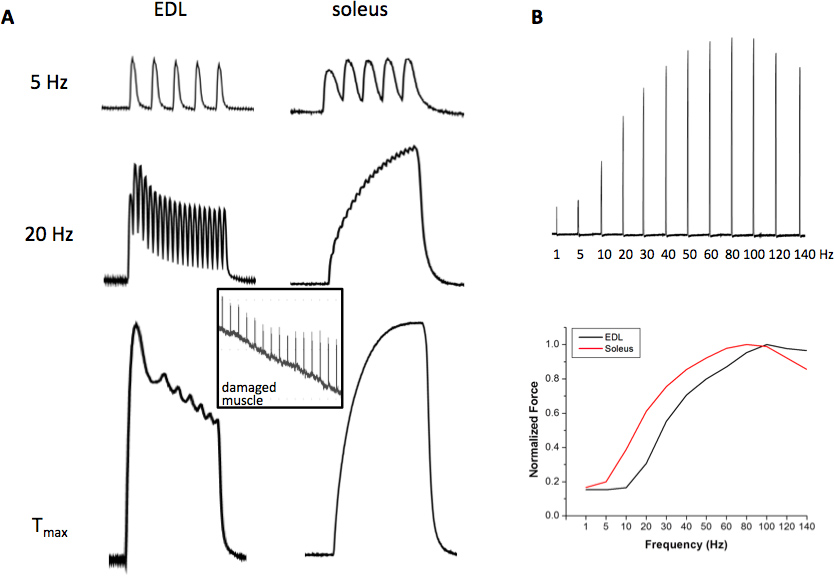
- השג את יחסי הכח מול תדירות (FF) על ידי גירוי השרירים עם תדרי הגירוי הבאים: 1-140 הרץ (במרווחים של 5-10 הרץ), במחזוריות של 30-60 שניות בעת ביצוע ניסויים ב 25 ° C. בעת ביצוע ניסויים על 37 מעלות צלזיוס, להאריך את FF לתדרים גבוהים יותר של עד 300 רץ לשריר סרעפת, 180-200 הרץ לsoleus ו220-250 הרץ לאד. בשלב בא, לזהות את התדר של גירוי שיוצר כוח מקסימאלי tetanic (ט מקסימום) וכ ½ כוח tetanic מקסימאלי (1/2 T מקס), לפעמים זה קשה להשיג את מקסימום T ½ המדויק עבור שניהם edl ובלעדישרירינו אם רק מקור אחד ממריץ נמצא בשימוש, בגלל ההבדלים המהותיים בין השרירים הללו. פתרון בר קיימא הוא לזהות תדר שמייצר 30-70% מT מקס. הרציונל לתדרים אלו של גירוי הוא כי T מקס מספק מידע חשוב של אירועי מכונות התכווצות / אפנון, בעוד ½ T מקס מספק מידע רלוונטי יותר לCa 2 + הרגולציה ותהליך ECC. כדי להיות בטוח שכוח מקסימאלי שהושג, קפאין 10-20 מ"מ ניתן להוסיף לפתרון הרחצה תוך גירוי השריר. אם כוח מקסימאלי הושג, הכח לא יגדיל בנוכחות קפאין. FF מוסט ימינה וכוחות נוטים להיות מעט גבוהה יותר בטמפרטורות גבוהות יותר ניסיוניות. באמצעות מערכת זו, כל תדרי גירוי אחרים יכולים להתבצע אם רצוי. הפרופיל מובחן ההתכווצות של שריר אדי המהיר glycolytic ושרירי soleus איטיי חמצונים הואמוצג באיור 2.
- לאחר איזון, עייפות השרירים ב½ T מקס ל5 דקות, עם מרווח של 2 של גירוי ומחזור עבודת 25% (איור 3). פרוטוקול מתיש ספציפי זה הוא האמין כדי לשקף טוב יותר את התרומה של reticulum Ca 2 + sarcoplasmic השחרור להתכווצות שרירים 31.
- לשחזר את השריר ב½ T מקס למשך 30 דקות או עד שהכח הוא יציב, במרווח דקות 1. פרוטוקול מייגע נוסף ניתן לבצע כיום באמצעות גירוי T מקס, שהוא האמין למשקף את המצב של מכונות כויץ 31. אפשרות נוספת היא להאריך את הפרוטוקול מתיש לintercalate רכבות גירוי שיוצר T מקס ו½ T מקס בעייפות ולהתאושש בשרירים עם אותו הסוג של גירוי.
- חזור על FF כמתואר בשלב 3.4; הרציונל הוא שהבדלים פנוטיפי בין ההבדלזני erent, מודלים למחלות או טיפולים תרופתיים ניתן לראות על ידי ניתוח FF לפני ואחרי העייפות. למשל יש לנו דיווח בעבר כי לאחר עייפות FF של שרירים מסוג פרא צעירים הוא העביר לשמאל, ואילו בשרירים משרירים בגיל, הוא עבר לצד ימין, מה שמרמז השפעות עצבות פרשים של עייפות שרירי שלד כפונקציה של הזדקנות.
- כדי לבחון את התרומה של Ca 2 + הכניסה התאי בהתכווצות שרירים, פתרון רחצה ניתן לשנות לתוך תמיסה המכילה שום Ca 2 + אבל 0.1 המ"מ EGTA (0 mM Ca 2 + פתרון Tyrode) 32. לחלופין, חוסמים שונים של ערוץ חנות המופעל ניתן ליישם את פתרון הרחצה. כמה דוגמאות הן: 2-aminoethyl diphenylborinate (2-APB), SKF96365, 3,5-BIS (trifluoromethyl) pyrazole 2 (BTP-2) וazumolene, 33 וכו ', 34. קפאין יכול לשמש כדי לחקור את תפקידו של הקולטן ryanodine, וניתן להשתמש בו כדי להעריך KCl הכלליתכונות קיטוב של תכשירים אלה. תרופות אחרות יכולות גם לשמש כדי לחקור אפנון החשוב בכח בעייפות על ידי Na +, K +, וNa +-K + המשאבות 35. רוב התרופות אלו יש גדלים קטנים יחסית ונראה לנטרל במהירות להכנות השרירים האלה כפי שרואים מההשפעות המיידיות שלהם. חוסר השפעה של כל תרופות שניתנו לא בהכרח אומר כי התרופה אינה יעילה ובדיקות נוספות באמצעות מינון גבוה הרבה יותר מזו המשמשת בניסויי סיבי שריר בודדים הן לפעמים הכרחיות.
- יישום ייחודי אחד של מערכת vivo לשעבר זו הוביל לגילוי האחרון של alternans המכונאי בtric-- / - שרירים 4, 30. Alternans מוגדרים כפרקי פרץ תנודות של כוח התכווצות בשלב הירידה של הפרופיל עייף אותו. במהלך אירוע כוח כויץ רגע יכול להגדיל מעל רמתו הקודמת בכח במהלךלעייף גירוי משום ש2 + Ca או יותר מתפרסם או מכונות כויץ הפכו רגישות יותר לCa 2 +. התפרצות כוח ההתכווצות צריכה להיות 50% גבוהים יותר מכוחו הקודם ואת ההתפרצויות יש לראות לפחות 10 פעמים במהלך תהליך גירוי עייפות 5-דקות. alternans המכונאי אינו נפוץ בשריר שלד מהעכברים מהסוג פרא, אבל ניתן לראות בכמה שרירים מוטנטים עם הפרעה של התאי Ca + 2 תהליך האיתות כגון סוג Trimeric התאי קטיון הערוץ (tric-) - / - שריר 4. alternans מכונאי יכול להיגרם באמצעות גירויים מעייפים, טיפול בחומצת קפאין וcyclopiazonic (רו"ח), ראה איור 3 להקלטת נציג alternans המכאני אלה. טבעה של תופעה זו הוא מסקרן למדי, ואילו שריר שמסוגל לעייף נראה לרגע כדי לייצר יותר כוח. בפרסום הקודם שלנו, שילוב עם התא הבודד Ca 2 + ניתוח מגלה שמראה של alternans הוא תוצאה של SR Ca 2 + עומס ולא יציב SR. אנו מאמינים כי הבנה עמוקה יותר של alternans עשויה להוביל להבנה טובה יותר של תהליך ECC.
- בסוף הניסוי, למדוד את האורך והמשקל של שרירים בודדים באמצעות קליפר מכויל ואיזון האנליטי, פלאש קפוא השריר בחנקן נוזלי ולשמור ב-80 ° C, שכן ניתן לבצע ניתוחים ביוכימיים בשרירים אלה. שרירים אלו יכולים להיות גם עורו מכאני לחיטוט מפורט של תהליך ECC, או להיות כימי עורו לקביעת מאפייני התכווצות מהותיים בהיעדר מנגנוני פיקוח ECC.
- כוח שרירי הקלטות (mV) מומר 1 גרם לכפות על סמך תוצאות כיול ולאחר מכן מנורמל שטח החתך הפיזיולוגי (PCSA) תוך שימוש בנוסחא הבאה: כוח השריר (N / 2 סנטימטר) = (כוח (ז) x שריר lengtשעות (סנטימטר) x 1.06) / (משקל שריר (ז) x 0.00981) 5,36. לחלופין, כוח השרירים יכול להיות מנורמל לחלבון מוחלט או תוכן יקטין כולל של שרירי הפרט באמצעות בדיקת חלבון ברדפורד / כימות מכתימה Commossie כחולה. בתנאים מסוימים ואילו מסת שריר יכול להיות מושפע באופן חמור כתוצאה ממחלות, הזדקנות, טיפולים תרופתיים, נורמליזצית כוח המבוססת על מסת שריר, חלבון ו / או תוכן יקטין יכולים לספק readout יציב יותר.
4. נציג תוצאות
כוחות אופייניים טמפרטורת חדר התכווצות של אדי וsoleus מגיבים לגירויים נמוכים, בינוניים וגבוהים בתדר מוצגים באיור 2. התכווצות האד הנגרמת על ידי גירוי 5 הרץ נשארת כמו פרכוסים בודדים עקב הפעולה המהירה של SERCA Ca 2 + ATPase והמהותי Ca 2 + מאפייני רגישות של מכונות התכווצות, בעוד התכווצות soleus של 5 הרץ מתחילה פתיל (ATPase האיטיד רגישות גבוהה יותר לCa 2 + של מכונות התכווצות), אבל כוחות השיא עדיין נפרדים. בגירוי רץ 20, התכווצויות אדי מתמזגות באופן חלקי ואילו זה של soleus מהווה כוח tetanic התמזג לחלוטין. בתדירות של גירוי שמייצרת גירוי T מקס, שיכול להשתנות בטמפרטורת חדר 80-110 הרץ ולאד 60-90 הרץ לsoleus, upstroke המהיר והרפיה מהירה של כוח tetanic באד הם ציינו, שהוא בניגוד ל המאפיינים האיטיים של השרירים. soleus 2B האיור מראים כי עקומת כוח בתדירות של שריר אדי מוסטת לימין בהשוואה לשריר soleus, המציין כי שרירי soleus רגישים יותר לשחרור Ca + 2 בכל תדר נתון גירוי בשל הנוכחות של השרירן האיטי וisoforms טרופונין. בנוסף, מנגנון ההתכווצות מגיב בעצמה רבה יותר יחסית בשריר soleus בתדרים נמוכים. איור 3 תכניתפרופיל sa רגיל עייפות של אדי (פנל העליון) וsoleus השריר (פנל באמצע). שים לב לירידה המהירה יותר בכוח ההתכווצות תחת גירוי מתיש בשריר edl והירידה הגבוהה יותר בכוח בסוף פרוטוקול עייפות 5-דקות. לבסוף, פרופיל alternan מכאני אופייני לשרירי מוטציה שמוצג באיור 3 (פנל תחתון), שהוגדר כהתפרצויות כוח רגעיות בשלב הירידה של פרופיל של עייפות שרירים. התפרצות כוח ההתכווצות צריכה להיות 50% גבוהים יותר מכוחו הקודם ואת ההתפרצויות יש לראות לפחות 10 פעמים במהלך תהליך גירוי עייפות 5-דקות.

איור 1. ציור סכמטי של מערכת contractility ערוץ 4-vivo לשעבר. פולסים כיכר גל מופק על ידי מחשב שליטת ממריץ גראס. שתי יחידות בידוד גירוי לסנן את הגירויים שמקורם elecיחידת הממריץ trical להסיר כל תנודות באות החשמלי ולהקים אות גל מרובעת יציבה. אות חשמלי זה מסונן נשלח ל4 חדרי הרחצה המכילים את האלקטרודות תיל הפלטינה מקיפה כל שריר מבודד. סופו של דבר, זה נוכחי על פני שתי אלקטרודות גירוי (הנקרא שדה) שיוצרים פוטנציאל פעולה, גרימת ההתכווצות של השריר. התכווצות זו מזוהית על ידי מתמרי כוח ספציפיים, ששודרו למגברי הגשר, מסונן, בממוצע (מיזוג אות) ונרשם על ידי תוכנת מחשב דרך ממיר / D.

איור 2. . כוחות התכווצות יציגים של אדי וsoleus () הנגרמים על ידי כוחות התכווצות 5 רץ (פנל העליון), 20 רץ (אמצע פנל) וכוח tetanic מקסימאלי (T מקס) (פנל תחתון); כניסה מראה שמץ של כוח ההתכווצות של ניזוק שריר; (ב)נציג להגדיר מראה את ההתכווצויות הבודדות של כוח לעומת יחסים בתדירות אדי (FF, פנל העליון) והעקומה זממה כתוצאה מFF (פנל תחתון). לחץ כאן לצפייה בדמות גדולה.
{kind=link}

איור 3. נציג לעייף פרופיל וalternans המכאני. פרופיל טיפוסי מהר ירידה מתישה של שריר אדי (פנל העליון) והירידה האיטית לעייף פרופיל של שריר soleus (פנל באמצע). לעייף גירוי מוביל להופעתו של alternans המכאני בtric-- / - שריר עם Ca 2 + מאפייני טיפול מופרע (פנל תחתון).
Discussion
המדידה של כוח ועיפנות כויץ חשובה להערכה הכוללת של תפקוד שרירי שלד. המטרה העיקרית של בדיקה זו היא לזהות שינויים בכוח השרירים ומאפיינים מעייפים תחת מצבים פתולוגיים מסוימים, כגון sarcopenia ועייפות שרירים, ולבדוק את ההשפעה של תרופות / ריאגנטים על התכווצות שרירים. מאז כוח השר...
Disclosures
אין ניגודי האינטרסים הכריזו.
Acknowledgements
עבודה זו נתמכה על ידי AHA SDG 10SDG2630086 לזאו X, RO1-AR061385 למא י וללכת גרנט RC2AR05896 לBrotto מ '
Materials
| Name | Company | Catalog Number | Comments |
| שם המגיב | חברה | מספר קטלוגים | תגובות (אופציונלי) |
| 2-APB | Tocris | 1224 | חוסם של מספר Ca 2 + ערוצי כניסה כולל SOC וTRP וכו ' |
| SKF96365 | סיגמא | SKF-96365 | חוסם של מספר Ca 2 + ערוצי כניסה כולל SOC וקולט בתיווך Ca 2 + כניסה וכו ' |
| BTP-2 | Millipore | 203890-5mg | חוסם SOC יחסית ספציפי |
| רו"ח | סיגמא | C1530 | חוסם SERCA הפיך |
| קפאין | סיגמא | C0750 | אגוניסט RyR פעולה המהיר |
| מערכת מיקלוש רדנוטי כבר ארבע יחידת רקמות איברי אמבטיה | מיקלוש רדנוטי כבר | 159920 | |
| תמיכת רקמות שילוב / אלקטרודה מגרה | מיקלוש רדנוטי כבר | 160151 | אנכי זגזג סוג עם תמיכת רקמות |
| Quad הגשר Amp | ADInstruments | FE224 | |
| PowerLab/400 | ADInstruments | מוצר זה אינו זמין עוד. בחר גרסה אחרת של מערכת איסוף הנתונים. | |
| מתמר כוח (5 מ"ג - 25 ז) | ADInstruments | MLT0201/RAD | |
| תרשים v4.02 | ADInstruments | LabChart 7.3 היא הגרסה האחרונה של תוכנת תרשים. | |
| S8800 כפול Pulse דיגיטלי ממריץ | טכנולוגיות GRASS | מוצר זה אינו זמין עוד. S88X כפול פלט כיכר דופק הגירוי הוא גירוי חדש יותר. | |
| יחידת בידוד שנאי RF | טכנולוגיות GRASS | דגם SIU5 | |
References
- Winegrad, S. Role of intracellular calcium movements in excitation-contraction coupling in skeletal muscle. Fed. 24, 1146-1152 (1965).
- Sandow, A. Excitation-contraction coupling in skeletal muscle. Pharmacol. Rev. 17, 265-320 (1965).
- Thornton, A. M. Store-operated Ca(2+) entry (SOCE) contributes to normal skeletal muscle contractility in young but not in aged skeletal muscle. Aging. 3, 621-634 (2011).
- Zhao, X. Ca2+ overload and sarcoplasmic reticulum instability in tric-a null skeletal muscle. J. Biol. Chem. 285, 37370-37376 (2010).
- Brotto, M. A. Defective maintenance of intracellular Ca2+ homeostasis is linked to increased muscle fatigability in the MG29 null mice. Cell Res. 14, 373-378 (2004).
- Florkin, M. Machina carnis. The Biochemistry of Muscular Contraction in its Historical Development. Med. Hist. 17, 316-317 (1973).
- Galvani, A., Aldini, J. De viribus electricitatis in motu musculari commentarius. ApudSocietatem Typographicam. , (1792).
- Fulton, J. F., Fulton, J. F., Wilson, L. G. . Selected Reading in the History of Physiology. , (1930).
- Piccolino, M. Luigi Galvani and animal electricity: two centuries after the foundation of electrophysiology. Trends Neurosci. 20, 443-448 (1997).
- Hodgkin, A. L. The Croonian Lecture: Ionic Movements and Electrical Activity in Giant Nerve Fibres. Proceedings of the Royal Society of London. Series B, Biological Sciences. 148, 1-37 (1958).
- Hodgkin, A. L. . The Sherrington Lectures VII the Conduction of the Nervous Impulse. , 71964 (1965).
- Ringer, S. A further contribution regarding the influence of the different constituents of the blood on the contraction of the heart. J. Physiol. 4, 29-42.3 .
- Ringer, S. Further experiments regarding the influence of small quantities of lime, and other salts on muscular tissue. J. Physiol. 7, 291-308 .
- Ringer, S., Buxton, D. W. Concerning the action of calcium, potassium and sodium salts upon the eel's heart and upon the skeletal muscles of the frog. J. Physiol. 8, 15-19 .
- Ringer, S. Regarding the action of lime, potassium and sodium salts on skeletal muscle. J. Physiol. 8, 20-24 (1887).
- Campbell, A. K. . Intracellular Calcium its Universal Role as Regulator. , (1983).
- Mol, J. . Cell Cardiol. 16, ll3-ll6 (1984).
- Ridings, J. W., Barry, S. R., Faulkner, J. A. Aminophylline enhances contractility of frog skeletal muscle: an effect dependent on extracellular calcium. J. Appl. Physiol. 67, 671-676 (1989).
- Fitts, R. H. The cross-bridge cycle and skeletal muscle fatigue. J. Appl. Physiol. 104, 551-558 (2008).
- Kolbeck, R. C., Speir, W. A. Diaphragm contactility as related to cellular calcium metabolism: Influence of theophylline and fatigue. American Review of Respiratory Disease. 139, 495 (1989).
- Kolbeck, R. C., Nosek, T. M. Fatigue of rapid and slow onset in isolated perfused rat and mouse diaphragms. J. Appl. Physiol. 77, 1991-1998 (1994).
- Moore, B. J. Diaphragm atrophy and weakness in cortisone-treated rats. J. Appl. Physiol. 67, 2420-2426 (1989).
- Lannergren, J., Westerblad, H. Force decline due to fatigue and intracellular acidification in isolated fibres from mouse skeletal muscle. J. Physiol. 434, 307-322 (1991).
- Westerblad, H. Spatial gradients of intracellular calcium in skeletal muscle during fatigue. Pflugers Arch. 415, 734-740 (1990).
- Zhao, X. Enhanced resistance to fatigue and altered calcium handling properties of sarcalumenin knockout mice. Physiol. Genomics. 23, 72-78 (2005).
- Wang, X. Cardioprotection of ischemia/reperfusion injury by cholesterol-dependent MG53-mediated membrane repair. Circ. Res. 107, 76-83 (2010).
- Cai, C. MG53 nucleates assembly of cell membrane repair machinery. Nat. Cell Biol. 11, 56-64 (2009).
- Shen, J. Deficiency of MIP/MTMR14 phosphatase induces a muscle disorder by disrupting Ca(2+) homeostasis. Nat. Cell Biol. 11, 769-776 (2009).
- Romero-Suarez, S. Muscle-specific inositide phosphatase (MIP/MTMR14) is reduced with age and its loss accelerates skeletal muscle aging process by altering calcium homeostasis. Aging (Albany NY). 2, 504-513 (2010).
- Yazawa, M. TRIC channels are essential for Ca2+ handling in intracellular stores. Nature. 448, 78-82 (2007).
- Brotto, M. A., Nosek, T. M., Kolbeck, R. C. Influence of ageing on the fatigability of isolated mouse skeletal muscles from mature and aged mice. Exp. Physiol. 87, 77-82 (2002).
- Zhao, X. Compromised store-operated Ca2+ entry in aged skeletal muscle. Aging Cell. 7, 561-568 (2008).
- Pan, Z. Dysfunction of store-operated calcium channel in muscle cells lacking mg29. Nat. Cell Biol. 4, 379-383 (2002).
- Zhao, X. Azumolene inhibits a component of store-operated calcium entry coupled to the skeletal muscle ryanodine receptor. J. Biol. Chem. 281, 33477-33486 (2006).
- Renaud, J. M. Modulation of force development by Na+, K+, Na+ K+ pump and KATP channel during muscular activity. Can. J. Appl. Physiol. 27, 296-315 (2002).
- Brotto, M. A. Functional and biochemical modifications in skeletal muscles from malarial mice. Exp. Physiol. 90, 417-425 (2005).
- Brotto, M. A. Hypoxia and fatigue-induced modification of function and proteins in intact and skinned murine diaphragm muscle. Pflugers Arch. 440, 727-734 (2000).
- Smith, M. A., Reid, M. B. Redox modulation of contractile function in respiratory and limb skeletal muscle. Respir Physiol Neurobiol. 151, 229-241 (2006).
- Bagni, M. A., Cecchi, G., Colomo, F. Myofilament spacing and force generation in intact frog muscle fibres. J. Physiol. 430, 61-75 (1990).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved