
Method Article
בחקירתו vivo של Translatome מערכת עצבים המרכזי על ידי Polyribosome היפוך חלוק
In This Article
Summary
פרוטוקול זה ממחיש שינויים מהותיים בחלוקת polyribosome כדי ללמוד translatome של in vivo דגימות במערכת העצבים המרכזית. זה מאפשר הערכה הגלובלית של רגולציה תרגום ותעתיק דרך הבידוד והשוואה של רנ"א הכל לריבוזום שברי RNA מאוגד.
Abstract
תהליכים רבים מעורבים בביטוי גנים, כולל שעתוק, תרגום ויציבות של mRNAs וחלבונים. כל אחד מהצעדים הללו בחוזקה מוסדרת, המשפיע על הדינמיקה הסופית של שפע חלבון. מנגנוני פיקוח שונים קיימים בשלב התרגום, עיבוד רמות ה-mRNA לבד אינדיקטור אמין של ביטוי גנים. בנוסף, רגולציה מקומית של תרגום mRNA הייתה מעורבת בעיקר בפונקציות עצביות, הסטה 'translatomics' למוקד תשומת הלב בנוירוביולוגיה. השיטה שהוצגה יכולה לשמש כדי transcriptomics הגשר ופרוטאומיקה.
כאן אנו מתארים שינויים חיוניים לטכניקה של חלוקה polyribosome, שחוקרת את translatome המבוסס על הקשר של mRNAs תורגם באופן פעיל לריבוזומים רבים וההשקעה שלהם בהפרש הדרגתיים סוכרוז. באופן מסורתי, עבודה עם in vivo דוגמאות, במיוחד של CENTRAמערכת העצבים l (CNS), הוכיחה מאתגרת בשל הכמויות מוגבלות של חומר ואת נוכחותם של מרכיבי רקמת שומן. על מנת לטפל בזה, הפרוטוקול המתואר הוא מותאם במיוחד לשימוש עם כמות מינימאלית של חומר במערכת העצבים המרכזית, כפי שהודגמו על ידי השימוש בכבל יחיד עכבר השדרה ובמוח. בקצרה, רקמות מערכת העצבים המרכזית המחולצים והריבוזומים תרגום הם משותקים על mRNAs עם cycloheximide. הנפקת המיאלין מתבצעת לאחר מכן להסיר רכיבים עשירים שומנים בדם. חלוקה מתבצעת על שיפוע סוכרוז בי mRNAs מופרדים על פי טעינת ריבוזומלי שלהם. שברים מבודדים מתאימים למגוון רחב של מבחני במורד הזרם, כוללים טכנולוגיות assay הגנום רחב חדשות.
Introduction
ביטוי גנים נקבע על ידי הפעולה המשולבת של שעתוק, תרגום ויציבות של mRNA וחלבונים, עם תרגום הנושא את ההשפעה הדומיננטית ביותר 1. עכשיו זה ברור שכל אחד מהשלבים הבאים הוא פיקוח הדוק. מיקרו-RNAs, היווצרות של גרגרי RNA, אלטרנטיבי וpolyadenylation cytoplasmic כמה דוגמאות לרגולציה שלאחר תעתיק של ביטוי גני 2,3. כל אחד ממנגנונים אלה uncouples תעתיק מתרגום ומשפיע proteome במערכת הביולוגית של עניין. לכן, באופן לא מפתיע, רמות ה-mRNA לבד readout מושלם של רמות חלבון 4. פרוטאומיקה כמותיים מספקת הערכה הישירה ביותר של ביטוי גנים, עם זאת, למרות ההתקדמות, עדיין יש מגבלות רבות על רגישות ורזולוציה רצף חלבון. לכן פונה translatome, הרפרטואר של mRNAs תרגום, מציע פשרה מצוינת בין לומדtranscriptome וproteome. זה יותר מדויק מאשר transcriptomics בהערכת ביטוי גנים סופי, ומספק גם כיסוי גבוה יותר ורזולוציה רצף מ פרוטאומיקה.
במערכות של יונקים, הרוב המכריע של אירועי תרגום יתחיל באמצעות ייזום כובע תלוי. קבוצה של גורמי חניכה אוקריוטים יחד עם מקטע ריבוזומלי הקטן 40S להרכיב בכובע '5 של ה-mRNA. המורכב ואז סורק את ה-mRNA ועם הגיע לתחילת קודון לאוגוסט, מגייס למקטע ריבוזומלי 60S הגדול כדי ליצור 80S מלא הריבוזום. תמורת שלב התארכות עם הריבוזום נע לאורך ה-mRNA, עם גורמי התארכות סיוע לשילוב של חומצות אמינו מtRNAs הטעון לשרשרת פפטיד המתהווה. הריבוזומים מרובים יכולים להמשיך לאורך mRNA אחד בו זמנית ובמספר הריבוזומים הקשורים הוכח לתאם עם השיעור של סינתזת חלבון 5,6. זה עושה את טעינת ריבוזומלי אינדיקציה אמינה לטראןslation ומאפשר הפרדה פעילה תרגום mRNAs המבוסס על קצב שקיעה. בנוסף לכימות של תרגום mRNAs, ניתן לקבל מידע רצף לזהות מוטיבים מעורבים בויסות תרגום. גם RNA מחייבים חלבונים ויכולים להיות מבודדים משברים שונים גורמי תרגום אחרים, להקל על המחקר וגילוי של חלבונים רגולטוריים בנושא.
במערכת העצבים, בקרת translational נקשרה לתהליכים כגון אחסון ה-mRNA, תחבורה וסינתזת חלבון מקומית. קונוסים צמיחה הוכחו נמל בריכה מקומית ספציפית של mRNAs שונה משאר האקסון 7. בנוסף, האקסונים להחזיק את היכולת לסנתז באופן מקומי חלבוני 8,9. כתוצאה מכך, שליטה מקומית של תרגום הפכה לנושא חיוני של מחקר בנוירוביולוגיה. הפוטנציאל של חלוקה polyribosome כדי לטפל בזה כבר הודגם במספר מחקרים, שבו הטכניקה שימשהלחקור האקסון הדרכה בפיתוח חוט השדרה, והפגין את תרגום הפעילות תלויה של BDNF ב10,11 המוח.
Protocol
כל הניסויים בבעלי החיים בוצעו בהתאם להנחיות מוסדיות של DKFZ.
זהירות: כדי למנוע זיהום RNase של דגימות, לנקוט באמצעי זהירות בסיסית למניעת הזיהום RNase ולהכין את כל המאגרים במי DEPC שטופל.
1. הכנת עירובי סוכרוז
- הכן 17.5, 25.6, 33.8, 41.9 ופתרונות סוכרוז 50% (ראה טבלה 1). התחל בהכנה על ידי הוספה הדרגתיים לאט 2 מיליליטר של 50% תמיסת סוכרוז לחלק התחתון של צינור ultracentrifuge polyallomer. להקפיא את השכבה על ידי הנחת צינורות עבור 20 דקות ב -80 ° C.
- הוסף את השכבות הבאות בהפחתת אחוזי סוכרוז בצעדי הקפאה שביניהם, שהסתיימה עד סוכרוז 17.5%. שמור הדרגתיים על קרח במהלך תוספת של פתרונות סוכרוז כדי למנוע הפשרה של שכבות שמתחת.
- הכן הדרגתיים סוכרוז טרי יום אחד לפני השימוש ולשמור על 4 ° C. Alternatively, להכין הדרגתיים מראש, חנות ב -80 מעלות צלזיוס והפשרה על 4 מעלות צלזיוס בלילה שלפני הניסוי.
2. הכנת רקמה (כבל ו / או מוח השדרה)
- הרדימי עכבר על ידי מנת יתר של Xylazin וKetamin בNaCl וtranscardially תנקב עם 20 מיליליטר של התמיסה של האנק מאוזן מלחים (HBSS) המכילה 200 CHX מיקרוגרם / מיליליטר שמשתק את הריבוזומים בתמלילים קשורים. לאשר הרדמה נכונה על ידי אי - היענות לצובט הבוהן.
- לחלץ רקמות במהירות. פתח את dorsally הגולגולת ולחלץ את המוח. בצע laminectomy של כל בית החזה ועמוד השדרה המותני ולחלץ את חוט השדרה. השתמש במוח כולו (~ מ"ג 400) או חתיכת חוט השדרה (~ מ"ג 90) 4 סנטימטר, בהתאמה.
- העבר את הרקמה לתוך מאגר המגון קר כקרח (ראה טבלה 1) (בחוט השדרה: 1 מיליליטר; מוח: 4 מיליליטר) וקוביות לחתיכות קטנות עם אזמל נקי כדי לאפשר ספיגה מוגברת של cycloheximide. Perfo rm כל צעדים נוספים על קרח.
- דגירה במשך 15 דקות וhomogenize רקמה באופן מכאני באמצעות homogenizer Dounce. המגון מבוקר בקפידה יש צורך להבטיח שגרעינים יישארו ללא שינוי. לטפל בכל הדגימות בדיוק באותו אופן, כדי להבטיח השוואתיים. הערה: לרקמת מוח: לשבש את רקמה על ידי 5 משיכות עם העלי בחוזקה הולם. לרקמה בחוט השדרה: לשבש ידי 5 משיכות עם העלי רפוי, ואחרי 5 משיכות נוספות עם העלי בחוזקה הולם.
- קח aliquots (400 μl למוח ו200 μl לחוט השדרה), הקפאת הבזק ולאחסן ב -80 ° C לסך בידוד RNA.
- הסר את הגרעינים ותא בלא הפרעה ושברי רקמה על ידי צנטריפוגה ב XG 500 10 דקות ב 4 ° C. צנטריפוגה במהירות נמוכה מונעת אובדן של הריבוזומים.
- ברי Lyse קרום בsupernatant בחינם הגרעינים ל30 דקות על ידי הוספת NP-40 וחומרי ניקוי deoxycholate נתרן (כל אחד לריכוז סופי של 1%).
"Jove_title"> 3. הנפקה המיאלין
- צעד זה מסיר רכיבי שומן מן המדגם שאחרת יהיה להסוות את האות בפרופיל polyribosome. ראשית, דליים מראש צינת ultracentrifuge והרוטור על 4 מעלות צלזיוס ולהכין 2 M, 1.1 M ופתרונות סוכרוז 0.9 M (ראו טבלה 1).
- מערבבים lysate עם 1.22 כרכים של 2 תמיסת סוכרוז M ולהעביר לתוך צינור ultracentrifuge פוליאתילן. למלא את כל הצינורות לנפח שווה של 10 מיליליטר עם 1.1 M תמיסת סוכרוז, לאחר מכן כיסויו בזהירות עם 0.9 M תמיסת סוכרוז.
- מניחים את צינורות ultracentrifuge לסלים ultracentrifuge טרום צוננים. צנטריפוגה במשך 3 שעות ב 100,000 XG ב 4 ° C. במהלך תהליך זה הריבוזומים להיות מופקד בגלולה ואילו רכיבי שומן יצופו למעלה ויישארו בsupernatant.
4. סוכרוז Gradient היפוך חלוק
- הסר את supernatant ו לפזר גלולה במאגר הומוגניזציה ב '(ראה טבלת מס' 1 ). מדוד את הספיגה ב 260 ננומטר עבור כל דגימה באמצעות Nanodrop או מכשירים שווי ערך ולנרמל את הכמויות כדי לטעון לפי הערך ספיגת.
- הנח הדרגתיים סוכרוז (הכנה מתוארת בסעיף קודם) לתוך דליי ultracentrifuge טרום צוננים. דגימות שכבה בזהירות על גבי השיפוע. הוספת שיפוע סוכרוז ריק אחד כמו שליטה טכנית.
- התאם את המשקל של כל אחד מהסלים עם חיץ הומוגניזציה. צנטריפוגה דגימות ל1.5 שעות ב285,000 XG ב 4 ° C.
- להתחיל בהכנות דקות ISCO fractionator 30 לפני סוף צנטריפוגה. אם דגם אחר של fractionator משמש, בצע את הפרוטוקול של היצרן.
- הגדר את הרגישות המתאימה (0.2 AUFS למוח או 0.05 AUFS לחוט השדרה) למנורת UV ולעבור אותו לחימום. להרכיב את מדקר הצינור, לחבר אותו באמצעות המשאבה מתגלגלת 60% תמיסת סוכרוז דרך מדקר הצינור.
- בדוק את ההתקנה על ידי שאיבת סוכרוז (זרימת עכברושדואר: 1 מיליליטר / דקה), ובמיוחד לוודא שאין דליפות בtubings אשר להכניס בועות לתוך השיפוע.
- קליטת רקע ריקה על ידי שאיבה ידנית חיץ שיפוע לתוך גלאי UV ותיקון קו הבסיס.
- מוציא בזהירות את דליי ultracentrifuge המכילים סוכרוז הדרגתיים עם דגימות משקעים מהרוטור ומניח אותם על קרח. הימנע מכל הקפצת של הדרגתיים על מנת למנוע אובדן של רזולוציה.
- הדרגות לרוץ על fractionator. הפעל הדרגות ריקות ראשון להעריך ספיגת רקע ולהבטיח התקנה טכנית מתאימה.
- צרף שיפוע לגלאי UV ולנקב את החלק התחתון של הצינור עם מדקר הצינור. התחל את השאיבה של סוכרוז 60%, שתחליף את השיפוע עם דגימות משקעים כלפי מעלה דרך גלאי UV ולתוך מתקן הטיפה.
- הספיגה ב 260 ננומטר מתועד על מנת ליצור את פרופיל הקליטה לדוגמה, עם פסגות מציינות sedimentation של mRNAs הקשורים ליחידות משנת ריבוזומלי, monosomes, ולאחר מכן מספר גדל והולך של הריבוזומים.
- לאסוף דגימות ל20 שברים באמצעות מתקן הטיפה (600 μl כל אחד, ב2 צינורות מיליליטר).
5. בידוד RNA מן הברים בודדים
- הוספת SDS 10% לריכוז סופי של 1% לשברים פרט ומערבבים היטב על מנת להתפתח חלבונים ולנתק הריבוזומים. בשלב זה ניתן להקפיא דגימות ב -80 ° C. בהתאם לשאלה יש לטפל, ניתן ונקווה שברים בהתאם לפרופיל הספיגה.
- בודד RNA עם מיצוי פנול / כלורופורם חומצי, אשר גם מסיר עקבות זיהום של ה-DNA.
- הוספת נפח אחד של פנול / כלורופורם חומצי (prewarmed לRT) לכל דגימה, חום 10 דקות ב 65 מעלות צלזיוס ולחץ שחרורו מתחת למכסת מנוע קטר לאחר מכן.
- צנטריפוגה דגימות עבור 20 דקות ב 17,000 XG ב RT. בזהירות העברת שלב מימית ל1.5 מיליליטר חדשצינור. להיות מודע להיפוך שלב בשברי סוכרוז צפופים, שבו השלב המימית עשוי להיות בתחתית לאחר צנטריפוגה.
- הוספת נפח אחד של isopropanol, 1/9 אצטט נפח נתרן (pH 5.2) וGlycoblue μl 1 לכל דגימה על מנת לזרז RNA. שמור דגימות שעה לפחות 1 ב -80 ° C.
- צנטריפוגה במשך 30 דקות ב 17,000 XG ו4 ° C. Glycoblue מספק הדמיה של גלולה. הסר supernatant, לשטוף גלולה פעם אחת עם קרח קר אתנול 80% וגלולים אוויר יבש. ממיסים גלולה במי RNase ללא.
- לכמת כמות הרנ"א על ידי Nanodrop ולנתח שלמות RNA על ידי שבב Bioanalyzer. RNA אשר מתקבל על ידי הפרוטוקול שהוצג מתאים לכל המצב של מבחני תפוקה גבוהה אמנות כולל microarray ורצף עמוק.
תוצאות
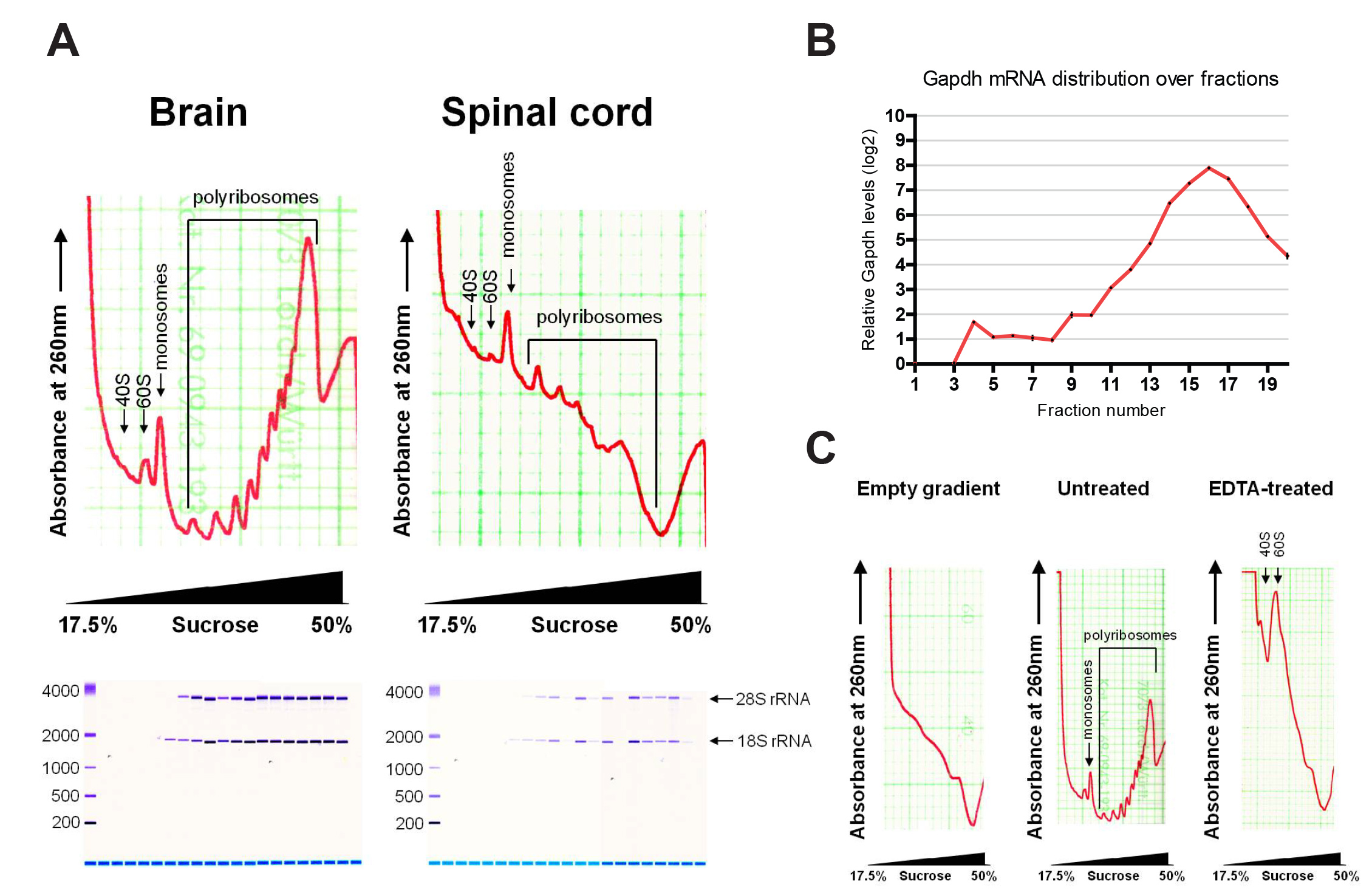
איור 2 מראה פרופילי polyribosome נציג לאחר חלוקה. הפרופילים של מוח וחוט השדרה להראות עקומות קליטה אופייניות כפי שתוארו קודמים לשורות תאים. mRNAs חייב יחידות משנה קטנות ריבוזומלי משקעים בשברים קלים יותר (40S) ומופיע לראשונה כשיא על הפרופיל, ואחריו למקטע הגדול ריבוזומלי (60S) וmonosome (80S) mRNAs כבולות. mRNAs חייב הריבוזומים מרובים משקעים שבברים כבדים יותר, עם הפסגות בהמשך מצביעים על מספר גדל והולך של הריבוזומים מאוגדים. תרגום עולמי ניתן להעריך מהפרופיל. כדוגמא, יש רקמת מוח polyribosome גבוהה יותר לmonosome יחס מאשר רקמה בחוט השדרה, מה שמעיד תרגום פעיל יותר.
RNA חילוץ משברים בודדים נבדקו על ידי Bioanalyzer, מראה הפצה של 18S ו28S rRNA. 18S rRNA מופיע מוקדם יותר בפרופיל, בהתאם ליחידות המשנה הקטנה ריבוזומלי סדימנטציה בsucr הקל שברים OSE. תשואות אופייניות של רנ"א הכל הן 10-20 מיקרוגרם למוח ו2-4 מיקרוגרם לחוט השדרה. תשואות RNA משברים בודדים הן עד 4 מיקרוגרם ו0.8 מיקרוגרם למוח וחוט השדרה, בהתאמה, בהתאם לחלק הקטן. שיפוע סוכרוז ריק כבר מראה כמה קליטת רקע ב 260 ננומטר, בשל נוכחותם של DTT. על רקע זה יכול להיות מופחת במהלך ניתוח הנתונים על מנת לנרמל את ערכי מדגם. טיפול EDTA קרס פסגות polyribosome, הוכחת פרופיל שקיעה נובע מתרגום.

איור 1. זרימת עבודה ויישומים אפשריים של in vivo חלוקה polyribosome.> לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

פרופילי איור 2. () פרופילים אופייניים polyribosome של מוח וחוט השדרה, עם הערכה של RNA שחולץ על ידי Bioanalyzer. הפצה של GAPDH mRNA על שברים ידי qRT-PCR (ב '). (C) Polyribosome של שיפוע ריק ומוח עם ובלי טיפול EDTA. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
| 1.1 | חיץ שיפוע 2x | 30 מ"מ טריס-HCl pH7.4, 30 מ"מ MgCl 2, 600 mM NaCl, 200 מיקרוגרם / מיליליטר cycloheximide (CHX), 2 המ"מ dithiothreitol (DTT) | |
| 1.1 | תמיסת סוכרוז 17.5% | 8.67 סוכרוז גרם, 25 מיליליטר 2x חיץ שיפוע, עד 50 מיליליטר H DEPC 2 0 | |
| 1.1 | תמיסת סוכרוז 25.6% | 12.8 סוכרוז גרם, 25 מיליליטר 2x חיץ שיפוע, עד 50 מיליליטר H DEPC 2 0 | |
| 1.1 | תמיסת סוכרוז 33.8% | 16.9 סוכרוז גרם, 25 מיליליטר 2x חיץ שיפוע, עד 50 מיליליטר H DEPC 2 0 | |
| 1.1 | תמיסת סוכרוז 41.9% | 20.95 סוכרוז גרם, 25 מיליליטר 2x חיץ שיפוע, עד 50 מיליליטר H DEPC 2 0 | |
| 1.1 | תמיסת סוכרוז 50% | 25 סוכרוז גרם, 25 מיליליטר 2x חיץ שיפוע, עד 50 מיליליטר H DEPC 2 0 | |
| 2.3 | המגוןחיץ | 0.25 M סוכרוז, 50 מ"מ טריס / HCl, pH7.4), 5 מ"מ MgCl 2, 25 מ"מ KCl | 200 CHX מיקרוגרם / מיליליטר, מעכב 1x רוש שלם פרוטאז, 1 מ"מ DTT, phenylmethanesulfonylfluoride 1 מ"מ (PMSF), 100 U / ml RNasin |
| 3.1 | תמיסת סוכרוז 2M | סוכרוז 68.4%, 50 מ"מ טריס-HCl, pH7.4, 5 מ"מ MgCl 2, 25 מ"מ KCl | 100 CHX מיקרוגרם / מיליליטר, מעכב 1x רוש שלם פרוטאז, 1 מ"מ DTT, PMSF 1 מ"מ |
| 3.1 | תמיסת סוכרוז 1.1M | סוכרוז 38.5%, 50 מ"מ טריס-HCl, pH7.4, 5 מ"מ MgCl 2, 25 מ"מ KCl | 100 CHX מיקרוגרם / מיליליטר, מעכב 1x רוש שלם פרוטאז, 1 מ"מ DTT, PMSF 1 מ"מ |
| 3.1 | תמיסת סוכרוז 0.9M | סוכרוז 30.8%, 50 מ"מ טריס-HCl, pH7.4, 5 מ"מ MgCl 2, 25 מ"מ KCl | 100 CHX מיקרוגרם / מיליליטר, מעכב 1x רוש שלם פרוטאז, 1 מ"מ DTT, PMSF 1 מ"מ |
| 4.1 | שעותomogenization חיץ B | 0.25 M סוכרוז, 50 מ"מ טריס / HCl, pH7.4, 5 מ"מ MgCl 2, 25mm KCl, 1% NP-40, deoxycholate נתרן 1% | 200 CHX מיקרוגרם / מיליליטר, מעכב 1x רוש שלם פרוטאז, 1mm DTT, 1 מ"מ PMSF, 100 U / ml RNasin |
טבלת 1. רשימה של מאגרים ופתרונות.
Discussion
למרות polyribosome חלוקה היא לא טכניקה חדשנית, הוא נשאר אחד מאתגר במיוחד. בהתבסס על חומר ההזנה, אופטימיזציה משמעותית יכולה להיות נחוצה. הדבר נכון במיוחד במקרה של in vivo דגימות של מערכת העצבים המרכזית, שבו כמות החומר היא לעתים קרובות הגבלה ושומן מרכיבי רקמה לעכב בידוד של mRNAs תרגום. פרוטוקולים שפורסמו חלוקה ביותר להתמודד עם שמרים או שורות תאי יונקים, ויש פרוטוקולים שהוקמו למוח 12,13,14. לעומת זאת, בקושי יש פרסומים המתארים חלוקה של מיתרי השדרה, ופרוטוקולים קודמים דורשים חוט השדרה ממספר רב של בעלי חיים שאספו 15. מסיבות אלה, כמה שינויים מהותיים שנעשו על מנת להתאים את פרוטוקול חלוקה לרקמות מערכת העצבים המרכזית, כוללים חוט השדרה עכבר אחת. חוסר תנועת הריבוזום עם cycloheximide מבוצעת במהלך זלוף בעלי החיים, כדי למנוע ניתוק של הריבוזומים דוריןg התהליך הארוך של מיצוי רקמה. בהמשך לכך, RNAs polysomal המחולצים באופן ספציפי על מנת למקסם את תשואת polyribosome. ראשית, בשליטת המגון של הרקמה על ידי douncing, שומר על הגרעין שלם ומונע זיהום ה-DNA. הגרעין לאחר מכן הוסר באופן מהימן על ידי צנטריפוגה. השילוב של חומרי הניקוי NP-40 וdeoxycholate נתרן מבטיח תמוגה של reticulum ושחרורו של הריבוזומים הקרום הקשורים אליו endoplasmic. בנוסף, רכיבים סופג UV בתוך המיאלין העשיר השומנים לטשטש את פרופילי polyribosome. הנפקת המיאלין לכן יש צורך, שבו תרכובות צפופות כגון polyribosomes הם pelleted ותרכובות קלות כמו לצוף המיאלין ויוסרו 16. אז Polyribosomes הם משקעים על שיפוע סוכרוז 17.5% 50%. במקום ידני שכבות והקפאה כל שכבת סוכרוז, יכול גם להיות מוכן הדרגתיים בעזרת מערבל שיפוע. מיצוי של RNA משברים באמצעות פנול / כלורופורם חומציים מאפשר DNA שיש להסירו עם אובדן RNA מינימאלי. עם זאת, היפוך שלב עלול להתרחש בברים צפופים סוכרוז (מעל שבריר 16).
פרוטוקול זה מספק יתרונות על פני שיטות קונבנציונליות אחרות המטפלים ברמת התרגום. לדוגמא, שתי המדידה של S6 פוספורילציה (סמן תרגום בולט), כמו גם התיוג של רשתות המתהוות עם רדיואיזוטופים לתת מידע על רמות התרגום העולמיים אבל מגלה מעט על מה שדווקא הוא להיות מתורגם. Polyribosome חלוקה, ומצד שני, מאפשרת לא רק הערכה של תרגום העולמי, אלא גם בזהות של mRNAs תרגום והחלבונים רגולטוריים בנושא. יכול להתבצע בזמן אמת PCR כמו בRNAs המבודד לקריאה מהירה מmRNAs שנבחר, וmicroarrays ורצף RNA דור הבא ניתן לבצע ללימודים רחבים הגנום. אופטימיזציות שהוצגו כאן מאפשרות את הטכניקה לשימוש עם כמויות מינימליות של רקמות במערכת העצבים המרכזית, ולכן תקטנהing את מספר בעלי החיים הדרושים ושיפור איכות ניסיונית הכוללת על ידי הקטנת זמן עיבוד רקמה. מצד השני, טכניקה זו לא יכולה להבדיל בין הריבוזומים נתקע מאלו תרגום. תעתיקי ביצוע הריבוזומים נתקע יהיו משקעים שבברים הכבדים, ואת זה צריך לזכור כאשר לפרש את הנתונים.
קיימות טכניקות האחרונות שדווחו, כלומר RiboTaq ומלכודת, שבו הריבוזומים מסומנים עם תגי epitope או כתבי בהתאמה באופן ספציפי לסוג תא וקשור mRNAs מבודד על ידי immunoprecipitation 17,18. טכניקה זו יכולה להיות יחד כדי polyribosome חלוקה להציע ברזולוציה גבוהה הקריאו לאוכלוסיות תאים ספציפיות. רגל הדפסת ריבוזום היא טכניקה נוספת שרומן כרוך עיכול nuclease ליצור שברים קטנים, או "עקבות", של mRNAs שמוגנים על ידי הריבוזומים. לאחר מכן ספריות נוצרות מהעקבות הללו ורצף. prov שיטה זואידו ניתוח הגנום כולו קודון הספציפי על תרגום, והוא מסוגל לזהות הריבוזומים נתקע, לא באוגוסט להתחיל קודונים ומסגרות קטנות במעלה הזרם פתוחות קריאה, שלא יכולים להיות מושגת על ידי חלוקה polyribosome 19. עם זאת, polyribosome חלוקה יכולה להיות מצמידים את פרופיל הריבוזום לבידוד של שברים מתעכלים המכילים ריבוזומים בודדים, ובכך מעשירה למינים המולקולריים הנחוצים להכנת ספרייה שבה הוא נדרש לעתים קרובות טוהר גבוה מדגם. יחדיו, חלוקה polyribosome היא שיטה גמישה עם משמעות ארוכת טווח ויכולה להיות מצמידים את יישומים במורד הזרם שונים, כולל מבחני רחבים הגנום כמו הדור הבא של רצף.
Disclosures
אין ניגוד האינטרסים הכריז.
Acknowledgements
עבודה זו נתמכה על ידי המשרד הפדרלי הגרמני לחינוך ולמחקר (BMBF), הביולוגיה מערכות איתות בסרטן (הלמהולץ ברית במערכות ביולוגיה) והמרכז לחקר הסרטן הגרמני (DKFZ).
Materials
| Name | Company | Catalog Number | Comments |
| Hank’s balanced salts solution | Gibco | 14170-138 | |
| Tube, Thinwall, Polyallomer, 14 ml, 14 x 95 mm | Beckman Coulter | 331374 | |
| Diethylpyrocarbonate | Sigma | D5758 | caution |
| Cycloheximide | Sigma | C7698 | danger |
| Dithiothreitol | Sigma | 43815 | warning |
| Complete Protease Inhibitor Cocktail Tablets | Roche | 11697498001 | |
| Phenylmethanesulfonyl fluoride solution | Sigma | 93482 | danger |
| RNasin Plus RNase Inhibitor | Promega | N2611 | |
| Nonidet P-40 | Roche | 11332473001 | danger |
| Sodium deoxycholate | Sigma | D6750 | warning |
| Acid-Phenol:Chloroform, pH 4.5 (with IAA, 125:24:1) | Ambion | AM9722 | danger |
| GlycoBlue | Invitrogen | AM9516 | |
| Equipment | |||
| Dounce Tissue Grinder, 1 ml/7 ml | Wheaton | 357538/357542 | |
| Nanodrop 2000 | Thermo Scientific | ||
| SW 40 Ti Rotor, Swinging Bucket, Titanium, 6 x 14 ml, 40,000 rpm, 285,000 x g | Beckman Coulter | 331302 | |
| Density Gradient Fractionator | Teledyne Isco | ||
| 2100 Bioanalyzer | Agilent Technologies |
References
- Schwanhäusser, B., et al. Global quantification of mammalian gene expression control. Nature. 473 (7347), 337-342 (2011).
- Lau, A. G., et al. Distinct 3'UTRs differentially regulate activity-dependent translation of brain-derived neurotrophic factor (BDNF). Proceedings of the National Academy of Sciences of the United States of America. 107 (36), 15945-15950 (2010).
- Piqué, M., López, J. M., Foissac, S., Guigó, R., Méndez, R. A combinatorial code for CPE-mediated translational control. Cell. 132 (3), 434-448 (2008).
- Ingolia, N. T., Ghaemmaghami, S., Newman, J. R. S., Weissman, J. S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science. 324 (5924), 218-223 (2009).
- Grolleau, A., et al. Global and specific translational control by rapamycin in T cells uncovered by microarrays and proteomics. The Journal of Biological Chemistry. 277 (25), 22175-22184 (1074).
- Rajasekhar, V. K., Viale, A., Socci, N. D., Wiedmann, M., Hu, X., Holland, E. C. Oncogenic Ras and Akt signaling contribute to glioblastoma formation by differential recruitment of existing mRNAs to polysomes. Molecular Cell. 12 (4), 889-901 (2003).
- Zivraj, K. H., et al. Subcellular profiling reveals distinct and developmentally regulated repertoire of growth cone mRNAs. The Journal of Neuroscience. 30 (46), 15464-15478 (2010).
- Brittis, P. a, Lu, Q., Flanagan, J. G. Axonal protein synthesis provides a mechanism for localized regulation at an intermediate target. Cell. 110 (2), 223-235 (2002).
- Tcherkezian, J., Brittis, P. a, Thomas, F., Roux, P. P., Flanagan, J. G. Transmembrane receptor DCC associates with protein synthesis machinery and regulates translation. Cell. 141 (4), 632-644 (2010).
- Colak, D., Ji, S. -J., Porse, B. T., Jaffrey, S. R. Regulation of axon guidance by compartmentalized nonsense-mediated mRNA decay. Cell. 153 (6), 1252-1265 (2013).
- Lau, A. G., et al. Distinct 3'UTRs differentially regulate activity-dependent translation of brain-derived neurotrophic factor (BDNF). Proceedings of the National Academy of Sciences of the United States of America. 107 (36), 15945-15950 (2010).
- Del Prete, M. J., Vernal, R., Dolznig, H., Müllner, E. W., Garcia-Sanz, J. A. Isolation of polysome-bound mRNA from solid tissues amenable for RT-PCR and profiling experiments. RNA. 13 (3), 414-421 (2007).
- Esposito, A. M., et al. Eukaryotic polyribosome profile analysis. J. Vis. Exp. (40), (2010).
- Sampath, P., Lee, Q. Y., Tanavde, V., et al. Identifying translationally regulated genes during stem cell differentiation. Current Protocols in Stem Cell Biology. Schlaeger, T. 1, (2011).
- Chiu, F. C., Smith, M. E. Studies on rat spinal cord polysomes: postnatal development and experimental demyelination. Journal of Neurochemistry. 31 (4), 835-844 (1978).
- Larocca, J. N., Norton, W. T., et al. Isolation of myelin. Current Protocols in Cell Biology. Bonifacino, J. S., et al. Chapter 3, (2007).
- Heiman, M., et al. A translational profiling approach for the molecular characterization of CNS cell types. Cell. 135 (4), 738-748 (2008).
- Sanz, E., Yang, L., Su, T., Morris, D. R., McKnight, G. S., Amieux, P. S. Cell-type-specific isolation of ribosome-associated mRNA from complex tissues. Proceedings of the National Academy of Sciences of the United States of America. 106 (33), 13939-13944 (2009).
- Ingolia, N. T., Brar, G. a, Rouskin, S., McGeachy, A. M., Weissman, J. S. The ribosome profiling strategy for monitoring translation in vivo by deep sequencing of ribosome-protected mRNA fragments. Nature Protocols. 7 (8), 1534-1550 (2012).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved