
Method Article
מסך משתיק קול עמוק רצף-בסיוע, ספונטני ב ביקוע שמרים שמר הביקוע
In This Article
Summary
אנו מציגים את פרוטוקול מסך משתיק קול פשוט בהתכה שמרים. שיטה זו היא יעילה ללא מוטגן, סלקטיבי מוטציות מתרחשות לעיתים קרובות באתר גנומית יחיד. הפרוטוקול מתאים לבידוד מדכאי על פגמים צמיחה בתרבות נוזלי שנגרמות על ידי מוטציה או תרופה.
Abstract
המסך עבור אללים mutant לדיכוי פנוטיפי ליקויים שנגרמו על ידי מוטציה גנטית היא גישה רב עוצמה לזיהוי גנים השייכים מסלולים ביוכימיים הקשורים קשר הדוק. שיטות קודמות כגון ניתוח מערך גנטי סינתטי (התלמידים) וכן באמצעות אולטרה סגול (UV) או כימיקלים כמו אתיל methanesulfonate (EMS) או N-אתיל-N-nitrosourea (ENU), טכניקות מוטגנזה מכוונת אקראי היה בשימוש נרחב, אבל הם בדרך כלל יקרים, מפרך. כמו כן, שיטות סינון שמבוסס מוטגן אלה קשורות לעתים קרובות תופעות לוואי חמורות על האורגניזם, גרימת מוטציות מרובות מוסיפים למורכבות של לבודד את מדכאי. כאן, אנו מציגים פרוטוקול פשוט ויעיל כדי לזהות מוטציות משתיק קול מוטציות אשר מעניקים פגם צמיחה שמר הביקוע. ניתן לנטר את הכושר של תאים עם חוסר צמיחה סטנדרטיים מדיה נוזלי עשיר או סינתטי נוזלי מדיה עבור שחזור משתמש בקורא צלחת 96-ובכן אוטומטית על פני תקופה ארוכה. ברגע תא רוכשת מוטציה משתיק קול בתרבות, שצאצאיו לגבור של התאים הורים. התאים התאושש שיש יתרון תחרותי צמיחה על פני התאים הורים יכולים להיות לאחר מכן מבודד, backcrossed עם תאי הורים. מוטציות משתיק קול ואז ניתן לזהות באמצעות רצף הגנום כולו. באמצעות גישה זו, יש בהצלחה בודדנו מדכאי מרובים על הליקויים צמיחה חמורה נגרמת על ידי אובדן של Elf1, AAA + משפחה של ATPase חשוב גרעיני mRNA תחבורה ותחזוקה של יציבות גנומית. כיום יש מעל 400 גנים ב ס הביקוע עם מוטציות היוועצות פגם צמיחה. כמו רבים מן הגנים האלה הם uncharacterized, אנו מציעים כי השיטה שלנו לזרז את הזיהוי של אינטראקציות תפקודית הרומן עם גישה זו ידידותי למשתמש, תפוקה גבוהה.
Introduction
הבסיס להבנת פונקציונלי קישורים בין הגנים מסתמכת על היכולת לזהות את mechanism(s) המולקולרי שבאמצעותו גנטיים מורכבים מתפצלות לייצר פנוטיפים שונים1. שמרים ביקוע, שמר הביקוע (ס' הביקוע), הרוב המכריע של חלבונים הם לוותר על הכדאיות2. תוצאה זו לא מדברים על unimportance של גנים אלו, אלא מעדיף המנגנונים לפיצוי הסבוך של מסלולים ביוכימיים שאליו שייכים גנים כאלה. לנתח את המנגנונים לפיצוי האלה יצר מפות אפיסטזה, אשר חשפו אינטראקציות גנטי מקיף הרחיבה את ההבנה שלנו של משעולים הביוכימי פונקציונלי3,4.
פותחו שיטות תפוקה גבוהה (למשל, ניתוח מערך גנטי סינתטי או מועצת התלמידים) כדי לזהות את האינטראקציות גנטי הגנום כולו ניצני שמרים ולאחר שהורחבו לשימוש בביקוע-שמרים-5,-6. גישות כאלה לעיתים קרובות לסמוך על ספריה של זנים המכילים כל קיימא יחיד חלבון-קידוד גנטי מחיקות (בסביבות 3,300 מוטציות מחיקה הפלואידי מכסה מעל 92% של הגנום שמרים ביקוע), דורשים זרוע רובוטית לבצע חוצה גנטי בין זן של עניין כל זנים אפשריים ב- library6. עוד יותר, התלמידים טכניקות תלוי היכולת של ספריית זנים נכונה ויעילה ההזדווגות, פנוטיפ שנמצא חריג כדי 444 כעת המאופיינת בגנים הביקוע ס'2.
למרות המורכבות של אינטראקציות גנטי, משווה את פנוטיפ של זן נשיאת מוטציות בגנים שני פנוטיפ של שני זנים נשיאת מוטציות בודדות של כל הגן יכול לקבל שתי תוצאות בולטים: 1) היא מוטציה כפולה גן # מושגים בסיסיים גרוע יותר הצפוי אוילר הורים פנוטיפים בצורה של מחלה או, במקרה הקיצוני ביותר, הקטלניות. זה מכונה של אינטראקציה גנטית השלילית, היא בדרך כלל סימן לכך שני גנים לפעול במקביל מסלולים ביולוגיים. 2 פנוטיפ מוטציה כפול) עדיף השילוב הצפוי של פנוטיפים הורים, המכונה גם השפעה חיובית הדדית גנטי. השפעה חיובית הדדית גנטי הוא מעניין במיוחד משום שהוא מציין כי גנים אלו מתפקד באותו תהליך. שני גנים אינטראקציה חיובית יש שלושה קשרים פוטנציאליים: הגן המוטנטי עשוי למעלה-לווסת את הביטוי של הגן השני במסלול מקביל, שני גנים עשויים לעבוד ביחד בתוך מסלול אותו במורד הזרם אחת לאחרת או לקודד שני גנים חלבונים לתקשר ישירות עם כל אחד את השני. לכן, אינטראקציה חיובית גנטי ניתן למפות צמתים רגולטוריות ג'ין ולסווג גנים uncharacterized משעולים הביוכימי7,8.
משתיק קול הוא מוטציה אשר יכול להקל על בחילות פנוטיפ של המוטציה של גן אחר, בדרך כלל המייצג השפעה חיובית הדדית גנטי בין שני גנים9,10. מוטציות משתיק קול על לוקוס שונה מזו של המוטציה שהם לדכא נקראות מדכאי extragenic. . הם יקר במיוחד בלימוד מוטציות גנטיות כלכלית בהצלה לאכסון קטלני פנוטיפ (הידוע גם בשם אפקט לזרוס)11. יש להם גם יישומים פוטנציאליים טיפולית בטיפול מחלות תורשתיות12,13.
מכל הסיבות האלה, זיהוי מוטציות משתיק קול אורגניזמים שונים דגם יש כבר נרחב מנוצל כדי להקל על ההבנה שלנו של מסלולים ביוכימיים שונים14,15,16. הקרנה של מדכאי מבוסס בדרך כלל על פנוטיפ של המוטציה המדובר ודורש ניצוח אקראי מוטגנזה מכוונת לבודד מוטציות זה להקל על פנוטיפ. כמעט כל האורגניזמים דגם הקימו שיטות מוטגנזה מכוונת אקראי. לדוגמה, N-אתיל-N-nitrosourea (ENU) ו ethylmethanesulfonate (EMS), שני מוטגנים המסוגלים גרימת נקודת מוטציות ב- DNA, מועסקים נרחב במודלים שונים של חיידקים עכברים17,18,19 . בנוסף, מנגן כלורי זמן כבר בשימוש שמרים על היכולת של הקטיון מנגן לעכב את ה-DNA תיקון מסלולים20. גישה נפוצה אחרת היא מוטגנזה מכוונת UV-induced, אשר מפיק פירימידין מוטגניות הגנום כולו הדימרים21,22.
למרות הניצול של כימי מוטגנזה מכוונת כדי לזהות מוטציות משתיק קול היה פופולארי, לשיטה חסרונות רבים, כולל השימוש של כימיקלים מסוכנים, שיעורי הצלחה משתנה מאוד המבוא של המשתנים מבלבלים במיוחד שהציג תופעות הלוואי השליליות של מוטגן על תהליכים תאיים מרובים23,24. בנוסף, מוטגנזה מכוונת כימית גורם לעיתים קרובות מספר מוטציות בגנום המוסיף המורכבות של שימוש הגנטית ואת רצף טכניקות כדי לזהות את המוטציה המדויק זה הוענקו על פנוטיפ משתיק קול האורגניזם25.
לפנות את החסרונות של גישות מוטגנזה מכוונת הנוכחי, אנו מציגים שיטה למסך משתיק קול ספונטנית מוטציות שמרים ביקוע כי אין להסתמך על כל מוטגנים או מחיקת ספריה. השיטה מבודד מדכאי דרך וזמינותו הבחירה חיובי. העיקרון של שיטה זו מבוססת על היתרון צמיחה subpopulation משתיק קול מוטציה בתרבות נוזלי, אשר ניתן יהיה פיקוח על ידי בקורא צלחת אוטומטית. הזדווגות של מיוזה משמשים רק אם אחד רוצה לנקות את הרקע הגנטי או לאשר הנוכחות של אללים monogenic של מדכאי לפני רצף הגנום כולו. אם פנוטיפ דיכוי נגרם על ידי מוטציה יחידה, פנוטיפ משתיק קול הפרדת 2:2 לאחר backcrossing עם זנים הורים. ניתן לזהות מוטציות משתיק קול ואז באמצעות רצף הגנום כולו. אנו מציעים כי שיטה זו ישימה הקרנת מדכאי ב כל המיקרואורגניזמים יכולים לגדל אוכלוסיה גדולה בתרבות נוזלי.
Protocol
1. זן בנייה והכנות
- ליצור מוטציה או מחיקה של הגן (yfm, מוטציה האהוב שלך) באמצעות תקן מוטגנזה (SDM) כפי שתואר לעיל26.
- לפני שמתחילים את המסך, backcross (אופטימלי) זנים מוטציה עם זן פראי-סוג כדי לנקות את הרקע הגנטי ולהפיק טריים-נולד מוטציה תאים זנים הורים. צובעות את המתח הורים למושבות בודדים על לוחות סטנדרטיים מדיה עשירים. נבחר באקראי שמונה 16 מושבות עצמאית (משכפל ביולוגי) עם מוטציות הרצוי עבור וזמינותו קורא צלחת (ראה 3.1).
הערה: פרוטוקול זה יעיל רק כאשר זנים הורים יש גידול לערוק בתקשורת נוזלי (מינימלי או עשיר, עם או בלי סמים, או עם משמרות בטמפרטורה, הגורמות את הפגם צמיחה). כל זני הורים צריך להיות הפלואידי ובכך מסוגל גנטית חצה עם זנים אחרים הפלואידי בעל סוג ההזדווגות משלימים.
2. צלחת קורא assay
- עם המוליך סטרילי, לקחת כמות קטנה של כל המושבות מוכן בשלב 1.1 (הסכום המדויק אין צורך לחסן ההתחלה תרבות), מניחים microplate 96-ובכן פוליסטירן. להשעות כל אחת המושבות ב 200 µL של המדיה המתאימה נוזלי (עשיר או מינימלי, עם או בלי סמים). כוללים גם ריק עבור כל שורה בצלחת המכילה 200 µL של המדיה (ללא תאים).
- בניהול הפרוטוקול. הבא על הקורא לוחית זיהוי תוכנה מחובר בקורא-microplate אוטומטיות: הגדר תוכנית קינטי במשך 24 שעות ביממה ואת הטמפרטורה ב 30 מעלות צלזיוס, ברעידות רציפה מהר מסלולית (425 cpm, משרעת 3 מ מ). להגדיר את קורא אופטי למדידת פיזור אור באורך-גל של 600nm עבור צפיפות אופטית, והגדר את האור כדי לקרוא מתחת לצלחת בתדר הקריאה של 2 דקות (721 קריאות סה כ במשך 24 שעות ביממה לכל טוב).
- אחרי 24 h, להקליט את הקריאות צפיפות אופטית חסמה סופית (OD600 נמחק), השתמש בנוסחה הבאה כדי לקבוע את אמצעי האחסון הדרושים כדי לדלל את כל הדגימות אל יתר = 0.1:

הערה: לייצא את הנתונים בתוכנת reader צלחת, להשתמש בתכנת להוספת הנוסחה לעיל כפי פונקציה אצווה, תהליך האחסון דילול כדי לשמש מכל קידוח ניסיוני. - כל 24 שעות, לדלל את כל הדגימות באמצעות המדיה כמו יום 0 לקבל מנת יתר = 0.1 (בערך 1.5 x 106 תאים למ"ל) תוך שימוש בנוסחה המצוין בשלב 2.3. שמור כל הצמיחה עקומות שנוצרו מדי יום ורשום בכל המושבה בודדים המציג את שיעור צמיחה מוגברת, נשפט יתר הסופי הוא גבוה משמעותית מזה של קבוצה עם רקע גנטי זהה או על ידי עקומת צמיחה הדומה לזה של פראי-סוג מושבות.
הערה: assay זה בדרך כלל לוקח בערך 7-14 ימים. בצע את כל השלבים בתנאים סטריליים.
3. מבחר של מושבות משתיק קול ואישור של פנוטיפ.
- מן היום האחרון של וזמינותו קורא צלחת (שלב 2.4), שמור תרבויות נוזלי, יש קצב גדילה התאושש באופן ניכר, ככל הנראה על ידי קבלת מוטציה משתיק קול אשר יכול להקל על פנוטיפ של המוטציה הורים. העברת ומערבבים µL 250 נוזלי תרבות cryotube המכילה 250 µL של 50% גליצרול. פלאש להקפיא את התאים בחנקן נוזלי ולשמור הזנים ב- 80oC ללא הגבלת זמן.
- כדי לאשר המוטציה משתיק קול הוא רכיב גנטי תורשתי, השתמש בשיטות המעבר גנטי רגיל לחצות yfm P (עבור המתח הורים, בשימוש בתחילת וזמינותו קורא צלחת) עם yfm S (עבור משתיק קול, המתח נשמר בקצה של צלחת קורא וזמינותו). אם המוטציה משתיק קול הוא אכן רכיב גנטי תורשתי, yfm P × yfm S אמור להניב tetrads שבה אוחדו שתי המושבות של פנוטיפ בחילות של המתח הורים ויש אוחדו שתי המושבות קצב הגידול התאושש משתיק קול זן.
- מן הצלב של צעד 3.2, לאסוף שלוש מושבות בעלות פנוטיפ משתיק קול (זן S) ואת שלוש מושבות בעלות פנוטיפ הורים (זן P) מאותו גנטי לחצות (3 ביולוגי משכפל עבור כל אחד), והמשך עם מיצוי הדנ א הגנומי ו רצף השלבים שלהלן.
הערה: השלבים 3.2 ו- 3.3 מומלץ מאוד, אך אינם נדרשים. לחלופין, אחד יכול להתפשט התאושש תרבית נוזלית שנאסף 3.1 באמצעי עשיר לתוך המושבות יחיד ולאחר מכן נבחר באקראי שלוש מושבות כמו triplicates ביולוגי עבור רצף הגנום כולו ללא אישור גנטי נוסף. במקרה זה, שלושה triplicates הביולוגי של זן הורים אמור לשמש להשוואה רצף גנטי.
4. גנומית DNA החילוץ, ספריית ייצור, רצף.
- עבור מיצוי הדנ א, ספריית הכנה, רצף, באקראי משכפל ביולוגי שלוש לכל זן yfm P , ומשוכפלת ביולוגית שלושה זנים כל אחד בנפרד שהתעוררו yfm S חוצה גנטי (שלב 3.2) או צלחות שהופצו כדי להשיג אחת המושבות של המתח S (הערה שלב 3.3).
- גדלים זנים בתרבויות 10 מ"ל במדיה עשירים לשלב יומן אמצע (יתר = 0.5-0.8, על 0.75 – 1.2 x 107 תאים למ"ל), ולהשתמש של חממה חזק לגדול תרבויות נוזלי ב 30 מעלות צלזיוס ברעידות ברציפות ב- 250 סל"ד. איסוף תאים על ידי צנטריפוגה ב 4 מעלות צלזיוס למשך 5 דקות ב x-1000 g.
- להשעות מגורען בתאים µL 400 מאגר הפקת דנ א (2% טריטון X-100, 1% מרחביות, 100 מ מ NaCl, 10 מ מ טריס-קלרנית (pH 8.0), 1 מ מ נה2-EDTA), לאחר מכן להוסיף 400 µL של חרוזי זכוכית, µL 400 של אלכוהול פנול: כלורופורם: isoamyl 25:24:1. מערבולת נמרצות למשך 2 דקות ב 4 º C.
- להוסיף µL 200 נוספים של מאגר הפקת דנ א, מיקס על ידי היפוך מספר פעמים. צנטריפוגה למשך 5 דקות ב 4 ° C ב 20,000 x g.
- להעביר את פאזה מימית צינור נקי, להוסיף 20 µg של RNase A/T1 מיקס, דגירה ב 37 מעלות צלזיוס למשך 15 דקות.
- להוסיף אמצעי שווה של אלכוהול פנול: כלורופורם: isoamyl 25:24:1, ספין בשביל 5 דקות ב 4 ° C-x 20,000 g, ולאחר מכן להעביר את פאזה מימית צינור נקי.
- להוסיף אמצעי שווה של כלורופורם, לערבב על ידי היפוך מספר פעמים, ואז לסובב למשך 5 דקות ב 4 ° C ב 20,000 x g, ואז להעביר את פאזה מימית צינור נקי.
- DNA precipitate עם שני כרכים של 100% אתנול בתוספת 10% נפח של 3 מ' NaOAC (pH 4.3) ב-20 ° C כבר לפחות שעתיים, ואז לסובב למשך 5 דקות ב 4 מעלות צלזיוס בגובה 20,000. רגל x g ולאסוף בגדר.
- לשטוף גלולה (DNA זירז) פעמיים עם 70% צוננת אתנול (צנטריפוגה 20,000 x g, 5 דקות, 4 ° C), להשעות את צניפה ב- 50 µL 10 מ מ טריס מאגר (pH 7.4).
- שימוש הכנה ספריית קיט (ראה טבלה של חומרים) לפי ההמלצות של יצרן כדי להכין את ספריית רצף הגנום כולו.
הערה: מומלץ ערכת המופיעים בטבלה של חומרים משום שהיא מאפשרת הבנייה של ספרייה גנומית ללא הגברה PCR, אשר מצמצם את השגיאה מוטציות שנוצרו במהלך ה-PCR-הגברה. בנוסף, במהלך הכנת ספריה גנומית, אינם מאפשרים את החרוזים כדי יבש לחלוטין על ידי קיצור החרוז ייבוש זמן 1 – 2 דקות. - עבור הגז פרמטרים במהלך הכנת ספריה, להשתמש sonicator ממוקד (ראה טבלה של חומרים) מוגדר הגורם חובה 20%, הכוח שיא 175 W, עם 200 מחזורים לכל פרץ, ואת תדירות למצב גורף ב 5.5oC עד 6 מעלות צלזיוס במשך 45 s. לחילופין, להשתמש DNA ו chomatin הטיה המערכת (ראה טבלה של חומרים) עם ההגדרות הבאות: 50% משרעת ב 4 ° C עם מצב הדופק, 15 s s על ו-15 הנחה למשך 10 דקות, עם זמן העיבוד הכולל של 20 דקות.
- זה חיוני כדי להתמודד עם החומרים המסוכנים בשימוש בשלב זה בזהירות. התייעץ עם המתאים גליונות נתונים חומר בטיחות של המוסד בריאות וסביבה ואת הבטיחות במשרד לטיפול NaOAC, אתנול, אלכוהול פנול: כלורופורם: isoamyl 25:24:1, כלורופורם.
- רצף של ספריות גנומית וכתוצאה מכך. קורא הרצף כולו אמור לכסות לפחות שלוש פעמים של הגנום כולו עם הפתרון בטווח נוקלאוטיד יחיד. מומלץ לזווג-הסתיים רצף (או הטכנולוגיה העדכנית ביותר).
5. ביואינפורמטיקה ניתוח לזיהוי מוטציות משתיק קול
- לבצע את הניתוח ביואינפורמטיקה להתמקד על השינויים גנומית שמזוהים באופן עקבי בין הורים ומשוכפלת מודחקים yfm זנים של כל ביולוגית.
הערה: תהליך צינור מלא המתוארת להלן, אבל, בנוסף, קבצי BASH script טקסט רגיל שני, fastq_to_vcf.sh, vcfprocess.sh, כלולים כשפות תוספת חומרים כדי להציג דוגמאות של זרימת העבודה מן העיבוד הקריאות VCF משתנה הקבצים עיבוד של הצטלבות VCF קבצים, בהתאמה. - לקצץ קצר-קריאות באמצעות הטיה (https://github.com/jbpease/shear) ביצוע קווי הפקודה (כל שאר אפשרויות ברירת המחדל):
shear.py - fq1 $FASTQ1 - fq2 $FASTQ2 - out1 $OUTFQ1--out2 $OUTFQ2 \
--barcodes1 $BARCODE - פלטפורמה TruSeq - trimqual 20:20 \
--trimpolyat 0 - trimambig - filterlength 50 - filterunpaired - מפת מקריאה הפניה ס הביקוע הגנום v2.30 המתקבל PomBase (ftp://ftp.ebi.ac.uk/pub/databases/pombase/pombe/Chromosome_Dumps/fasta/) באמצעות BWA v0.7.1527. השתמש את שורת הפקודה הבאה (כל שאר אפשרויות ברירת המחדל):
bwa mem -t 8 $GENOME $OUTFQ1 $OUTFQ2 > $SAM1 - תעביר קבצים יישור סאם GATK מומלצות צינור28 שהתקשרת משתנה באמצעות GATK v3.629, PicardTools v2.5.0 (http://broadinstitute.github.io/picard) SAMtools v1.3.130. השתמש את שורות הפקודה הבאות ואת הפרמטרים (כל שאר אפשרויות ברירת המחדל):
java-Xmx30g-ג'אר picard.jar AddOrReplaceReadGroups קלט = $SAM1 \
פלט = $BAMMARKED RGID = 1 RGLB = lib01 RGPL = אילומינה \
RGPU = $BARCODE RGSM = $SAMPLENUMBER
bam - O fixmate samtools $BAMMARKED $BAMFIXED
samtools למיין bam - O -o $BAMSORTED /home/peasejb/tmp -T $BAMFIXED
samtools מדד $BAMSORTED
java-Xmx30g-GenomeAnalysisTK.jar -T צנצנת HaplotypeCaller \
-R $GENOME-אני $BAMSORTED - genotyping_mode גילוי \
-stand_emit_conf 10 - stand_call_conf 30 -o $VCFRAW - לדחיסה של אינדקס קבצי VCF באמצעות tabix:
bgzip $VCFRAW.vcf
tabix $VCFRAW.vcf.gz - השוואת קבצי VCF בקרב הורים, משתיק קול זנים רצף ושכפול באמצעות BCFtools v1.3.127. השתמש את שורות הפקודה הבאות ואת הפרמטרים (כל שאר אפשרויות שמאל ברירת המחדל):
bcftools isec - n + 1 $VCFPARENTAL1.gz $VCFPARENTAL2.gz $VCFPARENTAL3.gz \
$VCFMUTANT1.gz $VCFMUTANT2.gz $VCFMUTANT3.gz > common_variants.list
הערה: פקודה זו הניבה קובץ מקודד עם דפוסי בינארי בו רצף גרסאות המופיעים המוטציה הראשונה רק יהיה בקידוד בינארי "000100", המוטציה השנייה רק בתור "000010", כל שלוש המוטציות כמו "000111", ועוד. קבצים אלה נוצרו עבור כל ערכה של הורים, מוטציה לשכפל את קבצי VCF. - לקמפל את הקבצים הרשימה לצומת variant שם הקובץ מצורף לשם בכל שורה באמצעות הפקודה grep UNIX:
grep "." *.list > all.list - הפניה מקושרת variant רשימה מלאה עם הקובץ ביאור GFF3 הנוכחי (ftp://ftp.ebi.ac.uk/pub/databases/pombase/pombe/Chromosome_Dumps/gff3/schizosacchar omyces_pombe.chr.gff3) באמצעות קובץ script מותאם אישית פייתון (variant_characterize.py) כדי לזהות אתרים עקבי הסנ פ חלבונים אזורים (נרדף ושם נרדף), יונקות, 3′ UTRs ו ncRNA.
python3 variant_characterize.py - רשימת common_variants.list \
schizosaccharomyces_pombe.chr.gff3 - מנכ ל החברה \
-fasta Schizosaccharomyces_pombe. ASM294v2.30.dna.genome.fa \
-תבנית 000100.... all.list.filter.000100
חזור התסריט הזה שינוי התבנית--ואת הסיומת של קובץ הפלט (....) באמצעות הקובץ הבינארי
דוגמאות מילוי: 000010 000001, 000110, 000011, 000101, 000111 - לשלב את הפלט של כל הרצפים האלה script בקובץ המופרד, לתצוגה של גיליון אלקטרוני. הטבלה מוערת של משתנים כולל אלה המופיעות strain(s) מוטציה באחד או בשני ביחס לרקע. השדה flag בינארי מציין גם הופעתו זן מוטציה בודדת (000100, 000010, 000001), שני זנים מוטציה (000011, 000101, 000110), או כל שלושה זנים מוטציה (000111).
- לנתח את הרשימות פלט מוערת של משתנים לא נמצא בדגימות הורים, אך נמצאו אחת, שתיים או שלוש הדוגמאות מוטציה. הביאור מציין מיקום גנומית והן ברמה של variant (נרדף/אי-שם נרדף באזור קידוד, 3/5' UTR, ללא קידוד, וכו '.). מתוך רשימה זו של המועמד מוטציות, דוגמה מועמדים רלוונטיים חריפה עשויה להיות משתנה קידוד-שם נרדף המופיעים באופן עקבי כל שלושה זנים. סוג נוסף של מועמד חזק תהיה הצטברות של שונים-שם נרדף או בשם רגולטוריות מוטציות בגן זנים מוטציה המופיעים ביחד או בתוך אותו גן.
תוצאות
מוטציות הגדלים לאט להציג שחזור פנוטיפי בתרבות נוזלי
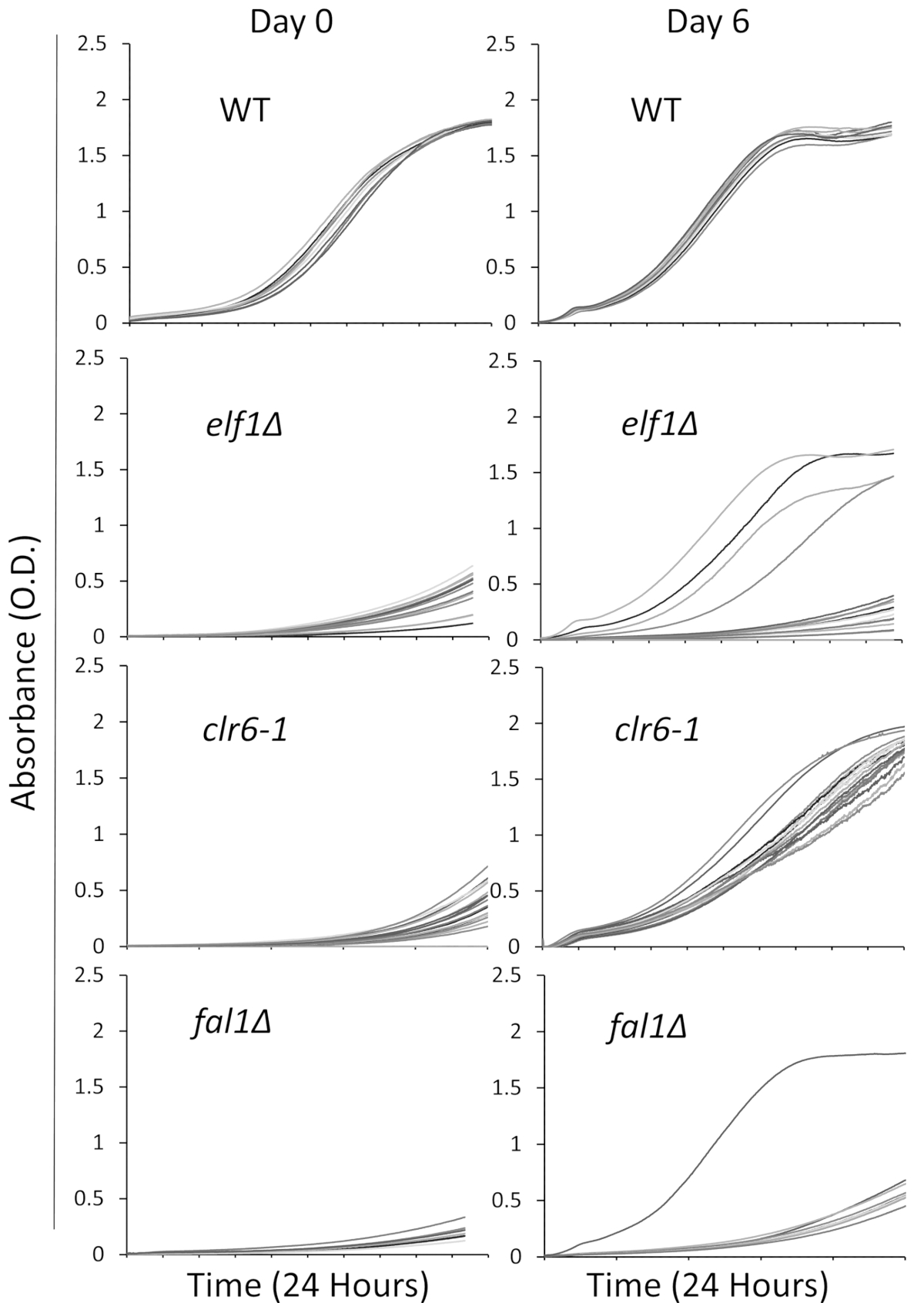
בחרנו שלוש מוטציות מעורבת במגוון מסלולים ביולוגיים עם פנוטיפ חולה, הגדלים לאט,: AAA משפחה ATPase Elf1, Deacetylase Clr6 היסטון, ומורכבים אקסון צומת רכיב Fal1. זן פראי-סוג וללחצים נשיאת מוטציות של גנים אלה שלוש זה היה כבר backcrossed עם פראי-סוג זנים היו קווצות שער למושבות בודדים ולאחר 16 מושבות יחיד נבחרו באופן אקראי להיות בתרבית בתקשורת נוזלי עשיר באמצעות הלוח 96-ובכן כמו המתואר לעיל. עקומות גדילה של מושבות בודדות נרשמו בנקודת הזמן הראשוני (יום 0) ו 6 ימים עם ניטור רציף באמצעות קורא צלחת. כצפוי, מושבות פראי-סוג הצג שינויים בולטים לא עקומות גדילה שלהם לאורך כל הניסוי31 (איור 1). ראוי לציין, ארבע המושבות עם רקע elf1∆ והמושבה fal1∆ אחד מראים תפנית דרמטית צמיחה הגדלים לאט כמה רמות משתנות של צמיחה דומה או קרוב לזה של מושבות פראי-סוג. באופן דרמטי, כל המוטציות clr6-1 הצג החלמה פנוטיפי עקבית, גדל בקצב מהיר יותר עד סוף assay31 (איור 1). כדי לאפיין את פנוטיפים שונים, אנו מתייחסים זנים המקורי כי הם הגדלים לאט "זני P" (או זנים הורים) זנים מציג שחזור פנוטיפי "S זנים" (או זנים מודחק). אנא שימו לב כי איור 1 דוגמה של סיבוב אחד של הניסוי ההקרנה, אינו מייצג את סך מדכאי שאינם משלימים מזוהה, וסודרו בתוצאות הבאות נציג.
שחזור פנוטיפי מיוחסות תכונות heritable
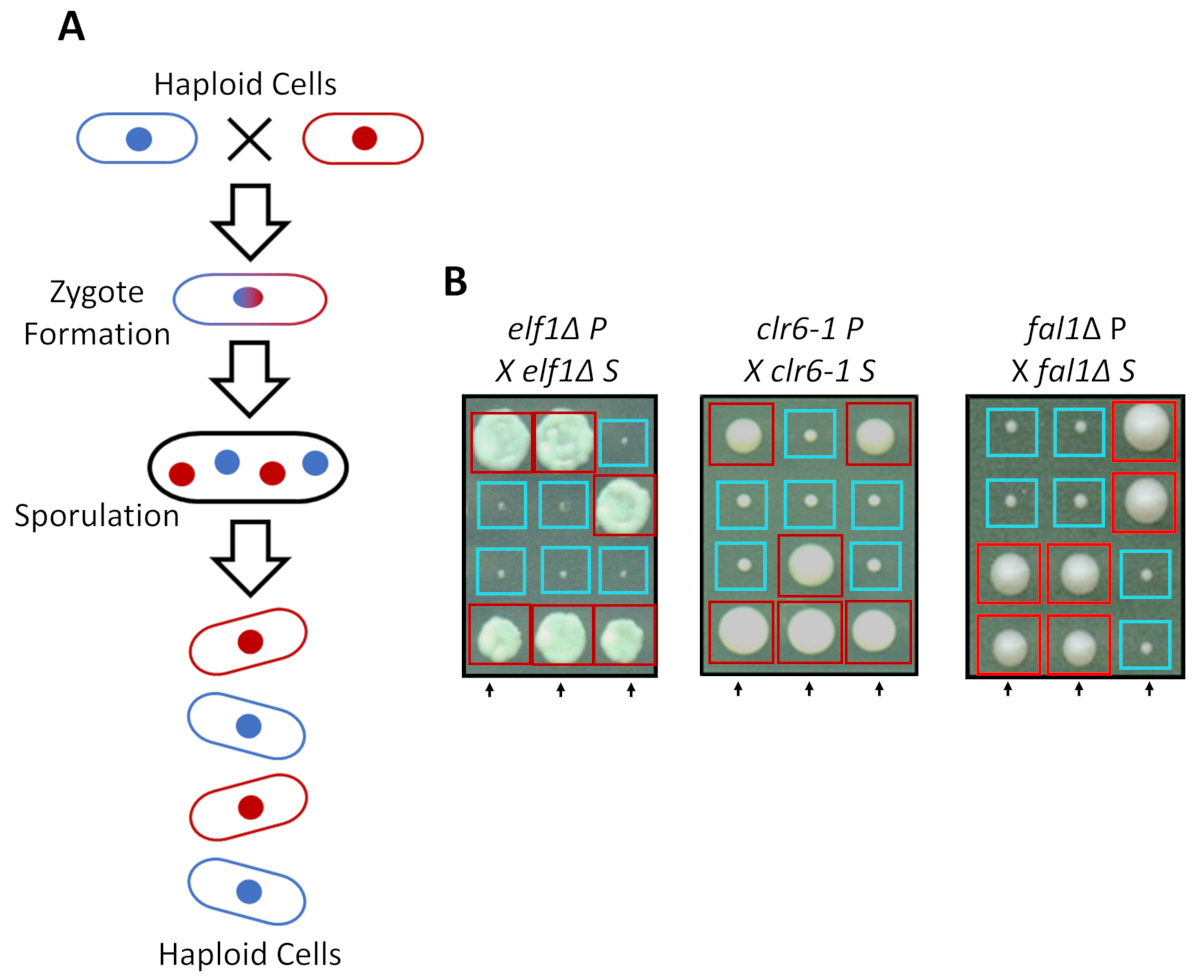
ס הביקוע גדלים ככל הפלואידי מדיה עשירה, אבל שני זנים הפלואידי עם חבר סוגי ההזדווגות משלימים תחת חנקן הרעבה. מיוזה בביקוע שמרים כרוך סיבוב של שכפול ואחריו שני סיבובים של חלוקת התא. מחזור מינית גורמת להיווצרות של ארבעה נבגים הפלואידי נושא החומר הגנטי של המתח הורים עם 2:2 סגרגציה של תכונות גנטיות לפי הכללים של גנטיקה Mendelian קלאסית (איור 2 א). כאשר גדל באותה צלחת עבור אותה כמות של זמן, אישרנו ההפרדה 2:2, כאשר כל הגב-המעבר דיכא זנים (S זנים) עם שלהם זנים הורים (P זנים), אשר הביא 2 קטנים (צמיחה פגם) ו-2 גדולים (משתיק קול פנוטיפ) מושבות. דוגמאות בודדות עבור תאים elf1∆, clr6-1 ו- fal1∆ מודחק מוצגים באיור 2B. יש לנו אישר כי כל זני S מבודד לבצע רכיב גנטי monogenic מעלימה את הגדלים לאט פנוטיפ של זנים שלהם P (נתונים לא מוצג).
רצף הגנום כולו בהצלחה מזהה מוטציות משתיק קול
לדוגמה, השתמשנו רצף הגנום כולו לזווג-קצה כדי לזהות מרכיבים גנטיים אחראי על התאוששות פנוטיפי ב- elf1∆ S זנים. תיאור מפורט יותר של ניתוח נתונים הוא זמין באינטרנט31. בקיצור, אנחנו מועסקים triplicates הביולוגי של שני זני elf1∆ P שנוצר באופן עצמאי, כפילויות הביולוגי של חמש קבוצות שאינן משלימות של זנים elf1∆ S , שכל אחד מהם מכיל מדכאי שונים. לאחר בידינו רשימה של משתנים המבואר מניתוח לביואינפורמטיקה (6.1-10), אנחנו עדיפות לשיעורים מסוימים של גרסאות שהיו רלוונטיים הניתוח שלנו. התמקדנו זיהוי של שינויים עקביים גנומית שהיו זהים בכל משכפל הביולוגי של הפרט זנים elf1∆ S לעומת זנים elf1∆ P הורים שלהם (איור 3 ו משלימה בטבלאות 1-4 ). זוהו חמישה שינויים תקליטורים אזורים בכל חמש elf1∆ S . זנים, כולל rli1 +, SPBPJ4664.02, cue2 + rpl2702 +nonsynonymous. S-A1 ו- S-A2 מכיל מוטציה SPBPJ4664.02, למרות מוטציות מתרחשות בחומצות אמינו שונות. מכיוון SPBPJ4664.02 הוא גן ארוך (נוקלאוטידים 11,916) עם מאות חזרה, מוטציות לא היו מסוגלים להיות מאושרות על ידי ביצוע PCR ואחריו רצף. S-A3 מכיל מוטציה מחיקה ב rli1 עקבי שבו הן כפילויות ביולוגי. עם זאת, החשבונאי לא שותף הפרדת עם פנוטיפ S רקע elf1∆ . זיהינו cue2 מוטציה (cue2-1) ב- S-B1, עם חומצות אמינו 396-400 חסרים. S-B2 מכיל מוטציה rpl2702 (rpl2702-1), אשר משתנה חומצת אמינו במיקום 45 גליצין אספרטט31. Cue2-1 ו- rpl2702-1 אומתו כמו מדכאי elf1∆ כמוצג להלן.
אישור גנטי של מוטציות משתיק קול מזוהה מאמת תורשתיות של פנוטיפ התאוששות
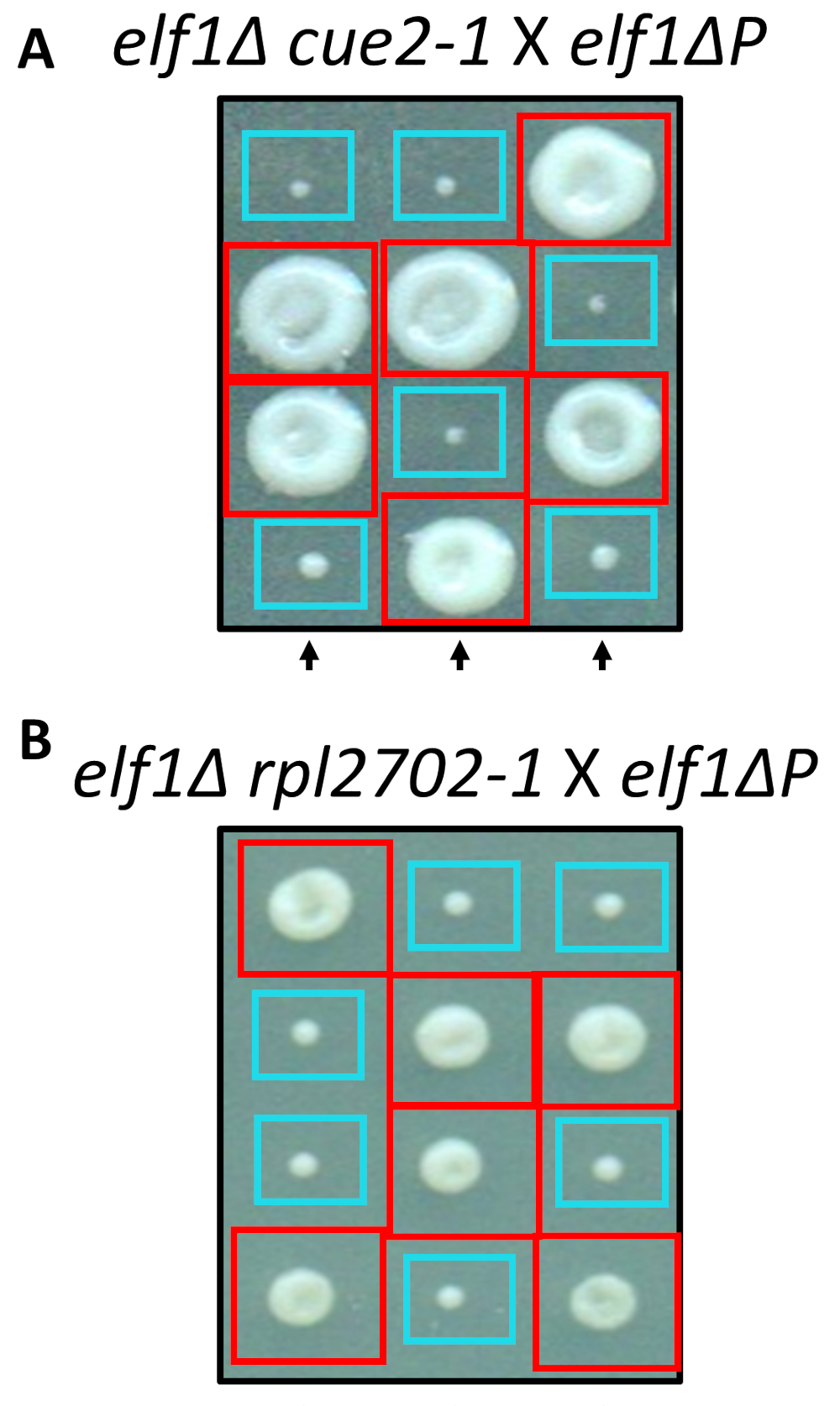
שני השינויים nonsynonymous מזוהה, cue2-1 ו- rpl2702-1, שוחזרו במעבדה באמצעות פרוטוקולים סטנדרטיים עבור מוטגנזה. מוטציה כפולה זנים cue2-1 elf1∆ P , rpl2702-1 elf1∆ P היו שלובות עם חינם elf1∆ P זן31 (איור 4). אם מוטציות nonsynonymous, זיהו דרך המסך הזה, היו מספיקות לדכא elf1∆ P, ואז tetrads שנוצר יראה 2:2 קטן יחס גדול במושבות הנובע הנבגים 4 ב כל דטרמיניזם. ואכן, מעבר גנטי הראה כי מוטציות משתיק קול מזוהה מצליחים לדכא את הגדלים לאט פנוטיפ של elf1∆ P , הן heritable.

איור 1: שחזור פנוטיפי ניתן יהיה פיקוח על ידי הקלטה עקומות גדילה קורא צלחת- 16 מושבות יחיד פראי-סוג (WT), elf1∆, clr6-1, fal1∆ היו ממוקמים בתוך צלחת 96-ובכן. עקומות גדילה הוקלטו על פני תקופה של 24 שעות ביממה, המושבות היו מדולל מחדש מדי יום במדיה עשירים. הפגם צמיחה ניכרת על ידי ספיגת נמוך (יתר) בסוף תקופת 24 שעות ביום 0. זני phenotypically התאושש הם אלו המציגים את עקומת גדילה דומה, או קרוב לזה של פראי-סוג על התקופה 24 שעות ביום 6. ארבע המושבות של elf1∆, מושבה אחת של fal1∆ושל כל מושבות של clr6-1 הראו רמות שונות של שחזור פנוטיפי לאחר 6 ימים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2: מעבר גנטי יכול לאשר כי ההתאוששות פנוטיפי מיוחס אלל heritable יחיד. (א) בעת ביקוע שמרים התאים ותוחלת חנקן הרעבה, שני תאים הפלואידי בעל סוג ההזדווגות משלימים ניתן להפיק זיגוטה אשר sporulates כדי להפיק של דטרמיניזם טכנולוגי 4 נבגים. החומרים גנטי הורים הפרדת במהלך המיוזה לפי הכללים של גנטיקה Mendelian. (B) Phenotypically התאושש מושבות (S שכותרתו, כי דיכאו) עם המצוין הורים אחרים היו חצה בחזרה עם למושבתם הורים ללא תשלום (המציגה לא שחזור פנוטיפי, שכותרתו P, עבור הורים). גנטי צלבים מציג 2:2 קטנים (כושר המסכן) למושבות (התאושש כושר גדול) מדגימים כי ההתאוששות פנוטיפי heritable, ניתן לייחס רכיב גנטי יחיד. התיבות האדומות הן מושבות נושאת אלל משתיק קול, הקופסאות הכחולות הם מושבות נושאת אלל הורים. איור זה השתנה מ Marayati ואח ', 201831. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3: ניתוח של נתוני רצף הגנום כולו כדי לזהות מרכיבים גנטיים אחראים של התאוששות פנוטיפי. ביולוגית שלושה ושכפול של שני זנים "P" הורים (P-A, P, B), ואת שני משכפל הביולוגי של חמש דקות phenotypically התאוששה החלפת זנים "S" (S-A1, S-A2 ו- S - A3 התאוששה P-A; S-B1 ו- S-B2 מ P, B), היו וסודרו מוטציות היו מאורגנים כמו רשימה של כל מוטציה בהמתח התאושש לעומת הגנום הורים זן זה נגזר מן (למשל, P-A לעומת S-A1, וכו '.). המספר הכולל של מוטציות שזוהו ברחבי הגנום כולו של כל השוואות pairwise היה 660. סכום כולל של 44 מוטציות אותרו כאשר מוטציות היחידה המתרחשים בשתי משכפל הביולוגי של המתח "S" באותו נבחרו. מתוך 44 מוטציות, מוטציות 12 היו הוספה/מחיקה (ויאסין) או אי - שם נרדף מוטציות. מתוך 12 ויאסין או מוטציות שם נרדף, חמש אירעה בהחלבון קידוד רצף. חמש מוטציות שעשויות להיות לתאם עם רכיב גנטי יחיד אחראי על זנים phenotypically התאושש: מוטציה שם נרדף בגן SPBPJ4664.02 שנמצאו ב S-A1 ו- S-A2, ויאסין ב rli1 שנמצאו ב S-A3, ויאסין ב- cue2 נמצאו ב S-B1 ושם נרדף מוטציות בגן rpl2702 שנמצאו ב S-B2. רצף מפורט מידע על מוטציות, על רקע מסוננים כלולה משלימה בטבלאות 1-4. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 4: אישור מדכאי מזוהה באמצעות רצף הגנום כולו. תוצאות של רצף הגנום כולו שאושרו על-ידי יצירת מוטציות באופן עצמאי ביצוע חוצה גנטית כדי לאשר את ההתאוששות פנוטיפי על ידי חציית זן elf1∆ cue2-1 עם זן elf1∆ P , ו elf1∆ rpl2702-1 עם זן elf1∆ P . שלושה tetrads אנכי נציג מוצגים. התיבות האדומות הן מושבות כפול-מוטציה (elf1 cue2-1, או elf1 rpl2702-1); הקופסאות הכחולות הם מושבות elf1∆ . איור זה השתנה מ Marayati ואח ', 201831. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
משלימה טבלה 1- אנא לחץ כאן כדי להוריד את השולחן הזה.
משלימה בטבלה 2 . אנא לחץ כאן כדי להוריד את השולחן הזה.
3 טבלה משלימה . אנא לחץ כאן כדי להוריד את השולחן הזה.
4 טבלה משלימה . אנא לחץ כאן כדי להוריד את השולחן הזה.
משלימה קידוד קבצים . אנא לחץ כאן כדי להוריד את הקבצים.
Discussion
הפרוטוקול המתואר כאן מייצג את הרומן של המסך פשוט מוטציות משתיק קול ספונטנית לזיהוי באמצעות שחזור פנוטיפי של מוטציות היוועצות צמיחה איטית בביקוע שמרים, הפנוטיפ האופייני של גנים מעל 400 ס הביקוע, הפונקציה של רבים מהם נשאר לא ידוע2,32. שיטות קודמות לקחו גישות אחרות על המסך עבור מוטציות משתיק קול מיקרואורגניזמים, לרבות השימוש מוטגנים21, או את היישום של שינוי טמפרטורה הרגיש רקע מוטציה33. לעומת זאת, פרוטוקול זה מראה התאוששות פנוטיפי מתרחשת ללא התערבות סביבתית/כימי נוסף מדגיש את היתרון כושר של עלייתו של דיכוי מוטציות בסופו של דבר משתלטים על המשאבים הזמינים בנוזל תרבות. מסך זה מאפשר בידודו של שניהם מדכאי מעקף או אינטראקציה עם מדכאי כי זה יעיל עבור שתי מוטציות אובדן-של-פונקציה כגון elf1∆ או fal1∆ , מוטציות נקודה כגון clr6-1, כל עוד המוטנטים להפגין כושר פגמים בתרבות נוזלי.
עד כה, כל זני S התאושש אנו חקרו הראו בדרגות שונות של שחזור פנוטיפי. כפי שאותרו במעבר גנטי, פנוטיפ התאושש הוא לייחס רכיב גנטי יחיד והוא heritable (דוגמאות באיור 2). זהו אחד היתרונות המשמעותיים של שיטה זו לעומת מסכי משתיק קול המבוססות על חומרים כימיים או מבוסס-UV, אשר לעתים קרובות יעד מרובים לוקוסים גנומית. נהוג לקיים מושבות אחד או שניים התאושש מתוך 16 מושבות/זן (בסביבות 10%) בתוך שבוע אחד. עם זאת, לב כי מוטציות מסוימות, כגון האובדן-של-הפונקציה של Rrp6, יחידת משנה הגרעין ספציפית exosome, מעולם לא החלימה לקצב גדילה פראי-סוג כמעט נצפו תאים elf1Δ 31. סביר להניח כי הפונקציה של Rrp6 יכולים רק חלקית יפוצו על ידי מדכאי, בניגוד הפונקציה של אחרים המוטציות שנבדקו, כולל fal1∆, אשר הוצגה לגרום פגם חמור meiotic דרך תפקידה החשוב של רגולציה שחבור34. אנו מאמינים כי משתיק קול אלטרנטיבי הקרנה שיטות תהיה נתונה באותה הבעיה כאשר yfg יש תפקידים ייחודי, שאינו ניתן להחלפה בצמיחת תאים.
לפני ביצוע רצף גנטי, רצוי הגב-צלב המושבות phenotypically המשוחזר, זוהה דרך הצלחת-הקורא, עם זנים הורים כדי לנקות את הרקע הגנטי להשיג משכפל ביולוגי. בנוסף, רצף הגנום כולו עמוק מזהה מאות שינויים נוקלאוטיד יחיד, אשר רובם אינם זהים בין הביולוגי משכפל יש עניין קטן להקרנה. לדוגמה, מצאנו בסך של 660 גנומית. שינויים על פני כל שלושת הכרומוזומים בין שני elf1Δ P את חמש S זנים (איור 3). לא נפוץ נתבונן מוטציות זהים בין הביולוגי ברצף ושכפול של כל זן, רומז כי מוטציות חדשות שעשויות להתעורר במהלך culturing של תאים elf1Δ לפני בניית ספרייה גנומית או שגיאות אקראיים עשויים להיות הציג במהלך בניית ספריה ואת רצף. ומכאן, לבודד מוטציות יהיו עקביים ברחבי משכפל הביולוגי הוא היבט חשוב בזיהוי מוצלחת מדכאי באמצעות רצף הגנום כולו.
אנו מזוהה, אישר מדכאי שני באזורים תקליטורים חמישה זנים S ברצף. למרות מוטציות בגן SPBPJ4664.02 התגלו ב S-A1 ו- S-A2 זנים, סביר להניח כי SPBPJ4664.02 הוא משתיק קול בתוקף כי S-A1 ו- S-A2 מכיל משתיק קול על אותו גן כפי שהם אינם משלימים עם אחד את השני ( נתונים לא מוצג). אנחנו גם לא אימת rli1 ב S-A3, אשר לא הפרדת במשותף עם פנוטיפ S כאשר backcrossed עם elf1Δ. לחלופין, מצאנו מוטציות ספציפיות באזורים ללא קידוד ב S-A1, S-A2 ו- S-A3. זה אפשרי כי שינו ללא קידוד גנומית אזורים אלה להקל על פנוטיפ elf1Δ , אשר יטופלו מחקרים עתידיים שלנו. לעומת שיטות מסורתיות כמו assay הצמדה, אשר עשויה להימשך שנים כדי למפות מוטציה גנטית, זיהינו שני מדכאי בתוך חודשיים לאחר שנוכח כי רכיב monogenic גרם של פנוטיפ S. עם מהירות הפיתוח הטכנולוגי רצף הגנום כולו, אנחנו אופטימיים כי שיטה זו תהיה יעילה יותר לזהות מוטציות גנטיות עקבי בעתיד הנראה לעין.
לסיכום, פרוטוקול זה מספק הנחיות צעד אחר צעד בהצלחה לזהות מוטציות משתיק קול עבור ג'ין כל עניין עם פגם הגדלים לאט בתרבות נוזלי. הפשטות של assay הזה מאפשר ההקרנה בקנה מידה גדול של רקע גנטי מרובים עניין עם קצת הדרכה מעשית. יש חדר נוסף למכן את התהליך באמצעות רובוט טיפול בנוזלים כדי לבצע את דילולים יומי. מאז מעבדה מניפולציה של מיקרואורגניזמים דורשת צמיחה בתרבות נוזלי, תהליך סלקטיבי מטבעו ופיטנס, אנו מציעים כי ניתן להחיל בהרחבה בפרוטוקול זה על אורגניזמים אוכלוסייה גדולה דגם אחרים כגון חיידקים, מינים אחרים שמרים.
Disclosures
המחברים מצהירים ללא חסויות על ידי יצרני הכלים לשימוש בשיטה זו אין אינטרסים כלכליים מתחרים.
Acknowledgements
עבודה זו נתמכה על ידי הלאומית המכון כללי רפואי למדעים, מענקים 1R15GM119105-01 כדי K.Z. אנו מודים כל הבודקים להערות תובנה. אנו מודים גם ג'יימס טאקר, אלישיה אנדרסון, אליזבת שחור, גלן Marrs הדיון והערות על כתב היד הזה.
Materials
| Name | Company | Catalog Number | Comments |
| Adenine, Powder | Acros Organics | 147441000 | Use at 75 mg/L to make liquid and solid rich media (YEA) |
| Bacteriology Petri Dish | Corning, Falcon | C351029 | 100 ×15 mm, use to grow strains to single colonies on solid rich media |
| D-Glucose Anhydrous, Powder | Fisher Chemical | D16-1 | Use at 30 g/L to make liquid and solid rich media (YEA) |
| Difco Agar, Granuated | Becton, Dickinson and Co. | 214530 | Use at 20 g/L to make solid rich media (YEA) |
| DNA extraction buffer | 2% Triton X-100, 1% SDS, 100 mM NaCl, 10 mM Tris-Cl (pH 8.0), 1mM Na2-EDTA | ||
| Focused-ultrasonicator | Covaris Inc. | S220 | Alternatively, use QSonica Q800R sonicator/DNA and chromatin shearing system |
| Gen5 Data Collection and Analysis Software | Biotek, Inc. | GEN5SECURE | Or equivalent, must be compatible with the micro-plate reader, use to export data readings from the micro-plate reader |
| Hydrochloric Acid 1N, Liquid | Fisher Chemical | SA48-4 | Use to adjust pH to 5.5 in liquid and solid rich media |
| Liquid Rich Media (liquid YEA) | 30 g/L D-Glucose, 5 g/L Yeast Extract, 75 mg/L Adenine, pH adjusted to 5.5 with 1 M HCl | ||
| Microplate Reader, Synergy H1 Hybrid Multi-Mode Reader | Biotek, Inc. | BTH1MG | Or equivalent, must read visible light at 600 nm wavelength range |
| Rich Media agar plates (YEA plates) | 30 g/L D-Glucose, 5 g/L Yeast Extract, 75 mg/L Adenine, 20 g/L Agar, pH adjusted to 5.5 with 1 M HCl. | ||
| RNase A/T1 mix | Thermo Fisher Scientific | EN0551 | Use according to manufacturer recommendation |
| Sterile Polystyrene Inoculating Loop | Corning, Inc. | OS101 | Or equivalent, use to transfer colonies from agar plates to 96-well plate |
| Sterile workspace and burners | |||
| Tissue Culture Plate, 96-well Optical Flat Bottom with Low Evaporation Lid | Corning, Falcon | C353072 | Or equivalent, must have optical flat bottom for micro-plate ready |
| TruSeq DNA PCR-Free LT/HT Library Prep Kit | Illumina, Inc. | 20015962 | Use to prepare the whole-genome sequencing library |
| Yeast Extract, Powder | Fisher Chemical | BP1422-500 | Use at 5 g/L to make liquid and solid rich media (YEA) |
References
- McKay, J. K., Latta, R. G. Adaptive population divergence: Markers, QTL and traits. Trends in Ecology and Evolution. 17 (6), 285-291 (2002).
- Wood, V., Harris, M. A., et al. PomBase: A comprehensive online resource for fission yeast. Nucleic Acids Research. 40 (D1), (2012).
- de Visser, J. A. G. M., Cooper, T. F., Elena, S. F. The causes of epistasis. Proceedings of the Royal Society B: Biological Sciences. 278 (1725), 3617-3624 (2011).
- Sailer, Z. R., Harms, M. J. Detecting high-order epistasis in nonlinear genotype-phenotype maps. Genetics. 205 (3), 107911088(2017).
- Kuzmin, E., Costanzo, M., Andrews, B., Boone, C. Synthetic genetic arrays: Automation of yeast genetics. Cold Spring Harbor Protocols. 2016 (4), 326-332 (2016).
- Tong, A. H. Y., Boone, C. Synthetic genetic array analysis in Saccharomyces cerevisiae. Methods in Molecular Biology. 313 (1), 171-192 (2006).
- Dixon, S. J., Costanzo, M., Baryshnikova, A., Andrews, B., Boone, C. Systematic Mapping of Genetic Interaction Networks. Annual Review of Genetics. 43 (1), 601-625 (2009).
- Boone, C., Bussey, H., Andrews, B. J. Exploring genetic interactions and networks with yeast. Nature Reviews Genetics. 8 (6), 437-449 (2007).
- Bai, X., Yang, Z., Jiang, H., Lin, S., Zon, L. I. Genetic suppressor screens in haploids. Methods in Cell Biology. , 129-136 (2011).
- Manson, M. D. Allele-specific suppression as a tool to study protein-protein interactions in bacteria. Methods. 20 (1), 18-34 (2000).
- Motter, A. E., Gulbahce, N., Almaas, E., Barabási, A. L. Predicting synthetic rescues in metabolic networks. Molecular Systems Biology. 4, 168(2008).
- Peterson, R. T., Shaw, S. Y., et al. Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nature Biotechnology. 22 (5), 595-599 (2004).
- Giorgini, F., Guidetti, P., Nguyen, Q., Bennett, S. C., Muchowski, P. J. A genomic screen in yeast implicates kynurenine 3-monooxygenase as a therapeutic target for Huntington disease. Nature Genetics. 37 (5), 526-531 (2005).
- Forsburg, S. L., Patton, E., et al. The art and design of genetic screens. Nature reviews. Genetics. 2 (9), 659-668 (2001).
- Johnston, D. S. The art and design of genetic screens. Genetics. 3 (March), 176-188 (2002).
- Jorgensen, E. M., Mango, S. E. The art and design of genetic screens: Caenorhabditis elegans. Nature Reviews Genetics. 3 (5), 356-369 (2002).
- Gocke, E., Müller, L. In vivo studies in the mouse to define a threshold for the genotoxicity of EMS and ENU. Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 678 (2), 101-107 (2009).
- Suzuki, T., Hayashi, M., et al. A comparison of the genotoxicity of ethylnitrosourea and ethyl methanesulfonate in lacZ transgenic mice (Muta(TM)Mouse). Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 395 (1), 75-82 (1997).
- Uttam, J., Alberico, C., De Stasio, E. ENU Mutagenesis. International C. elegans Meeting. , (1995).
- Putrament, A., Baranowska, H., Ejchart, A., Prazmo, W. Manganese Mutagenesis in Yeast. Methods in Cell Biology. 20, 25-34 (1978).
- Bose, J. L. Chemical and UV mutagenesis. Methods in Molecular Biology. 1373, 111-115 (2016).
- Ikehata, H., Ono, T. The Mechanisms of UV Mutagenesis. Journal of Radiation Research. 52 (2), 115-125 (2011).
- Shrivastav, N., Li, D., Essigmann, J. M. Chemical biology of mutagenesis and DNA repair: cellular responses to DNA alkylation. Carcinogenesis. 31 (1), 59-70 (2010).
- De Stasio, E. A., Dorman, S. Optimization of ENU mutagenesis of Caenorhabditis elegans. Mutation Research - Genetic Toxicology and Environmental Mutagenesis. 495 (1-2), 81-88 (2001).
- Probst, F. J., Justice, M. J. Mouse mutagenesis with the chemical supermutagen ENU. Methods in Enzymology. 477 (C), 297-312 (2010).
- Bähler, J., Wu, J. Q., et al. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 14 (10), 943-951 (1998).
- Li, H., Durbin, R. Fast and accurate short read alignment with Burrows – Wheeler transform. Bioinformatics. 25 (14), 1754-1760 (2009).
- Van der Auwera, G. A., Carneiro, M. O., et al. From fastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Current Protocols in Bioinformatics. 43, 11.10.1-11.10.33 (2013).
- Mckenna, A., Hanna, M., et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Research. 20, 1297-1303 (2010).
- Li, H., Handsaker, B., et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics. 25 (16), 2078-2079 (2009).
- Marayati, B. F., Drayton, A. L., et al. Loss of Elongation-Like Factor 1 Spontaneously Induces Diverse, RNase H-Related Suppressor Mutations in Schizosaccharomyces pombe. Genetics. 209 (4), 967-981 (2018).
- Harris, M. A., Lock, A., Bähler, J., Oliver, S. G., Wood, V. FYPO: The fission yeast phenotype ontology. Bioinformatics. 29 (13), 1671-1678 (2013).
- Xu, X., Wang, L., Yanagida, M. Whole-Genome Sequencing of Suppressor DNA Mixtures Identifies Pathways That Compensate for Chromosome Segregation Defects in Schizosaccharomyces pombe. G3: Genes|Genomes|Genetics. 8 (3), 1031-1038 (2018).
- Marayati, B. F., Hoskins, V., et al. The fission yeast MTREC and EJC orthologs ensure the maturation of meiotic transcripts during meiosis. RNA. 22 (9), 1349-1359 (2016).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved