
Method Article
הדמיה תלת ממדית של תאים חיידקיים לייצוגים סלולריים מדויקים ולוקליזציה של חלבון מדויק
In This Article
Summary
פרוטוקול זה מסביר כיצד להכין ולטעון דגימות חיידקיים עבור הדמיה תלת ממדית חיה וכיצד לשחזר את הצורה תלת ממדית של E. coli מתמונות אלה.
Abstract
הצורה של חיידק חשוב עבור הפיזיולוגיה שלו. היבטים רבים של פיזיולוגיה התא, כגון תנועתיות תאים, מקדם, וייצור ביוilm יכולים להיות מושפעים מצורת התא. התאים חיידקי תלת מימדי (3D) אובייקטים, למרות שהם מטופלים לעתים נדירות ככאלה. רוב טכניקות המיקרוסקופיה כתוצאה מתמונות דו ממדיות (דו-ממדיות) המובילות לאובדן נתונים הנוגעים לצורה בפועל של תא תלת-ממדי ולוקליזציה של חלבונים. פרמטרי צורה מסוימים, כגון עקמומיות גאוסיאנית (התוצר של שני העקמומיות הראשיים), ניתנים למדידה רק ב-3D מכיוון שתמונות דו-ממדיות אינן מדידת שני העקמומיות הראשי. בנוסף, לא כל התאים משקרים כאשר הדמיה דו-ממדית של תאים מעוקלים עשויה שלא לייצג במדויק את הצורות של תאים אלה. מדידת מדויק לוקליזציה של חלבון ב-3D יכול לעזור לקבוע את הרגולציה המרחבית ותפקוד של חלבונים. טכניקה קונבולוציה קדימה פותחה המשתמשת טשטוש פונקציה של המיקרוסקופ לשחזר צורות 3D תא ו להתאים במדויק חלבונים. כאן, פרוטוקול עבור הכנת והרכבה דגימות עבור הדמיה של תאים חיים של חיידקים ב-3D הן לשחזר צורה מדויקת התא וכדי להתאים את החלבונים מתוארים. השיטה מבוססת על הכנה לדוגמה פשוטה, רכישת תמונת פלורסנט ועיבוד תמונה מבוסס MATLAB. מיקרוסקופים פלורסנט רבים באיכות גבוהה ניתן לשנות פשוט כדי לקחת את המידות האלה. שחזורים אלה של התא הם אינטנסיביים מבחינה חישובית וגישה למשאבים חישוביים בעלי תפוקה גבוהה מומלצת, למרות שאין צורך בכך. שיטה זו חלה בהצלחה על מינים חיידקיים מרובים מוטציות, שיטות דימות פלורסנט, ויצרני מיקרוסקופ.
Introduction
תאים מכל הסוגים מסדירים את הצורות שלהם עבור פונקציות ספציפיות. לדוגמה, נוירונים הם בצורת שונה מאשר תאי דם יש פונקציות שונות. באופן דומה, תאים חיידקיים מגיעים במגוון של צורות וגדלים, למרות המטרה של צורות אלה לא תמיד ידוע1,2. לכן, חשוב כי הצורה של תאים חיידקיים ייקבעו במדויק. השיטה שמתוארים בה מציגה דרך מיושמת בקלות לאיסוף נתונים המתאימים לניתוח תלת-ממדי של תאי החיידקים החיים או הקבועים ביותר.
השיטה המתוארת מאפשרת לאחד לקחת תמונות תלת-ממד של תאים חיידקיים כדי לייצג במדויק את צורת התא 3D של המדגם להתאים במדויק חלבונים בתוך צורות אלה. טכניקות מיקרוסקופ מסורתיות לקחת תמונות דו-ממדיות, אשר הוא בעייתי כאשר לומדים תאים בעלי צורות חריגות או לא סימטרי, כגון מוטציות של es,או חיידקים מעוקל כגון vibrio החאריאווים הליקובקטר . בסדר, פילורי בעוד שתמונות תלת-ממד ברזולוציה גבוהה הן הקלט העיקרי לשיטה זו, השיטה אינה מחזירה תמונה משופרת ברזולוציה. במקום זאת, שיטה זו משנה את הקואורדינטות של פני השטח התלת-ממדית ואת צורת התא תוך שימוש באלגוריתם קונבולוציה קדמי תוך שימוש בקווי מחשוב פעילים והפונקציה המטשטוש לכאורה של המיקרוסקופ3 (איור 1). הוא שימש לחקר הומוטין החיידקי של מורד ב -E. coli4,5,6,7, הרומן periskeletal האלמנט החדש ב -V. המרה8, ואת הפום bactofilin CcmA ב הליקובקטר פילורי9 (איור 2).
הלוקליזציה של חלבונים יכול לספק תובנה פונקציות שלהם. לדוגמה, חלבונים המעורבים בחלוקת תאים מותאמים בדרך כלל ל-midcell10,11. מחקרים בתפוקה גבוהה נעשו כדי להתאים את כל החלבונים של חיידק בתקווה לצבור תובנה לתוך הפונקציות שלהם12. למרבה הצער, מחקרים אלה נערכו עם הדמיה דו-ממדית ו-1D או 2D ניתוח, מה שהופך את זה בלתי אפשרי למדוד היבטים ספציפיים של לוקליזציה חלבונים, כמו לוקליזציה לתכונות גיאומטריות סלולר.
לדוגמה, מרבי, חלבון דינאמי הנדרש לצורת מוט של חיידקים רבים, הוא מיפותזה לעבודה על-ידי הנחיית הלוקליזציה של סינתזה של קיר התא, והלוקליזציה שלה מראות את הלוקליזציה של הקיר התאים סינתזה7,13. מרבי ממינים מרובים מציג העשרה גיאומטריים4,6,7,14,15. פולימרים משטח דינמי, כגון MreB, יכול לזוג את הגיאומטריה של פני השטח עד הזמן העשרה בממוצע פרופיל16 ויכול להיות מסוגל לכוון את הגיאומטריות הספציפיות על ידי מזעור האנרגיה הקשורה בכריכה לקרום17. בעוד שחשיבותה של הפיתול, הכיפוף, הכפיית והדינמיקה לא נפתרה לחלוטין עבור MreB, חשוב לציין כי מדידות מדויקות של שני עקמומיות הראשי של משטח דורשים ייצוג תלת-ממד מלא של התא. לכן, כדי למדוד במדויק ביותר את עקמומיות שאליו חלבונים לוקליזציה, זה מועדף להשתמש 3D, במקום הדמיה דו-ממדית. 3D הדמיה מבטלת את הצורך להעריך מבחינה חישובית אלה עקמומיות אשר לא ניתן למדוד ב-2D, הערכה שעשוי לא להיות מדויק בתאים סימטרי18.
למרות הדמיה דו-ממדית של תאים הוא מהיר יותר ואינו דורש כמו עבודה חישובית הרבה יותר, הדמיה 3D מספק ייצוג מדויק יותר של התא, כמו גם את היכולת למדוד תכונות משטח, כגון עקמומיות, אשר לא ניתן למדוד ב-2D. לכן, כאשר הדמיה תלת-ממדית הופכת להיות שגרתי יותר, תובנות חדשות לצורת תא ולוקליזציה של חלבון יהפכו לאפשריות.
Protocol
1. הכנה לדוגמא
- הפוך את החיידק פלורסנט עבור הדמיה או על ידי הנדסה אותם לבטא חלבון פלורסנט cytoplasmic6 או באמצעות צבע ממברנה5. סוגים אחרים של חיידקים ניתן להשתמש במקום E. coli.
- להפוך את התאים באמצעות אלקטרופורציה עם פלסטלינה המקודד חלבון פלורסנט cytoplasmic mCherry.
הערה: חלבונים פלורסנט שונים של צבעים אחרים של ניתן להשתמש. - לגדל את הtransformants על צלחת LB עם אנטיביוטיקה המתאים ב 37 ° c. להשיג מושבות יחיד ולזהות שיבוטים חיוביים על ידי מיקרוסקופ. מושבות עמידות לאנטיביוטיקה המופיעות באדום מתחת למיקרוסקופ מכילות את הפלסמיד בנשיאת mCherry.
- להפוך את התאים באמצעות אלקטרופורציה עם פלסטלינה המקודד חלבון פלורסנט cytoplasmic mCherry.
- גדל 2 מ ל של תרבות הלילה במדיה נוזלית ליברות עם אנטיביוטיקה מתאימה ב-37 ° c באינקובטור רועד. השתמש או מושבה מצלחת או כמות קטנה של מלאי המקפיא כמו האינוניות.
הערה: השתמש באמצעי התקשורת הנחוצים לתאים החיידקיים שנבחרו לניסוי. - תת תרבות לילה 1:1000 לתוך 5 מ ל של מדיה ליברות טריים ולאפשר לתאים לצמוח לשלב מעריכי ב 37 ° c באינקובטור רועד עבור 3 עד 4 h. מדוד את ה-OD600 של התאים כדי לאשר שהם בשלב האקספוננציאלי (כלומר , OD600 = 0.2-0.4 לאחד).
הערה: ניתן לבצע הדמיה בכל שלב הצמיחה בהתאם לניסוי שבוצע.
2. הכנת שקופיות
הערה: חשוב להשתמש במדיה עם פלואורסצנטית אוטומטי נמוך. כל מדיום שיש לו התאמה אוטומטית נמוכה ניתן להשתמש. בניסוי זה, 1% agarose M63 רפידות מדיה מינימליות איטום עם VaLaP19 נדרש לתאי התמונה ב-3d.
-
הכינו פתרון של 1% של הצמח ב 20 מ ל של מדיה מינימלית.
- מיקרוגל את הפתרון עד הצמח הוא התפרקה לחלוטין והפתרון מופיע ברור.
- שמרו על הפתרון באמבט מים ב60 ° c עד שהוא מוכן לשימוש.
הערה: פתרון זה יכול להיות מתחזק ונמס לפי הצורך או יכול להיות מצוטט לכמויות קטנות יותר כי הם נמס לפי הצורך. לאחר שימושים מרובים המדיה עשויה להיות דהוי ויש להחליפו.
- ממיסים את האיטום על צלחת חמה ב-80 עד 100 ° c.
הערה: הכינו VaLaP, לשמש איטום, על ידי התכה 50 g כל אחד של ג'לי, לנולין, ו פרפין יחד בגביע על צלחת חמה ב 80-100 ° c. כמות גדולה זו של VaLaP ניתן לאחסן בטמפרטורת החדר ללא הגבלת זמן יהיה מספיק עבור > 500 שקופיות. חימום ב> 100 ° צ' יגרום לו לבזות וצבעו יהיה להכהות. - מקום שתי ערימות כל אחד 3 20 מ"מ x 20 מ"מ מחליק לכסות על הקצוות הפוכים של שקופית (שש כיסוי סה כ).
- פיפטה 200 μL של הפתרון pad על השקופית.
הערה: אם לא משתמשים בכתם פלורסנט מהונדס, ניתן להוסיף צבע ממברנה לפתרון הלוח האגקם לפני שלב זה. להשעות מחדש את הצבע במים ולהוסיף לצבוע 1 מ ל של הפתרון pad agarose לריכוז הסופי של 5 μg/mL. - מיד ובחוזקה המקום השקופית השנייה על ערימת שוברי כיסוי כדי לשטח את אגר ולתת לו לגבש עבור 1 דקות בטמפרטורת החדר.
- החלק בזהירות את השקופית העליונה.
- השתמש בקצה הגדול של העצה 200 μL המשמש לחיתוך רפידות בודדות מהג (~ 5 מ"מ בקוטר) בשקופית ולהשליך ג'ל פגום או מיותרים.
הערה: אם הדמיה זנים מרובים או תנאים, כרית נפרדת יהיה צורך עבור כל אחד. אם רק זן אחד הוא להיות בתמונה, לעשות 3 על 4 רפידות כדי לעזור לתמוך את coverslip. - בתאי הפיפטה למעלה ולמטה מספר פעמים כדי לשבש את גושי התאים ולהבטיח שהתרבות מעורבת היטב. פיפטה 1 μL של תת-תרבות משלב 1.3 על כרית.
הערה: שחזורים של צורת תא דורשים תאים בודדים שאינם נוגעים בתאים אחרים. אם שימוש בשלב נייח או בתרבויות צפיפות התא גבוה, ייתכן שיהיה צורך לדלל את המדגם 1:10 לפני הצבת אותו על הלוח. - תנו לדגימה להתייבש במשך 5 עד 10 דקות. ודא כי ה-droplet נספג לחלוטין בתוך הפנקס. אם נשאר נוזל כלשהו, התאים יזוזו מסביב לנוזל ואינם ניתנים לדימות.
- הצב שובר כיסוי על החלק העליון של הרפידות.
- לאטום את שובר הכיסוי עם VaLaP נמס על ידי צחצוח בעדינות סביב קצה המכסה לכסות עם ספוגית כותנה. הקפד לשמור אותו הרחק מראש שובר הכיסוי שבו המטרה תיגע. , הוולאפ יושם בתוך שניות. סוגרים את הדגימה
- התמונה מיד את המדגם.
הערה: המדגם צריך להיות בתמונה ברגע שהשקופית מוכנה. התאים יכולים לצמוח על הרפידות, ואם הם מחלקים את שחזורים יהיה קשה יותר.
3. אימאינטיטיטיטיטיטיקיטיטיטיטי
- ודא z או מיקוד הציר של המיקרוסקופ יכול לעשות תנועות מדויקות של פחות מ 50 ננומטר. השתמש בשלבים z piezo (ראה טבלה של חומרים), זמין על מיקרוסקופים כיתה מחקר, משום אופייני התקנים מוטורי המוקד אינם יכולים לספק דיוק זה.
הערה: בהנחה שקריטריון הדגימה של נייקוויסט-שאנון20, האדם צריך את היכולת להעביר 0.5 x את גודל השלב הרצוי הקטן ביותר, או 2x התדר המרחבי הרצוי. עבור הצעדים 100 ננומטר הדרושים בפרוטוקול זה, שלב עם דיוק של 50 ננומטר או פחות נדרש. -
ודא כי המיקרוסקופ מכיל מטרה 100x עם צמצם מספרי מינימלי (NA) של 1.45.
- איסוף z-ערימות של בין 200-400 תאים שונים כדי להבטיח מספיק תאים מתקבלים עבור יישומים במטה.
הערה: מספר התאים הדרושים תלוי בהבדלים הבסיסיים של מדגם הריבית. חלק מהתאים שנאספו בשלב זה לא יעבור את הצעדים המחודשת.
- איסוף z-ערימות של בין 200-400 תאים שונים כדי להבטיח מספיק תאים מתקבלים עבור יישומים במטה.
- ודא שמחסנית ה-z תואמת להגדרות המשמשות עבור ערוץ הצורה אם ערוץ פלורסנט משני (חלבון, תווית מטבולית וכו ') נמדד.
4. הדמיה
- הכנס את השקופית האטומה אל המיקרוסקופ ואפשר לו לשבת 5 דקות כדי להחליק את הטמפרטורה עם הסביבה, כי חדר המיקרוסקופ עשוי להיות בטמפרטורה שונה מאשר חדר ההכנה לדוגמה.
-
קח מחסנית פלורסנט מדגם.
הערה: מחסנית z צריכה לכסות לחלוטין את המדגם עם מרווח z פחות מעומק השדה. עבור מטרת 1.45 NA 100x ו-~ 1 יקרומטר עובי E. coli תאים, 40 שלבים ב 100 nm לכל שלב עבודה היטב. עבור תאים או תאים גדולים שאינם שוכבים באופן מושלם על פני השטח, ייתכן ש50 או יותר מהשלבים יהיו נחוצים. כלול מספיק שלבים וודא שהמדגם מטושטש לגמרי מעל ומתחת.- השתמשו בתוכנה המשויכת למיקרוסקופ (ראו טבלת חומרים) כדי לשלוט במיקרוסקופ.
- התמקד באמצע התא באמצעות גלגלי מיקוד מיקרוסקופ. תחת הרכישה הND בדוק את תיבת z כדי לקחת מחסנית z. לחץ על לחצן הבית כדי להגדיר את אמצע התא כנקודת ההתחלה. הגדר את גודל השלב ל-0.1 יקרומטר והגדר את הטווח ל-4 יקרומטר. ודא שההתקן Z מוגדר לשלב ה-piezo.
- הגדר את ערוצי הפלורסנט תחת חלון למדא להגדרות עבור מולקולות הפלורסנט הנמצאות בתמונה. בניסוי זה השתמשו ב-GFP ו-mCherry.
הערה: קח מחסנית z נוספת עם גודל וטווח של אותו שלב בערוץ הצבע השני אם התפלגות התלת-ממדית של ערוץ פלורסנט נוסף רצוי. בניסוי זה, השתמשו cytoplasmic mCherry כדי לקבוע את צורת התא ו-Mcherry GFP שימש בצבע השני הערוץ21. - ודא שסדר הניסוי מוגדר כסדרה למדא (z) כך שהוא ייקח מחסנית z שלמה בכל ערוץ צבע לפני המעבר.
- לחץ על הפעל כעת כדי להתחיל את רכישת התמונה ולשמור את הקובץ onece one או שניהם z-ערימות הושלמו.
- מעבר לאזור חדש בלוח וחזור על שלבים 4.2.2-4.2.5.
5. שחזור התא
- חתוך תאים בודדים ושמור את התמונות כקובץ tiff מוערם כך שיהיה רק תא אחד לכל קובץ. ודא שתא זה מבודד היטב מתאים אחרים (כלומר, בערך 5x ברוחב מלא חצי מקסימום של הפונקציה טשטוש ב-xy.
הערה: ניתן לעשות זאת באמצעות תוכנת ניתוח תמונה זמינה באופן חופשי (ראה טבלת חומרים).- צייר תיבה סביב תא בודד ושכפל את התא 2x, פעם אחת עבור כל ערוץ. ודא שהתיבה הכפלת היפר-מחסנית מסומנת ומשנה את הערוץ ל-1 או 2, וודא שהפרוסות כוללות את כל מחסנית ה-z.
הערה: אם הדמיה רק צורת התאים ולא חלבון פלורסנט נוסף, רק ערוץ אחד יהיה נוכח. - פעם שתי ערימות זמינים ללכת לתמונות | ערימות | כלים | שרשר כדי לשלב את התמונות עם ערוץ החלבון תחילה ואת ערוץ הצורה השני.
- שמור את התמונה החדשה כקובץ tiff.
- צייר תיבה סביב תא בודד ושכפל את התא 2x, פעם אחת עבור כל ערוץ. ודא שהתיבה הכפלת היפר-מחסנית מסומנת ומשנה את הערוץ ל-1 או 2, וודא שהפרוסות כוללות את כל מחסנית ה-z.
- מדדו את מטשטש את תפקוד המיקרוסקופ באמצעות דיפרקציה מוגבלת חרוזי פלורסנט22. זה צריך להיעשות עבור כל מיקרוסקופ מטרה מיקרוסקופ אבל ניתן לבצע לפני או אחרי הדמיה דגימות של עניין.
- ממוצע יחד חרוזים עצמאיים מרובים עם התערבות ידנית מסוימת באמצעות תוכנה זמינה (ראה טבלת חומרים).
הערה: המוצר הסופי צריך להיות תמונה תלת-ממדית של טשטוש הפונקציה עם מרווח xyz זהה כמו דגימות של עניין.
- ממוצע יחד חרוזים עצמאיים מרובים עם התערבות ידנית מסוימת באמצעות תוכנה זמינה (ראה טבלת חומרים).
- הפעל את הקונבולוציה הקדמי של הצורה העברה שחזור סקריפטים באמצעות תוכנה זמינה. הגירסה העדכנית ביותר של סקריפטים אלה ניתן להוריד באופן חופשי מ https://github.com/PrincetonUniversity/shae-cellshape-public.
- להפוך תיקיה בתוך תיקיה בשולחן העבודה המכילה את התמונות שנחתכו ואת הקובץ cell_shape_settings_tri. txt מתוך shae-cellshape-הציבורי/exampleData_tri.
- ערוך cell_shape_setting_tri. txt כדי לקבל את ההגדרות הנכונות עבור ניסוי הריבית. עבור ניסוי זה, קובץ ההגדרות כולל את השורות הבאות:
nm_per_pixel 70
Z_scale 0.65
stack_z_size 41
stack_t_size
Fstack_z_size 41
Fstack_t_size 1
stack_seperation_nm 100
Fstack_seperation_nm 100
psfScript-999 osuPSF20180726
מעבר צבע 1
הערה: קובץ טקסט זה מאורגן במקטעים וכל מקטע מנותח למשתנה משלו. בעוד שרבים מההגדרות אינן צריכות להשתנות מהתנסות להתנסות, יש לוודא שהגודל של מחסנית z (stack_z_size עבור הצורה, Fstack_z_size עבור חלבון הריבית), מרווח של מחסנית z (stack_seperation_ nm, Fstack_seperation_nm), שינוי מוקד יחסי של המיקרוסקופ (Z_scale)23, גודל הפיקסל ב-xy (nm_per_pixel), ובשם ה-script הטוען את הפונקציה הטשטוש של המיקרוסקופ (psfscript ) מתאים לניסוי. השדה ' מעבר צבע ' צריך להיות מוגדר כ-1 אם ערוץ הצורה ממולא ב-cytoplasmically ו-0 אם הוא אובייקט מוכתם ברפידה. - הפעל את Cell_shape_detector3dConvTriFolder פונקציה (shae-cellshape-הציבור/כדוריות הדואר/) עם המחרוזת למיקום התיקיה ואחריו את מספר התא להתחיל, ואת מספר התאים שברצונך להפעיל.
הערה: דוגמה לקלט תיראה כדלקמן: Cell_shape_detector3dConvTriFolder (' נתיב לתיקיה עם תמונות שנחתכו ', התחלת אינדקס בתיקיה, של תאים). תא אופייני עשוי להימשך בין 5 ל -20 דקות לצורך בנייה מחודש להתכנס ולהסתיים.
- הדפס את שחזורים התא כדי לוודא שהם נכונים לפני השימוש בתאים עבור כל ניתוח סטטיסטי.
- הפעל מתאים (shae-cellshape-ציבורי/Shae-fitViewerGui/) למסך חזותי שחזורים של תאים בודדים.
- לחץ על לחצן התיקיה בחר כאשר ממשק המשתמש הגרפי (GUI) ייפתח ולאחר מכן בחר את התיקיה עם קבצי הנתונים של השחזור (TRI. mat) שנוצרו בשלב 5.3.
- בחר את התיבה שליד שחזור התא אם תא מופיע כשאינו מוצג כהלכה או שלא התכנסת במלואו (איור 3). זה יכול להיראות כמו תא עם חור, צד שטוח, או ענף יוצא ממנו. פעולה זו תצרף 'דגל' לשם הקובץ כך שניתן יהיה להוציא אותו מכל ניתוח במורד הזרם.
הערה: ניתן להשוות את התא המשוחזר לתמונות המקוריות. זה יכול להיות חשוב במיוחד אם התאים שלך מגיעים מאוכלוסיה הטרוגניים שונים של צורות.
- הפעל את enrichmentSmoothingSpline (shae-cellshape-ציבורי/) כדי ליצור פרופיל העשרה של הריכוז היחסי של חלבון הריבית כפונקציה של עקמומיות גאוס על פני השטח.
- בחר כל תיקיה עם קבצים TRI. mat שנוצרו בשלב 5.3.3.
- בחר בקובץ העקומה החדש שנוצר (5.5.1).
הערה: קובץ עקום. mat ייווצר עבור כל תיקיה שנבחרה ב-5.5.1. כל הפרופילים ההעשרה יוצגו בגרף אחד. אם גרפים בודדים נדרשים מאשר להפעיל 5.5 עבור כל תיקיה.
הערה: בנוסף ללוקליזציה הגיאומטרי המדויק של חלבון פלורסנט, קיימות דרכים רבות אחרות לנתח את הנתונים מהערוץ המשני, כולל ספירת המספר, הגודל והכיוון של אובייקטים4.
תוצאות
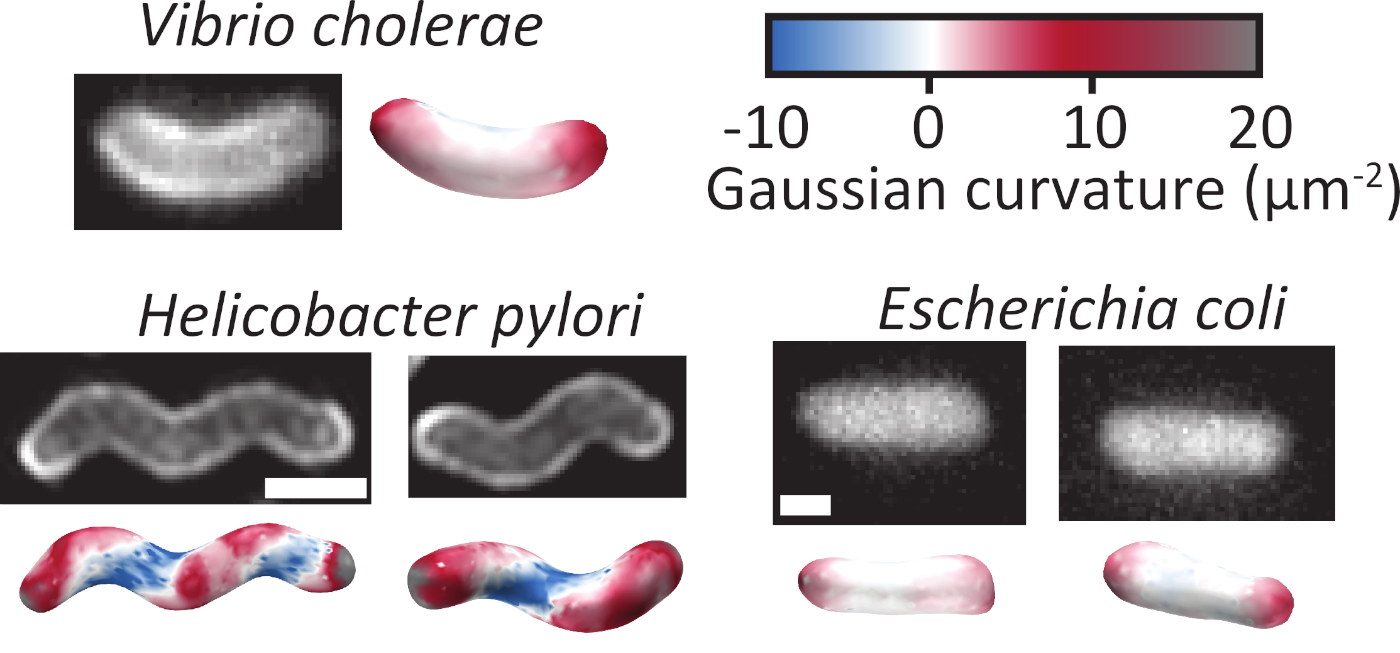
חיידקים מגיעים במגוון רחב של צורות וגדלים שעשויים לקבוע את התפקודים שלהם בטבע1. התוצאה של הליך זה היא ייצוג מדויק תלת-ממד של תאים מקונבולוציה קדימה של מחסנית z של תמונות (איור 1). שיטה זו חשובה במיוחד בהתמודדות עם תאים מעוקלים (איור 2), או עם תאים בצורת חריג (איור 4א), כייצוג דו-ממדי אינו משקף את העקמומיות של התאים במדויק. על מנת להשתמש בשיטה הקדמי קונבולוציה (איור 1א), התאים צריכים להיות מעניינת וכתם או יש כתם Cytoplasmic (איור 2B משמאל לעומת ימין).
איור 4 מציג את הלוקליזציה של mreb בתא. היתוך gfp נעשה לחלבון אקטין החיידקי מר21 על מנת ללמוד לוקליזציה המדויק שלה בשני פראי סוג המוטציות E. coli תאים4. מכיוון ש-MreB משויך לקרום, הדמיה תלת-ממדית נדרשת כדי לשחזר בנאמנות את מיקומו בתא. על-ידי ביצוע מדידות אלה ב-3D, הצלחנו לשחזר את הצורות של סוג פראי ותאי מוטציה Rodz (איור 4א). ניתן להעשיר את הלוקליזציה של רב-מימד בעקמומיות הקטנה של גאוס, תכונה גיאומטרית הניתנת למדידה רק ב-3D, באופן התלוי ב-RodZ (איור 4ב).

איור 1: השיטה הקונבולוציה קדימה משחזרת תאים ללא ידע מוקדם של צורת תא. (א) הפלט הסופי של השיטה הוא שחזור תא תלת-ממדי הנגזר על-ידי השוואת צורת בדיקה עם טשטוש תפקוד מכויל של המיקרוסקופ. זה מושווה עם מחסנית שנצפתה z עד התמונה 3D שנצפתה תואם היפותטי. התמונה המוצגת כאן היא עיבוד של שחזור תא אחד C. crescus . (ב) קו ההברחה של השחזור מעדכן את המיקומים המוערכים של רכיבי פני השטח בהתבסס על מחסנית התמונה הנצפתה. לפרטים אלגוריתמית מלאים על השיטה, ראו נגוין3. (ג) צינור השחזור מתחיל עם הידע לא פריורי של הצורה של התא ועדכון המיקום והמספר של קודקודים משטח להתאים את z מחסנית נצפתה. תמונות מייצגות מכל 30 שלבים במהלך שחזור תלת-ממד מוצגים מתוך שלוש זוויות שונות של תא V. המרה . מיקומי המשטח של צורה זו מתעדכנים כדי למזער את ההפרש בין הערימות המעובחנות והצפות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: נציג 3D שחזורים של שלושה מינים חיידקיים שונים בצורת. שחזורים תלת מימדיים ניתן לעשות מתאים עם הממברנה שלהם מוכתם (שמאל) או מלא עם fluoroplasmic (מימין). הצורה והעקמומיות של התא הפותח לא חשוב, כמוט מכופף, מוט מעוות, או מוט ישר כולם יכולים להיות משוחזר במדויק. המשטחים המשוחזר צבועים בהתאם לעקמומיות המקומי של המשטח. סרגל קנה מידה = 1 μm. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: שחזורים יכולים להיכשל ממספר סיבות. יש להקפיד על התאים כדי לוודא שאלגוריתם השחזור מתכנס לתוצאה סבירה. שמאל = סוג פראי מייצג (למעלה) ו- Rodz החשבונאי (למטה) התאים E. coli אשר עברו בקרת איכות. מימין = תאים שנכשלו לשחזר כראוי ולא עברו בקרת איכות. חמישה כיתות של התכנסות כושלים מוצגים: (i) תאים הקרובים מדי לקצה אזור החיתוך המוביל לתאים חדים, (ii), הקרובים מדי לתא אחר, (iii) שיצרו תאים מסוגים שאינם ממשיכים מעבר ל-s הראשוני מצב הסטה, ו (v) שגיאות לא ידועות אחרות. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: נתונים מייצגים המציגים לוקליזציה של חלבונים למטריות מסוימות של הסלולר. (א) תאים תלת מימדיים משוחזר של סוג פראי ו rodz E. coli תאים עם עקמומיות גאוסיאנית ועוצמת הקרינה של mreb מוצג. סרגל בקנה מידה = 1 μm. (ב) העשרה מגרשים מסוג פראי ו rodz E. coli תאים. ערכים > 1 מציגים העשרה וערכים < 1 מציגים דלדול של MreB ר מהאזורים הסלולריים האלה ביחס לכיסוי אחיד של התא. אזורים מוצללים מציינים את הממוצע של ± 90% ביטחון אתחול של הממוצע. עקומת כל זן הוא ליטת חיבור החלקה מעוקב נחתך באמצעות סף הסתברות עבור עקמומיות קיצונית של p > 5 x 10-3. איור זה שונה מ (בראטון ואח '). 4. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
שלב קריטי בפרוטוקול זה הוא רכישת תמונות באיכות גבוהה. כדי לשחזר את התאים כהלכה, חייב להיות טשטוש מספיק מעל ומתחת לתא. לכן, זה הכרחי כי מחסנית z נלקח מכסה מרחק גדול מספיק. מספר הצעדים שננקטו במהלך רכישת התמונה ניתן לכוונן עבור כל זן. לדוגמה, תאי קולי שנמחקו עבור rodz הם רחבים יותר ודורשים צעדים נוספים, ולכן, מרחק גדול יותר, מאשר תאים סוג פראי. אם המדגם הנמצא במהלך רכישת תמונה, השחזור יכול לקבל שגיאות מרכזיות. לכן, חשוב להניח לשקופית להגיע לשיווי משקל תרמי עם המיקרוסקופ לפני הדמיה כדי למנוע סחיפה במהלך רכישת מחסנית z. יש לאפשר התאמה של תאים על-גבי רפידות עם פלואורסצנטית אוטומטי נמוך. רכיבי מדיה, כגון אלה שנמצאו במדיום LB משותף, יש פלואורסצנטית אוטומטי שיכול לגרום לבעיות כאשר מנסים לשחזר את התאים. צפיפות התאים בלוח ההדמיה חשובה מכיוון שתהליך השחזור מבוצע באופן עצמאי בכל תא. תאים מעטים מדי יגדיל את הזמן הדרוש כדי להשיג תמונות של מספר מספיק של תאים, בעוד תאים רבים מדי יגרמו לשדות הדמיה כי הם צפופים מדי כדי לחתוך בקלות תאים בודדים. מכיוון שלא כל התאים משחזרים כראוי, יש לדמות תאים נוספים במהלך שלב הרכישה וכל התפוקות אמורות להיות מוקרן לפני המעבר קדימה עם ניתוח סטטיסטי (איור 3).
רבות מהמגבלות עבור שיטה זו הן טכניות. על המיקרוסקופ לשמש, אחד חייב להיות מטרה שיש לו צמצם מספרי גבוה (בדרך כלל > 1.4), כי זה מאפשר מעקב אופטי בקנה מידה של חיידקים. בנוסף, המיקרוסקופ צריך להיות מצויד בשלב piezo זה יכול לקחת צעדים קטנים, מדויקים בכיוון z. יתרה מזאת, בעוד שאין צורך בכך, גישה למשאבים חישוביים בעלי תפוקה גבוהה כדי להפעיל את תוכנת ניתוח התמונה מומלצת משום שהיא תקטין את זמן העיבוד לתאים משחזרים.
מגבלה רעיונית אחת לשיטה היא שצירי האנרגיה הנכונים לניפוח החלקות של השחזור ביחס ליחס האות לרעש של התמונות חייבים להיבחר. כדי לאמת מבחר פרמטרים, יש למדוד את הגדלים והצורות של התאים באמצעות שיטות עצמאיות כגון שידור אלקטרון מיקרוסקופ (TEM) או מיקרוסקופ כוח אטומי (AFM). כהוכחה העיקרון, 3D שחזורים של תאים בוצעו גם על AFM לבדוק z-דיוק (< 50 ננומטר) או על רשת TEM לבדוק xy-דיוק (< 30 ננומטר)3. גישה מקורלציה כזו היא זמן רב ויקר. גישה פשוטה יותר יכולה להיות דגימות סטנדרטיות של התמונה כגון תאים סוג פראי או חרוזים כדוריים 1 יקרומטר. הקוטר והספרוגניות של השחזורים יכולים לשמש כדי להבטיח שהגודל ומאזני האנרגיה המשמשים בשחזור יהיו נכונים.
זו אינה השיטה היחידה המבקשת לחלץ מידע מרחבי ברזולוציה גבוהה מתמונות מיקרוסקופ פלואורסצנטית. מאמרי סקירה רבים מתארים את ההתקדמות האחרונה בתחום המיקרוסקופיה הסופר-מיקרוסקופית24,25. טכניקות לשיפור הרזולוציה, כגון מיקרוסקופ לפירוק הדיסק26, מיקרוסקופ מיקרוסקופיה של דיסקים27, פיקסל שינוי28, ומיקרוסקופ תאורה מובנה (SIM)29 מבקשים לשפר את הרזולוציה של ה תמונות שנרכשו על ידי המיקרוסקופ. שיטות אלה אינן מתאימות לגישה המוצגת. לאחרונה שיטה זו הותאמה לאפשר תמונות מבוססות ה-SIM כתשומות9. בעוד שהשיטה הקונבולוציה קדימה חולקת חלק מהתפוקה שלה עם מיקרוסקופ פירוק, יש לו פלט שונה לחלוטין. בעוד שגישות כגון המיקרוסקופיה מנסות לשפר את הרזולוציה של הדימוי, גישה זו אינה יוצרת תמונה אלא שחזור של צורה תא עם דיוק של 50 ננומטר בקירוב. מולקולה בעלת שליטה מיקרוסקופית הפעילה על בסיס דגימות בדלילות, יכולה לספק רמות גבוהות אף יותר של דיוק מרחבי משיטה זו. במקרים רבים, אלה מולקולה בודדת גישות דורשים אופטימיזציה של מבנים פלורסנט והוא יכול לדרוש זמני רכישה ארוכים, מה שהופך אותם קשה לשימוש עם דגימות חי או דינמי. כל אחת מהשיטות הללו מגיעה עם אחת או יותר מהאזהרות ששיטה זו אינה. למשל, את היתרונות שפורסם על ידי ספינינג מיקרוסקופ הדיסק המכונה אינם ישימים monolayers של תאים חיידקיים, שם אין הרבה מחוץ לאור המטוס. יתר על כן, שיטה זו מספקת צינור לרכוש צורות תא מדויק 3D ולוקליזציה חלבונים ללא צורך של כל fluorophores מיוחדות. שיטה זו יש דרישות חומרה מינימליות (כלומר, z piezo, גבוהה NA מטרה) והוא דורש רק עשרות תמונות לכל נקודת זמן, ומאפשר לאחד בקלות לחקור מבנים תלת-ממד דינמי6.
יש כבר מספר גדל והולך של גישות ללמוד את הארגון של תאים חיידקיים במבנים תלת-ממד. אלה כוללים גישות מושגית דומה לזה כי לנצל איכות גבוהה, 3d תמונות פלואורסצנטית30,31,32. גישה זו מחייבת תאים מבודדים היטב ואינה מהווה הנחות פריורי בנוגע לגיאומטריה תאית. עם זאת, כדי לעבור לתוך אגרגטים סלולריים צפוף או ביואילאמים, התאים נחשבים כמו מוט. תצוגה זו ברזולוציה נמוכה עדיין מאפשר לבדוק את סידורי האריזה של התאים, למרות הצפיפות הגבוהה של תאים ב-ביוilm מונע ניתוח של הלוקליזציה subcellular של גורמים ספציפיים.
בעתיד, זה עשוי להיות מעניין לפתח מסגרת לשלב את המולקולה היחידה גישות השדה הרחב עם טכניקה זו שחזור תלת-ממד. יתר על כן, ייתכן שניתן לכלול גישה זו קדימה קונבולוציה עם מחשב פילוח כלי הראייה32 כדי לאפשר שחזורים של אשכולות תא צפוף יותר.
מדוע תאים התפתחו לצורות מסוימות היא בעיה מורכבת שחייבת לשקף את הסביבה המורכבת שבה הם חיים. הבנת האבולוציה והתפקוד של צורות התא מחייבת לקחת מדידות מדויקות ומדויקות של צורות אלה, שהיא מה ששיטה זו מספקת.
Disclosures
. למחברים אין מה לגלות
Acknowledgements
אנחנו רוצים להודות לג נגוין וליהושע שווביץ על שעזרו לפתח את השיטה.
מימון: RMM-NIH F32 GM103290-01A1, BPB-גלן מרכזים לחקר הזדקנות הקרן הלאומית למדע PHY-1734030.
Materials
| Name | Company | Catalog Number | Comments |
| 50 nm fluorescent beads | Invitrogen | F8795 | these are used to measure the blurring function of the microscope |
| Agarose | sigma-Aldrich | A9539 | |
| Cotton Swab | Puritan Medical Products Company LLC | S304659 | used to appy VaLaP |
| Cover Slips | VWR | 16004-302 | |
| Fiji | ImageJ | https://fiji.sc | used to cro cells |
| FM4-64 | Invitrogen | LST3166 | membrane dye used to stain cells |
| Huygens Software | Scientific Volume Imaging | Huygens essential or professional | Use to measure blurring function of microscope |
| Lanolin | Sigma-Aldrich | L7387 | combine with paraffin and petroleum jelly to make VaLaP |
| LB growth medium | BD Difco | DF0446173 | |
| M63 medium | US Biological | M1015 | |
| MATLAB | Mathworks | Needed to run forward convolution scripts | |
| Microscope Slides | Fisher | 12-550-133 | |
| NIS Elements | Nkon | ||
| Paraffin | Sigma-Aldrich | 327212 | |
| Petroleum Jelly | Equate | 49035-038-27 | |
| Piezo z stage | Nikon | 77011589 | |
| Pipet tips -p200 | USA Scientific | 1111-0730 | |
| Pipet tips- p10 | USA Scientific | 1111-3730 |
References
- Young, K. D. The Selective Value of Bacterial Shape. Microbiology and Molecular Biology Reviews. 70 (3), 660-703 (2006).
- Persat, A., Stone, H. A., Gitai, Z. The Curved Shape of Caulobacter crescentus Enhances Surface Colonization in Flow. Nature Communications. 5, 3824 (2014).
- Nguyen, J. P., Bratton, B. P., Shaevitz, J. W., Hong, H. J. . Bacterial Cell Wall Homeostasis: Methods and Protocols. , 227-245 (2016).
- Bratton, B. P., Shaevitz, J. W., Gitai, Z., Morgenstein, R. M. MreB Polymers and Curvature Localization are Enhanced by RodZ and Predict E. coli's Cylindrical Uniformity. Nature Communications. 9 (1), 2797 (2018).
- Ouzounov, N., et al. MreB Orientation Correlates with Cell Diameter in Escherichia coli. Biophysical Journal. 111 (5), 1035-1043 (2016).
- Morgenstein, R. M., et al. RodZ links MreB to Cell Wall Synthesis to Mediate MreB rotation and Robust Morphogenesis. Proceedings of the National Academy of Sciences. , 12510-12515 (2015).
- Ursell, T. S., et al. Rod-Like Bacterial Shape is Maintained by Feedback Between Cell Curvature and Cytoskeletal Localization. Proceeding of the National Academy of Science U.S.A. 111 (11), E1025-E1034 (2014).
- Bartlett, T. M., et al. A Periplasmic Polymer Curves Vibrio cholerae and Promotes Pathogenesis. Cell. 168 (1-2), 172-185 (2017).
- Taylor, J. A., et al. Distinct Cytoskeletal Proteins Define Zones of Enhanced Cell Wall Synthesis in Helicobacter pylori. bioRxiv. , 545517 (2019).
- Egan, A. J. F., Vollmer, W. The Physiology of Bacterial Cell Division. Annals of the New York Academy of Science. 1277 (1), 8-28 (2013).
- Adams, D. W., Errington, J. Bacterial Cell Division: Assembly, Maintenance and Disassembly of the Z ring. Nature Reviews Microbiology. 7 (9), 642-653 (2009).
- Werner, J. N., Gitai, Z., Melvin, I. S., Brian, R. C., Alexandrine, C. High-Throughput Screening of Bacterial Protein Localization. Methods in Enzymology. 471, 185-204 (2010).
- de Pedro, M. A., Quintela, J. C., Höltje, J. V., Schwarz, H. Murein Segregation in Escherichia coli. Journal of Bacteriology. 179 (9), 2823-2834 (1997).
- Hussain, S., et al. MreB Filaments Align Along Greatest Principal Membrane Curvature to Orient Cell Wall Synthesis. eLife. 7, e32471 (2018).
- Wong, F., et al. Mechanical Strain Sensing Implicated in Cell Shape Recovery in Escherichia coli. Nature Microbiology. 2, 17115 (2017).
- Wong, F., Garner, E. C., Amir, A. Mechanics and Dynamics of Translocating MreB Filaments on Curved Membranes. eLife. 8, e40472 (2019).
- Quint, D. A., Gopinathan, A., Grason, G. M. Shape Selection of Surface-Bound Helical Filaments: Biopolymers on Curved Membranes. Biophysical Journal. 111 (7), 1575-1585 (2016).
- Colavin, A., Shi, H., Huang, K. C. RodZ Modulates Geometric Localization of the Bacterial Actin MreB to Regulate Cell Shape. Nature Communications. 9 (1), 1280 (2018).
- Shannon, C. E. Communication in the Presence of Noise. Proceedings of the IRE. 37 (1), 10-21 (1949).
- Billings, G., et al. De Novo Morphogenesis in L-Forms Via Geometric Control of Cell Growth. Mol Microbiol. 93 (5), 883-896 (2014).
- . Measuring a Point Spread Function Available from: https://www.ibiology.org/talks/measuring-a-point-spread-function (2012)
- Bratton, B. P., Shaevitz, J. W. Simple Experimental Methods for Determining the Apparent Focal Shift in a Microscope System. PLOS ONE. 10 (8), e0134616 (2015).
- Moerner, W. E., Shechtman, Y., Wang, Q. Single-molecule Spectroscopy and Imaging over the decades. Faraday Discussions. 184, 9-36 (2015).
- Sahl, S. J., Hell, S. W., Jakobs, S. Fluorescence Nanoscopy in Cell Biology. Nature Reviews Molecular Cell Biology. 18, 685 (2017).
- Sarder, P., Nehorai, A. Deconvolution Methods for 3-D Fluorescence Microscopy Images. IEEE Signal Processing Magazine. 23 (3), 32-45 (2006).
- Oreopoulos, J., Berman, R., Browne, M., Waters, J. C., Wittman, T. . Methods in Cell Biology. 123, 153-175 (2014).
- Huff, J. The Airyscan detector from ZEISS: confocal imaging with improved signal-to-noise ratio and super-resolution. Nature Methods. 12, 1205 (2015).
- Gustafsson, M. G. L. Surpassing the Lateral Resolution Limit by a Factor of Two Using Structured Illumination Microscopy. Journal of Microscopy. 198 (2), 82-87 (2000).
- Lee, M. K., Rai, P., Williams, J., Twieg, R. J., Moerner, W. E. Small-Molecule Labeling of Live Cell Surfaces for Three-Dimensional Super-Resolution Microscopy. Journal of the American Chemical Society. 136 (40), 14003-14006 (2014).
- Yan, J., Sharo, A. G., Stone, H. A., Wingreen, N. S., Bassler, B. L. Vibrio cholerae Biofilm Growth Program and Architecture Revealed by Single-Cell Live Imaging. Proceedings of the National Academy of Sciences. 113 (36), E5337-E5343 (2016).
- Wang, J., et al. Bact-3D: A level set segmentation approach for dense multi-layered 3D bacterial biofilms. 2017 IEEE International Conference on Image Processing (ICIP). , 330-334 (2017).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved