
Method Article
学習と記憶のテストのための低コストの匂い-報酬関連課題
要約
匂い報酬の連想学習課題を使用して、生理学的操作が長期記憶と短期記憶に及ぼす異なる影響を調査しました。
要約
欲望的で連想的な記憶の堅牢で単純な行動パラダイムは、記憶の細胞および分子メカニズムに関心のある研究者にとって非常に重要です。この論文では、生理学的操作(薬理学的物質の注入など)が匂い報酬記憶の学習速度と持続時間に及ぼす影響を調べるための効果的で低コストのマウス行動プロトコルについて説明します。代表的な結果は、短期記憶(STM)と長期記憶(LTM)におけるチロシンキナーゼ受容体活性の異なる役割を調べた研究から得られたものです。雄マウスは、報酬(砂糖ペレット)を2つの匂いのうちの1つに関連付けるように条件付けされ、その関連性に対する彼らの記憶は、2時間または48時間後にテストされました。トレーニングの直前に、チロシンキナーゼ(Trk)受容体阻害剤またはビヒクル注入が嗅球(OB)に送達されました。学習率に対する注入の影響はなかったが、OBにおけるTrk受容体の遮断はLTM(48時間)を選択的に損なうものであり、短期記憶(STM;2時間)は損なわれなかった。LTM障害は、掘削時間の長さによって測定される臭気選択性の低下に起因していました。この実験の結果の集大成は、OBにおけるTrk受容体の活性化が嗅覚記憶の定着の鍵であることを示しました。
概要
連想記憶形成のメカニズムは、これまで主に1回の試行による恐怖条件付け研究に基づいて調査されてきた。しかし、多くの日常的なタスクは、通常、より複雑な取得パターンを持ち、繰り返し遭遇することに依存しています。このプロトコルの目標は、マルチトライアルの食欲学習と記憶の細胞および分子メカニズムを理解するために使用される、費用対効果の高いげっ歯類の行動パラダイムを提供することです。
主嗅球(OB)に依存する匂い学習は、複数回の食欲記憶の研究にいくつかの利点を提供します。まず、OB依存性記憶は持続時間(STM、LTM、中期記憶1)にばらつきがあり、神経調節4、長期増強5、成体神経新生6,7,8など、脳の他の場所と同じ分子2,3と構造メカニズムに依存しています.第二に、海馬のような高次領域とは対照的に、OB依存性記憶は、実験者の知覚環境の操作と学習に関与する神経回路の変化との間のより直接的な対応関係の観察を可能にする8,9,10,11 .この論文では、一般的な分子的および構造的メカニズムを研究するために使用できる、OB依存の連想学習および記憶パラダイムについて詳しく説明します。これは、研究者が記憶の細胞および分子メカニズムの研究のために嗅覚学習の利点にアクセスできるようにするために開発されました。
私たちの最近の出版物3では、ここに記載されているプロトコルを使用して、食欲の匂い学習の統合がOB内のTrk受容体の活性化に依存していることを実証しました。以下のプロトコルでは、さまざまな実験ニーズに合わせて行動パラダイムを調整できる領域についても説明します。
この研究では、カニューレ挿入時に8週齢の成人男性CD-1マウス合計87匹が使用されました。正確なグループ分布と臭気セットの使用については、以前の出版物3の方法のセクションを参照してください。エストロゲンレベルの大きな変動を避けるために雄マウスを使用したのは、以前の研究12 が嗅覚記憶がエストロゲンレベルの増加によって強化されることを示していたからである。これらのマウスは、常に12:12の逆明暗サイクルに保たれ、水にアクセスできました。行動実験中、マウスの食事は、マウスの自由摂食体重の~90%に維持するように制限されました。食事制限は、行動実験開始の3日前から始まりました。以下で説明するように、動物の使用を最小限に抑えながら適切なレベルの統計的検出力に到達するために、同じマウスのセットに異なる匂いセットが提示されます。統計分析のセクションでは、これによって生じる可能性のあるランダム分散を考慮する方法を示します。
プロトコル
以下のプロトコルは、アーラム大学のIACUCの動物ケアガイドラインに準拠しています。
1.嗅球カニューレ挿入
注:これらの手術は、大きな切開を行う必要がないため、滅菌技術を必要としません。ただし、各機関の要件は異なる場合があります。実験者が免疫不全マウス株でこの手術を行う場合、追加の考慮事項が必要になる場合があります。全体として、実験者は、使用前に獣医師や動物ケアチームとこのプロトコルについて話し合い、各手術の間にすべてのツールを洗浄および消毒することをお勧めします。
- セットアップ中に、カニューレとネジを32%クロルヘキシジンを含む小さなビーカーまたはシャーレに浸して消毒します。
- マウスをガス状の4%イソフルランで純酸素中で麻酔し、定位固定装置に固定します。手術中は、マウスがノーズコーンを介して供給される1.5〜2%のイソフルラン麻酔の下に維持されていることを確認してください。.手術中、呼吸をモニターします。麻酔下での目の乾燥を防ぐために、目の軟膏を使用してください。
- マウスが固定され、後足のしっかりとつまむことに反応しなくなったら、切開面をきれいにするために、32%クロルヘキシジンを使用して頭のてっぺんをこすります。
- 次に、吻側の頭頂部をリドカイン(局所鎮痛)で尾側にこすります。
- しっかりと押し下げて、きれいなメスの刃で正中線を1回切開します。
- 定位固定装置に取り付けられたドリルを使用して、ガイドカニューレ(26 G)用の嗅球の上に2つの穴を開けます。これは、ブレグマに対してAP + 5.0 mm、ML + /- 0.75 mmの座標を使用して行われます。
- 小脳層の上に2つの穴を開けます。
注意: これらのネジの位置は正確である必要はなく、正中線に沿って対称であることを確認してください。 - 小脳形成部の上の2つの穴にネジを入れ、ティッシュ接着剤を使用してこれらのネジを頭蓋骨に固定します。
- 定位固定装置を使用して、ガイドカニューレ(26 G)を嗅球に開けられた穴に挿入します(ステップ1.6)。カニューレDVを1.0mm下げます。
- デンタルセメントをペトリ皿で混ぜます。小さな金属製のスコップを使用して、カニューレの周りに歯科用セメントをゆっくりと積み上げます。5秒間乾燥させます。次に、ガイドカニューレを引き抜かないように注意しながら、脳定位固定装置のアームを取り外します。小さなキャップが形成されるまで、切開部全体に歯科用セメントを積み続けます(図1Aを参照)。
- この時点でダミープラグをガイドカニューレに配置して、詰まりを防ぎます。
- 手術直後、ケトプロフェン(0.2mg/kg)と生理食塩水(200μL)を注射して痛みを軽減し、水分を補給します。手術後のマウスに軟化した食品またはヒドロゲルを提供します。マウスが意識を取り戻し、胸骨の横臥を維持するまでマウスを放置しないでください。
注:マウスもこの時点から単独で飼育されます。 - 生理食塩水(200μL)とケトプロフェン(0.2mg/kg)を1日1回、手術後2日間注射します。
- 手術後2日間から最大5日間(必要に応じて)、マウスの体重を量り、体重を監視します。体重が2〜3日以内に手術前のレベルに戻らない場合は、適切な給餌方法について獣医師に相談してください。
- マウスが行動訓練を開始する前に、少なくとも7日間回復するのを待ちます。
2. 連想的差別課題
- 注入
- チロシンキナーゼ受容体阻害剤であるK252a(50μM;生理食塩水中の5%DMSO)またはビヒクル(生理食塩水中の5%DMSO)のOB特異的注入をマウスに次のように投与します。
- マウスのOBに両側で注入を送達します。デュアルインジェクターポンプを使用して、0.2 μL/分の注入速度と10分間の合計注入時間で、バルブあたり2.0 μLの最終容量を注入します。
注:操作のタイミングは、実施する行動研究の種類に応じて調整できます。輸液の正確な手順は、各インジェクターポンプに固有であり、メーカーのマニュアルに記載されています。 - インジェクターは、デリバリー後約5分間はカニューレ内に置いておくことで、逆流を抑制し、拡散を促進してください。(つまり、注入ごとに合計15分を計画します)。
- マウスのOBに両側で注入を送達します。デュアルインジェクターポンプを使用して、0.2 μL/分の注入速度と10分間の合計注入時間で、バルブあたり2.0 μLの最終容量を注入します。
- チロシンキナーゼ受容体阻害剤であるK252a(50μM;生理食塩水中の5%DMSO)またはビヒクル(生理食塩水中の5%DMSO)のOB特異的注入をマウスに次のように投与します。
- 臭いがします。
- 軽質鉱物油中のすべての臭気物質を、蒸気圧に基づいて事前に計算された比率を使用して1.0Paの分圧に希釈します(表1)。
- 表1の5つの別々の臭気ペアを使用します(表の数字は、1.0Paに対して50mLの鉱油を混合するための容量をμLで示しています)。
- 行動ステップ(セクション2.3および3)で使用する香りの砂を準備するには、プレイサンド100 gごとにステップ2.2.2の1.0 Pa臭物質を400 μL混合します。
- 形成
注:マウスは、以下に説明するように10日間成形を受ける必要があります。- マウスを処置室に持ち込み、手術からの回復後最初の2日間は1日10分間マウスを取り扱います。
- 3日目に、(+)-リモネンの香りの砂で満たされたシャーレをマウスのホームケージに入れ、それぞれ5 mgの質量で約10個のスクロースペレットを充填します。
注:報い臭として1.0 Pa(+)-リモネン(50 mLの鉱油に102μLを混合)を使用し、報いのない臭気としてプレーンな鉱油(試験臭の希釈剤)を使用してください。また、単分子の匂い物質はマウスにとって新規である可能性が高いため、選択することをお勧めします。 - 4日目に砂とペレットの両方を補充します。
- 5日目と6日目に、マウスを装置に入れて空間を探索させることにより、マウスをカスタムメイドの行動装置に順応させます(図1B、C)。標準的な家庭用ケージとポリ(メタクリル酸メチル)を使用して装置を作成し、2つの蓋と黒い中央の仕切りを構成します。蓋と中央の仕切りの両方がホームケージより1〜2cm大きいことを確認してください。
- リモネンの香りの砂のシャーレと、鉱物油を含む砂のシャーレを準備します。5日目と6日目の両方に順応した後、両方の砂の皿を試験チャンバーに入れ、10個のショ糖ペレットをリモネンの香りの皿に混ぜて報酬として使用します。各マウスを試験チャンバーに10分間入れ、報酬ペレットを自由に探索して消費できるようにします。
- 7日目に、リモネンの香りと鉱物油の香りの砂が入った皿を行動装置に入れることにより、最終的なテスト手順の要約版をマウスに導入します。今回は中央の仕切りが含まれています。
- リモネンの香りの砂の上に1つの報酬を置き、マウスを休息室に置きます。
- マウスを休息チャンバーに入れたら、中央の仕切りを持ち上げて、マウスがテストチャンバーに入り、砂で満たされた皿を調査して掘ります。マウスが報酬ペレットを回収した後、または5分が経過した後、マウスを休息室に戻します。
- このプロセスを、個々のマウスごとに合計 10 回の試行を繰り返します。ランダムナンバージェネレーターを使用して、左側または右側のいずれかに報酬のある料理の配置を相殺します。トライアル間の休憩時間はありません。
- 8日目は、7日目の試行を繰り返しますが、ペレットを砂の奥深くに徐々に埋めていきます。
注:ほとんどのマウスは、8日目の10回目の 試行までに、目に見えない報酬ペレットを掘っているはずです。 - 9日目に、スクロースペレットを完全に深く埋設し、マウスを試験室に1回の試行ごとに1分間導入するマウスごとに、試行回数を20回に増やします。ご褒美のために両方の皿をネズミに掘り起こさせてください。
- 10日目に、各マウスに対して20回の試行を繰り返しますが、報酬のある皿を掘る前に報酬のない皿を掘った場合は、次の試行を開始します。最初に報酬のある(リモネンの香りのする)皿を掘ったマウスが報酬ペレットを取り戻すのを待ってから、休息室に戻します。
3. トレーニングとテスト
注:マウスが目に見えない、臭いを原因とする報酬ペレットを確実に掘り始めると、実験を開始できます。
- 訓練
注:トレーニングフェーズは、シェーピングが完了してから2日後に開始され、各マウスの20回の試行で構成されます。トレーニングの前に、 眼球内薬物/ビヒクル注入をすぐに提供し(注入の詳細についてはセクション2.1を参照)、注入直後にトレーニングを開始します。.- マウスを休息室に入れます。
- 新しい匂いペアの香りがする砂の皿を2つテストチャンバーに入れ、報酬ペレットを皿の1つに埋めます。
- 試験チャンバーの準備ができたら、不透明なバリアを持ち上げ、マウスを試験チャンバーに導入します。マウスが最初に報酬のない皿を掘った場合は、すぐにマウスを休息室に戻します(これらの試行を「0」として記録します)。ネズミが最初に報酬の臭いを掘り下げた場合は、ペレットを回収して休息室に戻します。これらの試行を「1」として記録します。マウスが報酬を取り戻さずに試用が1分間続く場合は、マウスを休息室に戻します。
- 食器をきれいにして補充し、次のトライアルを開始します。同じことを 20 回の試行で繰り返します。
- テスティング
注:記憶力テストは、研究者が関心のある任意の期間に実行できます。この実験では、K252aがSTMとLTMに及ぼす異なる影響に関心があることから、トレーニングの2時間後(STM)または48時間後(LTM)にマウスの2つの別々のグループをテストしました。- トレーニング(セクション3.1)で説明されているのと同じ臭気と手順を使用してテストを実施します。
注:研究課題によっては、研究者は対照群を含める必要がある場合があります。例えば、以前に発表された実験では、記憶の定着に対するTrk受容体遮断の効果が研究された3。したがって、48時間試験の前に対照群にK252aを注入し、その効果が回収の干渉によるものではないことを示しました。
- トレーニング(セクション3.1)で説明されているのと同じ臭気と手順を使用してテストを実施します。
- 統計分析
注:各ステップのSPSS 22.0構文は、例として 補足ファイル として提供されています。- 線形混合効果分析を使用して統計分析を実行します。分散分析(ANOVA)とは異なり、線形混合効果モデルは、変量効果と反復測定をより適切に説明できます。
- 従属メジャーを計算します: "比率が正しい"。3.1.3から、マウスが最初に報われない匂いを掘り下げた試行には「1」が割り当てられ、マウスが最初に報われない匂いを掘り下げた場合は「0」が割り当てられたことを思い出してください。4つの試行ブロックを作成するための5回の試行ごとの平均(TB;例えば、試行ブロック1またはTB1は試行1〜5の平均であり、試行ブロック2またはTB2は試行6〜10の平均であった、など)。
- 独立変数または固定効果を薬物グループ(K252aまたはVehicle;セクション 2.1.1) と試用ブロック (3.3.1 から) です。以下の代表的な結果では、各分析に用いる変数を指定しています。
- マウス内に配置された個々のマウスとマウス内にネストされた匂いセットを「ランダム効果」として含めて、マウスの本質的な行動の違いや複数の匂いセットを使用した場合の影響を補正します。
- 正しい比率でロジット変換を実行します。
注: 正しい比率は、連続的で束縛されていない従属変数ではありません。したがって、線形モデルの 2 つの仮定に違反します。したがって、ロジット変換が実行されます。 - 推定周辺平均を使用して、フルモデルによって特定された有意な交互作用に対して事後検定を実行します。多重ペアワイズ比較は、事後テストで修正する必要があります。通常、ボンフェローニまたはシダークが使用されます。
結果
説明したように、このプロトコルにより、研究者は学習、STM、およびLTMに対する何らかの操作の影響を評価できます。Tong et al, 20183 のサンプル結果をここに掲載します。この結果は、Trk受容体の遮断がLTMを選択的に阻害するが、学習やSTMは阻害しないという仮説を支持しています。
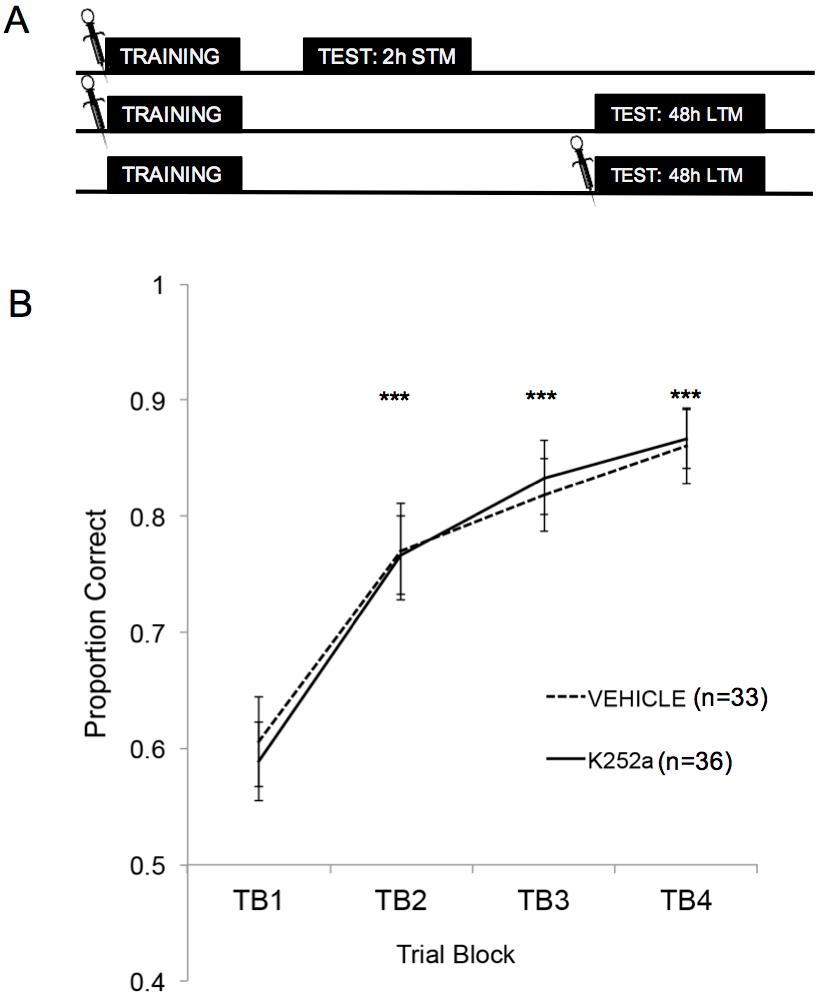
図 2A は、トレーニング、STM テスト、LTM テストの概略図を示しています。まず、K252aの注入は、匂いと報酬の関連付けの学習率に影響を与えないことが示されました。 図2B (セクション3.1)は、トレーニングからのK252aと車両グループの両方の学習率を示しています。線形混合モデルを使用した統計分析は、薬物群と試験ブロック(TB)の2つの固定効果で実行されました。マウスとマウス内にネストされた匂いセットはランダムな効果でした。テスト(セクション3.2)からのデータは分析に含まれていません。試験ブロックから有意な主効果が認められました(F(3, 183.692) = 43.735, p < 0.001)が、薬剤群からの効果は見られませんでした(F(1, 85.685) = 0.132, p = 0.717),有意な交互作用は認められませんでした(F(3, 183.692) = 0.111, p = 0.954)。Šidák調整を使用した事後テストでは、訓練中のどの試験ブロックでもK252aと車両グループに差がないことが確認されました(すべての比較でp > 0.05)。TB2、TB3、TB4は、すべての比較でTB1よりも有意に高かった(すべてのケースでp ≤ 0.001)ことから、両グループが20回のトレーニング試行の終わりまでに臭気と報酬の関連性を首尾よく学習したことが示されました。
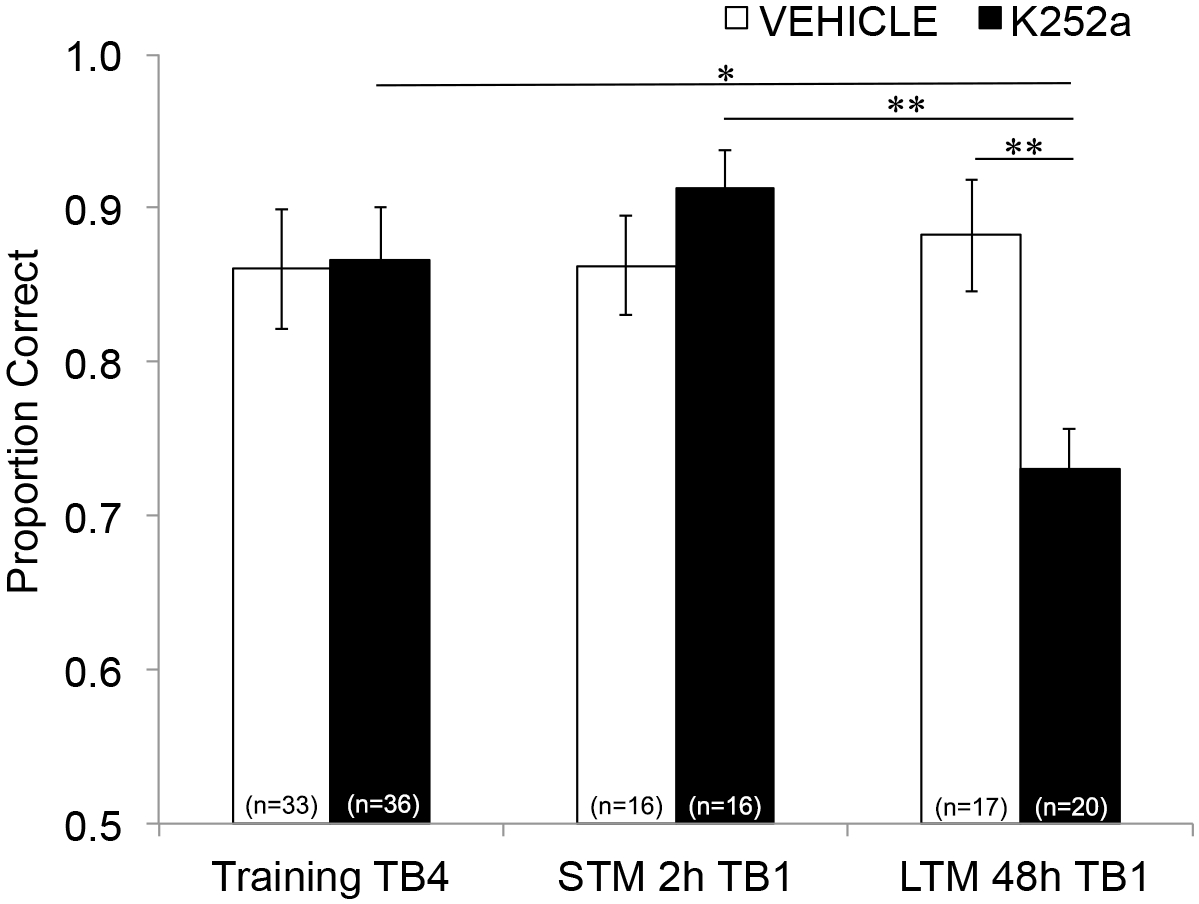
次に、注入がSTMおよびLTMに及ぼす影響を調べるために、同じ分析を実行し、テスト(セクション3.2)からのデータを含めました。薬剤群と試験ブロックとの間に有意な相互作用(F(2, 77.558) = 4.043, p = 0.021)が示され、薬剤群(F(1, 55.629) = 1.438, p = 0.236)または試験ブロック(F(2, 69.979) = 1.360, p = 0.263)の有意な主効果は認められませんでした。記憶を具体的に調べるために、Šidák補正との事後ペアワイズ比較では、トレーニングの最後の試行ブロック(セクション3.1)とテストの最初の試行ブロック(セクション3.2)の2時間後(STM)または48時間後(LTM)の違いを比較しました。ビヒクル注入マウスの場合、比較では、トレーニング後2時間と48時間の両方で連想記憶の保持が示されました(トレーニングパフォーマンスとのすべての比較でp > 0.05)。K252aを注入したマウスでは、2時間試験(STM)の最初の試行ブロックは、学習の最後の試行ブロックと異ならなかった(p > 0.05)。しかし、48時間後にはメモリ性能が有意に低下しました(P = 0.018)。また、48時間テストのメモリは、2時間テストのメモリ(p = 0.009)や48時間テストの車両群の性能(p = 0.006)と比較して大幅に減少しました。ビヒクルマウスとK252aを注入したマウスとの間にSTMに差はなかった(p = 0.356)。以上の結果から、K252aが嗅球におけるTrk受容体を阻害すると、匂い記憶は短期的ではなく、長期的に選択的に阻害されることが示されました(図3)。
| ニオイセット | 臭気1 | 臭気2 |
| 1 | ペンタン酸 | ブタン酸 |
| 225.1 | 63.6 | |
| 2 | ヘキサナル | ヘプタナール |
| 11.1 | 35.3 | |
| 3 | 酢酸プロピル | 酢酸ブチル |
| 3.1 | 10.9 | |
| 4 | 2-オクタノン | 2-ヘプタノン |
| 87.4 | 28.7 | |
| 5 | ペンタノール | ヘキサノール |
| 37.2 | 127.3 |
表1:臭気セットの混合量。 各行には、行動ステップのペアとして使用できる 2 つの臭気が表示されます。たとえば、最初の「臭気セット」を使用するには、ペンタン酸とブタン酸の混合物を作成します。表の数字は、各臭気の濃度が1.0Paの場合、50mLの鉱油を混合する容量をμLで示しています。トレーニングとテスト中、1つのシャーレにはペンタン酸の香りが、もう1つのシャーレにはブタン酸の香りがします。

図1:カニューレ留置と行動装置。 (A)カニューレ、歯科用セメントキャップ、およびネジとマウスの頭部との相対位置を示しています。カニューレの針は2つの嗅球に到達し、台座自体が歯科用セメントキャップに埋め込まれていることに注意してください。ネジは、小脳の形成上の頭蓋骨に開けられた2つの穴に配置されます。ネジは脳自体には触れませんが、歯科用セメントキャップの尾側アンカーとして機能します。この図は、セメントキャップを作るための相対的なサイズを示しています。(B)組み立てられた行動装置を示す。本体は典型的なマウスのホームケージです。空気用の穴が開いた蓋はプレキシガラスで作られました。中央の仕切りも黒のプレキシガラスで作られています。蓋は、試験中に中央の仕切りを持ち上げて配置するためのトラクトとして機能することができるように、十分に重いプレキシガラスで作られている必要があります。(C)行動装置の一面を示す。中央の仕切りは、持ち上げやすいようにケージよりもわずかに高いことに注意してください。砂のシャーレを端に沿って配置できます。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図2:学習デザインと学習結果。 (A)試験デザインの概略図を示しています。STMグループとLTMグループは独立している(つまり、マウスのグループが異なる)ことに注意してください。ブロックの先頭にある記号は、注入が行われた時間を示しています。(B) トレーニング中の試練1-20の正しい比率を示しています。その結果、K252aと車両群の学習率(線の傾き)に差はなかった。エラーバーは、平均の標準誤差(SEM)を表しました。アスタリスクは、ビヒクルとK252aの両方のコホートでTB1と比較して、正しい比例の有意な増加を示しています(すべての比較でp ≤0.001)。この図は、Tong et al. 2018 の許可を得て3 から引用したものです。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図3:STMとLTMに対するTrk受容体遮断の異なる効果。 トレーニングの試行ブロック 4 と、STM テストと LTM テストの両方の試行ブロック 1 の正しい比率を示します。つまり、線形混合モデルを使用して、トレーニング フェーズの最後の試行ブロックで正しい比率を比較します (図 2;Training-TB4)を、短期(2時間テスト)および長期(48時間テスト)記憶テストの最初の試行ブロック(Testing-TB1)中のものに適用します。線形混合モデルには、薬物群と試験ブロック (Training-TB4、STM-TB1、LTM-TB1) の 2 つの固定効果がありました。ランダム効果は、マウスとマウス内にネストされた臭いセットでした。事後比較の結果、K252aマウスはLTMが有意に損なわれていることがわかりました(トレーニングTB4との比較; p = 0.018) ですが、STM (p > 0.05) は除くことができます。また、K252aを注入したマウスのLTM性能は、K252aを注入したマウスのSTMよりも有意に低く(p = 0.009)、ビヒクルマウスのLTM(p = 0.006)よりも低かった。エラーバーはSEMを表していました。この図は、Tong et al. 2018 の許可を得て3 から引用したものです。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

補足ファイル: 統計分析に使用される構文。 このファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
1回の試行による恐怖条件付けは、記憶の分子および細胞ダイナミクスを研究するための強力な行動プロトコルですが、自然学習の多くは漸進的であり、上記のようなパラダイムを通じてモデル化するのが最適です。OBにおけるTrk受容体の阻害は、以前に私たちのグループ3で示されたように、多試行の食欲学習パラダイムにおける嗅覚記憶の統合を妨げました。この知見は、ニューロトロフィンのような分子メカニズムの異なるタイミングを、食欲学習と嫌悪学習における研究に新たな道を開くものです。
この実験は、(1)カニューレ挿入と(2)連想識別タスク(シェーピング、トレーニング、テストに細分化)の2つの重要な部分で構成されていました。実験者は、このプロトコルを自分の特定の研究課題に適合させることができます。例えば、私たちは主に産婦人科に関心を持っていましたが、この確立されたプロトコルは他の産科研究にも簡単に適用できます。他の関心領域を持つ実験者にとって、パイロット研究で注入部位を検証することが重要になります。実験者は、注入するものの拡散率、空間浸透、および生物活性持続時間も考慮する必要があるかもしれません。
このプロトコルで概説されている整形手順は、このプロトコルの著者によって広く使用されてきました。説明されているようにそれらに従うことは、マウスにタイムリーにタスクを学習させるために重要であるように思われます。他のタイムラインを使用すると、著者たちは、この課題に対するマウスの習熟度により多くのばらつきがあることを観察し、これは、実験的な匂いでテストするための基準にすべてのマウスを到達させるための追加のトレーニングを意味しました。トレーニングとテストについては、研究者は、研究の関心に応じて、試行回数、実験臭気の濃度、および臭気の相互の類似性に柔軟性があります。実験に使用する動物の数を減らすために、説明したように、可能な限り複数の臭気セットを使用することをお勧めします。最終的な分析で複数の注文セットの使用を考慮する方法のガイダンスについては、「統計分析」セクションを参照してください。原則として、臭気ペアの類似性を変化させて、識別の難易度を調整することができます。私たちのグループ3から以前に発表された研究では、臭気ペアは、同じ官能基の2つの匂い物質で構成されていますが、互いに炭素の長さが1つ異なります。これらの識別は、炭素長が2つ以上異なるペアよりも困難ですが、エナンチオマー((+)-リモネンや(-)-リモネンなど)よりも簡単です。異なる官能基からの匂い物質は、知覚的に大きく異なります。Clelandら13 は、より多くの刺激のバリエーションと、それらが特定の学習パラメータに及ぼす影響について論じています。
このプロトコルの大きな制限の1つは、複数の動物を並行してテストできる自動連想学習タスクと比較して、実行にはるかに長い時間がかかることです。ある試験では、1人の研究者が1匹のマウスに対して20回の試行を完了するのに少なくとも20分かかります。しかし、この自動化の欠如は、プロトコルが財政的にアクセスしやすくなることを意味し、多くの機関にとって優先事項となっています。重要なことに、この実験の場合、このプロトコルは、行動神経科学に関心のある学部生の研究者を訓練するために非常に扱いやすく、効果的であることがわかりました。特に、これらの学生は、研究参加の通常の利点に加えて、強力な動物取り扱いスキルを開発します。
このパラダイムの採用に関心のある研究者は、いくつかのパラメーターを変えることができます。最も明らかに、メカニズムの薬理学的操作は多様であり、この行動プロトコルは、化学遺伝学的手法または分子および細胞経路を操作するさまざまな他の方法(例えば、光遺伝学)とともに使用することができる。パラダイム自体は、テストされる学習と記憶の種類を変えるように調整できます。例えば、研究者は学習率を制御するために、提示される2つの匂いの類似性を調整することができます。私たちの研究3では、臭気ペアは、同じ官能基の2つの匂い物質で構成されていますが、互いに炭素の長さが1つ異なります。これらの識別は、炭素長が2つ以上異なるペアよりも困難ですが、エナンチオマー(例:(+)-リモネンと(-)-リモネン)よりも簡単です。異なる官能基からの匂い物質は、知覚的に大きく異なります。Clelandら13は、より多くの刺激変動とそれらが特定の学習パラメータに及ぼす影響について議論し、類似した匂いが多いほど識別が難しく、したがって学習に時間がかかると結論付けています14。これらの操作は、記憶の強さにも影響を与えます。この流れに沿って、研究者は学習後のさまざまな時点で記憶をテストすることに興味があるかもしれません。例えば、2つの研究15,16では、1回の試行による嫌悪学習課題のLTM持続性におけるBDNFの役割を調べた。BDNFはTrkBのリガンドです。これらの研究では、学習の12時間後に海馬に抗BDNFアンチセンスオリゴヌクレオチドを注入すると、7日後にLTMがブロックされましたが、2日後にはブロックされませんでした。この研究は、初期学習後の分子メカニズムの時間スケールが、LTMにおいて興味深い役割を果たしているが、まだ理解されていないことを示しています。この論文では、これらのタイムスケールの調査を可能にする行動プロトコルについて説明します。将来のアプリケーションとして関心のある他のパラメータには、使用されるマウスモデルが含まれます。例えば、将来の研究では、雄のマウス(神経生物学の研究が著しく優れている)17を雌マウスに置き換えて、学習率、STM、およびLTMの変動を調べることは興味深いでしょう。なぜなら、雌の哺乳類は雄の哺乳類18よりも匂いに対する感度と選択性が高いからです。もちろん、このプロトコルでは、病気のげっ歯類モデルも効果的に使用できます。
開示事項
この論文の著者は、競合する金銭的利益を持っていませんでした。
謝辞
このプロジェクトは、Scantland Summer Collaborative Research GiftとStephen and Sylvia Tregidga Burges Endowed Research Fundの支援を受けました。著者らは、アーラム大学の心理学科と神経科学プログラムの同僚の支援と指導に感謝したいと思います。
資料
| Name | Company | Catalog Number | Comments |
| Double guide cannula | PlasticsOne | C235GS-5-1.5/SPC | Custom order |
| (-)-limonene | Sigma-Aldrich | 218367-50G | |
| (+)-limonene | Sigma-Aldrich | 183164-100ML | |
| 2-hetanone | Sigma-Aldrich | 537683 | |
| 2-octanone | Sigma-Aldrich | O4709 | |
| 5mg sucrose pellets | Test Diet | 1811560 | Custom size. Used for rewards |
| Butanoic acid | Sigma-Aldrich | B103500 | |
| butyl acetate | Sigma-Aldrich | 402842 | |
| Dental Cement Powder (Coral) | A-M Systems | 525000 | |
| Dental Cement Solvent | A-M Systems | 526000 | |
| Double connector assembly | PlasticsOne | C232C | |
| Double dummy cannula | PlasticsOne | C235DCS-5/SPC dummy dbl | Custom order |
| Double injector | PlasticsOne | C235IS-5/SPC | Custom order |
| Drill | Kopf Instruments | Model 1474 High Speed Stereotaxic Drill | This drill requires an additional "adaptor" piece in order to fit certain drill bits. We get by this problem by wrapping the drill bit with lab tape to increase the circumference of the drill it to fit. This may not be an option for surgeries requiring sterile technique. |
| Eye Ointment | Purchase from local pharmacy | ||
| Figure 1 illustration software | BioRender | ||
| heptanal | Sigma-Aldrich | W254002 | |
| hexanal | Sigma-Aldrich | 115606 | |
| hexanol | Sigma-Aldrich | H13303 | |
| Infusion pump model 11 | Harvard Apparatus | 4169D | Used pumps available via American Instrument Exchange |
| Isoflurane | Santa Cruz Animal Health | sc-363629Rx | Vet prescription needed for order |
| K252a | Sigma-Aldrich | K2015 | Mixed to 50uM in DMSO (5%) |
| Ketoprofen | Allivet | 25920 | Vet prescription needed for order |
| Lidocaine | Aspercreme | Purchased from Amazon | |
| Mounting Screws | PlasticsOne | 00-96 X 3/32 | |
| Mouse Anesthesia Mask | Kopf Instruments | Model 907 Mouse Anesthesia Mask | Used with the stereotaxic to allow oxygen and anesthesia while mouse in stereotax |
| Mouse Nose Adaptor | Kopf Instruments | Model 926 Mouse Adaptor | Used with the stereotaxic to allow for head of mouse to be secured. |
| Novalsan | Jeffers | 41375 | |
| Pentanoic acid | Sigma-Aldrich | 240370 | |
| pentanol | Sigma-Aldrich | 138975 | |
| Petri dish glass bottoms | VWR | 10754-804 | |
| Polycarbonate Café bottoms | Ancare | N10PCSEC | Use normal housing cages and custom fit a track in the middle to act as the track for an opaque plexiglass divider |
| propyl acetate | Sigma-Aldrich | 537438 | |
| Quikrete Premium Play Sand | Purchase from local hardware store | ||
| Saline | Insight Needles | N/A | Sterile saline for drug mixing |
| Stereotaxic apparatus | Kopf Instruments | Model 902 Small Animal Stereotaxic Instrument | |
| Testing chamber | Ancare | N10PCSEC | Our testing chambers are modified using the regular mouse housing cage. The manuscript details what was done. |
| Vetbond Tissue Adhesive | 3M | Purchased from Amazon |
参考文献
- Grimes, M. T., Smith, M., Li, X., Darby-King, A., Harley, C. W., Mclean, J. H. Mammalian Intermediate-Term Memory: New Findings in Neonate Rat. Neurobiology of Learning and Memory. 95 (3), 385-391 (2011).
- Grimes, M. T., Harley, C. W., Darby-King, A., Mclean, J. H. PKA Increases in the Olfactory Bulb Act as Unconditioned Stimuli and Provide Evidence for Parallel Memory Systems: Pairing Odor with Increased PKA Creates Intermediate- and Long-Term, but not Short-Term, Memories. Learning and Memory. 19 (3), 107-115 (2012).
- Tong, M. T., Kim, T. Y. P., Cleland, T. A. Kinase Activity in the Olfactory Bulb is Required for Odor Memory Consolidation. Learning & Memory. 25 (5), 198-205 (2018).
- Devore, S., Lee, J., Linster, C. Odor Preferences Shape Discrimination Learning in Rats. Behavioral Neuroscience. 127 (4), 498-504 (2013).
- Gao, Y., Strowbridge, B. W. Long-Term Plasticity of Excitatory Inputs to Granule Cells in the Rat Olfactory Bulb. Nature Neuroscience. 12 (6), 731-733 (2009).
- Bath, K. G., Mandairon, N., et al. Variant Brain-Derived Neurotrophic Factor (Val66Met) Alters Adult Olfactory Bulb Neurogenesis and Spontaneous Olfactory Discrimination. Journal of Neuroscience. 28 (10), 2383-2393 (2008).
- Lazarini, F., Lledo, P. M. Is Adult Neurogenesis Essential for Olfaction. Trends in Neurosciences. 34 (1), 20-30 (2011).
- Mandairon, N., Peace, S., Karnow, A., Kim, J., Ennis, M., Linster, C. Noradrenergic Modulation in the Olfactory Bulb Influences Spontaneous and Reward-Motivated Discrimination, but not the Formation of Habituation Memory. European Journal of Neuroscience. 27 (5), 1210-1219 (2008).
- Guérin, D., Peace, S. T., Didier, A., Linster, C., Cleland, T. A. Noradrenergic Neuromodulation in the Olfactory Bulb Modulates Odor Habituation and Spontaneous Discrimination. Behavioral Neuroscience. 122 (4), 816-826 (2008).
- Moreno, M. M., Bath, K., Kuczewski, N., Sacquet, J., Didier, A., Mandairon, N. Action of the Noradrenergic System on Adult-Born Cells Is Required for Olfactory Learning in Mice. Journal of Neuroscience. 32 (11), 3748-3758 (2012).
- Vinera, J., Kermen, F., Sacquet, J., Didier, A., Mandairon, N., Richard, M. Olfactory Perceptual Learning Requires Action of Noradrenaline in the Olfactory Bulb: Comparison with Olfactory Associative Learning. Learning and Memory. 22 (3), 192-196 (2015).
- Dillon, T. S., Fox, L. C., Han, C., Linster, C. 17β-estradiol Enhances Memory Duration in the Main Olfactory Bulb in CD-1 Mice. Behavioral Neuroscience. 127 (6), 923 (2013).
- Cleland, T. A., Narla, V. A., Boudadi, K. Multiple Learning Parameters Differentially Regulate Olfactory Generalization. Behavioral Neuroscience. 123 (1), 26 (2009).
- Cleland, T. A., Morse, A., Yue, E. L., Linster, C. Behavioral Models of Odor Similarity. Behavioral Neuroscience. 116 (2), 222-231 (2002).
- Bekinschtein, P., Cammarota, M., Igaz, L., Bevilaqua, L., Izquierdo, I., Medina, J. Persistence of Long-Term Memory Storage Requires a Late Protein Synthesis- and BDNF- Dependent Phase in the Hippocampus. Neuron. 53 (2), 261-277 (2007).
- Bekinschtein, P., et al. BDNF is Essential to Promote Persistence of Long-Term Memory Storage. Proceedings of the National Academy of Sciences. 105 (7), 2711-2716 (2008).
- Andreano, J., Cahill, L. Sex influences on the neurobiology of learning and memory. Learning and Memory. 16 (2009), 248-266 (2009).
- Kass, M. D., Czarnecki, L. A., Moberly, A. H., Mcgann, J. P. Differences in Peripheral Sensory Input to the Olfactory Bulb Between Male and Female Mice. Scientific Reports. 7 (1), (2017).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved