
Method Article
非トランスジェニック無脊椎動物における単一ニューロン分解能を用いたネットワークダイナミクスを記録するためのフォトダイオードベースの光学イメージング
要約
このプロトコルは、吸光度の電圧依存性色素とフォトダイオードアレイを用いた非トランスジェニック無脊椎動物種における単細胞分解能を用いたニューロン集団活性をイメージングする方法を提示する。このアプローチにより、迅速なワークフローが可能になり、イメージングと解析を1日にわたって追求できます。
要約
特定可能なニューロンセットの活性を光で記録し、操作することができるトランスジェニック無脊椎動物製剤の開発は、行動の神経基盤の研究のための革命的な進歩を表しています。しかし、この開発の欠点は、研究者が非常に少数の「デザイナー」生物(例えば 、C.エレガンス および ショウジョウバエ)に焦点を当てる傾向にあり、ネットワーク機能の一般原理を特定するために必要な多くの種にわたる比較研究の追求に悪影響を及ぼす可能性がある。本稿は、非トランスジェニック胃ポッド種の脳における電圧感受性色素による光記録が、単一細胞分解能を有するニューラルネットワークの機能的組織の特徴を迅速に(すなわち、単一の実験の時間内に)いかにして明らかにすることができるかを示している。我々は、神経科学に新しい1つを含む複数の胃ポッド種のCNSにおける行動関連の運動プログラム中に数十〜150ニューロンから行動電位痕を得るために我々の研究室で使用される解剖、染色、および記録方法を詳細に概説する - nudibranch Berghia stephanieaee.イメージングは、吸光度の電圧感受性色素と、記録されたニューロンによって生成されたすべてのアクション電位をキャプチャするのに十分な速さで、1,600フレーム/秒でサンプリングする464要素フォトダイオードアレイで行われます。複数の数分間の記録は、シグナルの漂白や光毒性をほとんどまたは全くなく、調製ごとに得ることができます。記載された方法を通して収集された生の光学データは、さまざまな例示された方法を通して分析することができる。私たちの光学記録アプローチは、様々な非トランスジェニック種のネットワーク活動を容易にプローブするために使用することができ、脳がどのように行動を生み出すのかの比較研究に適しています。
概要
ショウジョウバエやC.エレガンスなどの無脊椎動物のトランスジェニックラインの開発は、行動の神経塩基が光学的に尋問され、操作することができる強力なシステムを提供してきました。しかし、これらの特殊な製剤は、特に神経科学研究への新種の導入に関して、非トランスジェニック種の神経回路研究に対する熱意を低下させるという欠点を有し得る。比較研究は、そのような原理が発見される不可欠な経路を表すため、1つまたは2つのモデルシステムのみに焦点を当てることは、ネットワーク機能の一般的な原則の探索に有害である1、2、3、4。ここでの目的は、ニューラルネットワーク機能の比較研究を促進するために、胃ポッドニューラルネットワークの機能構造に関する迅速な洞察を得るための大規模なイメージングアプローチを実証することです。
Aplysia、Lymnaea、トリトニア、プリユーロブランチアなどの胃ポッド軟体動物は、神経管の表面に位置する大規模でしばしば識別可能なニューロンによってそれらの行動が媒介されているため、神経ネットワーク機能の原理を調査するために長い間使用されてきました。 1970年代には、プラズマ膜に溶け込むことができる電圧感受性色素(VSD)が開発され、すぐに複数のニューロン6によって生成された作用電位の最初の無電極記録が可能になった。ここでは、神経科学の新しい1種、ベルヒア・ステファニーエを含む、いくつかの種の胃ポッドにおけるネットワーク活動を調べるためにVSDを使用することを実証します。撮像装置は、1,600フレーム/秒でサンプリングする市販の464要素フォトダイオードアレイ(PDA)(図1)であり、これは、高速吸光度VSDで使用すると、記録されたすべてのニューロン7の作用電位を明らかにする。全てのダイオードによって記録された信号は、PDA取得ソフトウェアの中でガングリオンの画像に重ね合わせた直後に表示され、同じ準備8,9で鋭い電極を用いて目的のニューロンを調査することを可能にする。
生のPDAデータでは、多くのダイオードはより大きなニューロンを冗長に記録し、多くは複数のニューロンからの混合信号も含まれています。転機は、独立した成分分析を用いた自動スパイクソート法の開発で、各生の464チャネルPDAデータセットを新しいトレースセットに迅速に処理し、記録されたすべてのニューロンがその作用電位10,11のみを含む別のトレースに現れる。
この記事では、フォトダイオードアレイと高速吸光度VSDを備えた胃ポッド神経系からの大規模なアクションポテンシャル記録の取得に関する重要なステップを概説します。 さらに、光学的に記録されたニューロンを機能アンサンブルに対してクラスタリングおよびマッピングするために使用できる分析方法と、発射痕12,13の簡易検査を通じてしばしば明らかでない集団レベルの特徴を特徴付けるために用いることができる方法を示す。
プロトコル
注 : 以下のワークフローの概要を 図 2に示します。
1. 振動を最小限に抑える
- 可能であれば、リグが1階にあることを確認し、エアテーブルよりも広い範囲の振動周波数を減衰させるスプリングベースの分離テーブルを使用します。
- ばねベースのテーブルを使用する場合は、それがフローティングであることを確認します(テーブルに何かを追加したり取り外すたびに調整する必要があります)。
- 必要に応じて、撮像中の気流を遮断する程度であっても、撮像室の振動ベースの騒音を可能な限り低減します。灌流システムにおける流体乱流に起因する振動を最小限に抑えます。
注: 取得中に神経の準備を動かしてはいけません。あらゆる種類の動きは、ダイオードを横切るコントラストエッジのシフトを生じ、実際の信号を作り出します。この手順が取得時に刺激を伴う場合、準備の動きを誘発してはならない。
2. ピンホールテストを実行して、ガングリオン写真とPDAデータの適切な位置合わせを可能にする
- 小さな穴が3つ入ったアルミホイルを顕微鏡のスライドに置きます。そのパーフォーカルフォトポートに取り付けられたデジタルカメラで3つの穴の画像を撮ります。
- PDAに付属のイメージングソフトウェアを使用して、短いファイル(例えば、5 s)を取得します。取得の途中で、テーブルをタップしてピンホールの縁の周りに非常に目に見える振動アーティファクトを誘導し、ピンホールの画像を光学データと正確に整列させます。
- メニュー項目「表示」にあるイメージングソフトウェアで スーパーインポーズ 機能を使用|ページのスーパーインポーズ |外部イメージ|を使用してトレースを重ね合わせる「画像を重ね合わせる」は、ダイオードデータをピンホールの写真にオーバーレイし、ピンホールがダイオードデータのピンホールアーチファクトのすぐ上に置かれるまで、写真のx、y、倍率の設定を繰り返し調整します。
- これらの数字を保存して、カメラで撮影した準備画像を、今後の実験でダイオードデータと一致させます。
注: PDA のピンホールアライメントは、PDA を顕微鏡に取り付けた後、回転または取り外しが完了するまで 1 回だけ実行する必要があります。
- これらの数字を保存して、カメラで撮影した準備画像を、今後の実験でダイオードデータと一致させます。
3. 3種の海洋ガストロポッド種の切片
- トリトニアやアプリシアのような大きなサイズに成長する種の場合、より薄く、不透明でない神経節を持つ小さな個体から始まり、最適な信号対雑音に十分な光を得ることを緩和する。
- 分節および画像実験のための生理的な物として使用される準備ができる人工海水を持っている。
注: プロトコルの以降のすべてのステップでは、「生理」は人工海水を意味します。 - トリトニア・ディオメデアの解剖
- 動物を冷蔵庫に20分ほど置き、麻酔をします。
- 大きな動物の場合は、動物を片手に持って脳を露出させ、頭の端を人差し指の上にドレープして「首」を露出させます。小さな動物の場合は、脳を露出する前に、ワックスが並んだ解剖皿に後側をピン留めします。
- 解剖はさみを使用して、動物の側側、頬の塊(体壁を通して感じることができる)の上に3〜4センチメートルの正中線切開を行います。
注:CNSの必要な部分は、融合された両側性脳胸骨とペダル神経節からなり、オレンジ色で周囲の組織とは異なる外観です。それはサイの後部と頬の塊の上にあります。 - 鉗子とマイクロディセクチャを使用して動物の体を内面化する神経を切断し、中央神経節をつなぐすべての神経をそのまま維持することによってCNSを切除する。ペダル神経3(PdN3)の長い長さを残すか、または神経が刺激されるいずれかの。
- minutienピンを使用して、生理食糸で満たされたエラストマー裏地の皿の底にCNSを貼り付け、さらなる解剖を行います。蠕動ポンプを使用してフィードバック制御、インラインペルティエ冷却システムと配信された生理食物と料理を浸透させることによって、11 °Cで準備温度を維持します。
- 鉗子とマイクロディスセクションはさみを使用して、CNSの周囲から結合組織の緩やかに付着した層を慎重に取り除きます。細かい鞘をガングリアに密接に付着したままにしておきます。
- 簡単に (10 s) 生理食道中の 0.5% グルタルアルデヒドの溶液に神経節を浸します。食後に戻って神経節を食べ物を入れ、生理食類を使ってVSD染色を開始する前にグルタルアルデヒドを洗い流します。
注:結合組織とその固有の筋肉のこの光の修正は、イメージング中の動きを防ぐのに役立ちます。
- 時立性カリフォルニカの解剖
- 約40gの動物を、腹側表面(足)を通して約20mLの350mMMgCl2 を体内に注入して麻酔をします。
- ピンを使用して、動物の腹側をワックスで並べた解剖皿に並べます。
- 解剖はさみを使用して、足の最も前部の範囲に沿って2〜3センチメートルの正中線切開を行います。切開の両側の足のフラップをピン留めして、CNSと頬の質量の一部を明らかにします。
注:CNSの必要な部分は、融合した脳神経節からなり、密接にアパポーズされた両側胸膜とペダル神経節は黄色オレンジ色で、周囲の組織とは外観が異なります。それは球根性筋頬の塊に後部および後部に座る。 - 鉗子と解剖はさみを使用して、頬の塊を慎重に解剖し、脳神経節を明らかにする。
- 鉗子とマイクロディセクチャを使用して動物の体を内面化する神経を切断し、中央神経節をつなぐすべての神経をそのまま維持することによってCNSを切除する。ペダル神経9(PdN9)の長い長さを残すか、または神経が刺激されるいずれかの。
- minutienピンを使用して、生理食糸で満たされたエラストマー裏地の皿にCNSを配置します。ペルチェ冷却装置を通して生理食い物を皿に浸透させることによって15-16 °Cで準備温度を維持する。
- 鉗子とマイクロディセクチャを使用して、CNSから過剰な結合組織を取り除き、画像化されるガングリオンまたは神経節の鞘の表面的な部分を解剖する。このプロセス中に、鞘に穴が開かないように注意してください。
- 簡単に (20 s) 生理食道中の 0.5% グルタルアルデヒドの溶液に神経節を浸します。食後に戻って神経節を食べ物を入れ、生理食類を使ってVSD染色を開始する前にグルタルアルデヒドを洗い流します。
- ベルガ・ステファニーの解剖
- 動物を冷蔵庫に20分ほど置き、麻酔をします。
- 室温の生理食物で満たされたエラストマー裏地の皿を使用して、頭と尾の両方にminutienピンを置きます。
- マイクロディションハサミを使用して、5〜7mmの後部切開をCNS表面にします。
注:目は、CNSの横の動物の中に存在する暗い斑点は、二国間で融合した脳胸骨とペダル神経節から成り、頬の塊の上に座っている必要な部分CNSの位置を便利にマークします。 - 鉗子とマイクロディセクチャを使用して動物の体を内面化する神経を切断することによってCNSを切除する。吸引電極のために十分に長く刺激されている任意の神経を残します。
4. 分圧性染料で調製を汚す
- RH155(NK3041とも呼ばれる)またはRH482(NK3630またはJPW1132としても知られている)のいずれかのストック溶液を準備します。
- RH155: 5.4 mgの固形色素を1mLの100%EtOHに溶解し、29 μLを34マイクロ遠心チューブのそれぞれにピペット化します。各チューブの内容物を空気にさらして、暗闇の中で一晩乾燥させます。得られた固形アリコートを、それぞれ0.15mgを含むRH155の固形アリコートを、-20°Cの冷凍庫に入れる。
- RH482: 2 mg の固形色素を DMSO の 100 μL に溶解し、溶液を 20 個のアリコート 5 μL に分割し、それぞれ 0.1 mg の RH482 を含み、-20 °C 冷凍庫に保存します。
注意: トリトニア および 無呼吸の場合、浴灌流または圧力適用は、調製中のニューロンの膜にVSD RH155をロードするために使用することができる。圧力アプリケーションには、VSDに画像化されているガンギだけを公開するという利点があります。
- 浴灌流の場合、固体RH155と渦の2つのアリコートのそれぞれに生理食塩水を5 mL加えて溶液に入れ、0.03 mg/mL RH155を含む10 mLの結合溶液を製造する。
- 暗闇の中で(光の漂白を避けるために)トリトニアの場合は11°C、Aplysiaの場合は16°Cで1〜1.5時間のパーフューズ。 ペルチェ冷却システムを介して灌流液を通過させることにより、温度を維持します。
- 圧力適用のために、RH155の1つのアリコートに500 μLの生理食塩水を加え、渦を加え、0.3 mg/mLの染料濃度を作り出す。
- ハンドヘルドマイクロディスペンサーを使用して溶液の約200 μLをポリエチレンチューブに引き込み、チューブ径とガングリオンの直径との間に良好な一致が見られるようにします。
- マイクロマニピュレーターを使用して、チューブの端をターゲットの神経節の上に慎重に配置し、ガングリオンにぴったりしたシールを形成するまで下げます。上記の冷却システムの種類を使用して、江疽神経系を所望の温度に保ちます。
- 部屋のライトを暗くして光の漂白を避け、5分ごとにマイクロディスペンサーアプリケーターノブを回して、より多くの染料をガングリオンに押し付けます。
- 30分で良好な染色が行われていることを確認し、約1時間の完全染色時間を継続します。
- ベルヒアで染色する場合は、RH482の凍結アリコートに1mLの生理食塩水を加え、渦を溶かします。
- この溶液の200 μLを、800 μLの生理食糸と渦を含むマイクロ遠心分離チューブに溶液に移し、0.1%DMSOの生理食糸で0.02 mg/ml RH482の最終染色液を生成します。
- CNS全体をマイクロ遠心チューブに入れ、チューブをアルミニウム箔で包み、光の漂白を避け、5~6分ごとに手で1時間ほど振ります。最初の溶液の残りの800 μLを冷蔵庫に保管し、その後の調製物を染色するために最大3日間使用します。
5. 準備を平らにして神経刺激を設定する
注: このセクションの手順は、光の点滅を最小限に抑えるために、最小限の照明または緑色のライトで実行する必要があります。
- 染色後、CNSを撮像チャンバー内の生理食い物に浸し、解剖顕微鏡の下に置きます。
- シリコーン片( トリトニア または Aplysia用)または石油ゼリーの塊( ベルジア用)を、ガングリオン/神経節の左右に配置して画像化します。
- 適切なサイズのガラスまたはプラスチック製のカバースリップを準備に押し下げて平らにします。しっかりと押すが、ニューロンを損傷するほど難しくない。
注:この方法で準備の凸面を平らにすると、より多くのニューロンが共焦点を引き起こし、それによって記録されるニューロンの数が増え、さらにイメージング中に準備を固定するのに役立ちます。 - 神経を刺激して架空の運動プログラムを引き出す場合は、前先端が神経径とほぼ同じ幅の吸引電極を用意します。これを実現するには、PE-100ポリエチレンチューブのセグメントを炎の上で慎重に溶融させ、チューブセグメントの両端をそっと引っ張り、目的のポイントでテーパーを切断します。
- ポリエチレン吸引電極の先細り端部を通して少量の生理焼言を引き、続いて刺激される神経の端部を、電極の後端に厚い、柔軟なポリマーチューブの長さを取り付け、口の吸引を使用して負圧を加える。
- 電極の生理的な部分に、電気伝導を中断する気泡が欠けていることを確認します。
6. イメージングの準備と最適化
- チャンバーをイメージングリグに移動します。記録チャンバーを通して生理食糸灌流を開始し、準備の近くに温度プローブを置きます。画像化される種に必要な温度コントローラを設定します(トリトニア、11°C、Aplysia、15-16 °C、またはベルジア、26〜27°C)。
- 1本の塩化銀線を吸引電極の下に置き、電極内の生理食塩水に接触することを確認し、もう一方のAg-AgClワイヤー(リターンパス)を吸引電極の近くの浴塩水に入れます。
- 水浸漬レンズを生理食糸に下げます。ベースダイヤフラムを閉じてから、サブステージコンデンサを上げたり下げたりして、ダイアフラムのエッジが鋭い焦点になるまで焦点を調整し、ケーラーのイルミネーションを作成します。
- 画像化する準備の領域に焦点を当てます。大きなニューロンに由来する光学信号は、少し焦点が合っていない場合でも登録される可能性が高いため、より大きなニューロンよりも小さなニューロンに焦点を当てたバイアスが登録される可能性が高くなります。
- パーフォーカルデジタルカメラで撮影するガングリオンの写真を撮ります。
- コントロールパネルのゲインスイッチを1xに設定し、「RLI」ボタンをクリックし、ダイオードの平均RLIを確認して、イメージングソフトウェアの安静光強度 (RLI)を検査します。刺激装置からLEDランプ電源に送られる電圧レベルを調整し、希望の範囲(通常は3-4 V前後)になるまで平均RLIレベルのチェックを続けます。
注: PDA の約 3 ~ 4 V に対応する高い Rli が望ましいです。光が高いほど光信号の信号対雑音比が良くなりますが、これは高いRlIでの光漂白速度とバランスを取る必要があります。このリスクは、高NA対物レンズを使用することで最小化されます。使用される水浸し対物レンズは、10x/0.6 NA、20x/0.95 NA、40x/0.8 NA、および40x/1.15 NAです。
- 記録用にコントロールパネルのゲインスイッチを100倍に設定します。
- 神経を刺激する場合は、光レベルを設定するために使用される刺激装置とは別の刺激装置に、希望する電圧、周波数、および持続時間を設定します。コントロールパネルと刺激装置の間でTTLトリガが正しく設定されていることを確認します。
注:各種のサンプル神経刺激パラメータは次のとおりです: トリトニア PdN3、2 s、10 Hzパルス列5 ms、10 Vパルス; 時立 PdN9、2.5 s、20 Hzパルス列5 ms、8 Vパルス; ベルガの ペダル神経、2 s、5 ms、5 Vパルスの10 Hzパルス列。 - ばねまたは空気テーブルが浮いているか再確認してください。
7. 光記録
- 室内の照明をオフにするか、または暗くします(オーバーヘッド蛍光灯を含む)。
- 目的のファイルの継続時間、パス、およびファイル名を設定し、イメージング ソフトウェアの [データの取得] ボタンをクリックして、コンピュータの使用可能な RAM 容量までファイルを取得します。小さな振動が光記録データに大きなアーチファクトを導入する可能性がありますので、光記録中はそのまま残ります。
注: コンピュータの利用可能な RAM を超える取得については、ジョージタウン大学の Jian-young Wu 博士を通じて、カスタムメイドの C++ 取得プログラムを利用できます。 - 取得直後にデータを表示するには、撮像ソフトウェアで スーパーインポーズ 機能を使用して、464ダイオード全てによって収集されたデータを、前の準備7で撮影したガングリオンの画像に重ね合わせる。ソフトウェアに示されているダイオードのいずれかをクリックすると、別のトレース画面に記録された内容が展開されます。
- ピンホール試験であらかじめ決められたx、y、倍率を入力して、準備に対してダイオードの正確な位置合わせを実現します。
- アクションポテンシャルの可視性を最大化し、その後のスパイクソート14のニューロン収量を改善するために、5 Hzと100 Hzのカットオフ(イメージングソフトウェアで利用可能)を備えたバンドパスバターワースフィルタを適用して、低周波ノイズと高周波ノイズの両方を除去します。
- 科学コンピューティング プラットフォームで、さらに分析するためにフィルタ処理された光学データをテキスト ファイルとして保存するには、まずイメージング ソフトウェアの [ページ] 画面の下にある[TP フィルタ] ボックスを選択します。次に、[出力] タブの[ページを ASCII 形式で保存] を選択し、表示されるダイアログ ボックスに目的のファイル名を入力します。
結果
トリトニア
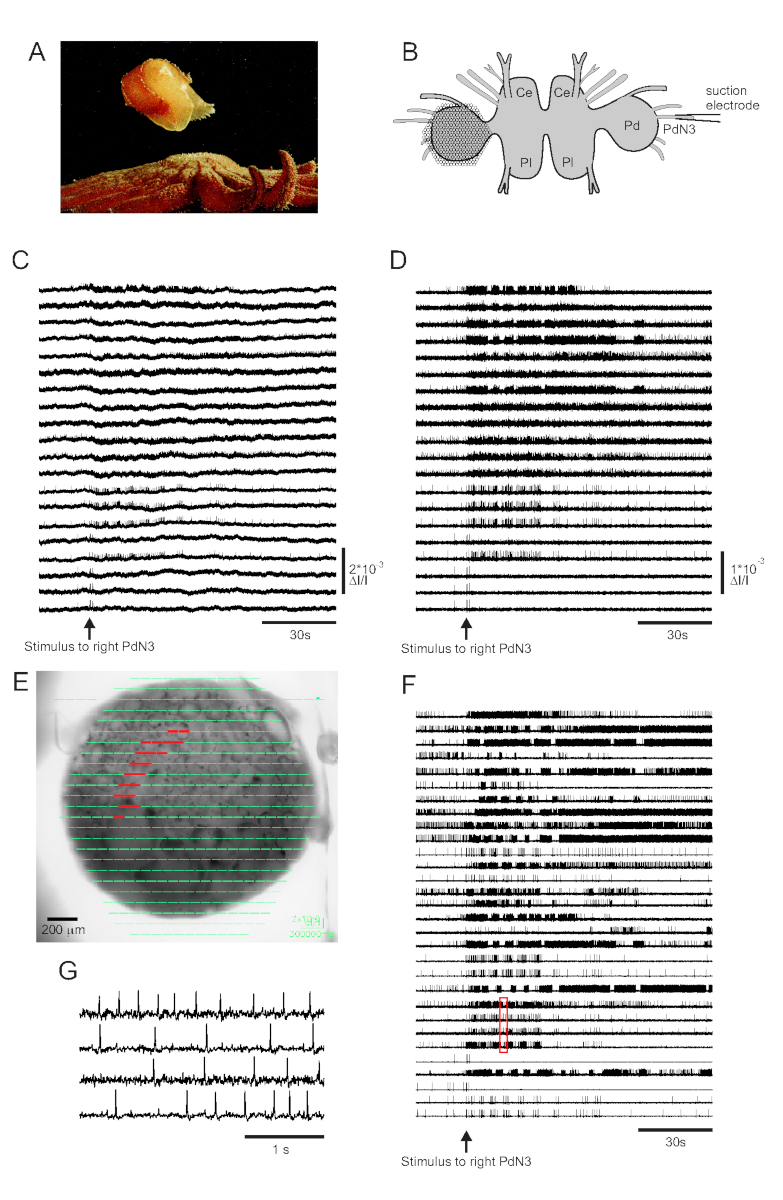
そのシースター捕食者との皮膚接触は、トリトニア・ディオメデアの脱出泳ぎを引き起こし、それを安全に推進する全身屈曲のリズミカルなシリーズからなる(図3A)。単離脳の準備では、ペダル神経3(PdN3)への短い刺激は、ペダル神経節からの光学記録で容易に認識可能であるこの行動のためのリズミカルな水泳運動プログラム(SMP)を引き出す。図3Bは、PDAが位置した左トリトニアペダル神経節の後面におけるニューロンの発火活性を記録するように設計されたVSDイメージング実験のレイアウトを、反側(右)PdN3がSMPを引き出す刺激として描いている。PdN3の刺激前、刺激中および後の20ダイオード記録活動からの生データおよびフィルタリングされたデータ(バンドパスバターワースフィルタ、5および100 Hzカットオフ)は、それぞれ図3C、Dに示されている。神経刺激は2分ファイルに20 sを送達した。取得直後に、記録アレイの464ダイオードの全てで測定された信号を、撮像ソフトウェアの準備の画像上に地形的に表示することができる(図3E)。この時点で、多くのトレースには同じニューロンから冗長的に記録されたスパイクが含まれ、いくつかのトレースには複数のニューロンからのスパイクが含まれています。フィルターされたダイオードトレースをICAでスパイクソートすると、53個のユニークなニューロントレースが生成され、そのうちの30個が図3Fに示されています。個々のスパイクの動態は、図3F(赤いボックス)からの4つの痕跡の抜粋を拡大する図3Gで理解することができます。ICAスパイク選別アルゴリズムの精度は、以前に同時に鋭い電極記録を使用して検証され、ソートされたトレース内のすべてのスパイクが個々のニューロン11、14からの細胞内に記録されたスパイクに対応することを示した。
時立
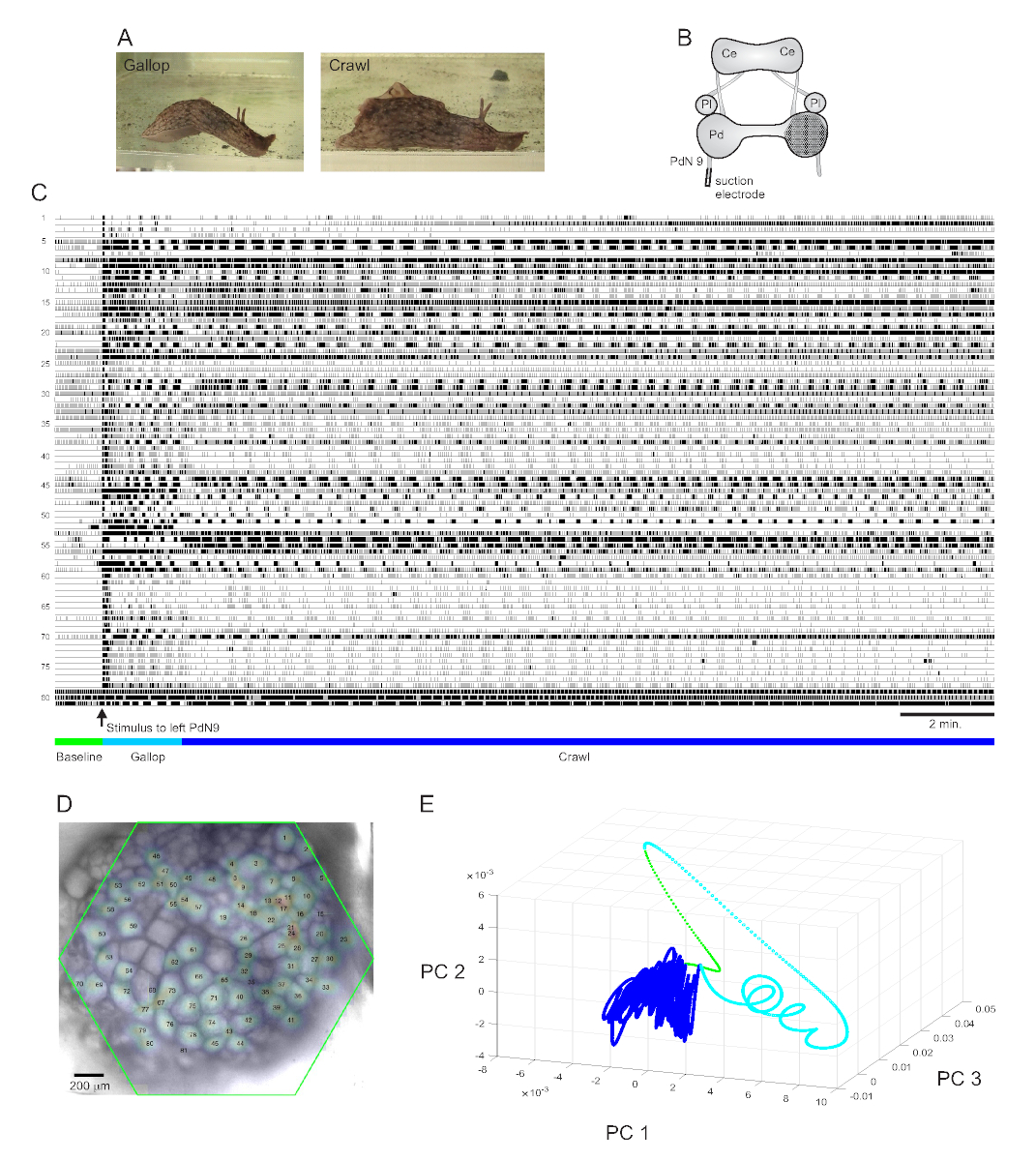
Aplysiaカリフォルニカへの強く回避的な尾の刺激は、ステレオタイプ化された2部のリズミカルエスケープ応答15を引き出す。応答の第1段階は、動物を素早く前進させる頭部肺および尾引きのいくつかのサイクルのギャロップである。これは通常、数分間遅い速度で動物を前方に動かす頭から尾への筋収縮の繰り返し波を伴うクロールの期間が続きます(図4A)。光学記録でこれらのエスケープモータープログラムをキャプチャするために、PDAは孤立した脳の準備で右ペダル神経節の後側表面に焦点を当て、吸引電極を対側(左)ペダル神経9(PdN9)に置いた。図4B)。連続的な20分の光学記録(図4C)に1分間、PdN9はギャロップ・クロール・モータ・プログラム・シーケンスを惹起するように刺激された。記録されたすべての81個のニューロンからの信号の確率的ガウス空間分布は、神経節にマッピングされた(図4D)。完全な記録に適用された次元減少は、エスケーププログラムのギャロップ(シアン)およびクロール(ダークブルー)フェーズが異なる領域を占め、それぞれ異なる軌道、スパイラル状およびループ状、主成分空間で形成されたことを明らかにした(図4E)。
図 4に示すAplysiaの記録に基づく 3 つのビデオは、このようなデータセットで実行できるさらなる種類の分析を示しています。ビデオ 1は、記録の全期間にわたって記録されたすべてのニューロンの発射をアニメーション化します。脱出運動プログラムの最初の刺激後の期間はギャロップによって特徴づけられ、ガングリオンの活動は異なる機能クラスタの交互破裂によってマークされた(Video 2)。ギャロップはその後、クロールに移行し、ニューロンクラスター全体の活動は広くフェージックなままであったが、神経節の反時計回りの回転軌道を仮定した(ビデオ3)。後者の2つのビデオにはコンセンサスクラスタリングも組み込まれており、エスケープ応答のギャロップとクロールフェーズの異なる機能的アンサンブルの発射と位置を別々に明らかにします。ギャロップとクロールの両方の段階で同じクラスターに割り当てられた多くのニューロンは、ガングリオン内で互いに物理的に近接し、以前の知見12と一致していることに注意してください。
ベルヒア
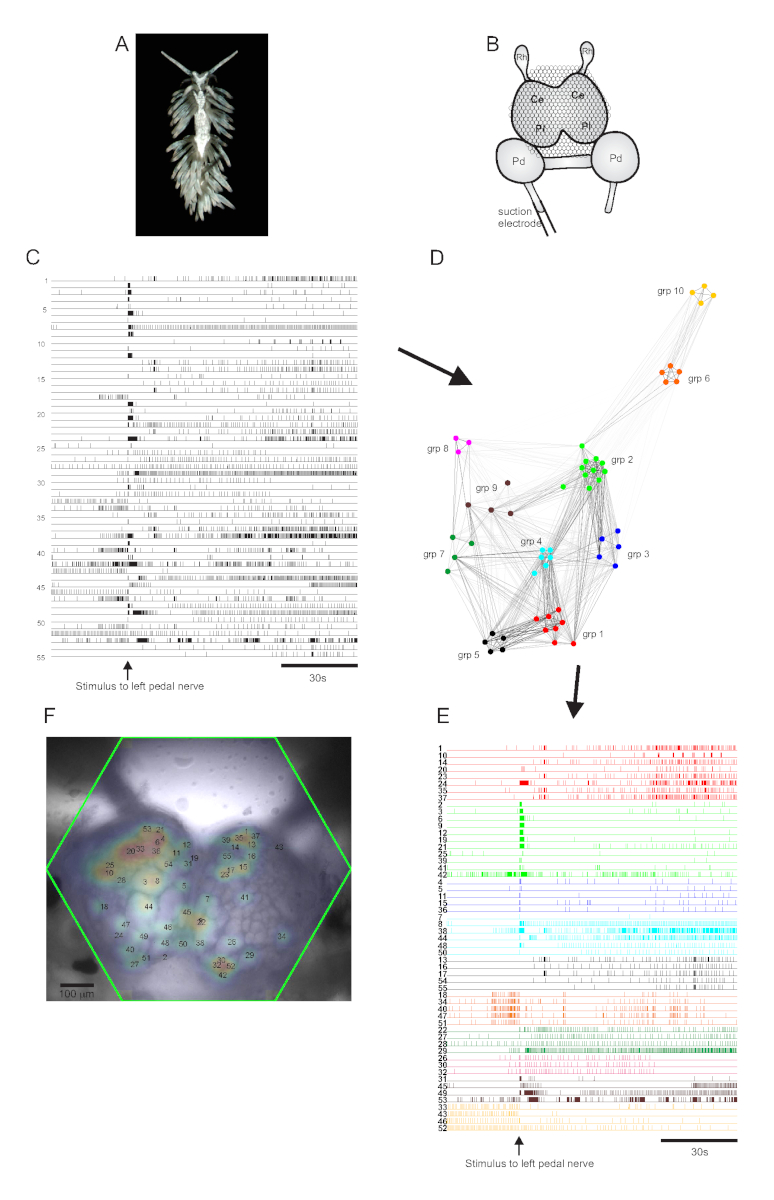
aeolid nudibranch ベルヒアステファニー (図5A)は、神経科学のための新しいモデルシステムを表しています。典型的な ベルグア 実験のイメージング設定を 図5Bに示します。広範囲の神経活動を惹起するために、最も顕著な左ペダル神経に吸引電極を置き、神経刺激を30sで2分の記録に送達した。ICA処理された痕跡は、55ニューロンにおける自発的および刺激誘発活性の両方を明らかにした(図5C)。コンセンサス クラスタリングによるコミュニティ検出では、図 5D に示されている 10 個の異なる機能アンサンブルを識別し、 図 5Eでは、クラスタリングの割り当てに基づいて 図 5C に示すトレースを再編成しています。すべての記録されたニューロンからの信号のガウス分布は、全55の記録されたニューロンの位置を示すために 図5F の準備の画像に重ね合わされる。

図1:光イメージングリグとフォトダイオードアレイ(PDA)の眺め。(A)PDA、デジタルカメラ、顕微鏡、およびステージを搭載した光学イメージングリグ。(B) 光ファイバーが464フォトダイオードに画像絞りを接続するPDAの内部設計。アンプの列は、フォトダイオードの上にあります。(C)撮像アパーチャーの六角形の面を、画像化される領域が焦点を合わせたもの。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2:光記録を得る上で不可欠なワークフローを示すフローチャート。 VSDイメージングプロトコルの重要なステップは、解剖と染色からイメージングの詳細まで、このフローチャートに示されています。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図3:トリトニア・ジオメデアの結果、生、フィルター、スパイクソートされたデータを示す。(A)トリトニアは、体の背側と腹側の屈曲を交互に構成する泳ぎを通して捕食海の星ピクノポディア・ヘリアントイドから脱出する。(B)イメージング設定の概略図。脳神経節のCe=脳神経節の葉;Pl =脳胸骨神経節の胸葉;Pd = ペダルガングリオン。(C)20枚のフォトダイオードからの生データは、逆側PdN3の刺激(矢印で示される刺激)に左ペダル神経節の活動を表示する。(D) Cと同じダイオードからのフィルタリングされたデータ (5 および 100 Hz バンドパス バターワース フィルター)。(E) 464ダイオードで収集された圧縮トレースが準備の画像に重畳されるイメージングソフトウェア出力。トレースがCとDで示されている 20 個のダイオードの位置は、赤で強調表示されます。(F) ICAを介したスパイクソートによって生成された30個の単一ニューロントレースを選択した。(G) Fの赤いボックスに対応する 4 つの単一ニューロン トレースの拡大図は、より高い時間分解能でそれらの作用電位を表示します。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図4: Aplysia カリフォルニカの結果、 長時間記録、シグナルマッピング、および次元の縮小を示す。(A) Aplysiaの逐次脱出モータープログラムの2つの段階、ギャロップとクロール。(B)イメージング設定の概略図。Ce = 脳神経節;Pl = 胸膜神経節;Pd = ペダルガングリオン。(C)右ペダルガンギオンの81個のニューロンの20分の記録が、逆方向PdN9(矢印で示される)への刺激に反応する。トレースの下にある緑、シアン、濃い青色のバーは、それぞれ、刺激前の期間、ギャロップ、およびエスケープモータープログラムのクロール段階を示します。(D) ICAによって同定された全81の神経信号源の位置のマッピングされた確率的ガウス分布を用いた準備の画像。緑色のアウトラインは、ガングリオンに対するPDAの六角形の面の位置を表します。各ガウスの数字は、Cのトレース番号に対応しています。(E) 主成分分析を用いた寸法縮小は、20分のファイルの間に最初の3つの主要成分を互いにプロットする。刺激前のベースライン、ギャロップ、およびクロールエポックは、それぞれ緑、シアン、濃い青色で示されています。この記録に対応する神経発火のアニメーションについては、ビデオ1-3を参照してください。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図5:ベルガ・ステファニーエ、神経科学の新種、ネットワークグラフ化、機能クラスタリング、および両側シグナルマッピングの結果。(A)ベルガアの標本。(B)イメージング設定の概略図。Ce = 脳神経節の脳葉;Pl =脳胸骨神経節の胸葉;Pd = ペダルガングリオン;Rh = 鼻泳子ガングリオン。(C) 脳神経節における55の両側ニューロンの自発的および刺激誘発活性を示す痕跡(刺激の送達は矢印によって示される)。(D) コンセンサス クラスタリングによって識別される、それぞれ固有の色を割り当てた 10 個の機能アンサンブルを表示するネットワーク グラフ。このプロットのノードはニューロンを表し、ネットワーク空間の距離はアンサンブル内およびアンサンブル間の発火相関の程度を表します。(E) Cのトレースは、(Dの配色に従って) 機能アンサンブルに並べ替え、色分けされます。(F) 記録されたすべてのニューロンからの信号のマップ位置と、それらが対応するCおよびEのトレース番号を示す準備の画像。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
ビデオ1:完全な、20分のアプリジアエスケープ運動プログラムのアニメーション。右のペダル神経節の81個のニューロン(左パネル)に位置する白い形状の不透明度は、対応するニューロンの痕跡(右パネル)によって駆動され、平均スパイクレート(記録中のリアルタイムの0.61秒ごとにビン分割)の関数として直線的に変化した。各ニューロンについて、完全な不透明度を記録期間中の最大焼成率に正規化した。ビデオの経過時間の 1 秒は、12.2 秒のリアルタイムを表します。スケール バーはリアルタイムに対応し、トレースの下にある緑、シアン、濃い青色の線が、それぞれエスケープ ロコモーター プログラムのプリ刺激ベースライン、ギャロップ、およびクロール の各フェーズを示します。ギャロップフェーズの周囲の黄色のボックスとクロールフェーズの一部は、ビデオ 2と3のアニメーションを生成するために使用される記録の抜粋を示します。このビデオを見るには、ここをクリックしてください。(右クリックしてダウンロードします。
ビデオ 2:Aplysia エスケープ運動プログラムのギャロップフェーズのアニメーション。 コンセンサスクラスタリングは、運動プログラムのギャロップ段階で記録されたすべての81個のニューロンに対して行われ、機能的アンサンブルを導出し、ref.12で説明されたアプローチとソフトウェアを使用して行われた。脱出プログラムのこの段階の間に主に強壮性または不規則な発火パターンを示すニューロンアンサンブルは、このビデオから省略された。黒とオリーブグリーンのアンサンブルに属するニューロンのアクションポテンシャルは、ビデオのオーディオトラックで聞こえ、対応するニューロンとトレースが強調表示されます。平均スパイクレートは 、ビデオ1 と同等の時間ビニングと同様に正規化されました。ビデオの経過時間の1 sは、リアルタイムの6.1 sに対応しています。 このビデオを見るには、ここをクリックしてください。(右クリックしてダウンロードします。
ビデオ 3:Aplysia エスケープ運動プログラムのクロール段階のアニメーション。 コンセンサスクラスタリングは、機能的アンサンブルを導出するために、モータプログラムのクロール段階だけで81個の記録されたニューロンすべてに対して行われた。モータープログラムのこの段階で主に強壮性または不規則な発火パターンを示すアンサンブルは、このビデオから省略されました。平均スパイクレートは 、ビデオ1 と 2 のように正規化され、同等の時間ビニングで正規化されました。ビデオの経過時間の 1 s は、リアルタイムの約 12.2 s に対応します。 このビデオを見るには、ここをクリックしてください。(右クリックしてダウンロードします。
ディスカッション
大規模なVSDイメージングアプローチを実装する上で最も重要な点の1つは、振動を最小限に抑え、ダイオード全体でコントラストエッジの動きを生じさせ、大きなアーティフィファクト信号を生成することです。吸光度VSDは、作用電位を有する光強度の非常に小さな割合の変化を生じるので、振動アーティファクトは、妨げられなければ、関心のある神経信号をあいまいにすることができる。振動の発生を最小限に抑える方法はいくつかあります。まず、当社のイメージングルームは、1階に位置し、空気処理装置や他の多くのソースの構築に関連する振動から準備を隔離しています。第二に、バネベースの分離テーブルが使用され、他のPDAユーザーは、より一般的な空気テーブル16よりも優れた振動減衰を提供することを確認した。第三に、水浸漬目的が使用され、表面の波紋から生じる画像の変動を排除しました。第4に、画像化された調製物を、シリコーンプラグまたは石油ゼリーによって所定の位置に保持されている上から押し下げたチャンバーカバースリップ底部とカバースリップ片との間に軽く押し付け、さらに調製を安定化させる。これはまた、画像化されている神経節または神経節の凸面を平らにし、その結果、目的の焦点面でより多くのニューロンが得られ、記録されたニューロンの数が増加する。
アクションポテンシャルに起因するVSD光吸光度の変化の非常に小さい変化に対する信号対雑音比を最大化するには、PDAへの調製を通してほぼ飽和光を達成すると同時に、色素の光の漂白を最小限に抑えるために不可欠です。この作業を行うために、1x位置のPDAコントロールパネルゲインスイッチで測定した3~4Vの静止光強度(PDAの464アンプは10Vの光で飽和します)で動作します。データ取得中に、このゲイン係数は100倍に変更されます。PDAで測定された3-4Vに達するのに十分な光を得ることは、いくつかの方法で達成することができる。まず、使用する吸光度染料の吸収特性に適した波長を供給する超明るいLED光源を使用する。したがって、735nm LEDコリメートランプを使用し、RH155およびRH482の最適な吸収波長と重なった。第二に、必要に応じて、LED光源からの光をより小さな領域に集中させるフリップトップサブステージコンデンサーを使用します。第三に、高い、明るさおよび最大のイメージの質を保障するケーラーの照明を達成するために凝縮器の高さを調節する。第4に、LEDランプの735nm波長を減衰させる光経路にヒートフィルターがないことを確認してください。第五に、ディフューザーを除去し、より多くの光が必要な場合は、光学経路から除去する。第六に、高い空間分解能を提供する高NA目標を使用し、低いランプ強度でPDAに到達するのに十分なレベルの光を可能にする。これにより、すべてのファイルで同じ光強度を使用し、信号振幅の大幅な損失や再染色の必要性をなくして、準備ごとに10〜20分の複数の取得ファイルを取得できる範囲で、光の漂白を最小限に抑えることができ、私たちは可能にしました。重要なことに、実験者がこれらの長いファイル全体のニューロンを追跡したい場合は、焦点面が変化せず、準備が動かないことを確認してください。最後に、PDAに十分な光をルーティングする追加の方法は、より薄く、したがって不透明な神経節を少なくする若い動物を使用することです。
光信号の信号対雑音比が劣化したり、モータのリズムが最適でない(遅い、異常など)ことが時々分かります。これが一貫して起こり始めると、VSDの新鮮なソリューションをミックスします。VSDのアリコートは、通常、-20°Cの冷凍庫で約6ヶ月間生存可能なままである。関連して、 ベルガにとって、これまで最良の結果は、吸光度VSD RH482で得られていることは注目に値する。RH482はRH155よりも親油性であるため、 ベルヒアの比較的小さいニューロンを染色する方が良いか、この熱帯種に使用されるより高い記録生理食塩水温度でより効果的に神経膜に残る可能性があります。
神経活動のPDAベースのイメージングの1つの制限は、100xプリアンプ化ステップの前にハードウェア内の電圧信号のACカップリングに関連する:これは、この技術で必要とされる高い静休み光レベルによって生成される大きなDCオフセットを除去するために必要な機能を表しているが、PDAに固有のAC結合は膜電位の遅い変化の測定を妨げる、 シナプス入力に関連するものなど。低速または定常状態の可能性の変化を記録することが望ましい場合は、DC結合CMOSカメライメージングシステムを使用してサブスレッショルドアクティビティをキャプチャできます。Byrneたちは最近、RH155でこのようなセットアップを使用して、Aplysia17, 18の頬神経節におけるニューロンの活動を画像化した。我々は、両方のシステムを使用し、検出器(128 x 128)の密度が非常に高いため、CMOSカメラが同じイメージング時間7で50倍大きなデータファイルを生成することを発見しました。PDAの小さなファイルは、処理と分析の高速化を促進します。また、拡張シングルトライアル記録(図4)と学習の研究が可能になり、複数の試行のデータをスパイクソートの前に1つの大きなファイルに連結し、学習が発展する19としてネットワーク組織を追跡することができます。
他のカメラベースの調査では、クリスタンと同僚が蛍光VSDを使用して、リーチのセグメント性神経節のネットワーク機能を調べている。ある有力な研究では、これは泳ぐか、またはクロールする動物の決定に関与するニューロンの同定につながった 20 .別の研究では、Kristanらはリーチの水泳とクロールの行動が多機能対専用回路21によってどの程度駆動されているかを調べた。最近では、Wagenaarたちは、リーチセグメント神経節22のほぼすべてのニューロンから記録することを可能にする電圧イメージングに両面顕微鏡を使用した。多くのカメラベースのイメージング方法とは対照的に、PDAベースのイメージング法の利点は、結果処理のための神経境界に関する決定を伴さない盲目のソース分離の一形態であるICAによる迅速かつ公平なスパイクソーティングです。
VSDの選択に関して、吸光度染料RH155およびRH482の1つの利点は、それらに関連するほとんどない光毒性23、24であり、蛍光VSDの典型的なよりも長い記録時間を可能にする。さらに、我々が使用する高速吸光度VSDは、通常振幅80 mVである胃ポッド調製におけるオーバーシューティング体細胞作用電位を記録するのに適しています。図3Gに示すように、私たちの光学的手法は、アクションポテンシャルのアンダーシュートを記録することができます(私たちの録音のどれもトレース平均ではありません):これは、私たちが使用するVSDがある程度減衰し、したがって、彼らがソーマに達するまでにオーバーシュートされていない他のモデルシステムのアクションポテンシャルを識別することができるべきであることを示唆しています。それにもかかわらず、我々の光学的アプローチは、相馬に記録された場合に高度に減衰する作用電位を示すことが知られている種にとって理想的ではないかもしれない。
ニューラルネットワークに関する現在の研究の多くは、少数のデザイナートランスジェニック種に焦点を当てています。しかし、神経科学は、多種多様な系統学的に異なる種の研究の恩恵を受けます。多くの異なる種を研究することは、回路がどのように進化するかについての洞察を提供します25,26, phyla全体で一般的である可能性のあるネットワーク機能の原理を照らします1,2,3,4,27.これまでに、我々のイメージング法を、Aplysiaカリフォルニカ8、11、12、13、14、28、トリトニア・ディオメデア8、9、11、14、19、28、トリトニアイバ28、及び胃ポッド種の数に適用してきました。 Pleurobranchaeaカリフォルニカ(未発表データ)、そして最近ベルヒアステファニー (図5)。このアプローチの魅力は、トランスジェニック動物を必要としない多くの種に容易に適用できることである。我々は、高速吸光度染料とPDAを用いたVSDイメージングの使用が、ナバナックス29とAplysia30の準備を行う半無傷でこれを達成した先駆的な作業の足跡をたどることを認めたい。我々のアプローチの迅速性に重点を置くことは、神経科学31に広範な関心を持つ科学的な問題を探求する前に、長年の研究が基本的なネットワーク組織を特徴付けるために何年も必要であるという懸念のために、多くの研究者が新種のネットワーク研究を開始することにますます消極的になるかもしれないという懸念に対する答えの一部である。したがって、ここでの目標は、単一の準備からネットワーク組織に関する重要な即日の洞察を得ることができるという点まで、プロセスを大幅にスピードアップする手法を実証することです。
開示事項
著者らは開示するものは何もない。
謝辞
この作業は、NSF 1257923とNIH 1U01NS10837によってサポートされました。著者らは、研究室でのジャン・ワンの支援を認めたい。
資料
| Name | Company | Catalog Number | Comments |
| Achromat 0.9 NA swing condenser | Nikon | N/A | |
| Bipolar temperature controller | Warner Instruments | CL-100 with SC-20 | Controls perfusion saline temperature |
| Chamber thermometer | Physitemp | BAT-12 with IT-18 microprobe | |
| Digital camera | Optronics | S97808 | |
| Dissecting forceps | Dumont | #5 | |
| Dissecting scissors | American Diagnostic Corp. | ADC-3410Q | |
| Imaging microscope | Olympus | BX51WIF | |
| Imaging perfusion chamber | Siskiyou | PC-H | |
| Instant Ocean | Instant Ocean | SS6-25 | Makes 25 gallons at a time |
| Master-8 pulse stimulator | A.M.P.I. | Master-8 | |
| Microdispenser | Drummond Scientific | 3-000-752 | Dye applicator for pressure staining |
| Microdissection scissors | Moria | 15371-92 | |
| Minutien pins (0.1 mm) | Fine Science Tools | NC9677548 | For positioning and stabilizing CNS |
| Motorized microscope platform | Thorlabs | GHB-BX | Gibraltar platform |
| NeuroPlex imaging software | RedShirtImaging | NeuroPlex | Compatible with the WuTech photodiode array |
| Objective lenses | Olympus | XLPLN10XSVMP, XLUMPLFLN20XW, LUMPLFLN40XW, UAPON40XW340 | |
| PE-100 polyethylene tubing | VWR | 63018-726 | Tubing to make suction electrodes |
| Perfusion pump | Instech | P720 with DBS062SDBSU tube set | |
| Petroleum jelly | Equate | NDC 49035-038-54 | |
| Photodiode array with control panel | WuTech Instruments | 469-IV photodiode array | Contact jianwu2nd@gmail.com for ordering information |
| RH155 | Santa Cruz Biotechnology | sc-499432 | Voltage-sensitive dye |
| RH482 | Univ of Conn. Health Center | JPW-1132 | Voltage-sensitive dye; special order from Leslie Leow |
| Silicone earplugs | Mack's | Model 7 | To be use for preparation compression |
| Staining PE tubing | VWR | 63018-xxx | Different sizes depending on fit |
| Sylgard 184 silicone elastomer kit | Dow Corning | Sylgard 184 silicone elastomer kit | |
| Thorlabs LED and driver | Thorlabs | M735L2-C1, DC2100 | LED lamp and driver |
| Tygon tubing | Fisher Scientific | 14-171-xxx | |
| Vibration isolation table | Kinetic Systems | MK26 | Spring-based |
参考文献
- Miller, C. T., Hale, M. E., Okano, H., Okabe, S., Mitra, P. Comparative Principles for Next-Generation Neuroscience. Frontiers in Behavioral Neuroscience. 13 (12), (2019).
- Brenowitz, E. A., Zakon, H. H. Emerging from the bottleneck: benefits of the comparative approach to modern neuroscience. Trends in Neuroscience. 38 (5), 273-278 (2015).
- Bolker, J. Model organisms: There's more to life than rats and flies. Nature. 491 (7422), 31-33 (2012).
- Carlson, B. A. Diversity matters: the importance of comparative studies and the potential for synergy between neuroscience and evolutionary biology. JAMA Neurology. 69 (8), 987-993 (2012).
- Chase, R. . Behavior and its neural control in gastropod molluscs. , (2002).
- Salzberg, B. M., Grinvald, A., Cohen, L. B., Davila, H. V., Ross, W. N. Optical recording of neuronal activity in an invertebrate central nervous system: simultaneous monitoring of several neurons. Journal of Neurophysiology. 40 (6), 1281-1291 (1977).
- Frost, W. N., et al. Monitoring Spiking Activity of Many Individual Neurons in Invertebrate Ganglia. Advances in Experimental Medicine and Biology. 859, 127-145 (2015).
- Frost, W. N., Wang, J., Brandon, C. J. A stereo-compound hybrid microscope for combined intracellular and optical recording of invertebrate neural network activity. Journal of Neuroscience Methods. 162 (1-2), 148-154 (2007).
- Frost, W. N., Wu, J. -. Y., Covey, E., Carter, M. Voltage-sensitive dye imaging. Basic Electrophysiological Methods. , 169-195 (2015).
- Brown, G. D., Yamada, S., Sejnowski, T. J. Independent component analysis at the neural cocktail party. Trends in Neuroscience. 24 (1), 54-63 (2001).
- Hill, E. S., Moore-Kochlacs, C., Vasireddi, S. K., Sejnowski, T. J., Frost, W. N. Validation of independent component analysis for rapid spike sorting of optical recording data. Journal of Neurophysiology. 104 (6), 3721-3731 (2010).
- Bruno, A. M., Frost, W. N., Humphries, M. D. Modular deconstruction reveals the dynamical and physical building blocks of a locomotion motor program. Neuron. 86 (1), 304-318 (2015).
- Bruno, A. M., Frost, W. N., Humphries, M. D. A spiral attractor network drives rhythmic locomotion. ELife. 6, 27342 (2017).
- Hill, E. S., Bruno, A. M., Vasireddi, S. K., Frost, W. N., Naik, G. R. ICA applied to VSD imaging of invertebrate neuronal networks. Independent Component Analysis for Audio and Biosignal Applications. , 235-246 (2012).
- Jahan-Parwar, B., Fredman, S. M. Neural control of locomotion in Aplysia: role of the central ganglia. Behavioral and Neural Biology. 27 (1), 39-58 (1979).
- Jin, W., Zhang, R. J., Wu, J. Y. Voltage-sensitive dye imaging of population neuronal activity in cortical tissue. Journal of Neuroscience Methods. 115 (1), 13-27 (2002).
- Neveu, C. L., et al. Unique Configurations of Compression and Truncation of Neuronal Activity Underlie l-DOPA-Induced Selection of Motor Patterns in Aplysia. eNeuro. 4 (5), 17 (2017).
- Cai, Z., Neveu, C. L., Baxter, D. A., Byrne, J. H., Aazhang, B. Inferring neuronal network functional connectivity with directed information. Journal of Neurophysiology. 118 (2), 1055-1069 (2017).
- Hill, E. S., Vasireddi, S. K., Wang, J., Bruno, A. M., Frost, W. N. Memory Formation in Tritonia via Recruitment of Variably Committed Neurons. Current Biology. 25 (22), 2879-2888 (2015).
- Briggman, K. L., Abarbanel, H. D., Kristan, W. B. Optical imaging of neuronal populations during decision-making. Science. 307 (5711), 896-901 (2005).
- Briggman, K. L., Kristan, W. B. Imaging dedicated and multifunctional neural circuits generating distinct behaviors. Journal of Neuroscience. 26 (42), 10925-10933 (2006).
- Tomina, Y., Wagenaar, D. A. A double-sided microscope to realize whole-ganglion imaging of membrane potential in the medicinal leech. ELife. 6, 29839 (2017).
- Chang, P. Y., Jackson, M. B. Interpretation and optimization of absorbance and fluorescence signals from voltage-sensitive dyes. Journal of Membrane Biology. 196 (2), 105-116 (2003).
- Parsons, T. D., Salzberg, B. M., Obaid, A. L., Raccuia-Behling, F., Kleinfeld, D. Long-term optical recording of patterns of electrical activity in ensembles of cultured Aplysia neurons. Journal of Neurophysiology. 66, 316-333 (1991).
- Katz, P. S. Evolution of central pattern generators and rhythmic behaviours. Transactions of the Royal Society of London, Series B. 371 (1685), 20150057 (2016).
- Moroz, L. L. Biodiversity Meets Neuroscience: From the Sequencing Ship (Ship-Seq) to Deciphering Parallel Evolution of Neural Systems in Omic's Era. Integrative and Comparative Biology. 55 (6), 1005-1017 (2015).
- Frost, W. N., Tian, L. -. M., Hoppe, T. A., Mongeluzi, D. L., Wang, J. A cellular mechanism for prepulse inhibition. Neuron. 40, 991-1001 (2003).
- Hill, E. S., Vasireddi, S. K., Bruno, A. M., Wang, J., Frost, W. N. Variable neuronal participation in stereotypic motor programs. PLoS One. 7 (7), 40579 (2012).
- London, J. A., Zecevic, D., Cohen, L. B. Simultaneous optical recording of activity from many neurons during feeding in Navanax. Journal of Neuroscience. 7 (3), 649-661 (1987).
- Wu, J., Cohen, L. B., Falk, C. X. Neuronal activity during different behaviors in Aplysia: A distributed organization. Science. 263 (5148), 820-823 (1994).
- Marder, E., North, G., Greenspan, R. J. Searching for insight. In Invertebrate Neurobiology. , 1-18 (2007).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved