ショウジョウバエにおける糖誘発局所探索行動の追跡
要約
このプロトコルはショウ ジョウバエのmelanogasterを使用して砂糖誘発された調査の行動を記録するための行動の試金を記述する。このアッセイは、摂食や採餌に関連する行動、および根底にあるニューロンのメカニズムを研究するために利用できます。
要約
採餌行動は、生物が不可欠な食料資源を見つけて獲得することを可能にするため、生物の生存に不可欠です。 ショウジョウバエでは、飢餓は少量の糖溶液の消費に続く明確な検索行動を引き起こします。本報告では、糖分を誘発する検索行動を研究するための簡単な実験方法を提示し、そのメカニズムを明らかにすることを目指している。微量の濃縮糖溶液は、ハエの持続的な探索行動を誘発します。この行動には経路統合が関与しており、ハエはその軌道を利用して糖の位置に戻ることが確立されています。最新の知見は、砂糖摂取後の探索行動の開始と強度における時間的調節の証拠を提供する。また、咽頭の特定の味覚受容体ニューロンを人工的に活性化し、探索行動を誘発するためにも、この装置を用いています。 ショウジョウバエ の神経遺伝学的ツールキットは、採餌の根底にある神経および遺伝的メカニズムを研究するために、糖分を誘発する探索行動パラダイムと組み合わせることができる多様なツールと技術を提供します。ハエの飢餓による探索行動の神経基盤を理解することは、神経生物学の分野全体に貢献し、他の生物だけでなくヒトの摂食行動を支配する制御メカニズムへの洞察を提供します。
概要
食物探索と採餌行動は、分類群全体の生物が示す基本的な生存戦略です。昆虫では、飢餓による餌探しと食後の局所探索の2種類の採餌行動が確認されています1。お腹が空くと、昆虫は感覚の手がかりを頼りに食料源を見つけます。小さな餌場に遭遇して食べると、入り組んだ道と餌の場所の周りを旋回することを特徴とする局所的な検索行動を開始します。
局所検索の一種である砂糖誘発検索行動は、60年以上前にアメリカの生物学者Vincent Dethierによって初めて研究されました2。飢餓状態になると、ハエは満腹にならないように少量の砂糖を与えられ、局所的な捜索を開始します。典型的な捜索行動は、非常に曲がりくねった歩行を特徴とし、移動速度が低く、回転率が高く、砂糖が落ちた場所に戻ります。その後の研究では、イエバエとショウジョウバエでこの行動が調査されました3,4。探索の開始、強度、および期間は、動物の内部状態(例えば、剥奪や動機付け)と、資源の利用可能性や質などの外的要因によって調節される1,5,6。
追跡技術の進歩により、研究者は制御された領域内の行動を捕捉して分析するための貴重なツールを手に入れました。ここでは、砂糖摂取後に自由に歩くハエを追跡するための行動パラダイムを提示します。この簡単なセットアップにより、アリーナに用意された濃縮糖溶液に反応したハエの動きを捕捉して分析することにより、ショウ ジョウバエ の糖誘発探索行動を研究できます。高度な追跡技術とデータ解析技術を用いて、自発運動パターン、空間探査、糖刺激に対する応答動態の定量化に成功しました。
このアッセイを用いて、糖が誘発する探索は経路積分の使用を含み、糖の摂取量から空間的に時間的に分離できることが実験的に実証された7,8。さらに、この行動は、咽頭味覚ニューロンの活性化によって引き起こされ得ることが示されている9。最近の研究結果では、糖刺激は生得的な放出機構ではなく、調節性であり、行動の開始を時間的に制御していることが示されている8。このパラダイムを用いて、ミツバチ(Apis mellifera)7,8のこの行動についても研究しています。
本研究の最終的な目的は、標的遺伝子操作とニューロイメージング技術により、検索行動の制御に関与する神経回路と新しい遺伝的構成要素を解明することです。食物探索行動は、昆虫のナビゲーションと空間記憶を研究するための非常に効果的な実験パラダイムであることが証明されています。これらの行動は、ハエの報酬となる食料源の探索に関与する感覚知覚、意思決定プロセス、および運動協調を調査するユニークな機会を提供します。さらに、これらの研究から得られた知見は、多くの基本的な遺伝的および神経メカニズムが進化的に保存されているため、ヒトを含む他の生物の摂食行動を理解する上でより広い意味を持ちます。摂食行動の調節不全は、さまざまな神経障害や代謝障害と関連しています10。したがって、ハエの探索行動の根底にある神経および遺伝的メカニズムは、これらの複雑なヒトの健康課題を理解し、対処するための新しい道を提供する可能性があります。
プロトコル
ショウ ジョウバエ(Drosophila melanogaster)カントン-S (CS)野生型株の成虫オスのハエを本研究に使用しました。
1. 実験準備
- ハエの飼育
- 12時間以内に出現する成虫のオスのハエを捕集し、標準的なハエ培地(社内で調製)で25°C、相対湿度75%、12時間の明暗サイクルで48時間維持します。
注:使用したショウ ジョウバエ 培地組成物は、(培地1 Lに対して)トウモロコシ粉(80 g)、D-グルコース(20 g)、砂糖(40 g)、寒天(8 g)、酵母粉末(15 g)、プロピオン酸(4 mL)、4-ヒドロキシ安息香酸メチル(3 mLのエタノール中1.25 g)、およびオルトリン酸(600 μL)でした( 材料表を参照)。最初の給餌期間は、ハエが飢餓段階の前に十分な餌と栄養素にアクセスできるようにします。オスとメスの両方のハエが探索行動を誘発しますが、飢餓時間はオスのハエの方が一貫しています。さらに、雌のハエは交尾後に摂食の好みを変える11.
- 12時間以内に出現する成虫のオスのハエを捕集し、標準的なハエ培地(社内で調製)で25°C、相対湿度75%、12時間の明暗サイクルで48時間維持します。
- 飢餓の手順
- 給餌期間の後、ハエの餌を飢えさせますが、水にアクセスできます。
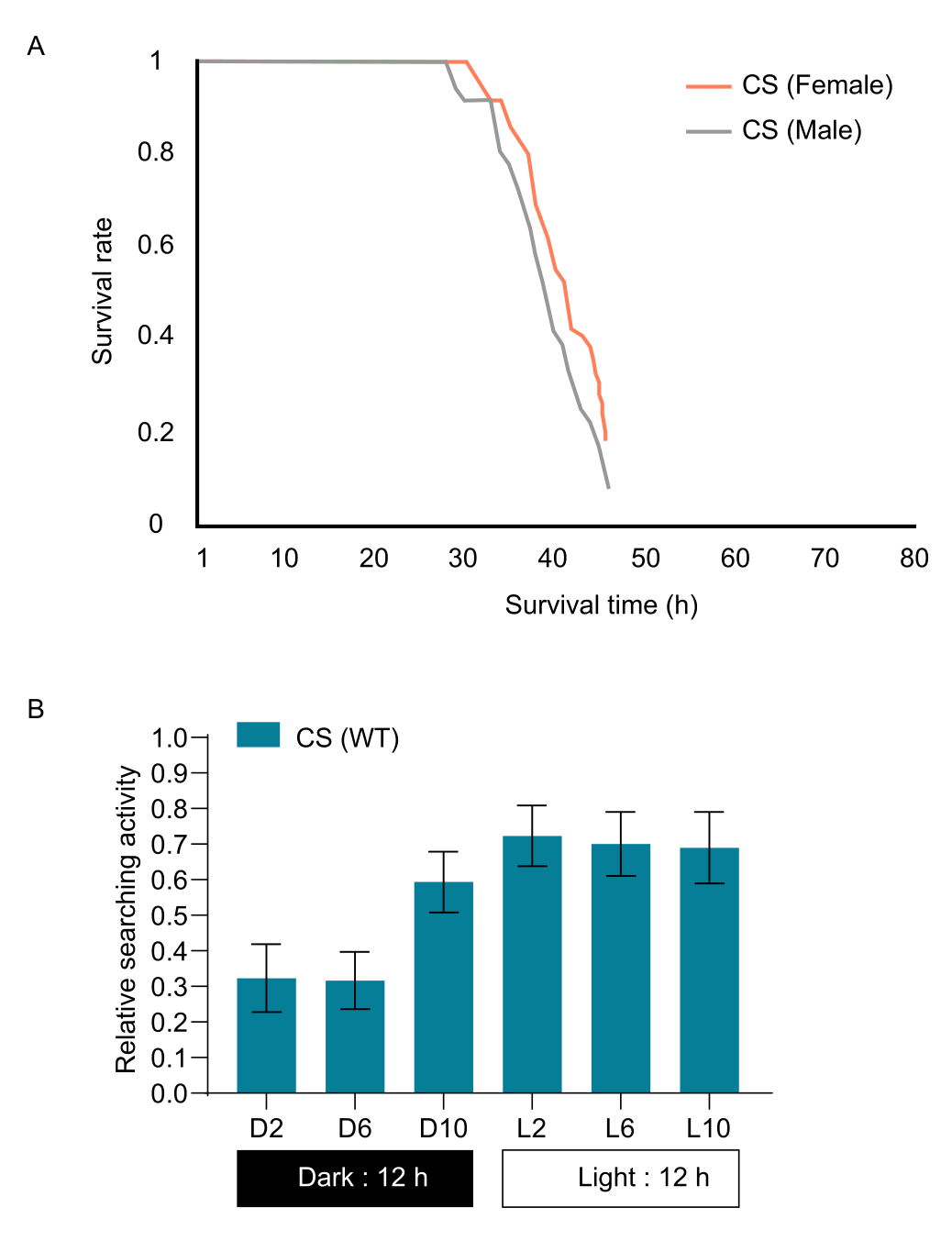
注:試行と系統間で飢餓状態を標準化するために、食料飢餓条件下での人口の90%の生存期間を決定することが提案されています。この結果に基づいて、CSバエは概日リズムやその他の要因が検索行動に影響を与える可能性があるため、28時間飢餓状態にあった(図1A)。また、一日のさまざまな時間帯におけるハエの探索活動の概日変化も測定されました(図1B)。その結果、ハエの光期と比較して、夜間の相対的な探索活動が減少することに気づきました。 - 生後2日のハエから餌を奪うことで、食物飢餓耐性を計算します。底にティッシュペーパーを浸したバイアルに15〜20匹のハエを入れます。これは基質として機能し、ハエが飢餓期間を通して水にアクセスできるようにします。
- 食物飢餓期間中のハエの数を1時間の一定の間隔で数えます。飢餓状態のハエの90%が生存する期間を飢餓期間として使用しました(図1A)。複数の反復(菌株ごとに3〜4回)を使用して、個々の変動の影響を最小限に抑え、飢餓耐性のより信頼性の高い評価を提供します。
注:各菌株の食物飢餓耐性は、異なる菌株と実験の間で標準化された飢餓状態を確立するために決定されました。これらの間隔で生き残ったハエを数えることで、時間の経過とともに生存率を監視し、各株が飢餓に屈するまでの食物欠乏にどれだけの期間耐えられるかを判断できます。
- 給餌期間の後、ハエの餌を飢えさせますが、水にアクセスできます。
- 行動を記録する手順
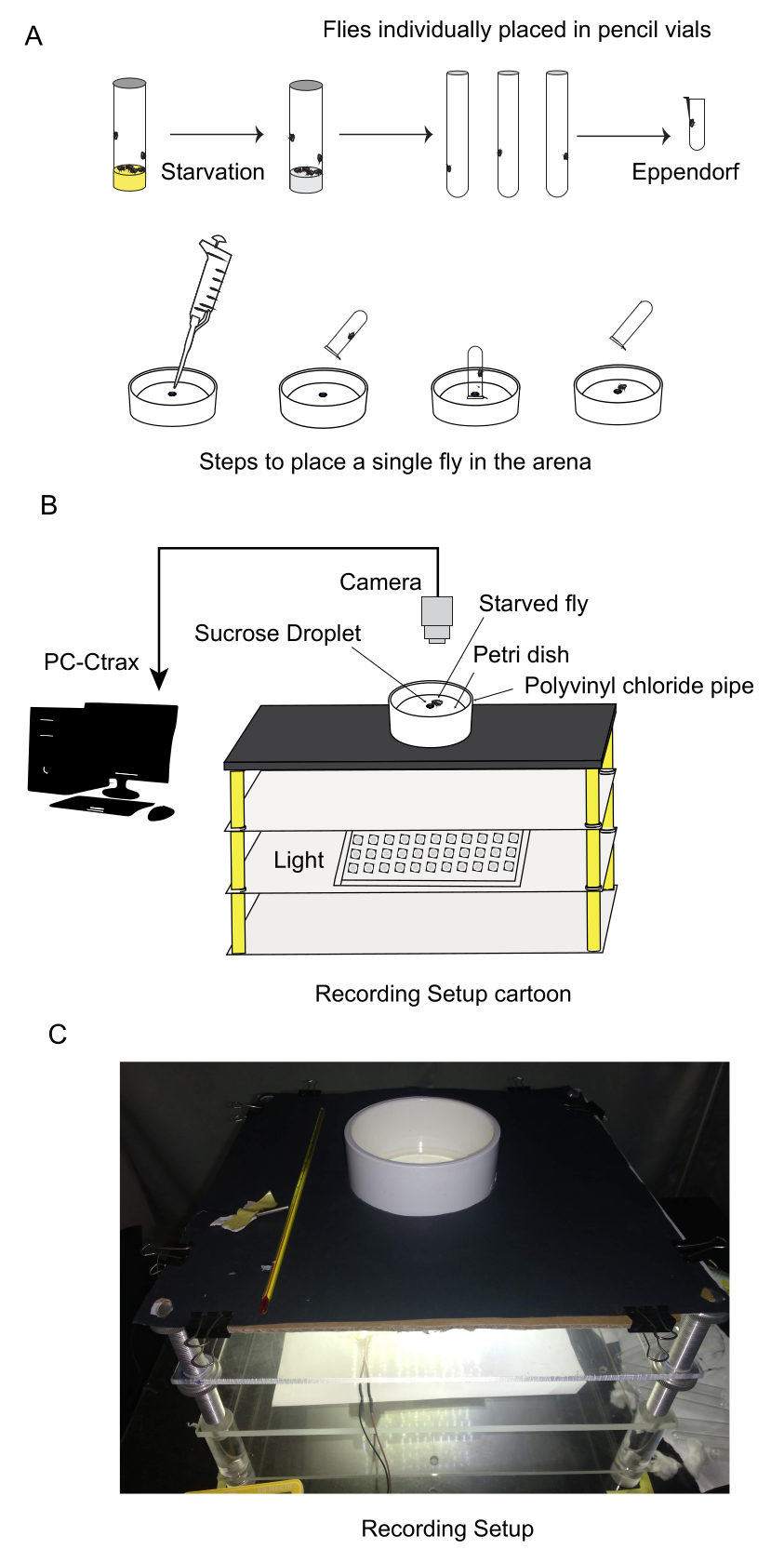
- 飢餓状態の個々のハエを小さなチューブに移します(図2A)。これを5〜6匹のハエのバッチで行い、分離時間を最小限に抑えることができます。1匹のハエをテストし、それぞれのハエを分離することで、観察された変化や行動は、他のハエとの相互作用の影響を受けることなく、観察されている特定のハエに起因する可能性があります。
- 90 mmシャーレを行動アッセイのアリーナとして使用します(より大きなシャーレを使用できます。大口径のシャーレでは挙動に差はなかった)。表面実装されたクールホワイトLEDのパネルでアリーナを下から照らします(図2B、C)。均一な視覚環境を維持し、外部からの気晴らしを最小限に抑えるために、実験場を白いポリ塩化ビニルパイプ(高さ51.5mm、内径114mm)で囲みます。
注:このパイプはバリアとして機能し、アリーナの外からの視覚刺激がハエの行動に影響を与えるのを防ぎます。外部からの気を散らすものを減らすことで、ハエと餌源との相互作用のみに集中し、実験全体を通して一貫性を保つことができます。 - アリーナの中央で320ルクスの光強度を使用します。
- 0.2 μLの糖溶液をアリーナの中央に配置します。報告された実験では500 mMのスクロース溶液が使用されましたが、これは変更することができます。2mLのマイクロ遠心チューブ(内径8.7mm、底に綿を詰めて長さを5mmに減らした。 図2A)シュガードロップの上に逆さまにされた1匹のハエを収容します。
- ハエが飛沫を摂取し始めたら、ハエの容器を取り外して、ハエが餌源に無制限にアクセスできるようにします。
- オーバーヘッドカメラでアリーナの2D位置を撮影します。
注:Flea3(ポイントグレー、1214 mmレンズ、 材料表を参照)を本研究に使用し、40フレーム/秒(fps)で記録しました。ただし、背景とのコントラストが良好なカメラであれば、どれでも使用できます。実験の性質に応じて、30〜60fpsで記録できます。追跡ソフトウェアと互換性があるため、.avi形式で記録します。 - ハエがアリーナから逃げ出すまでの裁判を記録します。ハエは自由に歩き回り、アリーナには蓋がありませんでした。
- ハエが探すのをやめるか、飛び去るか、アリーナの周辺まで歩くかを判断できるようにすることで、ハエの自然な行動と餌探しの戦略を観察します。試験の合間に70%エタノールでシャーレを拭き取り、完全に乾かすか、新しいシャーレを使用してください。
注:ハエが一貫して高い活動レベルを示すライトが点灯してから2時間から6時間の間にすべての実験を行うことが重要です。この時間枠により、ハエが活動状態にあることが保証され、ハエの自然な餌探し行動を観察する可能性が最大化され、概日リズムなど、ハエの行動に影響を与える可能性のある他の要因の影響が軽減されます。行動のセットアップは、温度と湿度が制御された部屋の中に収容されました。アリーナは振動のないテーブルの上に置かれました。この実験では、個々のハエを分離し、糖に対する反応を試験しました。飢餓状態のハエに砂糖を滴し、その行動をビデオで記録しました(図2B、C、ビデオ1)。実験は、ハエが一貫して高い活動レベルを示す特定の時間枠で行われました。
- 手順 2 に続く軌跡を分析して、検索動作を決定します。
2. 局所探索のための軌跡の解析
- Ctraxソフトウェア11 を使用して録画したビデオを分析します( 材料表を参照)。
注:ソフトウェアは、ビデオ内のハエの位置を追跡してx、y座標に変換し、その動きの正確な追跡と分析を可能にします。Ctraxの使い方については、 補足ファイル1 を参照してください。 - 軌跡を初期給餌段階と探索段階の2つの段階に分けます。給餌の終わりと歩行の開始を、3 つの連続したフレームで >4 mm s-1 の速度で移動するハエとして定義します。
注:給餌が終わった後、軌道の残りの部分がハエの捜索応答として使用されました。この研究では、VirtualDub( 資料表を参照)を使用して、追跡前のビデオから給餌フェーズを削除しました。 - 次のパラメーターで検索を定量化します。
- 経路の長さ: このパラメータは、餌の検索中にハエが開始点から歩いた距離(mm単位)を表します。
- 滞在時間:捜索中にハエが歩いた時間(単位:秒)。これは、検索の期間と、餌を探す際のハエの持続性を示します。
- 蛇行: パスのビーライン(パスの最初と最後のポイントの間の距離)をパスの全長に除算し、1から減算して比率として計算します。蛇行の値が高いほど、軌跡の曲がりくねりが大きいことを示します。
- リターン数:開発されたアルゴリズムを使用して、2つの同心円を使用してリターン数を識別してカウントします。
注:内側の円は検索の起点を示すRin (2.5 mm)と外側の円は、フライが原点から離れる必要があった最小距離Rout(4 mm)を示します。リターンは、外側の円(Rout)から出て、内側の円(Rin)に戻ることと定義されました。 - 活動率:フライの歩行速度が2mm/sを超える餌探しの時間を求め、活動率をパーセンテージで表します。このパラメータは、ハエの活動レベルと餌探しへの関与を反映しています。活動的な採餌者とそうでない個体を区別します。
注:本研究では、軌跡のさらなる解析のためにMATLABとPythonを使用しました。スクリプトには、https://github.com/eagermeagre/sugar_elicited_search からアクセスできます。分析手順では、Ctraxソフトウェアを使用してハエの動きを追跡し、餌の検索時間、経路の総長、検索時間、蛇行、リターンの回数、活動率などのパラメータを決定します。これらのパラメータは、記録された餌探し実験中のハエの行動と採餌効率に関する貴重な洞察を提供します。
代表的な結果
ハエは食物飢餓耐性によって推定された期間飢餓状態にする必要があり、砂糖に対する反応は個別にテストされました(図1A および 図2A)。この行動は、温度と湿度が管理された室内で記録されました。報告された実験には、0.2 μL の 500 mM スクロース溶液を使用しました。シュガードロップをアリーナの中央に配置し、ハエをシュガーに導入しました(図2B)。この行動は、ハエがアリーナから逃げ出すまで記録されました。ビデオを分析して、ハエの X、Y 座標と軌跡を抽出しました。行動反応を定量化するために、経路の長さ、滞在時間、蛇行、リターン数、活動率など、いくつかのパラメータを使用しました。
飢餓状態のハエの糖分摂取は、蛇行する経路とループによる局所的な探索につながります(図3A、C)。ネガティブコントロールとして、砂糖を与えられなかった飢餓状態のハエが記録された8。これらのハエは、アリーナに導入され、空になったとき、捜索を開始せず、アリーナから逃げ出しました(図3B、D)。このグループは、餌のないハエと呼ばれます。検索のパラメータである経路の長さ、滞在時間、蛇行、リターン回数は、糖分を滴下したハエと比較して、給餌されていないハエで有意に低かった(図4A-D)。

図1:野生型株 CSの飢餓曲線。 (A)野生型 CS の雄と雌の生存率を示す飢餓曲線。(B)一日の異なる時間帯におけるハエの捜索活動の概日変化。エラーバーは S.E.M. (n = 10) を表します。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図2:飢餓耐性の計算、行動記録手順、軌跡解析などの実験装置。 (A)標準化された飢餓状態を確立するために、食物飢餓耐性が決定された。行動記録のために、実験前に個々のハエを小さなチューブに隔離しました。行動アリーナである90mmのシャーレは、下から均一に照らされました。中央に一滴の砂糖溶液を置き、ハエは食料源に無制限にアクセスできました。(B)ハエがアリーナから逃げ出すまで、行動が記録されました。ビデオ解析では、トラッキングにCtraxソフトウェアを使用し、軌跡解析にMATLAB/Pythonを使用しました。(C)実験装置の写真。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図3:局所探索開始のための糖摂取の必要性 (A)500 mM、0.2 μLの糖溶液を給餌したハエの個々の軌跡。(B)糖分を与えられなかったハエの個々の軌跡。(C)検索軌跡のオーバーレイは、対照群(n = 11)から飛んでいます。(D)糖を与えられなかったハエ(n = 11)の経路の重ね合わせ。すべての軌跡は、歩行の開始点に正規化されます。この図は、Shakeel and Brockmann8 から引用したものです。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図4:餌を与えていないハエの行動パラメータの減少。 (A-D) 経路の長さ、滞在時間、蛇行、帰還回数は、糖分で刺激された対照ハエと比較して、糖報酬を与えなかった空腹のハエの方が小さかった。**p < 0.001、***p < 0.0001、****p < 0.00001、ウィルコクソン順位和検定。この図は、Shakeel and Brockmann8 から引用したものです。この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
動画1:ハエの探索行動と経路のリアルタイム軌跡このビデオをダウンロードするには、ここをクリックしてください。
補足ファイル1:CTRAXを使用してビデオファイルを追跡するための段階的な手順。このファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
本研究は、Dethier2によって最初に記述されたショウジョウバエの糖誘発性探索行動を調査するための簡単なパラダイムを導入しています。この生来の行動により、ハエは餌の報酬に遭遇した後、追加の食料資源を求めて局所的に探索することができます。実験プロトコルの最も重要な側面は、ハエを適切に動機づけることです。まず、ハエは空腹の状態にあり、水にアクセスできるにもかかわらず餌を奪われ、砂糖の摂取を確実にする必要があります。実験的試行全体で飢餓状態を均一にするために、人口の90%が生き残る期間を飢餓期間として使用しました。重要なことは、給餌後の検索反応の誘導には、十分な質の食物刺激を提供する必要があるが、ハエを完全に満腹させるには十分ではない。したがって、糖の濃度と量、および飢餓期間を標準化することは時間がかかるかもしれませんが、堅牢で信頼性の高い行動のために不可欠です。
この研究では、500 mM、0.2 μLのスクロース溶液を飢餓状態のハエの刺激として使用しました。砂糖の摂取は、旋回行動の増加と砂糖滴の位置への頻繁な帰還を特徴とする特徴的な局所探索行動を誘発します(ビデオ1)。逆に、砂糖を与えられていない空腹のハエは、検索応答を示さない。注目すべきは、経路の長さ、滞在時間、蛇行、リターンの回数など、行動に関連するすべてのパラメータが、餌を食べていないハエで有意に低かったことです。我々は以前、水の摂取だけでは検索応答を引き出さないことを実証した9。
このセットアップは、この生来の行動を研究するための費用対効果が高く、メンテナンスの少ないアプローチを提供します。この研究ではバックライト付きアリーナを使用していますが、ハエと背景の間に十分なコントラストがある限り、トップライトも使用できます。使用される追跡ソフトウェアは、静的な背景13に対するハエの動きの検出に依存している。カメラと解像度の設定は、調査中の特定の行動の規模に基づいて調整できます。重要なことは、この方法論により、採餌中の感覚的手がかりの注意、餌の関与と摂食、探索の自発運動制御、搾取と探索に関連する意思決定プロセスなど、採餌行動のさまざまな要素の研究が可能になることです。さらに、このパラダイムは、さまざまな生態学的文脈で多様な分類群に共通して観察される行動である局所検索の調査を容易にします6。ショウジョウバエのこの行動を研究することで、採餌に関与する神経経路を理解することを目的とした科学的調査への道が開かれます。我々はミツバチの局所探索を研究し、その行動がハエに似ていることを示した7,8。
最近の研究では、ハエの様々な糖感覚ニューロンの光遺伝学的活性化によって局所探索行動が引き起こされることが実証されている14,15,16。しかし、これらの研究で観察された局所的な検索が、実際の砂糖摂取に反応するハエの自然な行動をどの程度正確に表しているかは不明のままである。ハエの摂食行動は厳密に制御されており、これらの知見は、咽頭糖受容体の活性化が探索行動を開始することを示している。足根味覚は糖を検出して口吻伸展反射を誘発する役割を担っており、咽頭味覚ニューロンは摂食を進めるべきかどうかを決定します17,18。一旦摂取されると、糖溶液は食道を通って前腹部に移動し、その膨張が反回神経によって監視された状態で作物に入る19。さらに、前述の研究のいくつかはハエの利用または閉じ込めに関係していましたが、この方法では実験中、動物が自由に歩くことができることは注目に値します。私たちの実験のハエは、蓋をせずにアリーナ内にとどまり、探索するのに十分な動機を持っていました。
ハエの探索行動を支配する神経経路、遺伝的要因、環境的手がかりの間の複雑な相互作用を理解することで、情報処理、学習、記憶形成の基本原理を明らかにすることができます。さらに、採餌行動の調節不全は、摂食障害や肥満など、さまざまなヒトの障害に関与しています。ショウジョウバエで利用可能な広範な神経遺伝学的ツールは、糖が誘発する探索行動を調査し、採餌の根底にある神経および遺伝的メカニズムを解明するための貴重なリソースを提供します。このパラダイムは、光遺伝学的操作および機能イメージングと組み合わせることで、強力で有望なアプローチを提示します20,21,22。しかし、オプトジェネティクスを用いてニューロンの活動をリアルタイムに操作するためのセットアップを変更することは、困難な場合があります。ハエが探索行動を行っている間に脳内のニューロン活動を監視するには、トレッドボールにつながれたハエなど、別のセットアップが必要になります。摂食調節や意思決定プロセスなど、採餌行動の多くの側面は、種を超えて高度に保存されています。したがって、ハエの採餌の神経メカニズムを研究することで得られた知見は、ヒトを含む他の生物における同様のプロセスに関する貴重な知見を提供することができます。
開示事項
著者は、この記事の研究、著者、および/または出版に関して潜在的な利益相反がないことを宣言します。
謝辞
アリーナの設営を手伝ってくれたラヴィクマール・ボヤパティに感謝します。この研究は、ウェルカムトラストDBTインターミディエイトインドアライアンス助成金(助成金番号IA / I / 15/2 / 502074)によって資金提供されており、P.K. M.S.はインド医学研究評議会(ICMR)のフェローシップによって資金提供されました。ABは、NCBS-TIFR機関基金(No.12P4167)およびインド政府原子力省(No.12-R&D-TFR-5.04-0800および12-R&D-TFR-5.04-0900)。
資料
| Name | Company | Catalog Number | Comments |
| 2 mL Eppendorf tube | Sigma Aldrich | BR780546 | Used to introduce the fly to the sugar drop |
| Agar | SRL | 9002-18-0 | |
| Azure lens | https://www.rmaelectronics.com/azure-photonics-azure-1214mm/ | ||
| Camera | Logicool, Japan | ||
| Corn flour | locally available | ||
| Ctrax software | https://ctrax.sourceforge.net/ | ||
| D-glucose | SRL | 50-99-7 | |
| Flea3 | Sony | https://www.flir.com/products/flea3-usb3/?vertical=machine+vision&segment=iis | |
| glass tube | Borosil | Used to house the flies individually | |
| Kimwipe | Kimberly-Clark | 34155 | Used to provide access to water for flies during food starvation |
| LED light panel | custom-made in the workshop | ||
| Light Meter | TENMARS | TM-203 | |
| Methyl 4-hydroxybenzoate | Fisher Scientific | 99-76-3 | |
| Orthophosphotic acid | SRL | 7664-38-2 | |
| Petri dish (90 mm) | Tarsons | 460090 | |
| Propionic acid | SRL | 79-09-4 | |
| Sucrose | Qualigens | Q28105 | |
| Sugar | locally available | ||
| VirtualDub | https://www.virtualdub.org/ | ||
| White polyvinyl chloride pipe (67 mm inner diameter × 100 mm height) | custom-made in the workshop | ||
| Yeast powder | SRL | REF-34266 |
参考文献
- Jander, R. Ecological aspects of spatial orientation. Annu Rev Ecol Evol Syst. 6 (1), 171-188 (1975).
- Dethier, V. G. Communication by insects: Physiology of dancing. Science. 125 (3243), 331-336 (1957).
- White, J., Tobin, T. R., Bell, W. J. Local search in the housefly Musca domestica after feeding on sucrose. J. Insect Physiol. 30 (6), 477-487 (1984).
- Bell, W. J., Cathy, T., Roggero, R. J., Kipp, L. R., Tobin, T. R. Sucrose stimulated searching behaviour of Drosophila melanogaster in a uniform habitat: modulation by period of deprivation. Animal Behav. 33, 436-448 (1985).
- Dethier, V. G. Microscopic Brains. Science. 143 (3611), 1138-1145 (1964).
- Bell, W. J. Searching behavior patterns in insects. Annu Rev Entomol. 35 (1), 447-467 (1990).
- Brockmann, A., et al. Sugar intake elicits intelligent searching behavior in flies and honey bees. Front Behav Neurosci. 12, 280 (2018).
- Shakeel, M., Brockmann, A. Temporal effects of sugar intake on fly local search and honey bee dance behaviour. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. , (2023).
- Murata, S., Brockmann, A., Tanimura, T. Pharyngeal stimulation with sugar triggers local searching behavior in Drosophila. J Exp Biol. 220 (Pt 8), 3231-3237 (2017).
- Nishijo, H., Ono, T. Neural mechanisms of feeding behavior and its disorders new insights into metabolic syndrome. IntechOpen. , (2021).
- Carvalho, G. B., Kapahi, P., Anderson, D. J., Benzer, S. Allocrine modulation of feeding behavior by the sex peptide of Drosophila. Curr Biol. 16 (7), 692-696 (2006).
- Xu, K., Zheng, X., Sehgal, A. Regulation of feeding and metabolism by neuronal and peripheral clocks in Drosophila. Cell Metab. 8 (4), 289-300 (2008).
- Branson, K., Robie, A. A., Bender, J., Perona, P., Dickinson, M. H. High-throughput ethomics in large groups of Drosophila. Nat Methods. 6 (6), 451-457 (2009).
- Corfas, R. A., Sharma, T., Dickinson, M. H. Diverse food-sensing neurons trigger idiothetic local search in Drosophila. Curr Biol. 29 (10), 1660-1668.e4 (2019).
- Behbahani, A. H., Palmer, E. H., Corfas, R. A., Dickinson, M. H. Drosophila re-zero their path integrator at the center of a fictive food patch. Curr Biol. 31 (20), 4534-4546.e5 (2021).
- Titova, A. V., et al. Displacement experiments provide evidence for path integration in Drosophila. J Exp Biol. 226 (12), jeb245289 (2023).
- Stocker, R. F. The organization of the chemosensory system in Drosophila melanogaster: a review. Cell Tissue Res. 275 (1), 3-26 (1994).
- LeDue, E. E., Chen, Y. C., Jung, A. Y., Dahanukar, A., Gordon, M. D. Pharyngeal sense organs drive robust sugar consumption in Drosophila. Nat Commun. 6, 6667 (2015).
- Gelperin, A. Abdominal sensory neurons providing negative feedback to the feeding behavior of the blowfly. Zeitschrift für vergleichende Physiologie. 72 (1), 17-31 (1971).
- Simpson, J. H., Looger, L. L. Functional imaging and optogenetics in Drosophila. Genetics. 208 (4), 1291-1309 (2018).
- DeAngelis, B. D., Zavatone-Veth, J. A., Gonzalez-Suarez, A. D., Clark, D. A. Spatiotemporally precise optogenetic activation of sensory neurons in freely walking Drosophila. Elife. 9, e54183 (2020).
- Grover, D., Katsuki, T., Li, J., Dawkins, T. J., Greenspan, R. J. Imaging brain activity during complex social behaviors in Drosophila with Flyception2. Nat Commun. 11 (1), 623 (2020).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved