쥐 장 선와에서 오가노이드와 오가노이드 연구를 위한 단일 줄기 세포의 3D 배양
요약
우리는 쥐의 소장 선와와 배양 장 3D 오가노이드를 선와에서 분리하는 프로토콜을 설명합니다. 또한, 우리는 상피하 세포 틈새가 없는 단일 장 줄기 세포에서 오가노이드를 생성하는 방법을 설명합니다.
초록
현재 오가노이드 배양은 다양한 장기의 다양한 생물학적 측면과 질병에 대한 시험관 내 연구를 위한 중요한 도구입니다. 쥐 소장 선와(small intestinal crypt)는 3D 세포외 기질(extracellular matrix)에서 배양될 때 장 상피를 모방하는 오가노이드(organoid)를 형성할 수 있습니다. 오가노이드는 다양한 장 항상성 기능을 수행하는 모든 세포 유형으로 구성됩니다. 여기에는 Paneth 세포, 장 내분비 세포, 장 세포, 잔 세포 및 술 세포가 포함됩니다. 잘 특성화된 분자를 배양 배지에 첨가하여 G 단백질 결합 수용체 5를 포함하는 류신이 풍부한 반복으로 표지된 장 줄기 세포(ISC)를 풍부하게 하고 특정 계통을 따라 분화를 유도하는 데 사용됩니다. 이러한 분자에는 표피 성장 인자, Noggin(뼈 형태 형성 단백질) 및 R-spondin 1이 포함됩니다. 또한 단일 에리스로포이에틴 생성 간세포 수용체 B2(EphB2) 양성 ISC에서 오가노이드를 생성하는 프로토콜도 자세히 설명되어 있습니다. 이 방법 기사에서는 조직에서 소장 선와와 단일 ISC를 분리하고 오가노이드의 효율적인 확립을 보장하는 기술에 대해 설명합니다.
서문
2009년에 처음 설립된 장 오가노이드는 성숙한 조직과의 형태학적 및 기능적 유사성을 감안할 때 장 생물학을 연구하기 위한 강력한 체외 도구로 부상했습니다. 최근 성체 조직 줄기 세포에서 유래한 배양 오가노이드의 기술 발전으로 자가 재생 및 분화 가능성이 있는 장 줄기 세포(ISC)의 장기 배양이 가능해졌습니다. 이러한 오가노이드는 위장 생리학 및 병태생리학 1,2,3,4,5,6에 대한 기초 및 중개 연구에 널리 사용되었습니다. Clevers 그룹이 개발한 3D 오가노이드는 생리학적 관련성이 향상된 장 상피를 연구할 수 있는 강력한 도구를 제공합니다7. 장 오가노이드는 조직 줄기 세포에서 유래하고 여러 세포 유형으로 구성되어 있기 때문에 장 상피의 기능을 요약합니다. 주목할 점은, 단일 분류된 류신이 풍부한 반복 물질 함유 G 단백질 결합 수용체 5-양성(Lgr5+) 줄기 세포는 또한 Paneth 세포 또는 상피 틈새 또는 기질 틈새와 같은 ISC 틈새 없이 3D 오가노이드를 생성할 수 있다는것이다 7. 그러나, 단일 분류된 Lgr5+ 세포의 오가노이드 형성 능력은 crypt 및 ISC-Paneth celldoublet의 오가노이드 형성 능력에 비해 낮다8.
점점 더 많은 연구에서 에틸렌 디아민 테트라 아세트산 (EDTA) 배양 또는 콜라게나 제 해리 방법이 상피의 풀림과 지하실의 방출을 유발한다는 사실이 밝혀졌습니다. 효소 해리는 선와(crypt)의 세포 상태에 영향을 미칠 수 있으므로 일반적으로 조직을 해리하기 위해 기계적 분리 방법이 사용됩니다. 기계적 소화는 빠른 기술이지만, 이 방법은 일관성 없는 선와(crypt) 수율 또는 낮은 세포 생존율과 관련이 있을 수 있다9. 그러므로, EDTA 처리 및 기계적 해리는 더 나은 crypt 수율을 생성하기 위해 결합될 수 있습니다. 이 기사에 제시된 방법론의 특징은 EDTA 킬레이트화 후 조직 조각의 격렬한 진탕을 사용하는 것입니다10. 격렬한 흔들림은 소장의 crypt-villus 복합체에서 crypt를 효율적으로 격리 할 수 있습니다. 수동 흔들림의 정도에 따라 분리가 결정됩니다. 따라서 복합체에서 지하실을 얻는 것은 이 분야의 실험자에게 중요합니다. 또한 적절한 기술은 융모 오염을 최소화하고 지하실의 수를 늘릴 수 있습니다.
따라서 쥐 유래 소장 오가노이드를 사용하는 이 실험 프로토콜은 해리를 위해 EDTA로 처리한 후 물리적 힘으로 지하실을 더 잘 분리할 수 있습니다. 에리스로포이에틴 생성 간세포 수용체 B2(EphB2)의 발현 패턴은 부분적으로 선와 환경을 반영하는 것으로 알려져 있습니다. 예를 들어, EphB2 양성 세포는 아래에서 위로 조직화된다11. EphB2 발현을 기초로 형광 활성화 세포 분류(FACS)를 수행하였고, 얻어진 세포를 EphB2high, EphB2med, EphB2low 및 EphB2neg의 4개 그룹으로 나누었다. 이어서, 야생형(WT) 마우스에서 단일 분류된 EphB2고 세포로부터의 오가노이드 성장이 입증되었다.
프로토콜
모든 마우스 실험은 산토리 동물 윤리 위원회(APRV000561)의 승인을 받았으며, 모든 동물은 실험 동물의 관리 및 사용에 대한 위원회 지침에 따라 유지되었습니다. Mus musculus (C57BL6/J)의 표준 WT 균주를 사용했습니다. 10주령부터 20주령까지의 수컷 및 암컷 마우스를 모두 사용하였다. 마우스를CO2 질식으로 안락사시켰다.
1. 소장의 격리
- 십이지장과 공장의 근위 절반을 포함한 소장을 실험실 가위로 절제하십시오.
- 조직을 페트리 접시에 옮기고 5mL 주사기에 차가운 PBS-ABx 5mL(PBS + 페니실린-스트렙토마이신[1%] + 겐타마이신[0.5%])로 소장을 씻어내어 내강 함량을 제거합니다.
- 실험실 가위로 조직을 세로로 자르고 흔들면서 차가운 PBS-ABx로 수동으로 씻으십시오.
알림: 슬라이드로 융모를 긁어내면 융모 오염을 줄일 수 있습니다12. - 실험실 가위를 사용하여 약 5mm x 5mm 크기의 장 부분을 수집합니다. 단편을 핀셋을 사용하여 50mL 튜브로 옮기고 차가운 PBS-ABx 25mL를 추가합니다.
- 25mL의 차가운 PBS-ABx로 앞뒤로 10배 교반하여 절편을 세척하여 50mL 튜브에서 장 내용물을 제거합니다.
2. 토굴 격리
- 조각을 2mM EDTA를 함유하는 PBS-ABx에서 흔들림 없이 얼음 위에서 30분 동안 배양합니다.
- 세포외 기질(ECM)의 용이한 응고를 위해, 24-웰 플레이트를 미리 37°C 조직 배양 배양기에서 배양한다.
- 진공 펌프로 세포 배양 시스템에서 EDTA 용액을 흡인하고 신선하고 차가운 PBS-ABx 25mL를 추가한 다음 조각을 손으로 30x-40x로 위아래로 세게 흔들어 crypt-villus 복합체를 방출합니다.
참고: 분리된 선와와 융모는 4배 배율로 현탁액에서 25μL 물방울을 현미경으로 관찰하여 확인할 수 있습니다. - 다음으로, 현탁액을 70μm 스트레이너를 통해 한 번 여과합니다.
- 현탁액을 4°C에서 3분 동안 390× g 에서 원심분리합니다.
- 소르비톨 DMEM 20mL(고급 DMEM/F12 + 페니실린-스트렙토마이신[1%] + 겐타마이신[0.5%] + 소 태아 혈청[1%] + 소르비톨[2%])에 크립트 펠릿을 다시 현탁하고 피펫팅으로 크립트 현탁액을 두 개의 새로운 15mL 튜브로 옮겨 두 개의 10mL 용액으로 나누어 저속으로 원심분리합니다.
알림: 큰 세포 덩어리와 세포/파편은 저속 원심분리를 사용하여 분리할 수 있습니다. 큰 세포 덩어리는 펠릿에 있고 세포/파편은 상청액에 있습니다. - 2개의 crypt 현탁액을 80 × g 에서 4°C에서 3분 동안 원심분리한 다음, 상청액을 부드럽게 흡인한다.
알림: 펠릿 형성이 약하므로 너무 많이 흡인하지 마십시오. 각 튜브에 2mL의 상청액을 남겨 둡니다. - 10mL의 소르비톨 DMEM을 각 튜브에 다시 추가합니다. 현탁액을 80 × g 에서 4°C에서 3분 동안 원심분리한다.
- 상청액을 흡인하고, 각 튜브에 2 mL의 상청액을 남겨둔 후, 재현탁을 위해 10 mL의 소르비톨 DMEM을 첨가하고, 4°C에서 최종 3분 동안 80 × g 에서 크립트 현탁액을 원심분리한다.
- 상층액을 흡인 후 각 튜브에 상층액 2mL를 남기고 완전한 DMEM(고급 DMEM/F12 + 페니실린-스트렙토마이신[1%] + 겐타마이신[0.5%] + 소 태아 혈청[1%]) 위아래로 피펫팅하여 펠릿을 재현탁하고 1분 동안 그대로 둡니다.
알림: 떠 있는 지하실을 효율적으로 얻으려면 1분 동안 기다리십시오. - 1분 후 각 10mL 현탁액을 수집하여 총 20mL를 수집하고 70μm 세포 여과기로 한 번 여과하여 크립트를 정제합니다.
- 본질적으로 순수한 크립트를 파종하기 전에 여과된 완전한 DMEM에서 크립트 수를 세고, 4°C에서 3분 동안 290× g 에서 원심분리합니다.
- 25μL 물방울을 6cm 접시에 3개 지점에 떨어뜨립니다. 현미경으로 4x 배율로 선와를 세고 25μL 액적당 선와를 농도로 계산합니다.
- 웰당 40μL의 ECM으로 100개의 크립트를 현탁합니다. 피펫을 5x-10x 상하로 하여 ECM에서 크립트의 균질한 현탁액을 얻은 다음, 37°C로 예열된 24-웰 플레이트에서 시딩하였다.
알림: 중합을 피하기 위해 ECM을 항상 얼음 위에 두십시오. ECM에 기포가 생기지 않도록 조심스럽게 피펫팅합니다. - ECM의 중합을 위해 24-웰 플레이트를 37°C, 5%CO2 인큐베이터에서 15분 동안 배양한다.
- 마지막으로 실온에서 마우스 표피 성장 인자(EGF), 재조합 마우스 R-스폰딘 1 및 재조합 마우스 Noggin을 포함하는 배양액 500μL로 ECM을 덮습니다. 웰당 물질의 최종 농도는 다음과 같습니다: 페니실린-스트렙토마이신(1%), 각각 50U/mL; 겐타마이신(0.5%), 25μg/mL; EGF, 20ng/mL; 노긴, 100ng/mL; R-스폰딘 1, 500ng/mL; L- 글루타민, 2mM.
- 5%CO2 인큐베이터에서 37°C에서 크립트 배양을 시작한다.
참고: 24웰 플레이트의 오가노이드에 대한 배양 배지는 표 1을 참조하십시오. - 최대 7일 동안 3시간마다 20x 대물렌즈가 장착된 기록 타임랩스 이미지 현미경으로 오가노이드 형태 형성을 관찰하기 위해 장기 라이브 이미징을 수행합니다. 1 μm(1 μm x 5 스텝)의 z 스텝에서 연속 z 스택 이미지를 얻을 수 있습니다.
- 격일로 매체를 변경하십시오.
3. 형광 활성화 세포 분류(FACS)
- 생쥐에서 선와를 분리합니다(섹션 2 참조).
- 분리된 크립트를 37°C에서 30분 동안 2mL의 트립신으로 처리합니다.
- PBS 10 mL로 반응을 중지한 후 20 μm 세포 여과기를 통과시킨다.
- 용액을 4°C에서 3분 동안 390× g 에서 원심분리하고 100μL의 완전한 DMEM으로 재현탁합니다.
- 항-EphB2 APC 접합 항체(1/50)를 추가하고 얼음에서 30분 동안 배양합니다.
- 세포를 PBS로 3x 세척하고, 마지막으로 7-아미노-악티노마이신 D(7-AAD)(1/100)를 첨가한다.
- FACS를 통해 염색된 세포를 분류합니다.
- 영역 배율 인수를 조정하고 셀 크기(순방향 산란, FSC-A)와 입도(측면 산란, SSC-A)에 따라 정렬합니다.
- 488nm의 파장과 50mV 전력으로 설정된 레이저로 생존을 위해 7-AAD 음성 및 양성 세포를 분류합니다.
- 640 nm 및 100 mV 전력의 파장으로 설정된 레이저를 사용하여 EphB2-high(EphB2high), EphB2-medium(EphB2 med), EphB2-low(EphB2low) 및 EphB2-음성(EphB2neg) 세포를 분류하기 위해 게이트를 구분합니다.
- EphB2고 세포 배양을 5%CO2 배양기에서 37°C에서 시작한다.
4. 단세포 배양 오가노이드
- 등급이 매겨진 EphB2 표면 수준11에 따라 세포 분리 방법을 수행한 다음 4개의 별개의 집단(높음, 중간, 낮음 및 음성)을 얻습니다.
- 4°C에서 3분 동안 390× g 에서 원심분리하여 펠렛을 수집하고 피펫팅으로 ECM에 단일 분류된 EphB2high 세포를 삽입한 다음 24웰 플레이트(100 singlet/40μL ECM/well)에 파종합니다.
- 단계 2.14에서와 같이, ECM이 중합되도록 하고, EphB2높은 세포를 유지하기 위해 처음 2일 동안 Rho-associated kinase (ROCK) 억제제 (10 μM)를 함유하는 배양 배지로 ECM을 커버한다.
참고: ROCK 억제제는 anoikis에 효과적입니다. - 40x 배율의 도립 현미경을 사용하여 세포를 수동으로 검사하고 스페로이드 형성 및 선와돌출이 있는 생존 가능한 오가노이드를 관찰합니다.
대표적 결과
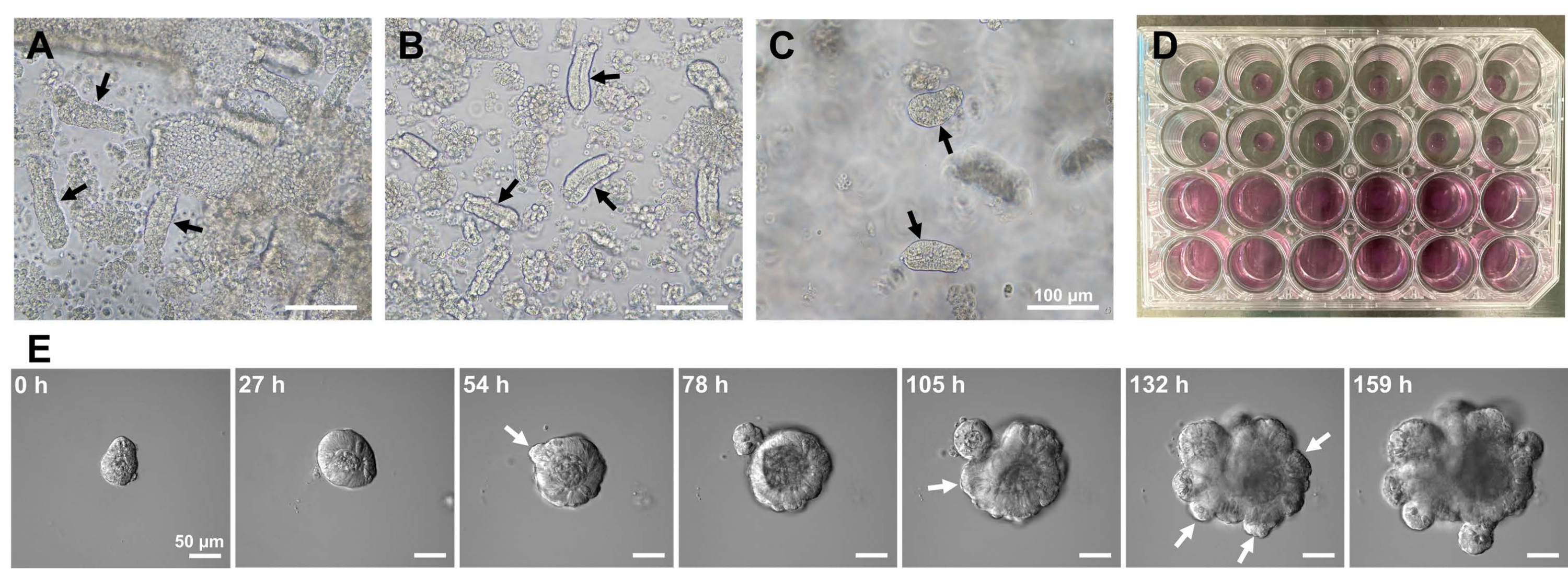
마우스 소장 오가노이드를 생성하기 위해, EDTA 처리와 기계적 격리 방법의 조합을 사용하여 크립드를 효율적으로 분리할 수 있다10,13. 이 연구의 결과는 거의 모든 격리된 선와가 즉시 봉인되어 상피 틈새에서 압착된 후 원뿔 모양으로 나타남을 보여주었습니다(그림 1A). 융모 오염을 최소화하기 위해, 생성된 현탁액을 70μm 세포 여과기에 통과시킨 다음, 여과액을 원심분리하였다. 일부 지하실은 여과 및 현탁 중에 중단되므로 이러한 단계를 신중하게 수행해야 합니다. 그 결과 최종 분획의 거의 모든 크립트가 통합되어 배양에 사용하기에 적합하다는 것을 보여주었습니다(그림 1B). 도금된 모든 크립트를 개별적으로 시각화하기 위해 웰당 100개의 크립트를 플레이팅했습니다(그림 1C). 특정 crypt 배양 배지를 첨가한 후(그림 1D), 오가노이드의 발달을 매일 현미경으로 모니터링했습니다. 또한, 선와로부터의 오가노이드 성장은 그들의 발달을 모니터링하기 위해 타임랩스 이미지로 관찰되었습니다(그림 1E 및 보충 비디오 S1). 교양 있는 지하실은 틀에 박힌 방식으로 행동했습니다. 오가노이드의 내부 내강은 아폽토시스 세포 덩어리로 채워졌습니다. ISC의 활성 증식 및 분화는 출아와 함께 선와(orbit) 영역에서 발생했습니다(그림 1E 및 보충 비디오 S1). 출아(毛國)는 ISC 이동 및 증식 및 파네스(Paneth) 세포 분화와 결합되었다. 분화된 파네스 세포는 항상 출아 부위에 위치하였다(보충도 S1). 오가노이드가 10x 배율에서 도립 현미경을 사용하여 배양에서 안정적인 것으로 확인됨에 따라 이 기술은 발달 중인 소장에서 선와 형성을 검사하고 새로운 장 상피 세포 생산을 위한 조직 재생 및 ISC 장기 생존 능력을 결정하는 데 사용할 수 있습니다14,15,16.
Lgr5는 ISC 마커로 정의되며, 쥐 Lgr5+ 세포는 3D 오가노이드를 형성한다7. 그러나 LGR5 단백질의 세포 표면 풍부도가 낮고 고친화성 항-LGR5 항체가 부족하기 때문에 FACS로 쥐 ISC를 효율적으로 분리하는 것이 어렵습니다. EphB2는 이전에 장 조직으로부터 쥐 및 인간 ISCs의 정제를 위한 표면 마커로서 확인되었다17,18. EphB2의 발현 패턴은 ISC 마커와 관련된 복잡성을 증가시킵니다. EphB2 양성 세포는 증식 구획 전체에 걸쳐 조직되어 있으며, 선와(orbit)의 바닥에서 정점을 찍는 반면, 선와(crypt)의 상단으로 갈수록 기울기가 감소한다(11). Paneth 세포와 전구 세포도 선와에 국한되어 있습니다. 파네스 세포는 주로 위치 지정에 필요한 EphB3를 발현하고, 선와에서 그 위의 전구 세포는 주로 EphB2를 발현합니다. 따라서, 두 세포 유형의 오염은 항-EphB2 항체를 사용하는 ISC 정제 과정 동안 발생할 수 있다. 따라서, 이들의 마커 유전자 발현 및 FACS에 의해 EphB2를 사용하여 분리된 세포의 오가노이드 형성 능력을 평가해야 한다.
이러한 사실에 기초하여, FACS 분석을 이용하여, EphB2 표면 표지된 세포는 WT 크립트로부터 분리될 수 있다19. EphB2 발현이 ISC 특이적 마커 유전자(Lgr5, Ascl2, Olfm4)와 전구세포 특이적 마커 유전자(Ki67, Myc, FoxM1)와 같은 특이적 마커의 발현으로 4개 군을 구별할 수 있는지 조사하였다. 이 실험은 EphB2상위 세포가 EphB2med 세포20,21과 달리 주로 ISC임을 입증했습니다. 마지막으로, 세포 분리 방법에 기초하여, 얻어진 세포를 4개의 그룹(EphB2high, EphB2med, EphB2low 및 EphB2neg cells)으로 나누었다(도 2). 이어서, 높은 수준의 EphB2를 발현하는 단일 세포를 FACS에 의해 분류하여 오가노이드 성장을 위해 배양하였다. 단일 EphB2고위 세포는 국소 치료에 독립적으로 적용될 수 있으며 정상 소장을 연상시키는 자가 조직화 섬와-융모 구조를 재현할 수 있습니다(그림 3). 그러나, 다른 그룹(EphB2med, EphB2낮음 및 EphB2neg)으로부터 유래된 세포는 오가노이드20을 생성하지 않는다.
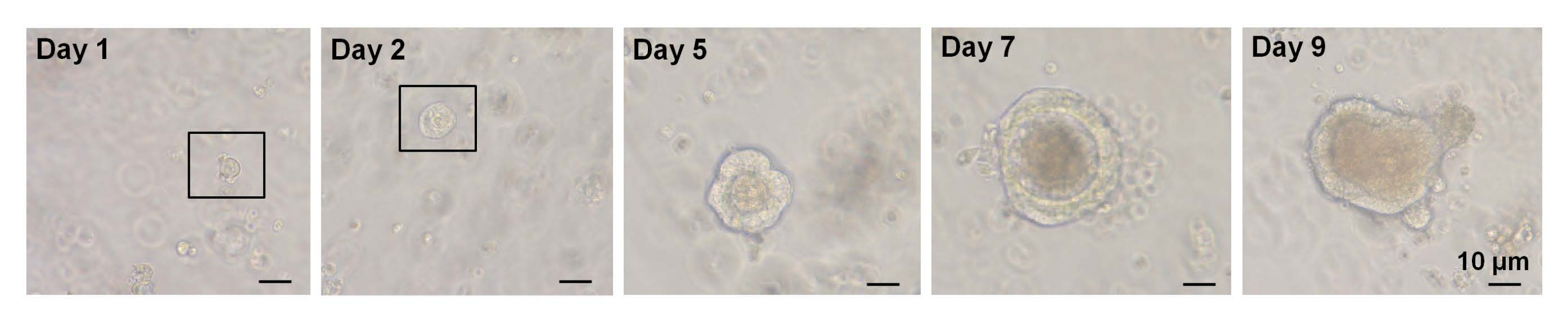
이전 연구에서, 단일 분류된 Lgr5-GFPhi 세포의 ~6%가 음와융모 오가노이드를 개시할 수 있었다7. 그러나 나머지 세포는 오가노이드를 생성할 수 없었고 처음 12시간이내에 사망했습니다. 저자는 이것이 격리 절차7에 내재된 신체적 및/또는 생물학적 스트레스의 결과라고 추정했다. WT 마우스의 단일 분류된 EphB2고 세포로부터 6% 미만의 오가노이드 성장이 또한 얻어졌다. 배양 5일째에 스페로이드와 같은 구조가 형성되었습니다(그림 3). 7일차부터 9일차까지 지하실을 형성하기 위한 반점의 대피가 발생했습니다(그림 3). 중요하게도, 선택된 ROCK 억제제를 단일 분류된 EphB2고 세포에 적용하면 해리 유도 세포자멸사가 감소하고 오가노이드 성장의 효율성이 증가한다는 것입니다.

그림 1: 마우스 소장 오가노이드의 생성. (A) EDTA 킬레이트화와 기계적 해리의 조합에 의해 제조된 크립트. (B) 결과 정제된 지하실. (C) 세포외 기질에 내장된 크립트. (ᄀ씨) 검은색 화살표는 지하실을 나타냅니다. (D) 토굴 및 오가노이드의 3차원 배양. (E) 지하실에서 파생된 성장하는 오가노이드의 대표 이미지. 흰색 화살표는 토굴 싹 트림을 나타냅니다. 스케일 바 = (A-C) 100 μm 및 (E) 50 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

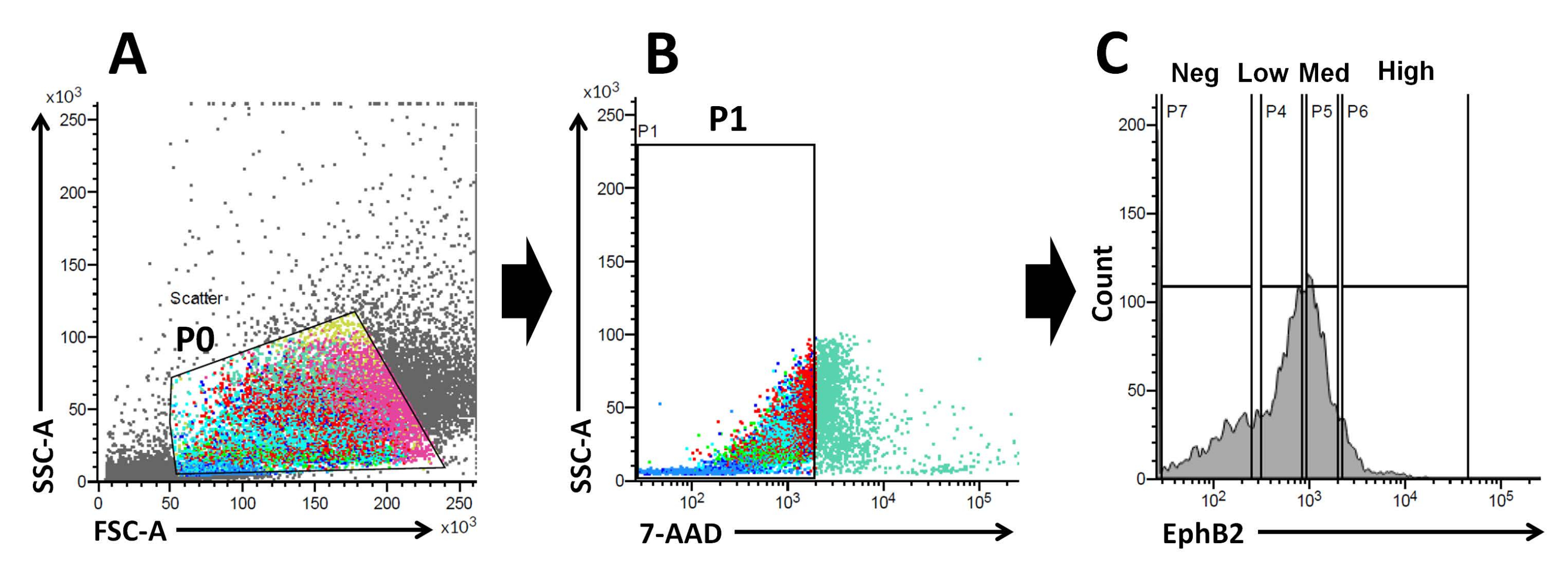
그림 2: 야생형 마우스에서 EphB2 양성(EphB2+) 세포 집단을 얻기 위한 유세포 분석 게이팅 전략. (A) 전방 및 측면 산점도는 각각 크기와 입도에 따라 세포를 분리하는 데 사용됩니다. (b) 형광 산란은 세포의 7-AAD(PerCP) 형광 강도에 따라 생존 세포를 분리하기 위해 사용된다. 7-AAD-음성 세포 집단에 대한 게이트가 선택되었습니다. (C) EphB2-high(EphB2high), EphB2-medium(EphB2med), EphB2-low(EphB2low) 및 EphB2-음성(EphB2neg) 세포 집단에 대한 게이트를 선택했습니다. 약어: FSC-A = 전방 산란 피크 면적; SSC-A = 측면 산란-피크 면적; 7-AAD = 7-아미노-악티노마이신 D. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 야생형 마우스에서 단일 정렬된 EphB2고세포 오가노이드 성장의 시간 경과. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
표 1: 24-웰 플레이트용 배양 배지. 이 표를 다운로드하려면 여기를 클릭하십시오.
보충 비디오 S1: 성장하는 오가노이드의 타임랩스 이미지. 스케일 바 = 50 μm. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S1: 오가노이드에서 항라이소자임 항체 염색의 대표 이미지. 흰색 화살표는 Paneth 셀을 나타냅니다. 약어: DIC = 미분 간섭 대비 현미경. 스케일 바 = 10 μm. 이 파일을 다운로드하려면 여기를 클릭하십시오.
토론
이 프로토콜은 소장 선와를 일관되게 분리하고 3D 오가노이드의 후속 배양을 설명하는 방법을 설명합니다. 지하실 방출 속도를 향상시키기 위해 EDTA 치료 후 격렬한 흔들림을 포함하는 기계적 격리 방법이 확립되었습니다. 배지 조성은 Sato et al.7의 원래 프로토콜과 다릅니다. 원래 매체는 상대적으로 비쌉니다. 따라서, 약리학적 억제제, 재조합 성장 인자 및/또는 컨디셔닝 배지를 함유하는 쥐 소장 오가노이드에 대한 배양 배지 및 맞춤형 배지가 표 1에 나와 있습니다. Wnt3A 및 N-아세틸시스테인은 이 프로토콜의 배양 배지에 포함되지 않습니다. Paneth 셀이 Wnt3를 발현함에 따라 셀은 Wnt3를 생성하고 ISC 유지를 지원합니다. 또한 크립트 격리 과정에서 컨디셔닝된 매체는 사용되지 않습니다. 오가노이드 모델은 동적이며 세포 및 구조적 이질성(파네스 세포, 장세포, 잔 세포, 장내분비 세포, 술 세포 및 ISC)을 가지고 있습니다. 따라서 이러한 오가노이드는 오가노이드 생물학의 근본적인 문제를 연구하는 데 대규모로 사용할 수 있습니다.
EphB2 구배는 성인 소장에서 섬와-융모 축을 따라 ISC 줄기 및 증식을 유지한다18. 분리된 선와와 비교하여 단일 EphB2 세포에서 오가노이드를 만드는 이점은 ISC가 다양한 인간 장 질환에서 핵심적인 역할을 하기 때문에 쥐 ISC의 생물학을 이해하는 것과 관련이 있습니다. 단일 EphB2 고발현 ISC는 단일 Lgr5발현 ISC에서 오가노이드를 개발하는 것과 유사한 방식으로 배양하여 오가노이드를 형성할 수 있습니다. 가장 중요한 단계는 FACS를 이용하여 정밀하게 세포들을 크립트에서의 EphB2 발현에 따라 4개의 그룹(EphB2high, EphB2 med, EphB2low, 및 EphB2neg)으로 나누는 것이다. 전방 대 측면 산란(FSC 대 SSC) 플롯은 일반적으로 크기와 입도에 따라 관심 있는 세포를 식별하는 데 사용됩니다. FSC는 셀 크기를 나타내고, SSC는 P0 게이트에서 셀의 복잡성 또는 세분성과 관련된다(도 2A). 이 작업에서, 정의된 게이트(P0) 내에 속하는 세포는 후속적으로 생존력에 대해 분석되었습니다. 다음으로, 이들의 생존력은 7-AAD 형광 신호의 음성 및 양성 집단에 따라 결정되었습니다. 7-AAD 음성 세포와 -양성 세포 사이의 경계는 최소한의 양성 세포 오염으로 음성 세포를 얻기 위해 엄격하게 결정되었습니다. EphB2 게이트는 EphB2 등급 발현에 기초하여 대략적으로 설정되었다.
4개의 군을 정밀하게 구분한 것을 확인하기 위해, 선별된 유전자의 mRNA 발현을 분석하였다. ISC 마커의 mRNA 수준은 EphB2세포 에서 높게 나타난다20. 또한, 전구 세포 특이적 마커의 mRNA 수준은 EphB2med 세포에서 상대적으로 높다20. 그러나, EphB2낮음 및 EphB2neg 세포에서의 EphB2 exdpression은 EphB2high 및 EphB2med 세포20에 비해 낮거나 음성이다. 도금 전에 EphB2고 세포 집단의 농축을 보장하기 위해 선행 조치를 취해야 합니다. 그러나, EphB2고 세포에서 6% 미만의 오가노이드 성장은 자와분리 동안 격렬한 진탕이 아니라 배양 과정 동안 줄기세포의 사멸에 기인할 수 있다. 인간 배아 줄기 세포에 선택적 Rho-associated kinase(ROCK) 억제제를 적용하면 해리 유도 세포자멸사가 현저히 감소하는 것으로 나타났습니다22. 따라서, 기술적 변화로서, 생존력을 향상시키기 위해 더 높은 농도와 더 긴 배양으로 ROCK 억제제를 첨가하는 것이 가치가 있다.
ISC 옆에 있는 Wnt3A-분비 파네스 셀은 ISC에 필수적인 지지를 제공한다(8). 실제로, ISC-Paneth 세포 이중선은 단일 ISC에 비해 크게 증가된 오가노이드 형성 능력을 나타낸다8. 또한, 배양 첫 3일 동안 100ng/mL 농도의 Wnt3A를 첨가하면 오가노이드 형성 능력이 증가하는 것으로 나타났다8. 따라서 또 다른 기술적 변화로 외인성 Wnt3A를 추가하면 단일 EphB2고발현 ISC의 오가노이드 형성 능력이 향상될 수 있습니다.
생체 내 접근법에 비해 오가노이드는 유전자 조작, 악성 표현형 분석 및 약물 스크리닝에 쉽게 사용할 수 있습니다20,23. EDTA 킬레이트화와 기계적 분리 방법의 조합은 선와에서 소장 오가노이드를 생성하는 데 효과적이고 재현 가능하며 시간 효율적이며 고급 경험이 없어도 실험실 직원이 쉽게 따를 수 있습니다. 따라서, EDTA로 치료한 후 격렬한 진탕을 통한 기계적 단리를 추가하면 생체 외에서 쥐 소장 오가노이드를 효율적으로 확립할 수 있고 다른 성인 상피 조직의 오가노이드 배양 및 질병 모델링을 위한 잠재적인 도구를 제공할 수 있습니다.
장 상피 세포는 분극화되고 정점 측면이 내강을 향하도록 배향됩니다. 그러나 3D 오가노이드의 루멘을 향한 정점 면은 내부에 있습니다. 따라서 이 조직은 영양분, 미생물 및 대사 산물과 같은 내강 성분이 상피 세포에 미치는 영향을 연구할 때 문제가 되는 정점 측면에 대한 접근을 방지합니다. 이러한 단점을 피하기 위해, 오가노이드 세포를 2D 단층으로 배양하는 방법이 개발되었다24. 미래의 응용 측면에서 오가노이드 세포 단층의 배양은 가장 효율적이고 다루기 쉬운 시스템을 나타내기 때문에 활용될 것입니다.
공개
저자는 선언할 이해 상충이 없습니다.
감사의 말
이 작업은 T.T.(보조금 번호 JP17K07495 및 JP20K06751)에 대한 과학 연구 보조금(C)의 지원을 받았습니다. 장기 타임랩스 이미징(LCV100; 올림푸스).
자료
| Name | Company | Catalog Number | Comments |
| 1.5 mL Eppendorf tube | Eppendorf | 0030 125.215 | |
| 5 mL syringe | TERUMO | SS-05SZ | |
| 15 mL Falcon tube | Iwaki | 2325-015 | |
| 20 μm cell strainer | Sysmex | 04-004-2325 | |
| 24-well plate | Iwaki | 3820-024 | |
| 50 mL Falcon tube | Iwaki | 2345-050 | |
| 60 mm tissue culture dish | FALCON | 353002 | |
| 70 μm cell strainer | Falcon | 352350 | |
| 100 mm Petri dish | Iwaki | 3020-100 | |
| 7-AAD | BD Biosciences | 559925 | |
| Advanced DMEM/F12 | Gibco | 12634-010 | |
| Alexa Fluor 568 Goat Anti-Mouse IgG (H+L) | Invitrogen | A-11004 | |
| Anti-EphB2 APC-conjugated antibody | BD Biosciences | 564699 | |
| C57BL6/J mice | Japan SLC, Inc. | ||
| Clean bench | HITACHI | CCV-1306E | |
| Confocal laser scanning microscope | Olympus | FV3000 | |
| EDTA (0.5 mol/L) | Nacalai Tesque | 06894-14 | 2 mM |
| FACSMelody | BD Life Sciences-Biosciences | 661762 | |
| Fetal bovine serum | Sigma | 173012 | 1% (v/v) |
| Fiji (software) | https://fiji.sc/ | ||
| Gentamicin (10 mg/mL) | Nacalai Tesque | 16672-04 | 25 μg/mL |
| Hammacher laboratory scissor | SANSYO | 91-1538 | |
| Incubator | Panasonic | MCO-170-PJ | |
| Laboratory tweezer | AS-ONE | 7-164-04 | |
| L-Glutamine 200 mM | Gibco | 25030081 | 2 mM |
| Matrigel | BD Biosciences | 354230 | ECM for 3D organoids |
| Mouse Anti-Human Lysozyme | LSBio | LS-B8704-100 | |
| Murine EGF (20 μg/mL stock solution) | PeproTech | 315-09 | 20 ng/mL |
| PBS 1x | Gibco | 10010-023 | |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140-122 | 50 U/mL |
| Pipetman (10 μL, 20 μL, 200 μL, and 1,000 μL) | GILSON | 1-6855-12, -13, -15, and -16 | |
| Recombinant murine Noggin (20 μg/mL stock solution | R&D Systems | 1967-NG-025 | 100 ng/mL |
| Recombinant murine R-Spondin 1 (250 μg/mL stock solution) | R&D Systems | 3474-RS-050 | 500 ng/mL |
| Sorbitol | Nacalai Tesque | 32021-95 | 2% (w/v) |
| TE2000-S (inverted microscope) | Nikon | 24131 | |
| Time-lapse image microscope | Olympus | LCV100 | |
| TrypLE Express 1x | Gibco | 12605-010 | |
| ULVAC | ULVAC KIKO Inc. | 100073 | |
| Y-27632 | Fujifilm | 331752-47-7 | 10 μM |
참고문헌
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Seidlitz, T., et al. Human gastric cancer modelling using organoids. Gut. 68 (2), 207-217 (2019).
- Nikolaev, M., et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature. 585 (7826), 574-578 (2020).
- Artegiani, B., Clevers, H. Use and application of 3D-organoid technology. Human Molecular Genetics. 27, R99-R107 (2018).
- Lancaster, M. A., Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 345 (6194), 1247125 (2014).
- Dedhia, P. H., Bertaux-Skeirik, N., Zavros, Y., Spence, J. R. Organoid models of human gastrointestinal development and disease. Gastroenterology. 150 (5), 1098-1112 (2016).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Sato, T., et al. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature. 469 (7330), 415-418 (2011).
- Aronowitz, J. A., Lockhart, R. A., Hakakian, C. S. Mechanical versus enzymatic isolation of stromal vascular fraction cells from adipose tissue. Springerplus. 4 (1), 713 (2015).
- Takahashi, T. New trends and perspectives in the function of non-neuronal acetylcholine in crypt-villus organoids in mice. Methods in Molecular Biology. 1576, 145-155 (2019).
- Batlle, E., et al. β-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell. 111 (2), 251-263 (2002).
- Baghdadi, M. B., Kim, T. -. H. Analysis of mouse intestinal organoid culture with conditioned media isolated from mucosal enteric glial cells. STAR Protocols. 3 (2), 101351 (2022).
- Takahashi, T., et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS Journal. 281 (20), 4672-4690 (2014).
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nature Reviews Molecular Cell Biology. 15 (1), 19-33 (2014).
- Fordham, R. P., et al. Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell. 13 (6), 734-744 (2013).
- Miyoshi, H., et al. Wnt5a potentiates TGF-β signaling to promote colonic crypt regeneration after tissue injury. Science. 338 (6103), 108-113 (2012).
- Jung, P., et al. Isolation and in vitro expansion of human colonic stem cells. NatureMedicine. 17 (10), 1225-1227 (2011).
- Merlos-Suárez, A., et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell. 8 (5), 511-524 (2011).
- Mao, W., et al. EphB2 as a therapeutic antibody drug target for the treatment of colorectal cancer. Cancer Research. 64 (3), 781-788 (2004).
- Takahashi, T., et al. Muscarinic receptor M3 contributes to intestinal stem cell maintenance via EphB/ephrin-B signaling. Life Science Alliance. 4 (9), e202000962 (2021).
- Jung, P., et al. Isolation of human colon stem cells using surface expression of PTK7. Stem Cell Reports. 5 (6), 979-987 (2015).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Schulte, L., Hohwieler, M., Müller, M., Klaus, J. Intestinal organoids as a novel complementary model to dissect inflammatory bowel disease. Stem Cells International. 2019, 8010645 (2019).
- Puzan, M., Hosic, S., Ghio, C., Koppes, A. Enteric nervous system regulation of intestinal stem cell differentiation and epithelial monolayer function. Scientific Reports. 8 (1), 6313 (2018).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유