호르몬, 대사 및 전기 조절 연구를 위한 표적 시상하부 뉴런의 분리
요약
여기에서 우리는 배양에서 특정 시상하부 세포 아형을 성장시키기 위한 프로토콜을 제시합니다. 세포는 적절한/고유한 막 마커를 기반으로 선택할 수 있으며 면역형광, 전기생리학 및 생화학적 분석을 포함한 많은 응용 분야에서 사용할 수 있습니다.
초록
시상하부는 음식 섭취, 체온 및 호르몬 방출과 같은 다양한 기능을 제어하여 기본적인 대사 과정을 조절합니다. 시상하부의 기능은 뉴런 집단의 특정 하위 집합에 의해 제어되기 때문에 이들을 분리하는 능력은 대사 메커니즘을 연구하기 위한 주요 도구를 제공합니다. 이와 관련하여 시상 하부의 신경 복잡성은 예외적 인 문제를 제기합니다.
이러한 이유로 MACS(Magnetic-Activated Cell Sorting)와 같은 새로운 기술이 연구되었습니다. 이 논문은 마이크로비드 기술을 사용하여 태아기 생쥐의 뇌에서 표적 뉴런 집단을 분리하는 자기 활성화 세포 분류(MACS)의 새로운 응용에 대해 설명합니다. 이 기술은 간단하며 높은 재현성으로 매우 순수하고 실행 가능한 1차 시상하부 뉴런 배양을 보장합니다. 시상 하부는 부드럽게 해리되고, 뉴런은 선택적으로 분리되어 신경교 세포로부터 분리되고, 마지막으로 세포 표면 마커에 대한 특이 항체를 사용하여 관심있는 집단이 선택된다.
일단 분리되면 표적 뉴런을 사용하여 형태학적, 전기적, 내분비 특성과 정상 또는 병리학적 상태에서의 반응을 조사할 수 있습니다. 또한, 섭식, 신진대사, 스트레스, 수면 및 동기 부여를 조절하는 시상하부의 다양한 역할을 감안할 때, 표적 및 지역별 뉴런을 자세히 살펴보면 이 복잡한 환경에서 그들의 작업에 대한 통찰력을 얻을 수 있습니다.
서문
시상하부는 섭식, 신진대사, 수면, 체온, 사회적 행동, 성욕 1,2,3,4,5를 포함한 내분비, 자율, 내장 및 행동 기능을 매개하는 뇌의 다각적인 영역입니다. 기능적 이질성은 생화학적 및 전기적 메커니즘의 시너지 조합에 의해 달성됩니다: 시상하부 뉴런은 활동 전위를 발화하고 신체의 뇌 영역과 기관을 조절하기 위해 호르몬과 신경 펩티드를 분비 및 방출합니다. 마지막으로, 시상 하부 뉴런은 신체의 항상성 메시지를 번역하여 장기 및 단기 피드백 및 피드 포워드 규정으로 반응합니다6.
시상 하부의 복잡한 신경 환경에는 옥시토신과 바소프레신을 방출하는 마그 노세포 내분비 뉴런이 포함됩니다. 주로 전신 호르몬 조절에 관여하는 파보세포 뉴런, 예를 들어 갑상선 자극 호르몬 방출 호르몬(TRH) 및 부신피질 자극 호르몬 방출 호르몬(CRH)을 뇌하수체로 방출합니다. 오렉신 및 멜라닌 농축 호르몬(MCH)을 방출하는 큰 펩티드성 투영 뉴런; 및 각각 ARC POMC 및 ARCAgRP로 명명된POMC(proopiomelanocortin) 및 AgRP(아구티 관련 단백질)를 방출하는 ARC 핵(ARC)의 파보세포 펩티드성 뉴런. 분비 세포와 함께, 도파민성 뉴런, 글루타민성 뉴런, GABA 성 뉴런7을 포함한 다른 흥분성 및 억제성 뉴런은 시상하부 내 및 시상하부 외 회로를 형성하는 데 관여하며, 이에 따라 상당한 세포 이질성8의 대규모 조정 네트워크를 형성한다.
시상 하부 다양성은 연구자들이 지난 50 년 동안 극복하려고 노력해 온 도전이었습니다. 시상하부의 발달, 성숙 및 노화에서 이러한 이질성을 연구하기 위해 연구자들은 한편으로는 단일 세포 RNA 시퀀싱을 사용하여 신경 조직과 분자 및 전사체 시그니처를 탐색해 왔습니다. 이러한 노력은 시상하부 뉴런의 다양한 역할에 대한 통찰력 있는 시각을 제공했으며 세포 정체성과 생리학적 시스템에서 가능한 역할 사이의 연결을 다루었습니다 8,9,10. 다른 한편으로, 신경 기능은 광유전학적 조작과 섬유 측광 행동 접근법에 의해 조사되어 회로 구조를 면밀히 관찰할 수 있습니다. 지난 20년 동안 Cre-recombinase 기술은 연구자들이 행동과 신체 반응의 변화를 관찰하면서 표적화된 뉴런 그룹을 존재유전학적으로 자극하거나 억제할 수 있게 해주었습니다 6,11,12.
그러나 이러한 접근 방식은 복잡한 시상하부 환경 내에서의 역할에 대한 특정 세포 메커니즘이나 생물학적 기초에 대해 더 깊이 파고들지 않고 일반적인 관점에서 시상하부 기능을 검사합니다. 이를 해결하기 위해 이질적인 일차 시상하부 배양을 활용한 분자, 생화학 및 전기적 특성을 조사하는 데 초점을 맞춘 연구는 거의 없습니다. 이 연구는 복잡한 환경에서 특정 신경 과정을 해부하고 생리적 메커니즘의 통합 모델을 생성했습니다13,14,15. 그럼에도 불구하고 비특정 문화는 중대한 문제를 제기합니다. 예를 들어, 뉴런의 생리적 연결성과 해부학적 분포는 일반적으로 상호 작용하지 않는 다른 시상하부 영역의 뉴런을 도금하여 교란 효과를 일으켜 방해를 받습니다. 또한 각 지역은 역할이 다르고 다양한 신경 집단을 가지고 있어 단순한 생물학적 과정을 연구하기 어렵습니다.
이러한 문제를 해결하기 위해 지난 10년 동안 면역 패닝, FACS(Fluorescent-Activated-Cell-Sorting) 및 MACS(Magnetic-Activated-Cell-Sorting)와 같은 관심 뉴런을 분리하기 위한 새로운 접근 방식이 구현되었습니다. 면역패닝은 일련의 비뉴런(음성) 및 뉴런(양성) 선택을 위해 항체 코팅 접시를 사용하여 표적 세포를 정제하는 데 사용되는 전략입니다. 이 기술은 원칙적으로 고수율 정제 세포 배양을 생성할 수 있지만, 실제로는 성상교세포와 희소돌기아교세포가 몇 시간의 조작에 저항할 수 있기 때문에 주로 사용됩니다16,17. FACS 기술은 유세포 분석 18,19,20을 사용하여 형광 마커 및 세포 특성을 기반으로 세포를 분류하는 강력한 도구입니다. 그러나 세포 배양을 위해 세포를 분리하기 위해 이 방법을 사용한 연구는 거의 없습니다. 이 기술은 비용이 많이 들고 사용 및 유지 관리에 고도로 숙련된 인력이 필요합니다. 추가적으로, 분류 절차(21)의 끝에서 생존 가능하고 멸균된 세포를 유지하는 것은 도전적이다. 전반적으로, MACS는 시상 하부 일차 뉴런의 매우 순수하고 실행 가능한 배양을 얻기위한 간단하고 비용이 들지 않는 기술로 보입니다. 이 방법은 항체를 통해 세포에 연결된 자성 비드를 이용한다. 이를 통해 컬럼의 자기장을 사용하여 셀을 분리할 수 있습니다.
여기에서는 일반적으로 피질 뉴런과 함께 사용되는 MACS 기술을 기반으로 하는 방법을 설명합니다. 이 프로토콜은 원칙적으로 생존 가능하고 매우 순수한 시상 하부 뉴런을 분리 할 수 있습니다. 본 연구에서는 ARCPOMC 및 ARCAgRP 뉴런과 같이 Arcuate Nucleus에만 존재하는 Leptin Receptor(LepR)를 발현하는 뉴런의 1차 배양을 준비합니다. 이 뉴런은 지방 조직에서 분비되는 식욕 부진 호르몬인 렙틴에 생화학적 및 전기적 방식으로 반응합니다. 따라서 배양에서 이 뉴런 그룹을 분리하면 체외에서 호르몬, 대사 및 전기적 특성을 연구할 수 있습니다.
프로토콜
알림: 일반적인 view 실험 절차는 그림 1A에 그래픽으로 설명되어 있습니다. 이 연구에서 수행된 모든 마우스 실험은 우리 기관의 동물 관리 및 사용 위원회(IACUC)의 승인을 받았습니다. 우리는 수의사의 보살핌 하에 AAALAC(Association for Assessment and Accreditation of Laboratory Animal Care International) 승인 사육장에 보관된 3개월 된 C57BL6/J 마우스를 사용했습니다. 생쥐는 12시간의 명암 주기로 큰 우리에서 살았고 임의로 먹이를 먹었습니다.
1. 임신 및 임신 확인
- 번식을 위해 관심있는 배경과 유전자형의 마우스를 배치하십시오. 임신하기 전에 여성의 날짜와 체중을 기록하십시오.
- 6 시간 후, 프로브로 플라크에 대한 여성을 검사하십시오. 플라크가 있으면 여성과 남성을 분리하십시오. 플라크가 없으면 다음날까지 새장에 암컷을 보관 한 다음 마우스를 분리하십시오.
- 임신 후 7일, 10일, 14일에 여성의 체중을 측정하여 임신을 확인합니다.
2. 매체, 24웰 플레이트 및 재료 준비
- 세포 분리 당일, 즉시 사용 가능한 폴리-D-라이신 코팅된 유리 커버슬립( 재료 표 참조)을 다음과 같이 24웰 플레이트에 넣습니다.
- 생물학적 후드 아래에서 70 % 에탄올로 15 개의 커버 슬립이 들어있는 단일 패키지를 멸균하고 건조시킵니다. 포장을 개봉하고 커버슬립을 60mm 플레이트에 넣습니다. 플레이트를 수평으로 흔들어 커버슬립을 분리합니다. 그런 다음 플레이트를 뒤집어 24웰 플레이트의 웰에 넣을 단일 커버슬립을 집어 올립니다.
- 1.0mL의 멸균 Hank's Balanced Salt Solution(HBSS)으로 5분 동안 커버슬립을 한 번 세척합니다.
- 그 동안 다음과 같이 20.0mL의 도금 매체를 준비합니다: 18.31mL의 BME(Basal Medium Eagle, + Earle's Salts)에 1.0mL의 열 비활성화 태아 소 혈청(FBS)을 보충하고, 200μL의 피루브산나트륨(100x 스톡에서), 200μL의 글루타민(200nM 스톡에서) 및 100μL의 페니실린/스트렙토마이신(200x 스톡에서).
- 웰의 HBSS를 1.0mL의 도금 배지로 교체하고 플레이트를 37°C의 인큐베이터에 놓습니다.
- 분젠 버너를 사용하여 파스퇴르 피펫 3개를 직경이 줄어드는 불 광택을 냅니다. 한 손으로 피펫을 잡고 팁을 화염에 삽입하고 신속하게 제거하십시오. 팁이 부드러워지고 직경이 원하는 직경(눈으로 평가)으로 줄어들 때까지 이 과정을 반복합니다.
3. 신경 조직 해리를 위한 시약 준비, 신경 해리 키트의 지시에 따름
- Digestion Buffer 1을 실온에서 데운 후 Enzyme Mix 1 50 μL와 Buffer 1 1.91 mL를 혼합하여 Enzyme Mix 1을 준비하고, Enzyme Mix 2 15 μL와 Digestion Buffer 2 30 μL를 혼합하여 Enzyme Mix 2를 준비합니다. 혼합물은 모든 배아의 뇌 조직에 사용하기에 충분합니다.
- HBSS에서 0.5% 소 혈청 알부민(BSA)을 준비합니다(예: HBSS 50.0mL에 0.25g).
4. 배아 추출
- 2 개의 직선 미세 집게, 1 개의 곡선 점 겸자 및 1 쌍의 미세 수술 용 가위를 오토 클레이브하고 사용하기 전에 70 % 에탄올로 살균하십시오. 그런 다음 페트리 접시를 HBSS로 채웁니다.
- CO2 챔버에서 E14-E16 임신 댐 1개를 안락사시키고 자궁경부 탈구를 수행합니다.
알림: 다음 단계는 무균 상태에서 후드 아래에서 수행해야 합니다. - 70 % 에탄올로 복부를 소독하십시오. 수술 용 가위와 집게로 흉곽의 xiphoid 과정까지 치골 symphysis에서 복강을 잘라냅니다.
- 자궁 뿔을 추출하고 얼음처럼 차가운 HBSS로 채워진 100mm 플레이트에 넣고 철저히 씻으십시오.
- 미세한 집게로 자궁에서 모든 배아를 추출하고 분리하십시오. 미세한 수술용 가위 및/또는 집게로 배아를 빠르게 참수합니다. HBSS로 채워진 60mm 페트리 접시에 헤드를 놓습니다.
5. 시상하부 추출, 수집 및 조직 해리
- 뇌를 고정하기 위해 안구강에 미세한 집게를 하나 놓습니다. 다른 미세한 집게를 사용하여 뇌가 보일 때까지 껍질을 벗겨 피부와 두개골을 제거합니다. 흰색 모양에 따라 뇌를 다른 조직과 구별하십시오. 피부와 두개골은 분홍색이며 혈관 구조가 풍부합니다.
- 구부러진 점 집게를 사용하여 두개골에서 뇌를 제거하고 후각 구근에서 뇌를 거꾸로 뒤집어 퍼냅니다.
- 이제 피질은 복부이고 시상 하부는 상부 표면에서 등쪽으로 보입니다 (그림 1B). 구부러진 집게를 사용하여 뇌가 하얗고 깨끗해 보일 때까지 수막과 혈관 층을 제거합니다.
- 구부러진 집게를 사용하여 시상 하부 영역을 뇌의 나머지 부분과 분리하십시오.
- 시상 하부를 3-4 개의 작은 조각으로 자르고 피펫으로 조각을 15mL 튜브에 옮깁니다.
- 튜브가 얼음 위에 있는 동안 다른 배아에 대해 단계를 반복합니다.
- 튜브에 6.0mL의 HBSS를 채우고 조직이 가라앉도록 하고 상층액을 제거한 다음 효소 혼합물 1을 추가합니다. 조직을 방지하기 위해 튜브를 부드럽게 혼합하고 교반하십시오.
- 튜브를 37°C 수조에서 15분 동안 인큐베이션하고 5분마다 조직을 교반하여 조직을 재현탁시킵니다.
- 15분 후, 효소 믹스 2 30 μL를 추가합니다. 가장 큰 직경(<1mm)의 피펫 파스퇴르를 사용하여 뇌 조직을 해리합니다. 기포를 형성하지 않고 10배 위아래로 피펫.
- 수조에서 37°C에서 10분 동안 배양합니다. 튜브를 부드럽게 저어 5분마다 조직을 다시 현탁시킵니다.
- 10분 후 나머지 15μL의 효소 믹스 2를 추가합니다. 기포를 형성하지 않고 직경이 감소하는 다른 두 개의 내화 연마 피펫과 조직을 위아래로 10배 분리합니다.
- 해리된 조직이 있는 튜브에 즉시 10.0mL의 HBSS-0.5% BSA를 넣고 실온에서 10분 동안 300× g 에서 원심분리합니다.
- 상청액을 흡인하고 세포 펠릿을 1.0mL의 HBS-0.5% BSA에 재현탁합니다.
6. 세포 계수
- HBSS-0.5% BSA를 사용하여 세포 현탁액을 1:5로 희석합니다.
- 희석된 세포 현탁액 10μL를 Neubauer 계수 챔버에 넣습니다.
- 명시야 현미경으로 4개의 챔버 모서리 사각형에 있는 세포만 계산합니다. 평균을 계산하고 5 × 104를 곱합니다.
알림: 셀 분리를 진행하려면 >106 개의 셀이 있는지 확인하십시오. 최적의 셀 번호는 107입니다.
7. 네거티브 선택
참고: 음성 선택을 통해 사용자는 신경 세포와 비신경 세포를 분리하여 순수한 1차 뉴런 배양을 얻을 수 있습니다. 미리 냉각 된 솔루션을 사용하십시오.
- 세포 현탁액을 300 × g 에서 3 분 동안 원심 분리합니다 (원심 분리는 최대 10 분까지 연장 될 수 있음). 상청액을 부드럽게 흡인하고 펠릿을 HBSS-0.5% BSA 80μL에 10 7개 세포의 농도 로 재현탁합니다.
- 20 μL의 비신경 세포 비오틴-항체 칵테일을 첨가하고 4 °C에서 5분 동안 배양합니다.
- 세포를 세척하여 유리 항체를 제거하고 2.0mL의 HBSS-0.5% BSA로 300× g 에서 3분 동안 원심분리합니다.
- 상층액을 부드럽게 흡인하고 펠릿을 80μL의 HBSS-0.5% BSA에 재현탁합니다. 20μL의 안티비오틴 마이크로비드를 넣고 잘 섞은 다음 4°C에서 10분 동안 배양합니다.
- 최대 10개의7 개 셀에 대해 0.5mL의 HBSS-0.5% BSA를 추가하고 마그네틱 컬럼이 준비될 때까지 기다립니다.
8. 자기 분리, 네거티브 선택
참고: 자기 분리는 신경 세포에서 비신경 세포를 분리할 수 있는 중요한 단계입니다. 뉴런 및 비뉴런 세포를 포함하는 샘플은 자기장을 통과하고 비오틴-항체-자성 비드 복합체에 결합된 비뉴런 세포는 컬럼에 갇힙니다(그림 1C). 유리 뉴런 세포는 컬럼을 통해 용리되고 15mL 튜브에 수집됩니다.
- 도 1C에 도시된 바와 같이 분리기 및 MS 컬럼을 사용하여 스탠드(키트에 포함됨)를 준비한다.
- 컬럼을 열고 셀을 분리할 준비가 된 경우에만 스탠드를 설치하십시오.
- 0.5mL의 HBSS-0.5% BSA로 컬럼을 헹굽니다. 용액이 떨어지지 않을 때까지 기다리십시오.
- 신경 세포를 수집하려면 컬럼 아래에 15mL 튜브를 놓고 0.5mL의 세포 현탁액을 컬럼에 통과시킵니다. 물방울이 멈출 때까지 튜브에 용리액을 수집합니다. 잔류 뉴런 세포를 포획하기 위해, 컬럼을 HBSS-0.5% BSA의 3 x 0.5 mL로 세척하였다.
- 비신경 세포를 수집하려면 자석에서 컬럼을 제거하고 새 15ml 튜브 안에 넣습니다. 1.0mL의 HBSS--0.5% BSA를 컬럼에 추가하고 플런저를 사용하여 자기적으로 표지된 비뉴런 세포를 수집합니다.
- 신경 세포와 비신경 세포를 300 × g 에서 3분 동안 원심분리합니다. 상청액을 부드럽게 흡인하고 1.0mL의 HBSS-0.5% BSA에 세포를 재현탁합니다. 섹션 6의 앞부분에서 설명한 대로 셀을 계산합니다.
- 필요한 경우, 비뉴런 세포를 24-웰 플레이트에 플레이트; 그렇지 않으면 폐기하십시오.
9. 포지티브 선택
참고: 순수한 신경 세포 현탁액이 얻어지면 표적 세포를 분리하기 위해 양성 선택이 수행됩니다. 세포는 표면 항원에 대한 특이적 비오틴 접합 항체를 사용하여 분리할 수 있다. 항체는 항-비오틴 마그네틱 비드에 의해 인식됩니다. 세포 현탁액을 컬럼을 통해 흐르게 하면 관심 있는 세포만 자기장에 갇히게 됩니다.
- 순수한 신경 세포 현탁액을 300 × g 에서 3분 동안 원심분리합니다. 상층액을 부드럽게 흡인하고 펠릿을 80μL의 HBSS-0.5% BSA에 재현탁합니다. 제조업체의 지침에 따라 특정 항체를 추가하고 4°C에서 10분 동안 배양합니다.
참고: LepR을 발현하는 세포를 찾는 경우 0.50μg/106개 세포의 농도로 Mouse Leptin R Biotinylated Antibody(재료 표 참조)를 제안합니다. - 과잉 항체를 2.0mL의 HBSS-0.5% BSA로 씻어내고 300× g 에서 3분 동안 원심분리합니다.
- 상층액을 제거하고 80μL의 HBSS-0.5% BSA에 펠릿을 재현탁하고 20μL의 항비오틴 마이크로비드를 추가합니다. 4°C에서 10분 동안 배양합니다.
- 107 셀마다 0.5 mL의 HBSS-0.5 % BSA를 추가하고 자기 컬럼이 준비 될 때까지 기다립니다.
10. 자기 분리, 포지티브 선택
- 분리기와 MS 컬럼이 있는 스탠드를 준비합니다. 0.5mL의 HBSS-0.5% BSA로 MS 컬럼을 헹굽니다. 물방울이 멈출 때까지 기다리십시오.
- 15 mL 튜브를 컬럼 아래에 놓고, 0.5 mL의 세포 현탁액을 컬럼을 통해 통과시키고, 비특이적 신경세포를 포함하는 용출액을 수집한다. 잔류 비특이적 뉴런 세포의 컬럼을 세척하기 위해, 3 x 0.5 mL의 HBSS-0.5% BSA로 세척한다.
- 자석에서 컬럼을 제거하고 새 15mL 튜브에 넣고 1.0mL의 HBSS-0.5% BSA를 추가합니다. 플런저를 사용하여 표적 세포를 씻어냅니다.
- 두 튜브를 300 × g 에서 3분 동안 원심분리합니다. 상층액을 부드럽게 제거하고 0.5mL의 도금 배지에 다시 현탁합니다.
- 섹션 6에서 설명한 대로 셀을 계산합니다.
- 섹션 2에 기재된 바와 같이 미리 제조된 24-웰 플레이트에서 양성 대조군으로서 표적 세포 및 음성 대조군으로서 비특이적 세포 모두를 플레이트하고, 5% CO2, 9% O2 및 95% 습도에서 37°C에서12시간 동안 배양한다.
11. 세포 배양 유지
- 19.2mL의 신경 배양 배지, 400μL의 B27 보충제(50x 스톡에서), 200μL의 글루타민(200μM 스톡에서) 및 100μL의 페니실린/스트렙토마이신(200x 스톡에서)으로 20mL의 배양 배지를 준비합니다.
- 뉴런 또는 비뉴런 세포를 포함하는 24웰 플레이트에서 도금 매체를 교체합니다.
- HBSS 2 x 1.0mL로 세척합니다.
- 배양 배지 1.0mL를 추가합니다.
- 0.5mL의 기존 배지를 0.5mL의 새 배지로 교체하여 2/3일마다 매체를 새로 고칩니다.
참고: 세포는 배양 중에 유지될 수 있으며 시험관 내에서 최대 21일 동안 사용할 수 있습니다(DIV21).
12. 뉴런 면역형광염색
- 염색하기 12시간 전에, 50/50 메탄올 및 아세톤으로 이루어진 용액을 제조하고, 이를 -20°C에서 밤새 냉각시켰다.
- 24웰 플레이트에서 뉴런을 2 x 1.0mL의 1x 인산염 완충 식염수(PBS)로 5분 동안 세척합니다.
- PBS 용액을 50/50 용액 1.0mL로 교체하고 얼음에서 20분 동안 배양합니다.
- 1x PBS로 3 x 5분 동안 세척합니다.
- 실온에서 1시간 동안 1x PBS에서 3% BSA로 뉴런을 차단합니다.
- 제조업체의 지침에 언급된 항체 농도를 사용하여 1x PBS의 3% BSA에서 1차 항체 용액을 준비합니다. 사용된 항체 및 농도는 재료 표에 나열되어 있습니다.
- 블로킹 용액을 1차 항체 용액으로 교체하고 4°C에서 밤새 인큐베이션합니다.
- 세포를 1x PBS로 3 x 10분 동안 세척합니다.
- 제조업체의 지침에 따라 항체 농도를 사용하여 1x PBS에 3% BSA가 포함된 2차 항체 용액을 준비합니다. 사용된 2차 항체는 재료 표에 나열되어 있습니다.
- 세포를 실온에서 1시간 동안 2차 항체 용액과 함께 배양합니다.
- 세포를 1x PBS로 3 x 10분 동안 세척합니다.
- 장착 절차 중에 뉴런을 1x PBS에 그대로 두십시오. 현미경 슬라이드에 장착 매체(핵 식별을 위한 4',6-diamidino-2-phenylindole 포함 또는 제외)를 작은 방울로 떨어뜨립니다. 집게로 뉴런이 들어 있는 유리 커버슬립 하나를 추출하고 커버슬립의 측면을 종이 티슈에 두드려 초과 PBS를 건조시킵니다. 장착 매체의 커버슬립을 뒤집어 뉴런이 현미경 슬라이드를 향하고 있는지 확인합니다. 티슈 페이퍼로 여분의 장착 매체를 부드럽게 누르고 제거합니다.
- 현미경 슬라이드는 명시야 또는 컨포칼 현미경으로 분석할 준비가 되었습니다.
결과
이 논문은 시상하부의 표적 뉴런을 분리하기 위한 프로토콜을 설명합니다 (그림 1). 이 방법의 범위는 통제되고 고립 된 맥락에서 특정 뉴런 특성을 연구하는 것입니다. 따라서, 마우스 배아는 E14-E16에서 임신한 댐으로부터 추출되었다. 수막이 제거되고 시상 하부가 뇌의 나머지 부분과 분리되었습니다. 조직은 참조된 해리 키트를 사용하여 갓 제조된 두 가지 효소 혼합물로 부드럽게 해리되었습니다. 먼저, 뉴런 세포로부터 비뉴런 세포를 분리하였다 - 신경교세포, 미세아교세포, 뉴런을 동일한 단일세포 현탁액에 채취하였다. 이를 위해, 비뉴런 세포는 비뉴런 표면 에피토프를 인식하는 항체의 칵테일을 사용하여 표지되었다. 인큐베이션 후, 항체-세포 복합체를 자성 마이크로비드와 접합시키고, 후속적으로 자기 컬럼을 통과시켜 비뉴런 세포를 포획하였다.
이 단계는 두 개의 세포 현탁액을 생성했는데, 하나는 비뉴런을 함유하고 다른 하나는 뉴런 세포를 함유합니다. 두 현탁액 모두 즉시 도금 할 수 있습니다. 대안적으로, 뉴런 세포 현탁액은 동일한 전략을 사용하여 나머지로부터 뉴런 서브-집단(표적 현탁액)을 분리하기 위해 추가로 조작될 수 있다. 관심 있는 실험에 기초하여, 뉴런 세포는 125,000 내지 200,000 cells/mm3로 도금될 수 있다. 밀도가 낮은 배양은 축삭 발달, 시냅스 형성 및 전달에서 전기 생리학에 이르기까지 단일 세포 분해능으로 뉴런을 분석하는 데 사용할 수 있습니다. 밀도가 높은 배양은 DNA 및 RNA 추출, 웨스턴 블롯, 서던 블롯, 노던 블롯, 실시간 PCR 및 RNA 시퀀싱을 포함한 생화학적 분석에 사용할 수 있습니다.
이 연구에서 LepR은 ARCPOMC 및 ARCAgRP 뉴런과 같은 멜라노코르틴 시스템에 관여하는 뉴런을 분리하는 것을 목표로 했습니다. 세포는 LepR+ 뉴런의 경우 120,000 cells/mm 3에서 일반 뉴런 집단의 경우 200,000 cells/mm3 범위의 밀도로 도금되었습니다. 48시간 후, LepR+ 뉴런은 신경돌기를 형성하기 시작했습니다(그림 2). DIV4에서 축삭 확장은 진행을 보였고 수지상 과정이 나타나기 시작했습니다. DIV6에서 뉴런은 충분히 발달되어 분석 준비가되었습니다. LepR+ 뉴런에 대한 면역형광 실험에서 LepR이 99% 발현되는 것으로 나타났습니다(녹색, 그림 3A). 신경교세포 또는 다른 비뉴런 세포는 관찰되지 않았으며, 이는 1차 뉴런 배양의 순도를 확인시켜 주었다. 세포의 신경 특성은 축삭 및 수지상 돌출부의 식별과 함께 미세소관 관련 단백질 2(MAP2) 염색에 의해 확인되었습니다(그림 3B). DIV10에서 LepR+ 세포의 30%가 POMC를 발현했습니다(빨간색). 이는 대부분의 LepR+ 세포가 POMC 또는 AgRP를 발현하기 때문에 예상됩니다. 그림 3C,D는 POMC와 LepR 신호 사이의 공동 위치 파악을 보여줍니다. 공동 국소화는 예상대로 핵과 그 주변에서 두드러졌습니다.
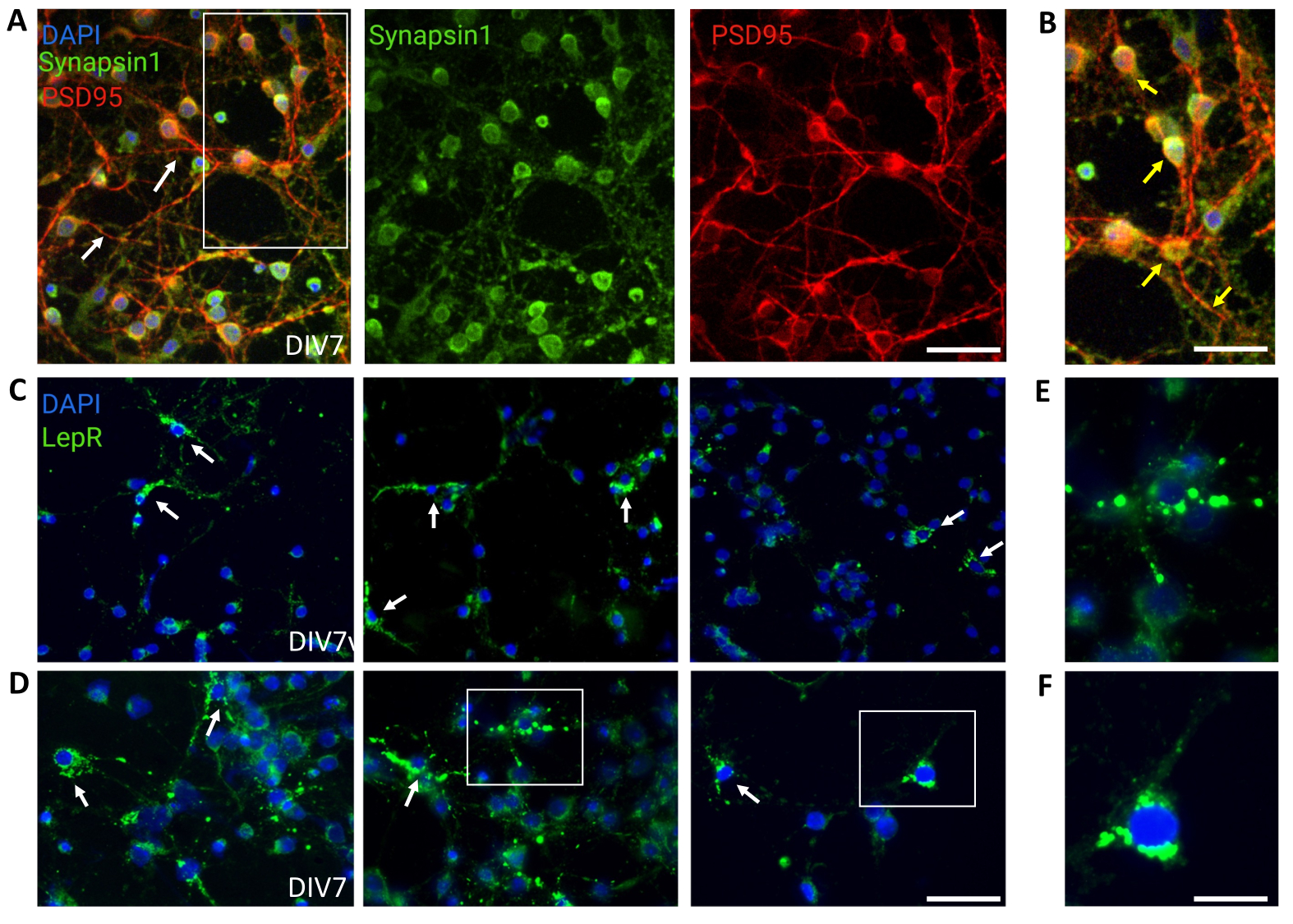
이질적인 시상하부 뉴런 집단을 포함하는 일반 배양이 대조군에 사용되었습니다. 면역형광은 Synapsin-1(녹색) 및 PSD 95(빨간색) 동시 염색으로 평가한 바와 같이 시냅스 연결성 및 기능을 입증했습니다(그림 4A,B). 일반 배양에 존재하는 LepR+ 뉴런의 수는 ~5%였으며, 이는 대부분의 LepR 발현 뉴런이 자기 분리 과정에서 선택되었다는 개념과 일치하는 비율입니다(대표적인 LepR+ 세포는 그림 4C, D에 설명되어 있음). 이 연구 중에 생성되거나 분석된 모든 데이터는 https://doi.org/10.5061/dryad.cnp5hqc9c 에서 사용할 수 있습니다.

그림 1: 실험 순서도 및 설정 . (A) 실험 절차의 그래픽 표현. Go-no-go: ≥106 개의 세포가 세포 분리를 진행하는 데 필요합니다. 최적의 셀 번호는 107입니다. (B) E16 배아 뇌의 대표 이미지. 시상 하부와 수막이 표시됩니다. (C) 표적 세포의 분리 및 분리에 사용되는 MACS 설정. 마그네틱 스탠드, 마그네틱 분리기 및 컬럼이 표시됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 2: DIV2와 DIV6 사이의 신경 배양 . (A) 양성 선택에서 얻은 LepR+ 세포는 세포 밀도와 연결성이 감소했지만 신경돌기(빨간색 화살표), 축삭돌기(파란색 화살표) 및 수상돌기(녹색 화살표)의 정상적인 발달을 보여줍니다. 세포는 120,000 cells/mm3의 밀도로 도말하였다. 스케일 바 = 100 μm. (B) 200,000 cells/mm3의 밀도로 도금된 일반 시상하부 뉴런은 정상적인 발달 및 성장 특징과 연결성(검은색 화살표)을 나타냅니다. 스케일 바 = 100 μm. 약어: DIV = 시험관 내 일수; LepR = 렙틴 수용체. 이 수치를 구성하기 위해 생성되거나 분석된 모든 데이터는 https://doi.org/10.5061/dryad.cnp5hqc9c 에서 확인할 수 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 배양된 LepR+ 뉴런에 의해 요약된 생체 내 뉴런. (a) LepR을 발현하는 뉴런의 배양물의 대표적인 이미지(녹색; DAPI는 파란색입니다. 세포의 99%가 LepR을 발현했습니다. 뉴런은 120,000 cells/mm3 밀도로 도금되었습니다. DIV7에서 뉴런은 길쭉한 축삭, 수지상 성숙 및 뉴런 연결성(화살표)을 나타냈습니다. 스케일 바 = 40 μm. (B) 세포의 신경 특성을 확인하기 위해 anti-MAP2를 사용한 면역형광을 사용했습니다. 축삭돌기(axon), 수상돌기(dendrites), 돌출부(protrusions)와 같은 뉴런 특이적 형태가 입증됩니다(화살표). 스케일 바 = 40 μm. (C) LepR+ 세포의 확대. 녹색의 LepR과 파란색의 DAPI. 스케일 바 = 10 μm. (D) LepR(녹색) 및 POMC(빨간색)와의 공동 염색. LepR+ 뉴런의 약 30%는 핵 수준(빨간색 화살표)에서 검출된 POMC에 대해 공동 면역반응이었습니다. 스케일 바 = 10 μm. 약어: DAPI = 4',6-디아미디노-2-페닐인돌; MAP2 = 미세소관 관련 단백질 2; POMC = 프로오피오멜라노코르틴; LepR = 렙틴 수용체. 이 수치를 구성하기 위해 생성되거나 분석된 모든 데이터는 https://doi.org/10.5061/dryad.cnp5hqc9c 에서 확인할 수 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 배양 된 일반 시상하부 뉴런에 의해 생체 내 세포가 요약됩니다. (A) Synapsin 1(녹색), PSD95(빨간색) 및 DAPI(파란색)로 염색된 일반 시상하부 신경 배양의 대표 이미지. 뉴런은 잘 발달 된 연결성과 시냅스 기능 (화살표)을 나타냈다. 스케일 바 = 40 μm. (B) Synapsin 1(녹색)과 PSD95(빨간색, 화살표)의 공동 국소화를 보여주는 (A)의 상자 확대. 스케일 바 = 20 μm. (씨, 디) 일반 배양액에서 LepR+ 세포를 보여주는 대표 이미지. LepR+ 세포(녹색)는 전체의 ~5%를 차지했습니다. 대표적인 LepR+ 세포는 화살표로 표시됩니다. 스케일 바 = 40 μm. (E,에프) (C,D)에 있는 상자의 배율은 소마에 국한된 녹색 반점 렙틴 수용체를 보여줍니다. 스케일 바 = 20 μm. 이 수치를 구성하기 위해 생성되거나 분석된 모든 데이터는 https://doi.org/10.5061/dryad.cnp5hqc9c 에서 확인할 수 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
토론
시상하부 뉴런의 생화학적 및 전기적 특성을 조사하는 것은 신진대사, 체온 조절, 기분 관리, 섭식 행동 등의 분자적 기초를 이해하는 데 중요합니다. 그러나 시상하부의 신경 이질성은 이러한 노력을 어렵게 만들고 특정 시상하부 하위 집단을 분리하고 연구하는 방법이 필요합니다.
생체 내 기술은 CRE-재조합효소, 광유전학, 섬유 광도 측정 및 칼슘 이미징을 사용합니다. 이러한 접근법은 주로 시상 하부 뉴런의 전기적 특성을 연구 할 수 있으며, 현재 비 전기적 특성을 조사하는 데 사용할 수있는 방법은 거의 없습니다. 이 연구에서 개발된 MACS 기술은 시험관 내에서 특정 시상하부 신경 하위 집단을 분리할 수 있는 기술을 제공하여 표적 치료 및 분석을 제공할 수 있습니다. 뉴런 배양은 다른 뉴런 집단의 공동 배양에 비해 관리가 더 간단합니다. 또한 순수 배양은 아교세포와 미세아교세포의 존재로 인한 교란 효과를 피합니다. 따라서 동일한 시상 하부 영역과 유형의 뉴런은 특정 대사 및 호르몬 입력에 대한 반응으로 연구 될 수 있습니다.
이 프로토콜에서 우리는 LepR을 발현하는 시상하부 뉴런을 선택했습니다. 분리된 LepR+ 세포를 배양하여 생체 내에서 연구하기 어려운 세포, 형태학적 및 분자적 특성을 조사했습니다.배양의 순도는 99%로 방법의 정확성을 뒷받침했습니다. 또한, LepR+ 세포는 DIV21까지 DIV7에서 건강하고 생존 가능했습니다.
그러나 이 기술에는 몇 가지 제한 사항이 있습니다. E18 이상의 순수 뉴런 배양은 유지하기가 어렵습니다. 따라서 추출 창은 E14-E16으로 제한됩니다. 이것은 E16 이후에 발생하는 세포 변화가 누락되었음을 의미합니다. 예를 들어, ARC 뉴런에서 렙틴 수용체의 발현은 출생 후 초기 기간 동안 증가한다22. 분리 절차는 세포 스트레스와 사망을 줄이고 수율을 향상시키기 위해 가능한 한 빨리 수행되어야합니다. 절차는 최대 5시간이 소요될 수 있습니다. 따라서 멸균 상태를 유지하고 필요한 최소한으로 조작을 줄이는 것이 필수적입니다. 양성 선택은 사용 가능한 조직의 양이 적기 때문에 수율이 낮아져 단일 제제로 수행할 수 있는 실험 횟수가 제한될 수 있습니다. 상승된 뉴런 사멸이 관찰되었는데, 아마도 낮은 세포 밀도와 감소된 뉴런 연결성 및 뉴런 내 지지로 인한 것입니다.
또한, 관심 항원을 표적으로 하는 항체는 정확한 분리를 보장하기 위해 세포 표면에 결합해야 합니다. 일반적으로 유세포 분석에 사용되는 항체는 MACS 기술에 적합합니다. 항체가 이전에 세포 분리 방법에 사용된 적이 없는 경우, 이상적인 용도와 농도를 결정하기 위해 검증 및 적정 실험이 필요합니다. 표적 세포를 추출하려면 세포 표면 마커가 필요합니다. 여기서는 비오티닐화 항체를 사용했지만 원칙적으로 FITC(플루오레세인 이소티오시아네이트) 및 PE(정제된 항-피코에리트린)와 같은 다른 분자와 접합된 항체도 사용할 수 있습니다. MACS 기술은 GFP 또는 다른 Tag 단백질과 같은 형광단을 발현하는 뉴런에도 적용되어 잠재적으로 특이성과 수율을 증가시킬 수 있습니다. 형광단을 사용하지 않는 경우 대안은 생세포 실험을 수행하기 전에 면역형광법으로 관심 분자의 발현을 확인하는 것입니다. 향후 연구에서는 이러한 대안의 타당성을 테스트할 것입니다.
이 연구가 다루지 않은 한 가지 중요한 측면은 하위 뉴런 집단의 "충실도"에 관한 것입니다. 우리는 배양된 LepR+ 뉴런이 천연 ARCPOMC 뉴런의 시그니처인 POMC를 발현한다는 것을 확인했습니다. 그러나 LepR+ 뉴런 배양이 생체 내 고유 대응물을 요약한다는 결론을 내리기 위해서는 더 많은 테스트가 필요할 것입니다. 전반적으로, 여기에 제시된 MACS 신경 분리 프로토콜은 생체 내에서 조사하기 어려운 시험관 내 시상하부 메커니즘을 연구하는 타당하고 효과적인 방법을 제공할 수 있습니다.
공개
저자는 이해 상충이 없다고 선언합니다.
감사의 말
그래픽 그림은 BioRender.com 로 만들어졌습니다. 이 작업은 FS에 대한 NIA 보조금(R01AG060919)과 NSF 보조금(2030348)의 지원을 받았습니다.
자료
| Name | Company | Catalog Number | Comments |
| Embryo extraction | |||
| 1 curved point forceps | Fine Science Tools | 11270-20 | Dumont |
| 1 fine surgical scissor | Fine Science Tools | 14058-11 | Dumont |
| 100 mm Petri dish | Corning | 430167 | |
| 2 straight fine forceps | Fine Science Tools | 11254-20 | Dumont |
| 60 mm Petri dish | Corning | 430196 | |
| 70% ethanol | Decon Laboratories, INC. | 2801 | Ethanol 190 Proof |
| Anti-Biotin MicroBeads 1mL | Miltenyi Biotec | 130-115-390 | |
| Anti-MAP2 antibody | Abcam | ab5392 | 1 : 800 |
| Bench pads | |||
| Bovine Serum Albumin | Sigma-Aldrich | A9418-50G | |
| Buffer Y | Miltenyi Biotec | 130-094-802 | |
| Buffer Z | Miltenyi Biotec | 130-094-802 | |
| Cell Culture | |||
| Anti-Biotin MicroBeads 1mL | Miltenyi Biotec | 130-115-390 | |
| Bovine Serum Albumin | Sigma-Aldrich | A9418-50G | |
| Buffer Y | Miltenyi Biotec | 130-094-802 | |
| Buffer Z | Miltenyi Biotec | 130-094-802 | |
| Enzyme A | Miltenyi Biotec | 130-094-802 | |
| Enzyme P | Miltenyi Biotec | 130-094-802 | |
| GG-12-1.5, 12 mm dia.#1.5 thick 100 pc cell culture tested German coverglasses | Neuvitro Corporation | GG-12-15 | |
| Gibco B-27 Supplement 10 mL | ThermoFisher | 17504-044 | |
| Gibco Basal Medium Eagle (BME) 500 mL | ThermoFisher | 21010046 | (+) Earle's Salts, (-) L-Glutamine |
| Gibco HBBS (1x) Hanks' Balanced Salt Solution 500 mL | ThermoFisher | 14025092 | Calcium, Magnesium, No phenol red |
| Gibco HI FBS 100 mL | ThermoFisher | 16140-063 | |
| Gibco L-Glutamine 200 mM (100x) | ThermoFisher | 25030-081 | |
| Gibco Penicilline/Streptomicine | ThermoFisher | 15140-122 | 10,000 U/mL |
| Gibco Sodium Pyruvate (100 mM) 100 mL | ThermoFisher | 11360070 | |
| MiniMACS Separator and Starting Kit | Miltenyi Biotec | 130-042-102 | |
| Mouse Leptin R Biotinylated Antibody | R&D Systems | ABAF497 | 0.25 μg/106 cells |
| MS Column | Miltenyi Biotec | 130-042-201 | |
| Neaubeaur-Improved Brightline 100 µm Chamber | Hausser Scientific | 3120 | |
| Neural Tissue Dissociation Kit - Postnatal Neurons | Miltenyi Biotec | 130-094-802 | |
| Neuronal Culture Medium 500 mL | ThermoFisher | 88283 | |
| Non-Neuronal Cell Biotin-Antibody Cocktail mouse 1 mL | Miltenyi Biotec | 130-115-389 | |
| Olympus SZ61 Zoom Stereomicroscope | Olympus Life Science | SZ61/SZ51 | |
| Pierce Primary Neuron Isolation Kit | ThermoFisher | 88280Y | |
| Staining | |||
| Anti-MAP2 antibody | Abcam | ab5392 | 1 : 800 |
| Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | ThermoFisher | A32766 | 1 : 500 |
| Donkey anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | ThermoFisher | A32790 | 1 : 500 |
| Dulbecco's Phosphate Buffered Saline (DPBS) | Sigma Aldrich | MFCD00131855 | |
| Goat anti-Chicken IgY (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 647 | ThemoFisher | A32933 | 1 : 500 |
| Goat anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 594 | ThermoFisher | A11037 | 1 : 200 |
| Invitrogen Leptin Receptor Recombinant Rabbit Monoclonal Antibody (JA73-01) | ThermoFisher | MA5-32685 | 1 : 500 |
| Mouse Leptin R Biotinylated Antibody | R&D Systems | ABAF497 | 1 : 500 |
| POMC Rabbit mAb | Cell Signaling Technology | D3R1U | 1 : 500 |
| PSD95 (D74D3) XP Rabbit mAb | Cell Signaling Technology | D74D3#3409 | 1 : 500 |
| Streptavidin, Alexa Fluor 594 conjugate | ThermoFisher | S11227 | 1 : 500 |
| Synapsin 1 Monoclonal Antibody (7H10G6) | ThermoFisher | MA5-31919 | 1 : 500 |
| Vectashield Plus Antifade Mountina Medium with DAPI 10 mL | Vector Laboratories | H-2000 |
참고문헌
- Cone, R. D. Anatomy and regulation of the central melanocortin system. Nature Neuroscience. 8 (5), 571-578 (2005).
- Clarke, I. J. Hypothalamus as an endocrine organ. Comprehensive Physiology. 5 (1), 217-253 (2015).
- Mignot, E., Taheri, S., Nishino, S. Sleeping with the hypothalamus: emerging therapeutic targets for sleep disorders. Nature Neuroscience. 5 Suppl, 1071-1075 (2002).
- Baird, A. D., Wilson, S. J., Bladin, P. F., Saling, M. M., Reutens, D. C. Neurological control of human sexual behaviour: insights from lesion studies. Journal of Neurology, Neurosurgery, and Psychiatry. 78 (10), 1042-1049 (2007).
- Caria, A., Dall, O. G. Functional neuroimaging of human hypothalamus in socioemotional behavior: a systematic review. Brain Sciences. 12 (6), 707 (2022).
- Andermann, M. L., Lowell, B. B. Toward a wiring diagram understanding of appetite control. Neuron. 95 (4), 757-778 (2017).
- Romanov, R. A., Alpar, A., Hokfelt, T., Harkany, T. Unified classification of molecular, network, and endocrine features of hypothalamic neurons. Annual Review of Neuroscience. 42, 1-26 (2019).
- Hajdarovic, K. H., Yu, D., Webb, A. E. Understanding the aging hypothalamus, one cell at a time. Trends in Neurosciences. 45 (12), 942-954 (2022).
- Zhang, Y. H., et al. Cascade diversification directs generation of neuronal diversity in the hypothalamus. Cell Stem Cell. 28 (8), 1483-1499 (2021).
- Chen, R., Wu, X., Jiang, L., Zhang, Y. Single-cell RNA-seq reveals hypothalamic cell diversity. Cell Reports. 18 (13), 3227-3241 (2017).
- Ma, C., et al. Neural pathways from hypothalamic orexin neurons to the ventrolateral preoptic area mediate sleep impairments induced by conditioned fear. Frontiers in Neuroscience. 17, 1122803 (2023).
- Wang, F., et al. A parabrachial to hypothalamic pathway mediates defensive behavior. Elife. 12, e85450 (2023).
- Cowley, M. A., et al. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature. 411 (6836), 480-484 (2001).
- Parekh, R. U., et al. Hypothalamic kinin B1 receptor mediates orexin system hyperactivity in neurogenic hypertension. Scientific Reports. 11 (1), 21050 (2021).
- Schmidt, C. X., Tsang, A. H., Oster, H. Generation of mouse primary hypothalamic neuronal cultures for circadian bioluminescence assays. Bio-protocol. 11 (5), e3944 (2021).
- Foo, L. C. . Purification of rat and mouse astrocytes by immunopanning. 2013 (5), 421-432 (2013).
- Emery, B., Dugas, J. C. . Purification of oligodendrocyte lineage cells from mouse cortices by immunopanning. 2013 (9), 854-868 (2013).
- Zhao, H., et al. Changes of constituents and activity to apoptosis and cell cycle during fermentation of tea. International Journal of Molecular Sciences. 12 (3), 1862-1875 (2011).
- Zhang, Z. M., et al. Down-regulation of human leukocyte antigens class I on peripheral T lymphocytes and NK cells from subjects in region of high-incidence gastrointestinal tumor. Chinese Medical Journal. 124 (12), 1813-1817 (2011).
- Drake, S. S., Zaman, A., Simas, T., Fournier, A. E. Comparing RNA-sequencing datasets from astrocytes, oligodendrocytes, and microglia in multiple sclerosis identifies novel dysregulated genes relevant to inflammation and myelination. WIREs Mechanisms of Disease. 15 (2), e1594 (2023).
- Mattanovich, D., Borth, N. Applications of cell sorting in biotechnology. Microbial Cell Factories. 5, 12 (2006).
- Cottrell, E. C., et al. Developmental changes in hypothalamic leptin receptor: relationship with the postnatal leptin surge and energy balance neuropeptides in the postnatal rat. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 296 (3), R631-R639 (2009).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유