
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Реактивация моделей демембранированных клеток в Chlamydomonas reinhardtii
В этой статье
Резюме
Реактивация подвижных клеток in vitro является решающим экспериментом в понимании механизмов подвижности клеток. Протокол описывает реактивацию демембранированных клеточных моделей Chlamydomonas reinhardtii, модельного организма для изучения ресничек / жгутиков.
Аннотация
Со времени исторического эксперимента по сокращению глицеринированных мышц путем добавления АТФ, который Сент-Дьёрдьи продемонстрировал в середине 20-го века, реактивация демембранированных клеток in vitro была традиционным и мощным способом изучения подвижности клеток. Фундаментальным преимуществом этого экспериментального метода является то, что состав реактивационного раствора может быть легко изменен. Например, среда с высокой концентрацией Ca2+ , которая возникает только временно из-за возбуждения мембраны in vivo , может быть воспроизведена в лаборатории. Эукариотические реснички (также известные как жгутики) являются сложным механизмом подвижности, регуляторные механизмы которого еще предстоит выяснить. Одноклеточная зеленая водоросль Chlamydomonas reinhardtii является отличным модельным организмом в области исследований ресничек. Эксперименты по реактивации с использованием демембранированных клеточных моделей C. reinhardtii и их производных, таких как демембранированные аксонемы изолированных ресничек, в значительной степени способствовали пониманию молекулярных механизмов цилиарной моторики. Эти эксперименты показали, что АТФ заряжает энергией цилиарную подвижность и что различные клеточные сигналы, включая Ca2+, цАМФ и активные формы кислорода, модулируют цилиарные движения. Здесь описан точный метод демембранации клеток C. reinhardtii и реактивации клеточных моделей.
Введение
Реактивация in vitro демембранированных подвижных клеток является ценным инструментом для изучения молекулярной основы регуляторного механизма подвижности клеток. Сент-Дьёрдьи впервые продемонстрировал in vitro сокращение волокон скелетных мышц кролика, извлеченных с 50% глицерином путем добавления аденозинтрифосфата (АТФ)1. Этот эксперимент был первым, доказавшим, что АТФ активизирует сокращение мышц. Методология вскоре была применена для изучения атф-энергетической цилиарной /жгутиковой подвижности, такой как жгутики сперматозоидов2, Paramecium cilia 3 и Chlamydomonas reinhardtii cilia (также называемые жгутиками)4 с использованием неионных детергентов для демембранации.
Одноклеточная зеленая водоросль C. reinhardtii является модельным организмом для изучения ресничек: она плавает с двумя ресничками, обыгрывая их, как брассомчеловека 5. Цилиарное движение управляется динеином, моторным белком на основе микротрубочек 6,7. Цилиарные динеины можно классифицировать на наружные динеины и внутренние динеины. Мутанты, лишенные каждого вида динеина, были выделены как медленно плавающие мутанты с различными аномалиями моторики. Подробный анализ подвижности этих мутантов in vitro значительно продвинул исследование динеина8.
Многие важные результаты были достигнуты с использованием этого метода и его производных с тех пор, как был создан эксперимент по реактивации in vitro демембранированных клеток C. reinhardtii (клеточные модели). Реактивация клеточных моделей в серии буферов Ca2+, например, показала9, что две реснички по-разному регулируются субмикромолярным Ca2+, и этот асимметричный контроль ресничек обеспечивает фототаксическую ориентацию C. reinhardtii10. Кроме того, обе реснички показывают преобразование формы сигнала из режима прямого плавания (называемого асимметричной формой сигнала) в режим обратного плавания (называемый симметричной формой волны, которая появляется в течение короткого периода, когда клетки фото- или механо-шокированы)11,12. Это преобразование формы сигнала регулируется субмиллимолярным Ca2+, что было показано реактивацией так называемого нуклеофлагеллярного аппарата (комплекса, содержащего две реснички, базальные тела, структуры, связывающие базальные тела с ядром, и остаток ядра)11 или демембранированные аксонемы изолированных ресничек13. Помимо Ca2+, окислительно-восстановительное (восстановительно-окислительное) равновесие является сигналом, регулирующим частоту цилиарного биения, что было показано реактивацией клеточных моделей в окислительно-восстановительных буферах, содержащих различные соотношения восстановленного глутатиона и окисленного глутатиона14. Кроме того, циклический аденозинмонофосфат (цАМФ) асимметрично регулирует две реснички, что было показано реактивацией аксонем с фоторасщепляемыми клетками цАМФ15. Эти результаты in vitro в сочетании с генетическими находками привели к более глубокому пониманию молекулярных механизмов регуляции ресничек у C. reinhardtii.
Протокол для реактивации моделей клеток описан здесь. Метод прост, допускает различные модификации и может быть применен к нескольким организмам, которые движутся с ресничками. Однако, поскольку демембранированные клетки являются хрупкими, требуются некоторые советы для реактивации подвижности клеточных моделей с хорошей эффективностью, предотвращая дециляцию.
протокол
Для настоящего исследования был использован штамм Chlamydomonas reinhardtii дикого типа, CC-125. CC-125 был получен из Ресурсного центра Хламидомонас (см. Таблицу материалов) и поддерживался на трис-ацетат-фосфате (TAP)16, 1,5% агарозной среде при 20-25 °C.
1. Клеточная культура
- Культура Chlamydomonas reinhardtii (CC-125) в среде TAP16 в светло-темный период 12 ч/12 ч (светлые условия для светового периода: ~50 мкмоль фотонов m−2 s−1 белого света) при 20-25 °C в течение 2 дней (рисунок 1).
ПРИМЕЧАНИЕ: Клетки должны находиться в среднелогарифмической фазе роста (фильм 1). Длительная культивирование (>4 дня, в позднелогарифмической фазе роста или стационарной фазе) снижает эффективность реактивации моделей демембранированных клеток.

Рисунок 1: Жидкая культура после 2-дневного культивирования. Из пластины агара TAP-1,5% кусок клеток дикого типа для заполнения платиновой петли инокулировали до ~ 150 мл жидкой среды TAP в колбе. Плотность клеток после 2-дневной культуры составляла 2,3 × 106 клеток/мл. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Фильм 1: Плавание живых клеток. Клетки наблюдались под микроскопом с 10-кратной объективной линзой и масляным погружным конденсатором темного поля. Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот фильм.
2. Подготовка моделей демембранированных клеток
ПРИМЕЧАНИЕ: Перед началом эксперимента держите промывочный буфер при комнатной температуре, а буферы демембранации, разбавления, реактивации и раствор АТФ на льду. Состав этих буферов приведен в дополнительной таблице 1.
- Центрифуга ~10 мл жидкой культуры при 1000 × г при 20 °С в течение 3 мин.
ПРИМЕЧАНИЕ: На протяжении всего протокола с этого шага не используйте автоклавную пластиковую посуду (трубки, наконечники пипеток и т. Д.), Снижая эффективность реактивации. Для конических труб предпочтительнее повторно используемые. Плотность клеток после 2-дневной культуры может составлять обычно 1,0-5,0 × 106 клеток/мл. Когда плотность клеток ниже этой, возьмите достаточный объем культуры, чтобы содержать ~ 5 × 107 клеток. - Отбросьте супернатант сначала декантацией, а затем пипеткой Пастера и повторно суспендируйте осадки в ~5 мл промывочного буфера.
- Центрифужные ячейки в промывочном буфере при 1000 × г в течение 3 мин при 20 °С.
- Осторожно выбросьте супернатант с помощью пипетки (рисунок 2).
ПРИМЕЧАНИЕ: Пипетки Pasteur предпочтительнее. При использовании микропипеток избегайте использования наконечников пипеток, которые были автоклавированы. - Наложите ~ 0,5 мл буфера демембранации на гранулу ячейки, аккуратно встряхните трубку вручную, чтобы примерно приостановить ячейки в буфере, и положите трубку на лед (рисунок 3A).
ПРИМЕЧАНИЕ: В данный момент нет необходимости полностью приостанавливать гранулу. Повышение концентрации MgSO4 до 15 мМ в буферах демембранации и реактивации при реактивации клеточных моделей с конечной концентрацией АТФ >1 мМ для стабильной реактивации17. - Осторожно приостановите оставшуюся ячейку с помощью пипетки и снова положите трубку на лед (рисунок 3B).
- Возьмите 5-10 мкл моделей клеток, разбавьте 10-кратное с буфером разбавления и наблюдайте под микроскопом (фильм 2), чтобы убедиться, что все модели клеток демембранированы и не плавают (рисунок 4).
ПРИМЕЧАНИЕ: Если некоторые клетки все еще плавают (живы), добавьте неионное моющее средство, используемое в буфере демембранации, непосредственно в раствор клеточной модели до конечной концентрации ~ 0,15%. Альтернативно, повторяют этапы 2.1-2.5 с раствором для демембранации, содержащим 0,15% моющего средства.

Рисунок 2: Выброс супернатанта. Остальная часть супернатанта была аккуратно удалена пипеткой Пастера после того, как супернатант был удален декантацией трубки. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Демембранация. (А) После наложения 0,5 мл демембранационного раствора на гранулу ячейки раствор смешивали рукой, чтобы грубо приостановить клетки. (B) После смешивания остальная часть гранулы ячейки была полностью суспендирована в растворе пипеткой Пастера. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

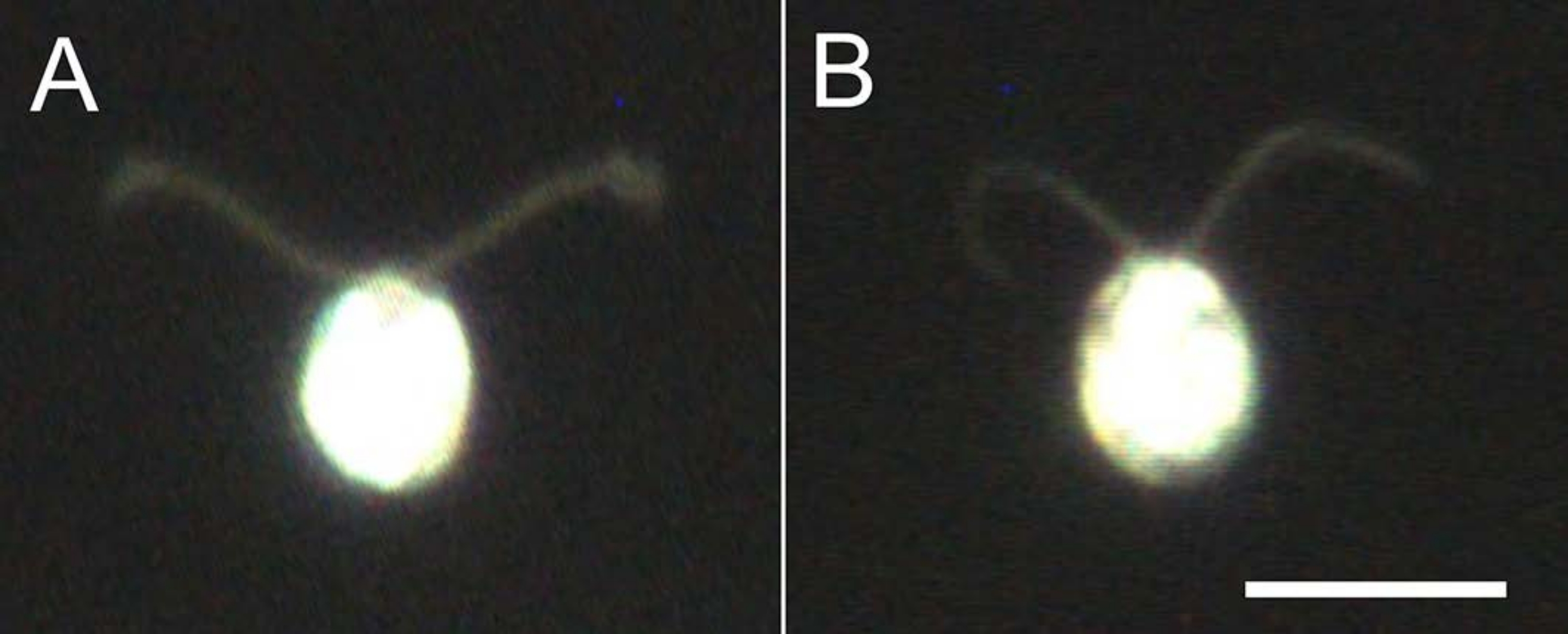
Рисунок 4: Эффекты демембранации. (А) Живая клетка, застрявшая на стеклянной горке. (B) Модель ячейки, наклеенная на стеклянный слайд. Обратите внимание, что в клеточной модели реснички стали немного тоньше. Изображения наблюдались под микроскопом с 20-кратным объективом и масляным погружным конденсатором темного поля. Шкала = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Фильм 2: Подтверждение демембранации. Суспензия клеточной модели наблюдалась под микроскопом с 10-кратной объективной линзой и масляным погружным конденсатором темного поля. Ни одна камера не плавала. Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот фильм.
3. Реактивация моделей демембранированных клеток
- Смешайте 80 мкл реактивационного раствора, 10 мкл раствора АТФ и 10 мкл клеточных моделей в пробирке объемом 0,5 мл, постукивая по трубке (рисунок 5).
ПРИМЕЧАНИЕ: Конечная концентрация АТФ должна составлять <3 мМ, поскольку частота цилиарного биения, являющаяся параметром цилиарного движения, увеличивается с концентрацией АТФ и насыщается при 2-3 мМАТФ-17.
ВНИМАНИЕ: Смешивание путем пипетирования или вихря вызывает дециляцию и снижает эффективность реактивации.- Для реактивации с <0,2 мМ АТФ добавляют систему регенерации АТФ, такую как 70 ЕД/мл креатинкиназы и 5 мМ креатинфосфата (см. Таблицу материалов).
ПРИМЕЧАНИЕ: Реактивационный раствор готовят в более высокой концентрации (1,125x, дополнительная таблица 1), чем раствор для разбавления, так что после смешивания АТФ, растворенного в воде, содержимое достигает следующих конечных концентраций: 30 мМ гепатита (рН 7,4), 5 мМ MgSO4, 1 мМ дитиотрейтола (DTT), 1 мМ EGTA и 50 мМ ацетата калия (см. Таблицу материалов).
- Для реактивации с <0,2 мМ АТФ добавляют систему регенерации АТФ, такую как 70 ЕД/мл креатинкиназы и 5 мМ креатинфосфата (см. Таблицу материалов).
- Нанесите ~30 мкл смешанного раствора на стеклянный предметный предмет и аккуратно поместите на него крышку с распорками, чтобы избежать механического удара о модели ячеек (рисунок 6).
ПРИМЕЧАНИЕ: Белые нефтяные или двусторонние клейкие ленты могут быть использованы для изготовления распорок. - Понаблюдайте за реактивированными моделями клеток под микроскопом (фильм 3).

Рисунок 5: Смешивание раствора путем постукивания по трубке. (А) До 80 мкл реактивационного раствора, 10 мкл раствора АТФ и 10 мкл моделей клеток добавляли в последовательном порядке. (B) Раствор смешивали путем постукивания по трубке пальцем. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6: Изготовление распорок по краям чехла. (А) Тонкий слой белой нефти наносили на тыльную сторону руки. (B) Небольшое количество белой нефти было соскоблено краем крышки. (C) Распорка была сделана на краю крышки. (D) Еще одна прокладка была сделана на противоположном краю. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Фильм 3: Плавание реактивированных моделей клеток. Подвижность клеточных моделей реактивировали путем добавления АТФ в конечной концентрации 1 мМ и наблюдали под микроскопом с 10-кратной объективной линзой и масляно-погружным конденсатором темного поля. Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот фильм.
Результаты
Процесс демембранации и реактивации в штамме дикого типа C. reinhardtii (CC-125) показан здесь. Культура через 2 дня после посева приобрела светло-зеленый цвет (этап 1.1) (рисунок 1). Клетки собирали (стадия 2.1), промывали (стадия 2.2) и демембранировали (стадия 2.5). После демембранац...
Обсуждение
В этом протоколе есть два важных шага. Первый — это процесс, известный как демембранация, который необходимо проводить мягко, но тщательно. Дециляция (т.е. отделение ресничек от тела клетки) индуцируется энергичным пипетированием или вихрем, что делает клеточные модели неподвижными даж...
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Это исследование было поддержано грантами Японского общества содействия развитию науки KAKENHI (https://www.jsps.go.jp/english/index.html) в Нью-Йорк (19K23758, 21K06295) и K.W. (19H03242, 20K21420, 21H00420), от Ohsumi Frontier Science Foundation (https://www.ofsf.or.jp/en/) до K.W. и от Динамического альянса за открытые инновации, соединяющие человека, окружающую среду и материалы (http://alliance.tagen.tohoku.ac.jp/english/) в N.U. и K.W. Мы благодарим г-жу Миюки Синохара (Университет Хосей) за ее помощь в подготовке цифр.
Материалы
| Name | Company | Catalog Number | Comments |
| 0.5 mL plastic tube | QSP | 502-PLN-Q | |
| 15 mL conical tube | SARSTEDT | 62.554.502 | |
| Adenosine 5'-triphosphate disodium salt hydrate (ATP) | Sima-Aldrich | A2383 | |
| Centrifuge | KUBOTA | 2800 | |
| Chlamydomonas strain CC-125 | Chlamydomonas Resource Center | https://www.chlamycollection.org/ | |
| Creatine kinase | Merck | CK-RO | |
| Creatine phosphate | Merck | CRPHO-RO | |
| Dithiothreitol (DTT) | Nakalai tesque | 14128-46 | |
| GEDTA(EGTA) | Dojindo | G002 | |
| Hepes | Dojindo | GB70 | |
| Igepal CA-630 | Sigma-Aldrich | I8896 | IUPAC name is octylphenoxypolyethoxyethanol: IGEPAL CA-630 is a substitute for Nonidet P-40 (NP-40); NP-40 is no longer available in Sigma-Aldrich. |
| MgSO4-7H2O | Nakalai tesque | 21002-85 | |
| Microscope | Olympus | BX-53 | |
| Pasteur pipette | fisher scientific | 13-678-20C | |
| Polyethylene glycol, Mr 20,000 | Merck | 8.18897.1000 | |
| Pottasium acetate | Nakalai tesque | 28434-25 | |
| Sodium Hydroxide | Nacalai | 31511-05 | |
| Sucrose | FUJIFILM Wako Pure Chemical Corporation | 196-00015 |
Ссылки
- Szent-Gyorgyi, A. Free-energy relations and contraction of actomyosin. Biological Bulletin. 96 (2), 140-161 (1949).
- Hoffman-Berling, H. Adenosintriphosphat als betriebsstoff von zellbewegungen. Biochimica et Biophysica Acta. 14, 182-194 (1954).
- Naitoh, Y., Kaneko, H. Reactivated Triton-extracted models of Paramecium: modification of ciliary movement by calcium ions. Science. 176 (4034), 523-524 (1972).
- Witman, G. B., Plummer, J., Sander, G. Chlamydomonas flagellar mutants lacking radial spokes and central tubules. Structure, composition, and function of specific axonemal components. Journal of Cell Biology. 76 (3), 729-747 (1978).
- Rüffer, U., Nultsch, W. Flagellar coordination in Chlamydomonas cells held on micropipettes. Cell Motility and the Cytoskeleton. 41 (4), 297-307 (1998).
- Sale, W. S., Satir, P. Direction of active sliding of microtubules in Tetrahymena cilia. Proceedings of the National Academy of Sciences of the United States of America. 74 (5), 2045-2049 (1977).
- Fox, L. A., Sale, W. S. Direction of force generated by the inner row of dynein arms on flagellar microtubules. Journal of Cell Biology. 105 (4), 1781-1787 (1987).
- Kamiya, R., Yagi, T. Functional diversity of axonemal dyneins as assessed by in vitro and in vivo motility assays of Chlamydomonas mutants. Zoolog Science. 31 (10), 633-644 (2014).
- Kamiya, R., Witman, G. B. Submicromolar levels of calcium control the balance of beating between the two flagella in demembranated models of Chlamydomonas. Journal of Cell Biology. 98 (1), 97-107 (1984).
- Okita, N., Isogai, N., Hirono, M., Kamiya, R., Yoshimura, K. Phototactic activity in Chlamydomonas 'non-phototactic' mutants deficient in Ca2+-dependent control of flagellar dominance or in inner-arm dynein. Journal of Cell Science. 118, 529-537 (2005).
- Hyams, J. S., Borisy, G. G. Isolated flagellar apparatus of Chlamydomonas: characterization of forward swimming and alteration of waveform and reversal of motion by calcium ions in vitro. Journal of Cell Science. 33, 235-253 (1978).
- Fujiu, K., Nakayama, Y., Yanagisawa, A., Sokabe, M., Yoshimura, K. Chlamydomonas CAV2 encodes a voltage- dependent calcium channel required for the flagellar waveform conversion. Current Biology. 19 (2), 133-139 (2009).
- Bessen, M., Fay, R. B., Witman, G. B. Calcium control of waveform in isolated flagellar axonemes of Chlamydomonas. Journal of Cell Biology. 86 (2), 446-455 (1980).
- Wakabayashi, K., King, S. M. Modulation of Chlamydomonas reinhardtii flagellar motility by redox poise. Journal of Cell Biology. 173 (5), 743-754 (2006).
- Saegusa, Y., Yoshimura, K. cAMP controls the balance of the propulsive forces generated by the two flagella of Chlamydomonas. Cytoskeleton. 72 (8), 412-421 (2015).
- Gorman, D. S., Levine, R. P. Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proceedings of the National Academy of Sciences of the United States of America. 54 (6), 1665-1669 (1965).
- Takano, W., Hisabori, T., Wakabayashi, K. Rapid estimation of cytosolic ATP concentration from the ciliary beating frequency in the green alga Chlamydomonas reinhardtii. Journal of Biological Chemistry. 296, 100156 (2021).
- Wakabayashi, K., Yagi, T., Kamiya, R. Ca2+-dependent waveform conversion in the flagellar axoneme of Chlamydomonas mutants lacking the central-pair/radial spoke system. Cell Motility and the Cytoskeleton. 38 (1), 22-28 (1997).
- Yueh, Y. G., Crain, R. C. Deflagellation of Chlamydomonas reinhardtii follows a rapid transitory accumulation of inositol 1,4,5-trisphosphate and requires Ca2+ entry. Journal of Cell Biology. 123 (4), 869-875 (1993).
- Wakabayashi, K., Ide, T., Kamiya, R. Calcium-dependent flagellar motility activation in Chlamydomonas reinhardtii in response to mechanical agitation. Cell Motility and the Cytoskeleton. 66 (9), 736-742 (2009).
- Ueki, N., Wakabayashi, K. Detergent-extracted Volvox model exhibits an anterior-posterior gradient in flagellar Ca2. Proceedings of the National Academy of Sciences of the United States of America. 115 (5), 1061-1068 (2018).
- Tanno, A., et al. The four-celled Volvocales green alga Tetrabaena socialis exhibits weak photobehavior and high-photoprotection ability. PLoS One. 16 (10), 0259138 (2021).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены