A subscription to JoVE is required to view this content. Sign in or start your free trial.
Spatiotemporal Subcellular Manipulation of the Microtubule Cytoskeleton in the Living Preimplantation Mouse Embryo using Photostatins
In This Article
Summary
Typical microtubule inhibitors, used widely in basic and applied research, have far-reaching effects on cells. Recently, photostatins emerged as a class of photoswitchable microtubule inhibitors, capable of instantaneous, reversible, spatiotemporally precise manipulation of microtubules. This step-by-step protocol details the application of photostatins in a 3D live preimplantation mouse embryo.
Abstract
The microtubule cytoskeleton forms the framework of a cell and is fundamental for intracellular transport, cell division, and signal transduction. Traditional pharmacological disruption of the ubiquitous microtubule network using, for instance, nocodazole can have devastating consequences for any cell. Reversibly photoswitchable microtubule inhibitors have the potential to overcome the limitations by enabling drug effects to be implemented in a spatiotemporally-controlled manner. One such family of drugs is the azobenzene-based photostatins (PSTs). These compounds are inactive in dark conditions, and upon illumination with UV light, they bind to the colchicine-binding site of β-tubulin and block microtubule polymerization and dynamic turnover. Here, the application of PSTs in the 3-dimensional (3D) live preimplantation mouse embryo is set out to disrupt the microtubule network on a subcellular level. This protocol provides instructions for the experimental setup, as well as light activation and deactivation parameters for PSTs using live-cell confocal microscopy. This ensures reproducibility and enables others to apply this procedure to their research questions. Innovative photoswitches like PSTs may evolve as powerful tools to advance the understanding of the dynamic intracellular microtubule network and to non-invasively manipulate the cytoskeleton in real-time. Furthermore, PSTs may prove useful in other 3D structures such as organoids, blastoids, or embryos of other species.
Introduction
The microtubule architecture varies widely across different cell types to support diverse functions1,2. Its dynamic nature of growth and shrinkage allows rapid adaptation to extra- and intracellular cues and to respond to the ever-changing needs of a cell. Hence, it can be considered as the "morphological fingerprint" playing a key role in cellular identity.
Pharmacological targeting of the microtubule cytoskeleton using small molecule inhibitors has led to a plethora of fundamental discoveries in developmental biology, stem cell biology, cancer biology, and neurobiology3,4,5,6,7. This approach, while indispensable, presents various limitations such as toxicity and off-target effects. For example, one of the most widely used microtubule-targeting agents, nocodazole, is a powerful microtubule-depolymerizing drug8. However, small-molecule inhibitors such as nocodazole are active from the time of application and, given the essential nature of the microtubule cytoskeleton to many critical cellular functions, global depolymerization of microtubules can produce off-target effects, which may be unsuitable for many applications. Additionally, nocodazole treatment is irreversible unless samples are washed free of the drug, preventing continuous live imaging and, thus, precise tracking of individual microtubule filaments.
The development of light-activated compounds began with the creation of photouncaged molecules and has heralded a new era in targeting and monitoring the effects of microtubule growth inhibition in a precise and spatiotemporally-controlled manner. One family of reversibly photoswitchable drugs, photostatins (PSTs), were developed by replacing the stilbene component of combretastatin A-4 with azobenzene9. PSTs are inactive until illumination with UV light, whereby the inactive trans-configuration converts to the active cis-configuration by reversible isomerization. Cis-PSTs inhibit microtubule polymerization by binding to the colchicine binding site of β-tubulin, blocking its interface with β-tubulin and preventing dimerization required for microtubule growth10. Among a cohort of PSTs, PST-1P has emerged as a lead compound as it has the highest potency, is fully water-soluble, and shows a rapid onset of bioactivity after illumination.
The most effective trans- to cis-isomerization of PSTs occurs at wavelengths between 360-420 nm, which enables dual options for PST activation. A 405 nm laser line on a typical confocal microscope can be administered for optimal spatial targeting of microtubule growth inhibition. The ability to pinpoint the location and timing of PST activation through 405 nm laser illumination facilitates precise temporal and spatial control, allowing disruption of microtubule dynamics on a subcellular level, within sub-second response times9. Alternatively, an affordable LED UV light allows whole organism illumination to induce organism-wide disruption of the microtubule architecture. This may be a cost-effective alternative for researchers for whom the precisely-timed onset of inhibition, rather than spatial targeting, is the goal. Another feature of PSTs is their on-demand inactivation by applying green light of a wavelength in the 510-540 nm range9. This enables tracing of microtubule filaments before, during, and after PST-mediated growth inhibition.
PSTs, while still a relatively recent design, have been used in numerous in vitro applications across diverse research fields11, including investigating new mechanisms of cell migration in amoeboids12, in neurons isolated from the brain of the newborn mouse13, and wing epithelium development in Drosophila melanogaster14. Other light-reactive drugs have proven to be valuable tools in targeted disruption of cellular function. For instance, an analog of blebbistatin, azidoblebbistatin, was used for enhanced myosin inhibition under illumination15,16. This highlights the potential for new discoveries owing to the ability for spatiotemporally controlled inhibition of cellular function.
Live 3D organisms present superb yet more delicate systems to manipulate microtubule dynamics on a whole-animal, single-cell, or subcellular level under physiological conditions. In particular, the preimplantation mouse embryo offers exceptional insight into the inner workings of the cell as well as intercellular relationships within an organism17. Temporally and spatially targeted consecutive cycles of activation and deactivation of PSTs contributed to the characterization of the interphase bridge, a post-cytokinetic structure between cells, as a non-centrosomal microtubule-organizing center in the preimplantation mouse embryo16. A similar experimental setup demonstrated the involvement of growing microtubules in the sealing of the mouse embryo to allow blastocyst formation18. Furthermore, PSTs were also used in whole zebrafish embryos to investigate neuronal cell migration by inhibiting microtubule growth in a subset of cells in the hindbrain19.
This protocol describes the experimental setup and use of PST-1P in the preimplantation mouse embryo. The instructions presented here can also guide the application of PSTs for a wide array of objectives such as studying chromosome segregation and cell division, trafficking of intracellular cargo, and cell morphogenesis and migration. Furthermore, such studies will assist the implementation of PSTs in organoid systems, blastoids, and other embryo models such as Caenorhabditis elegans and Xenopus laevis, as well as potentially expand the use of PSTs for in vitro fertilization technologies.
Protocol
Experiments were approved by the Monash Animal Ethics Committee under animal ethics number 19143. Animals were housed in specific pathogen-free animal house conditions at the animal facility (Monash Animal Research Platform) in strict accordance with ethical guidelines.
1. Preimplantation mouse embryo collection
- Superovulate and mate mice as described previously16,18, in compliance with the institutional animal ethics guidelines.
NOTE: The most commonly used strains for live embryo collection are C57BL/6 or FVB/N mice. All data shown here were generated using FVB/N mice. - On the morning after mating, flush zygotes from the oviduct using M2 medium as described20, or Human Tubal Fluid (HTF) medium. Using a mouth pipette apparatus as described21,22, transfer zygotes to fresh Potassium Simplex Optimised Medium (KSOM) droplets, prewarmed to 37 °C and equilibrated to 5% CO2, in a 35 mm culture dish overlaid with a sufficient volume of mineral oil to ensure media coverage.
- Microinject zygotes as described20 with cRNA encoding for a red fluorescently-tagged microtubule plus end marker. Here, cRNA for the End Binding protein 3 (EB3)-dTomato was used at 30 ng/µL concentration after preparing and purifying as described16,18 and diluting in microinjection buffer.
NOTE: It is recommended to prepare cRNA ahead of time and store it at -20 °C until required. - Culture embryos in dark at 37 °C and 5% CO2 until embryos have reached the desired developmental stage for PST-1P treatment.
NOTE: For a comprehensive resource of the culture times required for different embryonic stages see23. For 16-cell stage embryos used here, culture to embryonic day 3 (E3) post-fertilization.
2. Drug and imaging dish preparation
NOTE: For Steps 2.1-2.10, work exclusively in the dark or red-light conditions to avoid unintended PST-1P activation. Aluminum foil or dark covers should be used for all tubes and dishes containing PSTs.
- Prepare a stock concentration of 50 mM PST-1P in ultrapure water.
NOTE: The molecular weight of PST-1P is 440 g/mol. The stock solution is stable at -20 °C for up to 1 year. PST-1P is soluble in water or aqueous buffer but does not readily dissolve in DMSO. - From step 2.1, prepare an intermediate working concentration of 800 µM PST-1P in ultrapure water.
- Dilute PST-1P to a final concentration of 40 µM in fresh KSOM. As a typical experiment requires approximately 20 µL of PST-1P-treated KSOM, dilute 1 µL of 800 µM PST-1P in 19 µL of KSOM for a final volume of 20 µL to ensure sufficient medium is prepared in advance, using only red light for visibility.
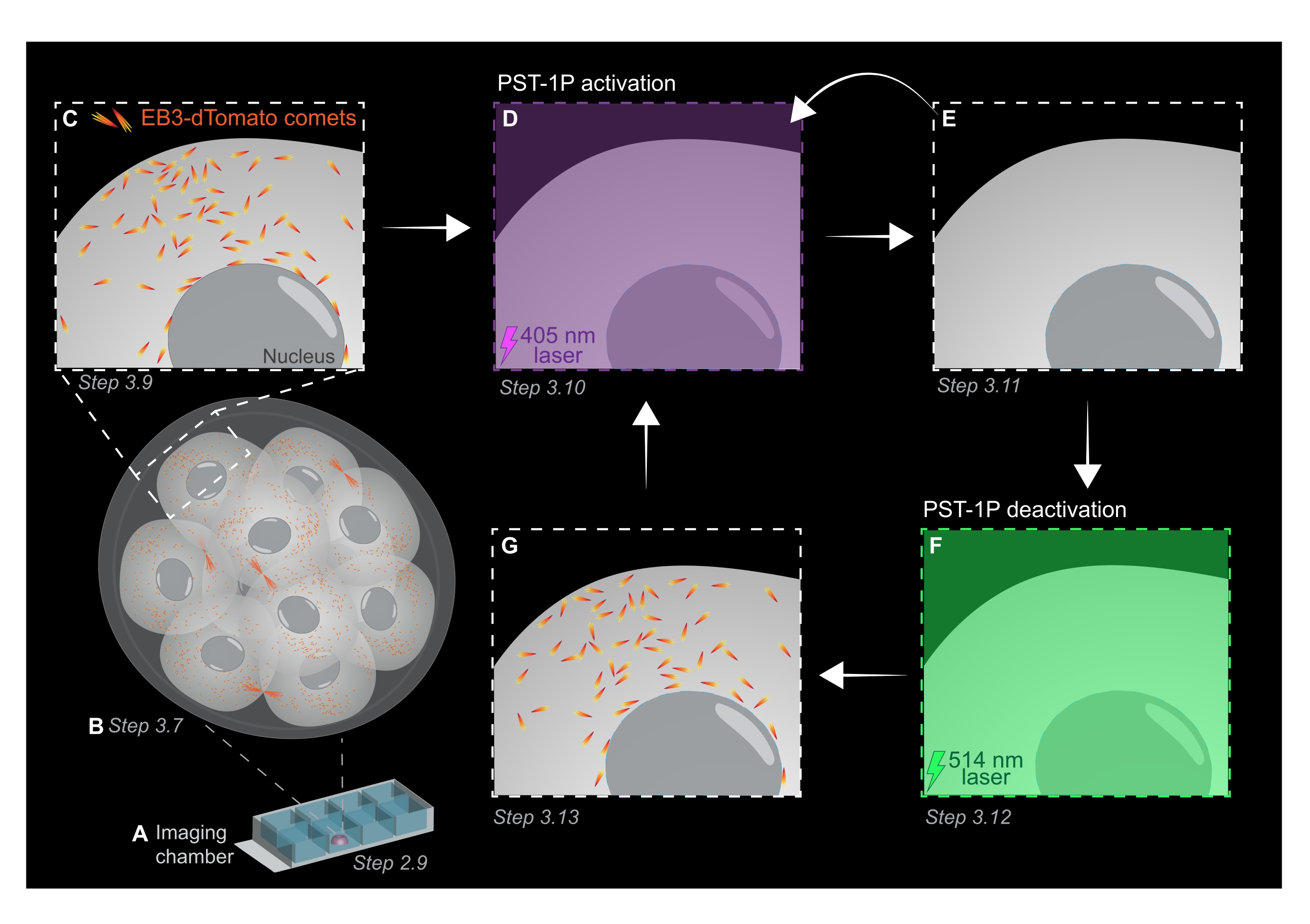
NOTE: Both the concentration and the activation status of a PST-1P dilution can be checked by taking a UV-Vis absorbance spectrum using a spectrophotometer which should be done during assay establishment. The absorbance at 380 nm (A380) of a fully-trans 40 µM dilution in a 1 cm cuvette should be approximately 0.8. Both cis- and trans-forms have the same absorbance at 455 nm (A455). When the ratio of the A380 to the A455 is approximately 9:1, the dilution is inactive (fully trans). When the ratio A380:A455 is 1:2, the dilution is fully activated (fully cis). Intermediate ratios reflect intermediate states of activation. - To prepare the chamber slide for live imaging, pipette 10 µL of PST-1P-treated KSOM into the center of one well to form a hemispherical droplet (Figure 1A).
- Gently add sufficient mineral oil to cover the droplet, ensuring that it does not disperse the media. This will ensure that the droplet does not evaporate.
- Pre-warm and CO2-equilibrate the chamber slide dish in an incubator at 37 °C and 5% CO2 for a minimum of 3 h or at most, overnight.
- At the end of the equilibration period in step 2.6, prepare a 35 mm culture dish with a 10 µL droplet of PST-1P-treated, prewarmed, and equilibrated KSOM as a wash step. Do not overlay with oil.
- Transfer embryos by mouth pipetting into the PST-1P-treated KSOM droplet from step 2.7.
NOTE: Steps 2.7 and 2.8 are recommended for mouth pipetting of embryos but are optional. - Immediately transfer embryos by mouth pipetting into the center of the PST-1P-treated KSOM droplet in the imaging chamber slide prepared in steps 2.4-2.6 (Figure 1A).
- Incubate embryos at 37 °C and 5% CO2 in PST-1P-treated KSOM in the imaging chamber slide for at least 1 h before imaging. If possible, mount the chamber slide on the microscope inside an environmental chamber at 37 °C and 5% CO2, and in complete darkness to ensure all PST-1Ps are in the inactive trans-configuration and that embryos can sink to the bottom of the dish.
3. Live imaging and PST-1P photoactivation
NOTE: Steps 3.1-3.13 are performed on a laser scanning confocal microscope fitted with avalanche photodiode detectors (APDs) and a dark environmental chamber. These instructions refer specifically to the imaging setup using the acquisition software described in the Table of Materials; however, they can also be applied to other confocal microscopy systems.
- Prepare a 63x/1.2 NA water oil immersion objective with the prescribed immersion medium.
- Using a red light torch to guide positioning, advance the objective to contact the immersion medium. At this stage, avoid using white or brightfield light to find the embryos as this could precociously activate PST-1P.
- Using the eyepiece and while under red light illumination, locate the edge of the droplet of medium and position the objective directly over this location. This can assist the user to establish orientation and find the focal plane.
- Next, through the eyepiece or on software-enabled live-mode scanning, use a red wavelength filter or 561 nm laser to locate the embryos within the droplet.
- Use stage controllers and live-scanning mode to set the start- and end-points for acquiring a z-stack of the whole embryo.
- Adjust the laser power settings (typically, with highly sensitive detectors such as the APDs, a 561 nm laser power of less than 5% is sufficient), digital offset (typically at -0.900) to optimize the appearance of EB3-dTomato comets and minimize background noise, pinhole at 2 µm, pixel resolution of 512 x 512, and pixel dwell time of 3.15 µs.
- Acquire a z-stack of the whole embryo with 1 µm section intervals to assess areas of microtubule growth in the whole organism (Figure 1B).
- Use the 3D z-stack image from step 3.7 to identify regions of interest (ROIs) for EB3-dTomato tracking experiments. Increase the zoom to 3x and draw a rectangular ROI around the specific subcellular area of interest.
- Acquire a time-lapse movie of a single z-plane using the typical values of imaging parameters: 561 nm laser power at 5%, digital offset of -0.900, pixel resolution of 512 x 512, pinhole at 3 µm, pixel dwell time of 3.15 µs, zoom of 3x, time interval of 500 ms.
NOTE: 120 time frames will provide a tracking movie of 1 min and should be sufficient for data analysis. The acquisition can continue for longer, provided that bleaching of the fluorophore is minimal (Figure 1C). - To activate PST-1P, switch to a 405 nm laser and acquire another time-lapse movie with the 405 nm laser set to 10% power, pixel resolution of 512 x 512, pinhole opened maximally, pixel dwell time of 3.15 µs, zoom of 3x, time interval of 500 ms, and a total of 20 frames (Figure 1D).
- Switch back to 561 nm laser and repeat acquisition as in step 3.9 to confirm the loss of EB3-dTomato comets after activation of PST-1P (Figure 1E). Ensure that this acquisition takes place as soon as possible following activation.
NOTE: Step 3.10 can be performed repetitively for longer inhibition of EB3-dTomato comets. However, embryos must be monitored carefully to avoid any harm caused by UV light. - To reverse PST-1P back to its inactive trans-state, engage a 514 nm laser at 10% power. Acquire a time-lapse movie with the 514 nm laser set to 10% power, pixel resolution of 512 x 512, pinhole opened maximally, pixel dwell time of 3.15 µs, zoom of 3x, time interval of 500 ms, and a total of 20 time frames (Figure 1F).
- Repeat step 3.11 to visualize recovery of EB3-dTomato comets (Figure 1G).
4. Image data analysis
- For analyzing and quantifying the inhibition of microtubule polymerization by PSTs, use software programs available to researchers to suit their specific needs. Those recommended for use will possess a tracking tool, that can manually or automatically track the movement, direction, and speed of EB3-dTomato comets16,18.

Figure 1: Schematic representation of PST-1P photoactivation and deactivation in the live 3D preimplantation mouse embryo. All experiments are performed in complete darkness (black background) or only by red light illumination. (A) Live preimplantation mouse embryos expressing EB3-dTomato are cultured to 16-cell stage and then transferred to a droplet of KSOM containing 40 µM PST-1P in an imaging chamber slide. (B) A 3D image of the whole embryo allows the assessment of microtubule growth by visualizing the distribution of EB3-dTomato comets. (C) To start the experiment, EB3-dTomato comets are tracked in a subcellular region using time-lapse imaging. (D) Subsequent PST-1P photoactivation in the same subcellular region using a 405 nm laser results in the loss of EB3-dTomato comets (E). Intensified PST-1P activation can be implemented, if necessary, by sequential 405 nm light illumination. (F-G) To reverse PST-1P back to its inactive state and restore EB3-dTomato comets, a 514 nm laser is applied to the same subcellular region. If required, multiple rounds of photoactivation and deactivation can be performed. Please click here to view a larger version of this figure.
{kind=link}
Results
In line with the protocol, preimplantation mouse embryos were microinjected with cRNA for EB3, tagged with red fluorescent dTomato (EB3-dTomato). This enables the visualization of growing microtubules as EB3 binds to polymerizing microtubule plus ends24.
The experiments were performed 3 days post-fertilisation (E3) when the mouse embryo is comprised of 16 cells. Any other preimplantation developmental stage can be used, depending on the scientific question to be investi...
Discussion
The microtubule network is integral to the fundamental inner workings of a cell. Consequently, this presents challenges in manipulating microtubule dynamics in living organisms, as any perturbation to the network tends to have widespread consequences for all aspects of cellular function. The emergence of photoswitchable microtubule-targeting compounds presents a way to precisely manipulate the cytoskeleton at a subcellular level, with superior control for the induction and reversal of microtubule growth inhibition
Disclosures
The authors declare no competing or financial interest.
Acknowledgements
The authors would like to thank Dr. Oliver Thorn-Seshold and Li Gao for providing us with photostatins and advice on manuscript preparation, Monash Production for filming support, and Monash Micro Imaging for microscopy support.
This work was supported by the National Health and Medical Research Council (NHMRC) project grant APP2002507 to J.Z. and the Canadian Institute for Advanced Research (CIFAR) Azrieli Scholarship to J.Z. The Australian Regenerative Medicine Institute is supported by grants from the State Government of Victoria and the Australian Government.
Materials
| Name | Company | Catalog Number | Comments |
| Aspirator tube | Sigma-Aldrich | A5177 | For mouth aspiration apparatus |
| Chamber slides - LabTek | Thermo Fisher Scientific | NUN155411 | |
| cRNA encoding for EB3-dTomato | N/A | N/A | Prepared according to manufacturers instructions using mMessage in vitro Transcription kit |
| Culture dishes - 35mm | Thermo Fisher Scientific | 150560 | |
| Human chorionic growth hormone | Sigma-Aldrich | C8554 | |
| Human Tubal Fluid (HTF) medium | Cosmo-Bio | CSR-R-B071 | |
| Imaris Image Analysis Software | Bitplane | ||
| Immersion Oil W 2010 | Carl Zeiss | 444969-0000-000 | For use with microscope immersion objective |
| LED torch - Red light | Celestron | 93588 | |
| M2 medium | Sigma-Aldrich | M7167 | |
| Mice - wild-type FVB/N, males and females | N/A | N/A | Females 8-9 weeks old. Males 2-6 months old. |
| Microcapillary Pipettes - Kimble | Sigma-Aldrich | Z543306 | For mouth aspiration apparatus |
| Microinjection buffer | N/A | N/A | 5 mM Tris, 5 mM NaCl, 0.1 mM EDTA, pH 7.4 |
| Mineral oil | Origio | ART-4008-5P | |
| mMessage In vitro Transcription kit | Thermo Fisher Scientific | AM1340 | |
| NanoDrop Spectrophotometer | Thermo Fisher Scientific | ||
| Potassium Simplex Optimised Medium (KSOM) medium | Cosmo-Bio | CSR-R-B074 | |
| Pregnant mare serum gonadotrophin | Prospec Bio | HOR-272 | |
| PST-1P | N/A | N/A | Borowiak, M. et al., Photoswitchable Inhibitors of Microtubule Dynamics Optically Control Mitosis and Cell Death. Cell. 162 (2), 403-411, doi:10.1016/j.cell.2015.06.049, (2015). |
| RNA purification kit | Sangon | B511361-0100 | |
| Ultrapure water | Sigma-Aldrich | W1503 | |
| ZEN Black Software | Carl Zeiss |
References
- Hawdon, A., Aberkane, A., Zenker, J. Microtubule-dependent subcellular organisation of pluripotent cells. Development. 148 (20), (2021).
- Sanchez, A. D., Feldman, J. L. Microtubule-organizing centers: from the centrosome to non-centrosomal sites. Current Opinion in Cell Biology. 44, 93-101 (2017).
- Galli, M., Morgan, D. O. Cell size determines the strength of the spindle assembly checkpoint during embryonic development. Developmental Cell. 36 (3), 344-352 (2016).
- Vazquez-Diez, C., Paim, L. M. G., FitzHarris, G. Cell-size-independent spindle checkpoint failure underlies chromosome segregation error in mouse embryos. Current Biology. 29 (5), 865-873 (2019).
- Baudoin, J. P., Alvarez, C., Gaspar, P., Metin, C. Nocodazole-induced changes in microtubule dynamics impair the morphology and directionality of migrating medial ganglionic eminence cells. Developmental Neuroscience. 30 (1-3), 132-143 (2008).
- Munz, F., et al. Human mesenchymal stem cells lose their functional properties after paclitaxel treatment. Scientific Reports. 8 (1), 312 (2018).
- Jordan, M. A., Wilson, L. Microtubules as a target for anticancer drugs. Nature Reviews Cancer. 4 (4), 253-265 (2004).
- Vasquez, R. J., Howell, B., Yvon, A. M., Wadsworth, P., Cassimeris, L. Nanomolar concentrations of nocodazole alter microtubule dynamic instability in vivo and in vitro. Molecular Biology of the Cell. 8 (6), 973-985 (1997).
- Borowiak, M., et al. Photoswitchable inhibitors of microtubule dynamics optically control mitosis and cell death. Cell. 162 (2), 403-411 (2015).
- Gaspari, R., Prota, A. E., Bargsten, K., Cavalli, A., Steinmetz, M. O. Structural basis of cis- and trans-Combretastatin binding to tubulin. Chem. 2 (1), 102-113 (2017).
- Thorn-Seshold, O., Meiring, J. Photocontrolling microtubule dynamics with photoswitchable chemical reagents. ChemRxiv. , (2021).
- Kopf, A., et al. Microtubules control cellular shape and coherence in amoeboid migrating cells. Journal of Cell Biology. 219 (6), 201907154 (2020).
- Sawada, M., et al. PlexinD1 signaling controls morphological changes and migration termination in newborn neurons. The EMBO Journal. 37 (4), 97404 (2018).
- Singh, A., et al. Polarized microtubule dynamics directs cell mechanics and coordinates forces during epithelial morphogenesis. Nature Cell Biology. 20 (10), 1126-1133 (2018).
- Kepiro, M., et al. Azidoblebbistatin, a photoreactive myosin inhibitor. Proceedings of the National Academy of Sciences of the United States of America. 109 (24), 9402-9407 (2012).
- Zenker, J., et al. A microtubule-organizing center directing intracellular transport in the early mouse embryo. Science. 357 (6354), 925-928 (2017).
- White, M. D., Zenker, J., Bissiere, S., Plachta, N. Instructions for assembling the early mammalian embryo. Developmental Cell. 45 (6), 667-679 (2018).
- Zenker, J., et al. Expanding actin rings zipper the mouse embryo for blastocyst formation. Cell. 173 (3), 776-791 (2018).
- Theisen, U., et al. Microtubules and motor proteins support zebrafish neuronal migration by directing cargo. Journal of Cell Biology. 219 (10), 201908040 (2020).
- Rulicke, T. Pronuclear microinjection of mouse zygotes. Methods in Molecular Biology. 254, 165-194 (2004).
- Greaney, J., Subramanian, G. N., Ye, Y., Homer, H. Isolation and in vitro culture of mouse oocytes. Bio-protocol. 11 (15), 4104 (2021).
- Subramanian, G. N., et al. Oocytes mount a noncanonical DNA damage response involving APC-Cdh1-mediated proteolysis. Journal of Cell Biology. 219 (4), 201907213 (2020).
- Mihajlovic, A. I., Bruce, A. W. The first cell-fate decision of mouse preimplantation embryo development: integrating cell position and polarity. Open Biology. 7 (11), 170210 (2017).
- Roostalu, J., et al. The speed of GTP hydrolysis determines GTP cap size and controls microtubule stability. Elife. 9, 51992 (2020).
- Gao, L., et al. In vivo photocontrol of microtubule dynamics and integrity, migration and mitosis, by the potent GFP-imaging-compatible photoswitchable reagents SBTubA4P and SBTub2M. BioRxiv. bioRxiv. , (2021).
- Tichy, A. M., Gerrard, E. J., Legrand, J. M. D., Hobbs, R. M., Janovjak, H. Engineering strategy and vector library for the rapid generation of modular light-controlled protein-protein interactions. Journal of Molecular Biology. 431 (17), 3046-3055 (2019).
- van Haren, J., Adachi, L. S., Wittmann, T. Optogenetic control of microtubule dynamics. Methods in Molecular Biology. 2101, 211-234 (2020).
- Adikes, R. C., Hallett, R. A., Saway, B. F., Kuhlman, B., Slep, K. C. Control of microtubule dynamics using an optogenetic microtubule plus end-F-actin cross-linker. Journal of Cell Biology. 217 (2), 779-793 (2018).
- Kogler, A. C., et al. Extremely rapid and reversible optogenetic perturbation of nuclear proteins in living embryos. Developmental Cell. 56 (16), 2348-2363 (2021).
- Maghelli, N., Tolic-Norrelykke, I. M. Laser ablation of the microtubule cytoskeleton: setting up and working with an ablation system. Methods in Molecular Biology. 777, 261-271 (2011).
- Bukhari, S. N. A., Kumar, G. B., Revankar, H. M., Qin, H. L. Development of combretastatins as potent tubulin polymerization inhibitors. Bioorganic Chemistry. 72, 130-147 (2017).
- Gilazieva, Z., Ponomarev, A., Rutland, C., Rizvanov, A., Solovyeva, V. Promising applications of tumor spheroids and organoids for personalized medicine. Cancers (Basel). 12 (10), 2727 (2020).
- Scherer, K. M., et al. Three-dimensional imaging and uptake of the anticancer drug combretastatin in cell spheroids and photoisomerization in gels with multiphoton excitation. Journal of Biomedical Optics. 20 (7), 78003 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved