A subscription to JoVE is required to view this content. Sign in or start your free trial.
Real-Time Detection of Reactive Oxygen Species Production in Immune Response in Rice with a Chemiluminescence Assay
In This Article
Summary
Here, we describe a method for the real-time detection of apoplastic reactive oxygen species (ROS) production in rice tissues in pathogen-associated molecular pattern-triggered immune response. This method is simple, standardized, and generates highly reproducible results under controlled conditions.
Abstract
Reactive oxygen species (ROS) play vital roles in a variety of biological processes, including the sensing of abiotic and biotic stresses. Upon pathogen infection or challenge with pathogen-associated chemicals (pathogen-associated molecular patterns [PAMPs]), an array of immune responses, including a ROS burst, are quickly induced in plants, which is called PAMP-triggered immunity (PTI). A ROS burst is a hallmark PTI response, which is catalyzed by a group of plasma membrane-localized NADPH oxidases-the RBOH family proteins. The vast majority of ROS comprise hydrogen peroxide (H2O2), which can be easily and steadily detected by a luminol-based chemiluminescence method. Chemiluminescence is a photon-producing reaction in which luminol, or its derivative (such as L-012), undergoes a redox reaction with ROS under the action of a catalyst. This paper describes an optimized L-012-based chemiluminescence method to detect apoplast ROS production in real-time upon PAMP elicitation in rice tissues. The method is easy, steady, standardized, and highly reproducible under firmly controlled conditions.
Introduction
Reactive oxygen species (ROS) comprise a series of chemically active oxygen derivatives, including superoxide anion radicals (O2-) and its derivatives, hydroxyl radicals (OH-), hydrogen peroxide, and products of singlet oxygen or oxidation-reduction reactions, which are constantly produced in plastids and chloroplasts, mitochondria, peroxisomes, and other subcellular locations1. ROS play important roles in many biological processes and are essential for all plants2,3,4. The broad spectrum of ROS functions varies from the regulation of growth and development to the perception of abiotic and biotic stresses5,6,7,8.
In the plant immune system, plant cell plasma membrane-localized receptors-so-called pattern recognition receptors (PRRs)-perceive pathogen-derived chemicals-pathogen-associated molecular patterns (PAMPs). This recognition triggers a series of fast immune responses, including calcium influx, ROS burst, and MAPK cascade; thus, this layer of immunity is named PAMP-triggered immunity (PTI). ROS burst is a hallmark PTI response, the determination of which is widely applied to PTI-related studies9,10. ROS production triggered by PAMPs is attributed to plasma membrane-resident NADPH oxidase, or respiratory burst oxidase homolog (RBOH) family proteins, which transfer electrons from cytosolic NADPH or NADH to extracellular oxygen to produce superoxide (O2-) which is spontaneously converted to hydrogen peroxide (H2O2) by superoxide dismutase8. PAMP-triggered ROS burst is quite rapid, appearing only a few minutes after PAMP treatment and peaking at ~10-12 min. The vast majority of the ROS molecules comprise hydrogen peroxide (H2O2), which can be easily and steadily detected with a chemiluminescence assay.
In chemiluminescence, the chemiluminescence reagent reacts with active oxygen, under the action of a catalyst, to produce the excited state intermediates. Then, the electrons in the product return to the ground state through non-radiative transition and emit photons. Common chemiluminescence reagents include luminol and L-012, with luminol dominating the application11,12,13. However, more researchers are choosing L-012 to detect ROS production, since L-012 has a much higher light emission efficiency under neutral or near neutral pH conditions compared to luminol.
This paper describes an optimized chemiluminescence method, based on L-012, for the real-time detection of ROS production after the elicitation of PAMPs in rice (Oryza sativa) tissues-leaf discs and sheath. The method provided here is simple, stable, and standardized, and is highly adaptable to meet different experimental needs. The data obtained with this method are highly reproducible under firmly controlled conditions.
Protocol
NOTE: The protocol is applicable to different plant tissues. Rice sheath and leaf discs were used in this protocol for ROS detection upon PAMP elicitation. As differences mainly arise due to the method of sampling, only the common procedures are described below, with specific steps being mentioned wherever necessary.
1. Plant culture
- Sterilize the dehusked rice seeds with 70% ethanol for 1 min, then with 40% sodium hypochlorite (NaClO) for 1 h. Then, rinse the seeds 5x with sterile water to remove residual chlorine.
- Plate the seeds aseptically on 1/2 MS medium (2.37 g/L Murashige and Skoog (MS) medium, 30 g/L sucrose, 2.1 g/L phytagel, pH 5.7, autoclaved).
- In the rice sheath method, directly plate the seeds in the sterile glass vessel with MS medium.
- In the leaf disc method, plate the seeds on MS plates for 5-7 days and transplant them to growth matrix or soil (Figure 1A).
- Grow the seedlings in a growth room with a 12 h light/12 h dark photoperiod.
2. Tissue preparation and pretreatment
- Rice sheath
- Cut the sheath from 10-day-old rice seedlings into 3 mm segments with a sharp razor blade or surgical blade for pretreatment 1 day before the ROS assay (Figure 1B).
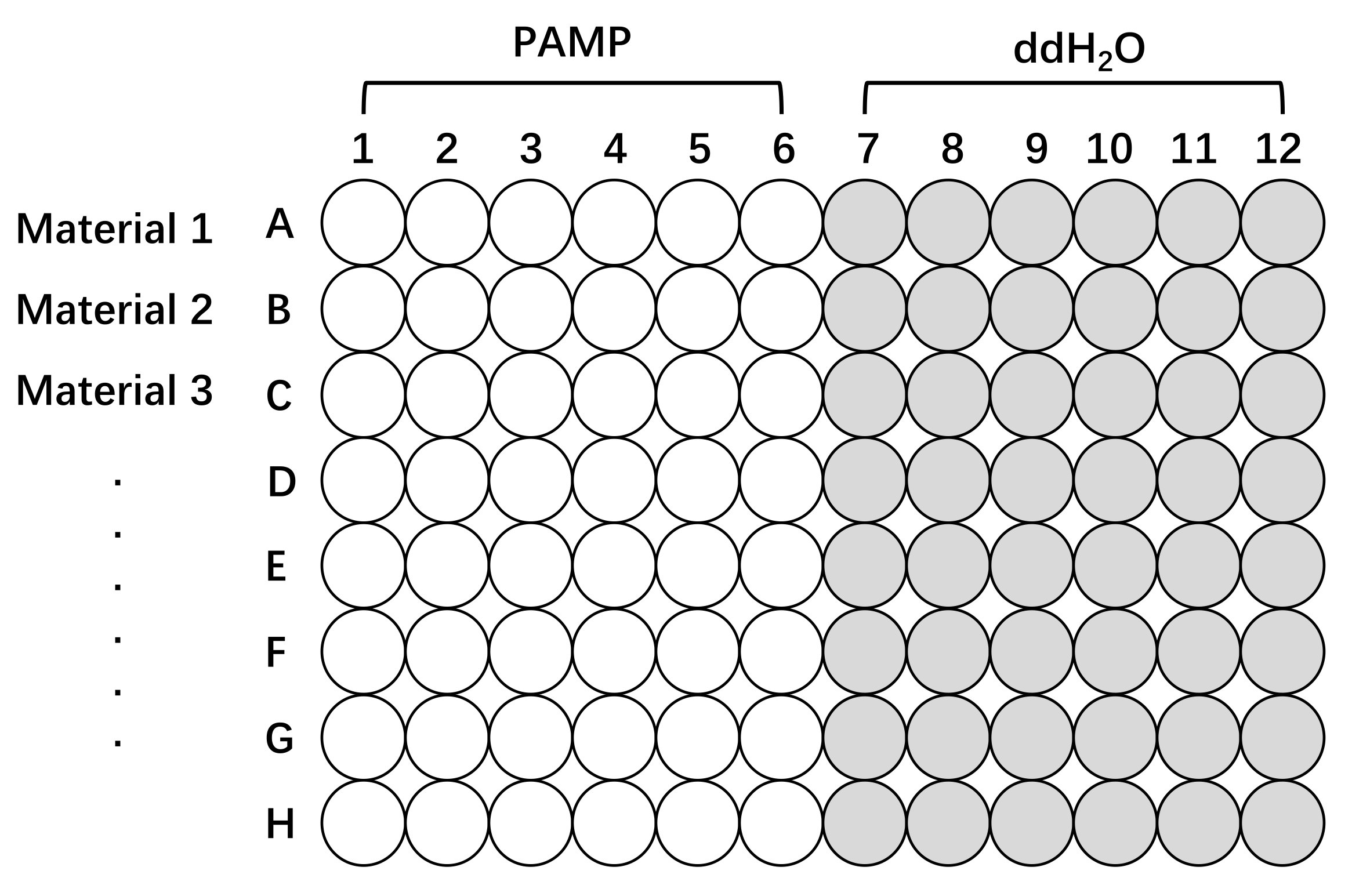
- Place five sheath segments in an individual well of a 96-well microtiter plate containing 100 µL of ddH2O for 10-12 h, in the dark at 25 °C, which allows wound injury-related ion leakage and defense responses to abate (Figure 2).
NOTE: Taking care to keep the cuts vertical to ensure a consistent cutting surface area exposed to the elicitation solution is an important step to obtain highly reproducible results. Move the segments gently. Do not make extra cuts or wounds on the segments, which could be a source of data variation. As a principle, each test must contain at least five replicates since the variation of the ROS value is large. The more replicates set, the more reliable the data are.
- Leaf disc
- Cut the leaf discs (4 mm in diameter) from 4-6-week-old rice plants using a biopsy punch with a plunger. Always cut leaf discs from the middle third of the second leaf (numbered from the top) of the main tiller to reduce data variation (Figure 1C).
- Place one leaf disc in an individual well of a 96-well microtiter plate containing 100 µL of ddH2O for 10-12 h for pretreatment, which allows wounding-related responses to abate as these could interfere with the induction of ROS by PAMPs (Figure 2).
NOTE: Operate the leaf discs gently. Do not make extra cuts or wounds on the discs in the experiment, which could result in data variation. The induction of ROS mostly occurs from the cells of the cut edge, since the surfaces of rice tissues (leaves or sheaths), are covered with hydrophobic layers. Only the cells of the cut edges are in contact with the elicitation solution (refer to the discussion section). - Keep all the leaf disks floating, with the abaxial surface facing up, in the wells of a microtiter plate for water pretreatment to avoid leaf side-associated variation.

Figure 1: The growth condition and stages of rice seedlings for sheath sampling and parts of the rice sheath and rice leaves used in the assay. (A) Rice seedlings grown on 1/2 MS medium under sterile conditions for 10 days can be sampled for ROS assay. Sterilized rice seeds were cultured on 1/2 MS medium and grown in a 12 h light/12 h dark photoperiod in clear glass vial, 8.5 cm in diameter and 15 cm in height. (B) Schematic diagram of the sampling parts of leaf sheaths. Leaf sheaths were cut from 10-day-old rice seedlings. The positions of leaf sheaths were above the roots and below the first leaf. (C) Schematic diagram of the sampling position of leaf discs. The leaf discs can be cut from the middle third of the second leaf (count from the top) of the main tiller of healthy rice plants at any growth stage. Abbreviations: ROS = reactive oxygen species; MS = Murashige and Skoog. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Schematic diagram of the plate setup for measuring ROS production with different lines of Oryza sativa. Pretreatment and test of rice tissues using a 96-well plate. Line 1, Line 2, and Line 3 (up to eight lines on one plate) can be any material of interest, different cultivars, mutants, or transgenic lines. The tissues were stimulated with elicitation solutions with PAMP (PAMP, white) or without PAMP (ddH2O, gray) to measure ROS response. It should be noted that the more the samples to be tested, the longer the time interval between readings. Abbreviations: ROS = reactive oxygen species; PAMP = pathogen-associated molecular pattern; ddH2O = double-distilled water. Please click here to view a larger version of this figure.
{kind=link}
3. Preparing the elicitation solution
- Dissolve L-012 powder into 20 mM (6.23 mg/mL) aqueous solution with ddH2O to make the stock solution. Then, dilute the stock solution with 50 mM Tris HCl buffer (pH 7.5) to make the working solution at the final concentration of 500 µM L-012. Keep the stock solution frozen and dilute to the working solution before use.
- Prepare the elicitation solution containing PAMP, L-012, and horseradish peroxidase (HRP; 10 mg/mL in ddH2O). For a 10 mL elicitation solution, mix 9.4 mL of 50 mM Tris HCl (pH 7.5) solution, 400 µL of the L-012 solution, 100 µL of HRP, and 100 µL of flg22 (PAMP; 10 mM in ddH2O). For the negative control, add 100 µL of ddH2O instead of PAMP.
NOTE: Keep the prepared elicitation solutions at room temperature to avoid cold stress to rice tissues. Other PAMPs can also be used for treatment as required, such as chitin (20 ng/mL in final concentration). Since L-012 is light-sensitive, cover all the tubes containing L-012 solution with aluminum foil.
4. Starting the software and setting up the protocol with the referenced microplate reader (see Table of Materials)
NOTE: It takes some time to set up the parameters of the microplate reader software. It is recommended to get the machine and protocol ready (one click to proceed) before adding the elicitation solution.
- Start the software. Click the Experiments button to create a new protocol or use an existing protocol.
- Click Procedure in the pop-up to set up the plate. Select the wells from the plate to be monitored.
- Click Start Kinetic to set up the total run time and reading interval. Set the Run time to 35 min or longer, depending on the experimental requirements. To obtain readings as frequently as possible, select Minimum Interval. For integration time, choose 1 s or longer, depending on the signal intensity.
NOTE: The reading interval depends on the number of samples and signal integration duration. - Click Validate | OK to confirm the settings.
- Click on Detect the new plate in the pop-up and wait for the software to prompt the load plate dialog box. Place the plate to be tested on the carrier.
- Stop here to wait for the elicitation system to be established (in the next section). As soon as the elicitation system is ready, click on Run to start the reading.
5. Establishing the elicitation system and measuring real-time ROS production
- Carefully remove the ddH2O from the wells containing the pretreated tissues, avoiding any tissue damage or desiccation.
- Use a multichannel pipette to add 200 µL of the elicitation solution to the wells containing the tissues.
- Gently shake to mix. Click Run to begin detection.
NOTE: With PAMP treatment, plant tissues respond and produce ROS very quickly. Therefore, it is suggested that the negative control without PAMP be treated first to reduce the operation time, when there are multiple treatments. Operate as quickly as possible to reduce the elicitation delay between treatments. The shorter the time between the addition of the elicitation solution and the start of detection, the better the capture of important experimental data will be.
Results
Here, we take rice material as an example to determine the ROS produced with flg22 treatment. The generation of ROS after elicitation is transient. In rice, the increase in ROS production was first detected in 1-2 min, peaked at 10-12 min, and returned to the baseline in ~30-35 min (Figure 3). Compared to the control test, in which PAMP was absent in the elicitation solution resulting in no obvious ROS induction, a specific ROS burst was induced only when the elicitation solution containing ...
Discussion
The purpose of this study was to establish a highly efficient method to quantify early ROS production in response to PAMP in rice tissues. This method provides a standardized procedure for the real-time determination of apoplast ROS produced from treated rice tissues. This method is simple in operation, low in cost, clear in composition, and independent of commercial kits. Using this method, researchers can study the real-time production of apoplast ROS when plants are subjected to biotic or abiotic stresses.
Disclosures
The authors have no conflicts of interest to disclose.
Acknowledgements
This work was supported by grants from Shanghai Natural Science Foundation (Grant Number: 21ZR1429300/BS1500016), Shanghai Jiao Tong University (Agri-X program, Grant Number:AF1500088/002), Shanghai Collaborative Innovation Center of Agri-Seeds (Grant Number: ZXWH2150201/001) to Jiangbo Fan, and by the Medical-Engineering Collaboration Project of Shanghai Jiao Tong Univesity (grant number: 21X010301734) to Can Li.
Materials
| Name | Company | Catalog Number | Comments |
| 96-well microtiter plate | WHB | WHB-96-01 | |
| Ethanol absolute | Innochem | A43543 | |
| flg22 | Sangon Biotech | p20973 | PAMP |
| Gen5 | BioTek | software | |
| L-012 | FUJIFILM | 120-04891 | 8-amino-5-chloro-7-phenyl-2,3-dihydropyrido [3,4-d] pyridazine-1,4-dione, CAS #:143556-24-5 |
| Microplate reader | BioTek | Synergy 2 | |
| MS Medium | Solarbio | M8521 | |
| NaCLO | Aladdin | S101636 | |
| Peroxidase from horseradish (HRP) | Sigma | P8375 | |
| Phytagel | Sigma | P8169 | |
| Sampler | Miltex | 15110-40 | |
| Sucrose | Sangon Biotech | A502792 | |
| Tris | Sangon Biotech | A610195 |
References
- Gechev, T. S., Van Breusegem, F., Stone, J. M., Denev, I., Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays. 28 (11), 1091-1101 (2006).
- Mittler, R. ROS are good. Trends in Plant Science. 22 (1), 11-19 (2017).
- Gilroy, S., et al. ROS, calcium, and electric signals: key mediators of rapid systemic signaling in plants. Plant Physiology. 171 (3), 1606-1615 (2016).
- Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. Reactive oxygen gene network of plants. Trends in Plant Science. 9 (10), 490-498 (2004).
- Marino, D., Dunand, C., Puppo, A., Pauly, N. A burst of plant NADPH oxidases. Trends in Plant Science. 17 (1), 9-15 (2012).
- Mittler, R., Zandalinas, S. I., Fichman, Y., Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology. 23 (10), 663-679 (2022).
- Suzuki, N., Koussevitzky, S., Mittler, R., Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant, Cell & Environment. 35 (2), 259-270 (2012).
- Suzuki, N., et al. Respiratory burst oxidases: the engines of ROS signaling. Current Opinion in Plant Biology. 14 (6), 691-699 (2011).
- Kadota, Y., Shirasu, K., Zipfel, C. Regulation of the NADPH oxidase RBOHD during plant immunity. Plant and Cell Physiology. 56 (8), 1472-1480 (2015).
- Segonzac, C., Zipfel, C. Activation of plant pattern-recognition receptors by bacteria. Current Opinion in Microbiology. 14 (1), 54-61 (2011).
- Roda, A., et al. Progress in chemical luminescence-based biosensors: A critical review. Biosensors and Bioelectronics. 76, 164-179 (2016).
- Hong, D., Joung, H. -. A., Lee, D. Y., Kim, S., Kim, M. -. G. Attomolar detection of cytokines using a chemiluminescence immunoassay based on an antibody-arrayed CMOS image sensor. Sensors and Actuators B: Chemical. 221, 1248-1255 (2015).
- Nishinaka, Y., et al. et al. new sensitive chemiluminescence probe, L-012, for measuring the production of superoxide anion by cells. Biochemical and Biophysical Research Communications. 193 (2), 554-559 (1993).
- Grundy, J., Stoker, C., Carre, I. A. Circadian regulation of abiotic stress tolerance in plants. Frontiers in Plant Science. 6, 648 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved