תרבית תלת-ממדית של אורגנואידים מקריפטים של מעי מורין ותא גזע יחיד למחקר אורגנואידים
In This Article
Summary
אנו מתארים פרוטוקול לבידוד קריפטים של מעי דק מורין ואורגנואידים תלת-ממדיים של תרבית מעיים מהקריפטים. בנוסף, אנו מתארים שיטה ליצירת אורגנואידים מתא גזע יחיד במעי בהיעדר נישה תאית תת-אפיתלית.
Abstract
כיום, תרבית אורגנואידים מהווה כלי חשוב לחקר חוץ גופי של היבטים ביולוגיים שונים ומחלות באיברים שונים. קריפטים של המעי הדק יכולים ליצור אורגנואידים המחקים את אפיתל המעי כאשר הם מתורבתים במטריצה חוץ-תאית תלת-ממדית. האורגנואידים מורכבים מכל סוגי התאים הממלאים פונקציות הומיאוסטטיות שונות במעי. אלה כוללים תאי Paneth, תאים enteroendocrine, enterocytes, תאי גביע, ותאי tuft. מולקולות מאופיינות היטב מתווספות למדיום התרבית כדי להעשיר את תאי הגזע של המעי (ISC) המסומנות בחזרות עשירות בלאוצין המכילות קולטן מצומד לחלבון G 5 ומשמשות להנעת התמיינות לאורך שושלות ספציפיות; מולקולות אלה כוללות גורם גדילה אפידרמלי, Noggin (חלבון מורפוגנטי עצם), ו- R-spondin 1. בנוסף, מפורט פרוטוקול ליצירת אורגנואידים מקולטן הפטוצלולרי B2 (EphB2)-חיובי יחיד המייצר אריתרופואיטין. במאמר שיטות זה מתוארות טכניקות לבידוד קריפטים של המעי הדק ו- ISC יחיד מרקמות והבטחת הקמה יעילה של אורגנואידים.
Introduction
אורגנואידים במעי, שהוקמו לראשונה בשנת 2009, התגלו ככלי רב עוצמה במבחנה לחקר ביולוגיה של המעי בהתחשב בדמיון המורפולוגי והתפקודי שלהם לרקמות בוגרות. לאחרונה, התקדמות טכנולוגית באורגנואידים בתרבית שמקורם בתאי גזע של רקמה בוגרת אפשרה תרבית ארוכת טווח של תאי גזע במעי (ISC) עם פוטנציאל התחדשות עצמית והתמיינות. אורגנואידים אלה נמצאים בשימוש נרחב במחקרים בסיסיים ותרגומיים על פיזיולוגיה של מערכת העיכול ופתופיזיולוגיה 1,2,3,4,5,6. אורגנואידים תלת ממדיים שפותחו על ידי קבוצת Clevers מספקים כלי רב עוצמה לחקר אפיתל המעי עם רלוונטיות פיזיולוגית משופרת7. מכיוון שאורגנואידי מעיים נגזרים מתאי גזע רקמתיים ומורכבים מסוגי תאים מרובים, הם משחזרים את הפונקציונליות של אפיתל המעי. יש לציין כי תא גזע 5-חיובי (Lgr5+) קולטן 5-חיובי (Lgr5+) הממוין יחיד עשיר בלאוצין ומכיל קולטן מצומד לחלבון G (Lgr5+) יכול גם לייצר אורגנואידים תלת-ממדיים ללא תאי Paneth או נישה ISC כגון נישה אפיתל או נישה סטרומלית7. עם זאת, יכולת יצירת האורגנואידים של תאי Lgr5+ ממוינים בודדים נמוכה בהשוואה לאלה של קריפטה ותא ISC-Paneth כפול8.
מספר גדל והולך של מחקרים הראו כי שיטות הדגירה של חומצה אתילאנדיאמין טטראצטית (EDTA) או דיסוציאציה של collagenase גורמות להתרופפות באפיתל ולשחרור קריפטות. מכיוון שלדיסוציאציה אנזימטית עשויה להיות השפעה על מצב התא של קריפטים, שיטת בידוד מכנית משמשת בדרך כלל לניתוק הרקמה. למרות שעיכול מכני הוא טכניקה מהירה, שיטה זו יכולה להיות קשורה לתפוקות קריפטה לא עקביות או לכדאיות תאים ירודה9. לכן, טיפול EDTA ודיסוציאציה מכנית יכולים להיות משולבים כדי לייצר תפוקות קריפטה טובות יותר. מאפיין של המתודולוגיה המוצגת במאמר זה הוא השימוש בטלטול נמרץ של שברי הרקמה לאחר כלציה EDTA10. טלטול נמרץ מאפשר בידוד יעיל של קריפטים ממתחמי קריפט-וילוס במעי הדק. מידת הרעידה הידנית קובעת את ההפרדה. לכן, קבלת crypts מקומפלקסים חשוב עבור נסיינים בתחום זה. בנוסף, מיומנות נכונה יכולה להפחית את זיהום villus למינימום ולהגדיל את מספר crypts.
לפיכך, פרוטוקול ניסיוני זה, המשתמש באורגנואידים של המעי הדק שמקורם במורין, יכול לבודד טוב יותר קריפטות עם כוח פיזי לאחר טיפול ב- EDTA לדיסוציאציה. ידוע כי דפוס הביטוי של קולטן הפטוצלול B2 (EphB2) המייצר אריתרופואיטין משקף בחלקו את סביבת הקריפטה. לדוגמה, תאים חיוביים ל- EphB2 מאורגנים מלמטה ל-11 העליונים. מיון תאים המופעלים על ידי פלואורסצנטיות (FACS) בוצע על בסיס ביטוי EphB2, והתאים שהתקבלו חולקו לארבע קבוצות: EphB2גבוה, EphB2med, EphB2נמוך, ו EphB2neg. לאחר מכן, הודגמה צמיחה אורגנואידית מתאיםגבוהים מסוג EphB2 חד-ממוין בעכברי בר (WT).
Protocol
כל הניסויים בעכברים אושרו על ידי ועדת האתיקה של Suntory Animal (APRV000561), וכל בעלי החיים הוחזקו בהתאם להנחיות הוועדה לטיפול ושימוש בחיות מעבדה. נעשה שימוש בזן WT סטנדרטי של Mus musculus (C57BL6/J). נעשה שימוש בעכברים זכרים ונקבות מגיל 10 שבועות עד 20 שבועות. העכברים הומתו בחנקCO2 .
1. בידוד המעי הדק
- הבלו את המעי הדק, כולל התריסריון ואת החצי הפרוקסימלי של jejunum, עם מספריים מעבדה.
- מעבירים את הרקמה לצלחת פטרי, ושוטפים את המעי הדק עם 5 מ"ל של PBS-ABx קר (PBS + פניצילין-סטרפטומיצין [1%] + גנטמיצין [0.5%]) במזרק של 5 מ"ל כדי לנקות את התוכן הלומינלי.
- יש לפתוח את הרקמה לאורכה עם מספריים למעבדה, ולשטוף ידנית עם PBS-ABx קר תוך כדי רעידה.
הערה: על ידי גירוד הווילי עם מגלשה, זיהום וילוס יכול להיות מופחת12. - לאסוף כ 5 מ"מ x 5 מ"מ חתיכות של קטע המעי באמצעות מספריים במעבדה. מעבירים את השברים לצינור 50 מ"ל עם פינצטה, ומוסיפים 25 מ"ל של PBS-ABx קר.
- שטפו את השברים על ידי התסיסה קדימה ואחורה 10x עם 25 מ"ל של PBS-ABx קר כדי להסיר את תוכן המעי בצינור 50 מ"ל.
2. בידוד קריפטה
- יש לדגור על החתיכות ב-PBS-ABx המכילות 2 mM EDTA למשך 30 דקות על קרח ללא רעידות.
- למיצוק קל של המטריצה החוץ תאית (ECM), דגרו מראש צלחת בת 24 בארות באינקובטור תרבית רקמה של 37 מעלות צלזיוס.
- שאפו את תמיסת ה-EDTA ממערכת תרבית התאים בעזרת משאבת ואקום, הוסיפו 25 מ"ל של PBS-ABx טרי וקר, ולאחר מכן נערו את החלקים מעלה ומטה במרץ ביד 30x-40x כדי לשחרר את מתחמי הקריפט-וילוס.
הערה: ניתן לבדוק את הקריפטים והווילי המופרדים על ידי תצפית מיקרוסקופית של טיפה של 25 μL מהמתלה בהגדלה פי 4. - לאחר מכן, סנן את המתלה דרך מסננת 70 מיקרומטר פעם אחת.
- צנטריפוגה את המתלה ב 390 × גרם במשך 3 דקות ב 4 ° C.
- השהה מחדש את גלולת הקריפטה ב 20 מ"ל של סורביטול DMEM (DMEM מתקדם / F12 + פניצילין-סטרפטומיצין [1%] + גנטמיצין [0.5%] + סרום בקר עוברי [1%] + סורביטול [2%]) עם פיפטינג, והעבר את מתלה הקריפטה לשני צינורות חדשים של 15 מ"ל לחלוקה לשני פתרונות 10 מ"ל לצנטריפוגה במהירות נמוכה.
הערה: ניתן להפריד את מסת התאים הגדולה ואת התאים/פסולת באמצעות צנטריפוגה במהירות נמוכה. מסת התא הגדולה נמצאת בכדורית, ותאים/פסולת נמצאים בסופרנטנט. - צנטריפוגו את שני מתלי הקריפטה ב-80 × גרם למשך 3 דקות ב-4°C, ואז שאפו את הסופרנאטנט בעדינות.
הערה: מכיוון שתצורת הכדורים חלשה, אל תשאפו יותר מדי. השאירו 2 מ"ל של supernatant בכל צינור. - הוסף 10 מ"ל של סורביטול DMEM לכל צינור שוב. צנטריפוגה את המתלה ב 80 × גרם במשך 3 דקות ב 4 ° C.
- לאחר שאיפת supernatant, משאיר 2 מ"ל של supernatant בכל צינור, להוסיף 10 מ"ל של DMEM סורביטול עבור השעיה, וצנטריפוגה את מתלה הקריפטה ב 80 × גרם במשך 3 דקות האחרונות ב 4 ° C.
- לאחר שאיפת הסופרנטנט, השארת 2 מ"ל של סופרנאטנט בכל צינור, הוסף 10 מ"ל DMEM מלא (DMEM מתקדם / F12 + פניצילין-סטרפטומיצין [1%] + גנטמיצין [0.5%] + נסיוב בקר עוברי [1%]) להשעיית הגלולה על ידי פיפטינג למעלה ולמטה, והשאר אותו למשך דקה אחת.
הערה: המתן דקה אחת כדי להשיג את הקריפטים הצפים ביעילות. - לאחר דקה אחת, אספו כל תרחיף של 10 מ"ל בסכום כולל של 20 מ"ל, וסננו פעם אחת עם מסננת תאים של 70 מיקרומטר כדי לטהר את הקריפטות.
- לפני זריעת קריפטים טהורים במהותם, ספור את מספר הקריפטות ב- DMEM המלא המסונן, ולאחר מכן צנטריפוגה ב 290 × גרם במשך 3 דקות ב 4 ° C.
- טפטפו טיפות של 25 מיקרוליטר לתוך צלחת בקוטר 6 ס"מ בשלוש נקודות. ספרו את מספר הקריפטות תחת מיקרוסקופ בהגדלה של פי 4, וחשבו את ריכוז הקריפטות לטיפה של 25 מיקרוליטר.
- להשעות 100 crypts עם 40 μL של ECM לכל באר. Pipet למעלה ולמטה 5x-10x כדי לקבל השעיה הומוגנית של crypts ב ECM, ולאחר מכן זרע בצלחת 37 ° C שחומם מראש 24 בארות.
הערה: תמיד לשמור את ECM על קרח כדי למנוע פילמור. פיפטה בזהירות כדי למנוע יצירת בועות אוויר ב- ECM. - לדגור את צלחת 24-באר במשך 15 דקות ב 37 ° C, 5% CO2 אינקובטור פילמור של ECM.
- לבסוף, לכסות את ECM עם 500 μL של מדיום תרבית המכיל גורם גדילה אפידרמיס עכבר (EGF), עכבר רקומביננטי R-spondin 1, ועכבר רקומביננטי Noggin בטמפרטורת החדר. הריכוז הסופי של חומרים לבאר הוא כדלקמן: פניצילין-סטרפטומיצין (1%), 50 U/mL כל אחד; גנטמיצין (0.5%), 25 מיקרוגרם/מ"ל; EGF, 20 ננוגרם/מ"ל; Noggin, 100 ng/mL; R-ספונדין 1, 500 ננוגרם/מ"ל; L-גלוטמין, 2 mM.
- התחל את תרבית הקריפטה ב 37 ° C באינקובטור 5% CO2 .
הערה: עבור מדיום התרבית עבור אורגנואידים בצלחת של 24 בארות, ראה טבלה 1. - בצע הדמיה חיה לטווח ארוך כדי לצפות במורפוגנזה אורגנואידית עם מיקרוסקופ תמונה בהילוך מהיר המצויד במטרה של 20x כל 3 שעות למשך עד 7 ימים. קבל תמונות טוריות עם ערימת z בשלבי z של 1 מיקרומטר (1 מיקרומטר x חמישה צעדים).
- החליפו את המדיום אחת ליומיים.
3. מיון תאים המופעלים על ידי פלואורסצנטיות (FACS)
- בודדו קריפטים מהעכברים (ראו סעיף 2).
- טפל crypts מבודדים עם 2 מ"ל של טריפסין במשך 30 דקות ב 37 ° C.
- עצור את התגובה עם 10 מ"ל של PBS, ולאחר מכן לעבור דרך מסננת תאים 20 מיקרומטר.
- צנטריפוגה את התמיסה ב 390 × גרם במשך 3 דקות ב 4 ° C, ו resuspend עם 100 μL של DMEM מלא.
- מוסיפים נוגדן מצומד APC נגד EphB2 (1/50), ודגרים במשך 30 דקות על קרח.
- לשטוף את התאים 3x עם PBS, ולבסוף להוסיף 7-amino-actinomycin D (7-AAD) (1/100).
- מיין את התאים המוכתמים באמצעות FACS.
- התאם את גורם שינוי קנה המידה של השטח ומיין לפי גודל התא (פיזור קדימה, FSC-A) לעומת גרעיניות (פיזור צד, SSC-A).
- מיין 7-AAD תאים שליליים וחיוביים לכדאיות עם לייזר מוגדר באורך גל של 488 ננומטר והספק של 50 mV.
- סמן את השערים כדי למיין את התאים EphB2-high (EphB2high), EphB2-medium (EphB2med), EphB2-low (EphB2low) ו-EphB2-negative (EphB2neg) כאשר הלייזר מוגדר באורך גל של 640 ננומטר ובהספק של 100 mV.
- התחל את תרבית התאיםהגבוהה EphB2 ב 37 ° C באינקובטור 5% CO2 .
4. אורגנואידים בתרבית חד-תאית
- בצע את שיטת בידוד התאים על פי רמות פני השטח של EphB2 מדורגות11, ולאחר מכן קבל ארבע אוכלוסיות נפרדות (גבוהה, בינונית, נמוכה ושלילית).
- אסוף, גלולה עם צנטריפוגה ב 390 × גרם במשך 3 דקות ב 4 ° C, ולהטמיע את התאיםהגבוהים EphB2 ממוין יחיד ECM על ידי pipetting, ולאחר מכן זריעה על צלחת 24 באר (100 סינגלים / 40 μL של ECM / באר).
- כמו בשלב 2.14, אפשרו ל-ECM להתפלמר, וכסו את ה-ECM במדיום תרבית המכיל מעכב קינאז הקשור ל-Rho (ROCK) (10 מיקרומטר) במשך היומיים הראשונים כדי לשמור על התאיםהגבוהים של EphB2.
הערה: מעכב ROCK יעיל נגד אנואיקיס. - בדוק ידנית את התאים באמצעות מיקרוסקופ הפוך בהגדלה של פי 40, והתבונן באורגנואידים ברי קיימא עם היווצרות ספרואידים ובליטה של קריפטה.
Representative Results
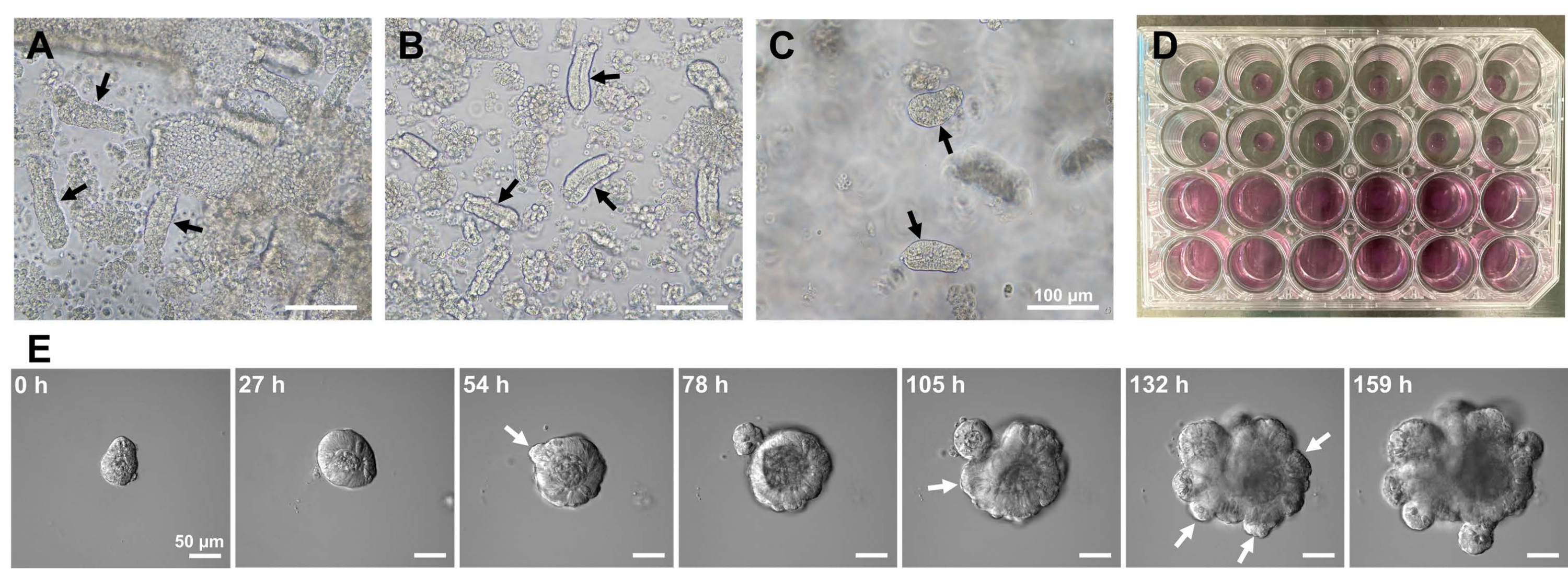
כדי ליצור אורגנואידים של עכבר במעי הדק, ניתן להשתמש בשילוב של טיפול EDTA ושיטת בידוד מכנית כדי לבודד ביעילות קריפטים10,13. תוצאות המחקר הזה הראו שכמעט כל הקריפטות המבודדות נאטמו מיד ונראו בצורת חרוט לאחר שנסחטו החוצה מגומחות האפיתל (איור 1A). כדי למזער את זיהום הווילוס, המתלה שנוצר הועבר דרך מסננת תאים של 70 מיקרומטר, ואז התסנין היה צנטריפוגה. מכיוון שחלק מהקריפטים משובשים במהלך סינון והשעיה, יש לבצע שלבים אלה בזהירות. התוצאות הראו שכמעט כל הקריפטות בשבר הסופי היו משולבות ומתאימות לשימוש בתרבית (איור 1B). כדי להמחיש את כל הקריפטים המצופים בנפרד, 100 קריפטים לכל באר היו מצופים (איור 1C). לאחר הוספת מדיום תרבית הקריפטה הספציפי (איור 1D), התפתחות האורגנואידים נוטרה במיקרוסקופ מדי יום. יתר על כן, צמיחת אורגנואידים מהקריפטים נצפתה על-ידי תמונות בהילוך מהיר כדי לעקוב אחר התפתחותן (איור 1E ווידאו משלים S1). הקריפטים המתורבתים התנהגו בצורה סטריאוטיפית. הלומן הפנימי של האורגנואיד היה מלא במסה של תאים אפופטוטיים. התפשטות פעילה והתמיינות של ISCs התרחשו באזור הקריפטה עם ניצנים (איור 1E ווידאו משלים S1). הניצנים היו יחד עם נדידה והתפשטות ISC והתמיינות תאי Paneth. תאי הפנתוס המתמיינים היו ממוקמים תמיד באתר הניצנים (איור משלים S1). מכיוון שהאורגנואידים אושרו כיציבים בתרבית באמצעות מיקרוסקופ הפוך בהגדלה של פי 10, ניתן היה להשתמש בטכניקה כדי לבחון היווצרות קריפטה במעי הדק המתפתח ולקבוע את היכולת להתחדשות רקמות והישרדות לטווח ארוך ISC לייצור תאי אפיתל מעי חדשים14,15,16.
Lgr5 מוגדר כסמן ISC, ותאי +Lgr5 יוצרים אורגנואידים תלת-ממדיים7. עם זאת, מכיוון ששפע פני השטח של חלבון LGR5 על פני התא נמוך ויש מחסור בנוגדנים נגד LGR5 בעלי זיקה גבוהה, מאתגר לבודד ביעילות ISCs מורין על ידי FACS. EphB2 זוהה בעבר כסמן פני השטח לטיהור ISCs מורין ואנושי מרקמות מעיים17,18. תבנית הביטוי של EphB2 מגדילה את המורכבות הכרוכה בסמני ISC. תאים חיוביים ל-EphB2 מאורגנים לאורך תא השגשוג, ומגיעים לשיאם בתחתית הקריפטים, בעוד שהם יורדים בשיפוע לכיוון החלק העליון של הקריפטים11. תאי Paneth ותאי אב ממוקמים גם הם בקריפטה. תאי Paneth מבטאים בעיקר EphB3, הנדרש למיקומם ותאי האב שמעליהם בקריפטה מבטאים בעיקר EphB2. לפיכך, זיהום של שני סוגי התאים יכול להתרחש במהלך טיהור ISC באמצעות נוגדן אנטי EphB2. בהתאם לכך, יש להעריך את ביטוי גני הסמן שלהם ואת יכולת יצירת האורגנואידים של תאים שבודדו באמצעות EphB2 על ידי FACS.
בהתבסס על עובדות אלה, באמצעות ניתוח FACS, ניתן לבודד תאים המסומנים על פני השטח EphB2 מקריפטות WT19. נחקר אם ביטוי EphB2 יכול להבחין בין ארבע קבוצות עם ביטוי של סמנים ספציפיים, כגון גנים של סמן ספציפי ל- ISC (Lgr5, Ascl2 ו- Olfm4) וגנים של סמן ספציפי לתאי אב (Ki67, Myc ו- FoxM1). ניסוי זה הראה כי תאיםגבוהים EphB2 היו בעיקר ISCs, בניגוד EphB2med cells20,21. לבסוף, בהתבסס על שיטת בידוד התא, התאים שהתקבלו חולקו לארבע קבוצות (EphB2גבוה, EphB2med, EphB2נמוך ותאיEphB2 neg) (איור 2). לאחר מכן, תאים בודדים המבטאים רמות גבוהות של EphB2 שמוינו על ידי FACS גודלו בתרבית לגידול אורגנואידים. תאגבוה יחיד מסוג EphB2 יכול להיות מיושם באופן עצמאי לטיפול מקומי וליצור מחדש מבנים קריפט-וויליים המתארגנים מעצמם ומזכירים את המעי הדק הרגיל (איור 3). עם זאת, התאים שמקורם בקבוצות אחרות (EphB2med, EphB2low ו-EphB2neg) אינם מייצרים אורגנואידים20.
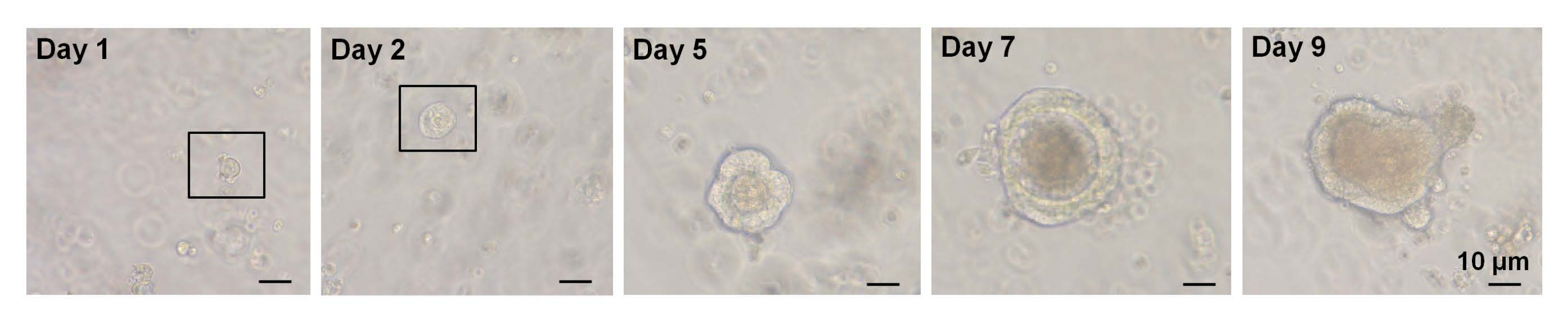
במחקר קודם, ~6% מתאי Lgr5-GFPhi חד-ממוינים הצליחו ליזום אורגנואידים7. עם זאת, התאים הנותרים לא היו מסוגלים לייצר אורגנואידים ומתו בתוך 12 שעות7 הראשונות. המחברים הניחו כי זו תוצאה של לחץ פיזי ו/או ביולוגי הטבוע בהליך הבידוד7. גידול אורגנואידים של פחות מ-6% התקבל גם מתאיםגבוהים מסוג EphB2 במיון יחיד בעכברי WT. ביום 5 של התרבית נוצרו מבנים דמויי ספרואידים (איור 3). מהיום ה-7 עד היום ה-9 התרחש אידוי של הכתמים ליצירת קריפטות (איור 3). חשוב לציין, היישום של מעכב ROCK נבחר על תאיםגבוהים EphB2 ממוינים יחיד הפחית אפופטוזיס המושרה על ידי דיסוציאציה והגדיל את היעילות של גידול אורגנואידים.

איור 1: יצירת אורגנואידים של עכברים במעי הדק. (A) קריפטים שהוכנו על-ידי שילוב של כלציה EDTA ודיסוציאציה מכנית. (B) כתוצאה מכך קריפטות מטוהרות. (C) קריפטים המוטבעים במטריצה החוץ תאית. (א-ג) החיצים השחורים מציינים קריפטות. (D) תרבית תלת-ממדית של קריפטות ואורגנואידים. (E) תמונות מייצגות של אורגנואיד גדל שמקורו בקריפטה. החיצים הלבנים מצביעים על ניצני קריפטה. פסי קנה מידה = (A-C) 100 מיקרומטר ו- (E) 50 מיקרומטר. לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

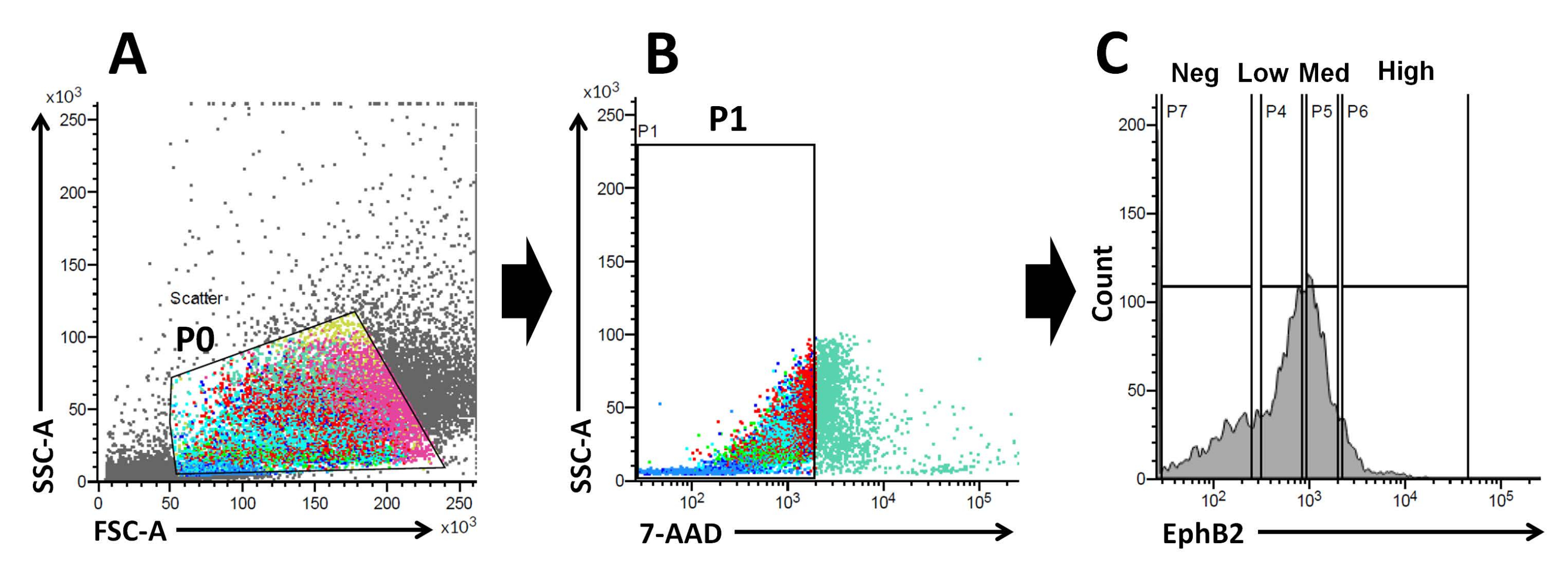
איור 2: אסטרטגיית גאטומטריית זרימה להשגת אוכלוסייה של תאים חיוביים ל-EphB2 (EphB2+) בעכברי בר. (A) חלקות פיזור קדימה וצד משמשות להפרדת התאים בהתאם לגודלם ולגרעיניותם, בהתאמה. (B) פיזור פלואורסצנטי משמש להפרדת תאים בני קיימא בהתאם לעוצמת הפלואורסצנטיות 7-AAD (PerCP) של התאים. נבחר השער לאוכלוסיית התאים השליליים 7-AAD. (C) נבחרו השערים לאוכלוסיות התאים EphB2-high (EphB2high), EphB2-medium (EphB2med), EphB2-low (EphB2low) ו-EphB2-negative (EphB2neg). קיצורים: FSC-A = אזור פיזור-שיא קדמי; SSC-A = אזור פיזור צדדי; 7-AAD = 7-amino-actinomycin D. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 3: מהלך הזמן של גידול אורגנואיד בתאיםגבוהים מסוג EphB2 במיון יחיד בעכברי בר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
טבלה 1: מדיום תרבית לצלחת בת 24 בארות. אנא לחץ כאן כדי להוריד טבלה זו.
וידאו משלים S1: תמונות קיטועי זמן של אורגנואיד גדל. סרגל קנה מידה = 50 מיקרומטר. אנא לחץ כאן להורדת קובץ זה.
איור משלים S1: תמונה מייצגת של צביעת נוגדנים נגד ליזוזים באורגנואיד. החצים הלבנים מציינים תאי Paneth. קיצור: DIC = מיקרוסקופ ניגודיות הפרעות דיפרנציאליות. סרגל קנה מידה = 10 מיקרומטר. אנא לחץ כאן להורדת קובץ זה.
Discussion
פרוטוקול זה מתאר שיטה לבידוד עקבי של קריפטים במעי הדק ואת התרבית הבאה של אורגנואידים תלת-ממדיים. כדי לשפר את קצב שחרור הקריפטה, נקבעה שיטת בידוד מכנית הכוללת טלטול נמרץ לאחר הטיפול ב- EDTA. הרכב המדיום שונה מהפרוטוקול המקורי של Sato et al.7. המדיום המקורי יקר יחסית. לפיכך, מדיום תרבית ומדיה מותאמת אישית לאורגנואידים של המעי הדק המכילים מעכבים פרמקולוגיים, גורמי גדילה רקומביננטיים ו / או מדיה מותנית מוצגים בטבלה 1. Wnt3A ו-N-אצטיל-ציסטאין אינם כלולים במדיום התרבית בפרוטוקול זה. כאשר תאי Paneth מבטאים Wnt3, התאים מייצרים Wnt3 ותומכים בתחזוקת ISC. בנוסף, במהלך בידוד קריפטה, המדיום המותנה אינו בשימוש. מודל האורגנואידים הוא דינמי ובעל הטרוגניות תאית ומבנית (תאי פאנת, אנטרוציטים, תאי גביע, תאים אנטרואנדוקריניים, תאי ציץ ותאי ISC). לפיכך, אורגנואידים אלה יכולים לשמש בקנה מידה גדול כדי לחקור סוגיות בסיסיות של ביולוגיה אורגנואידים.
שיפוע EphB2 שומר על גזע ISC והתפשטות לאורך ציר הקריפט-וילוס במעי הדק הבוגר18. היתרון של ייצור אורגנואידים מתא EphB2 יחיד בהשוואה לקריפטות מבודדות קשור להבנת הביולוגיה של ISCs מורינים, שכן ISCs ממלאים תפקידי מפתח בהפרעות מעיים אנושיות שונות. ניתן לתרבית ISCs יחידים בעלי ביטויגבוה של EphB2 ליצירת אורגנואידים באופן דומה להתפתחות אורגנואידים מספקי ISC בודדים המבטאים Lgr5. השלב החשוב ביותר הוא לחלק במדויק את התאים לארבע קבוצות (EphB2גבוה, EphB2med, EphB2נמוך ו- EphB2neg) על פי ביטוי EphB2 בקריפטות באמצעות FACS. תרשימי פיזור קדימה לעומת צד (FSC לעומת SSC) משמשים בדרך כלל לזיהוי תאים מעניינים בהתבסס על גודלם וגרעיניותם. FSC מציין את גודל התא, ו-SSC מתייחס למורכבות או לגרעיניות של התא בשער P0 (איור 2A). בעבודה זו, התאים שנפלו בתוך השער המוגדר (P0) נותחו לאחר מכן לכדאיות. לאחר מכן, הכדאיות שלהם נקבעה על פי האוכלוסיות השליליות והחיוביות של אותות פלואורסצנטיים 7-AAD. הגבול בין 7-AAD-שלילי ו -חיובי הוחלט בהחלט כדי להשיג את אלה שליליים עם זיהום תאים חיובי מינימלי. שערי EphB2 נקבעו באופן גס בהתבסס על הביטוי המדורג EphB2.
כדי לאשר שארבע הקבוצות חולקו במדויק, נותח ביטוי ה-mRNA של גנים נבחרים. רמות ה-mRNA של סמני ISC גבוהות בתאי EphB2גבוהים 20. בנוסף, רמות ה-mRNA של סמנים ספציפיים לתאי אב גבוהות יחסית בתאירפואה מסוג EphB220. עם זאת, דיכוי EphB2 בתאי EphB2נמוך ותאי EphB2 neg נמוך או שלילי בהשוואה לזה של EphB2גבוה ותאיEphB2 med 20. יש לנקוט בצעדים הקודמים כדי להבטיח את העשרת אוכלוסיית התאיםהגבוהה EphB2 לפני הציפוי. עם זאת, גידול אורגנואידים של פחות מ -6% מתאיםגבוהים EphB2 עשוי להיות תוצאה של מוות של תאי גזע במהלך תהליך התרבית, לא רעידות נמרצות במהלך בידוד קריפטה. הוכח כי היישום של מעכב קינאז סלקטיבי הקשור ל-Rho (ROCK) על תאי גזע עובריים אנושיים מפחית במידה ניכרת אפופטוזיס המושרה על ידי דיסוציאציה22. לכן, כשינוי טכני, כדאי לנסות להוסיף את מעכב ROCK בריכוז גבוה יותר ועם דגירה ארוכה יותר כדי לשפר את הכדאיות.
תאי Paneth מפרישי Wnt3A לצד ISCs מספקים תמיכה חיונית ל-ISCs8. ואכן, מכפילי תאי ISC-Paneth מציגים יכולת יצירת אורגנואידים מוגברת מאוד בהשוואה לתאי ISC בודדים8. יתר על כן, תוספת של Wnt3A בריכוז של 100 ננוגרם/מ"ל במשך 3 הימים הראשונים של התרבית הוכחה כמגדילה את יכולת יצירת האורגנואידים8. לכן, כשינוי טכני נוסף, הוספת Wnt3A אקסוגני יכולה לשפר את יכולת יצירת האורגנואידים של ISCs יחידים בעלי ביטויגבוה של EphB2.
בהשוואה לגישות in vivo, אורגנואידים יכולים לשמש בקלות למניפולציה גנטית, ניתוח פנוטיפים ממאירים, ובדיקת תרופות20,23. שילוב של כלציית EDTA ושיטת בידוד מכנית הוא יעיל, ניתן לשחזור וחסכוני בזמן ליצירת אורגנואידים במעי דק מקריפטות וניתן לעקוב אחריו בקלות על ידי צוות המעבדה ללא כל ניסיון מתקדם. לפיכך, תוספת של בידוד מכני עם רעד נמרץ לאחר הטיפול עם EDTA יכול ביעילות לבסס מורין אורגנואידים במעי דק ex vivo ולספק כלי פוטנציאלי לטיפוח אורגנואידים ומידול מחלות של רקמות אפיתל בוגרות אחרות.
תאי אפיתל המעי מקוטבים ומכוונים כאשר הצד האפי מכוון לכיוון הלומן. עם זאת, הצד האפי מול לומן של אורגנואידים 3D הוא הפנים שלהם. לפיכך, ארגון זה מונע גישה לצד האפיקאלי, שהוא בעיה כאשר לומדים את ההשפעות של רכיבים לומינליים, כגון חומרים מזינים, חיידקים ומטבוליטים על תאי אפיתל. כדי לעקוף את החיסרון הזה, פותחה תרבית של תאים אורגנואידים כחד-שכבות דו-ממדיות24. במונחים של יישומים עתידיים, תרבית של חד-שכבות תאים אורגנואידים תנוצל, מכיוון שהיא מייצגת את המערכת היעילה והניתנת למשיכה ביותר.
Disclosures
למחברים אין ניגודי עניינים להצהיר.
Acknowledgements
עבודה זו נתמכה על ידי מענקים בסיוע למחקר מדעי (C) ל- T.T. (מספרי מענקים JP17K07495 ו- JP20K06751). אנו מודים לפרופ' מינקו קנגאקו על השימוש בציוד לדימות בהילוך ארוך טווח (LCV100; אולימפוס).
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL Eppendorf tube | Eppendorf | 0030 125.215 | |
| 5 mL syringe | TERUMO | SS-05SZ | |
| 15 mL Falcon tube | Iwaki | 2325-015 | |
| 20 μm cell strainer | Sysmex | 04-004-2325 | |
| 24-well plate | Iwaki | 3820-024 | |
| 50 mL Falcon tube | Iwaki | 2345-050 | |
| 60 mm tissue culture dish | FALCON | 353002 | |
| 70 μm cell strainer | Falcon | 352350 | |
| 100 mm Petri dish | Iwaki | 3020-100 | |
| 7-AAD | BD Biosciences | 559925 | |
| Advanced DMEM/F12 | Gibco | 12634-010 | |
| Alexa Fluor 568 Goat Anti-Mouse IgG (H+L) | Invitrogen | A-11004 | |
| Anti-EphB2 APC-conjugated antibody | BD Biosciences | 564699 | |
| C57BL6/J mice | Japan SLC, Inc. | ||
| Clean bench | HITACHI | CCV-1306E | |
| Confocal laser scanning microscope | Olympus | FV3000 | |
| EDTA (0.5 mol/L) | Nacalai Tesque | 06894-14 | 2 mM |
| FACSMelody | BD Life Sciences-Biosciences | 661762 | |
| Fetal bovine serum | Sigma | 173012 | 1% (v/v) |
| Fiji (software) | https://fiji.sc/ | ||
| Gentamicin (10 mg/mL) | Nacalai Tesque | 16672-04 | 25 μg/mL |
| Hammacher laboratory scissor | SANSYO | 91-1538 | |
| Incubator | Panasonic | MCO-170-PJ | |
| Laboratory tweezer | AS-ONE | 7-164-04 | |
| L-Glutamine 200 mM | Gibco | 25030081 | 2 mM |
| Matrigel | BD Biosciences | 354230 | ECM for 3D organoids |
| Mouse Anti-Human Lysozyme | LSBio | LS-B8704-100 | |
| Murine EGF (20 μg/mL stock solution) | PeproTech | 315-09 | 20 ng/mL |
| PBS 1x | Gibco | 10010-023 | |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140-122 | 50 U/mL |
| Pipetman (10 μL, 20 μL, 200 μL, and 1,000 μL) | GILSON | 1-6855-12, -13, -15, and -16 | |
| Recombinant murine Noggin (20 μg/mL stock solution | R&D Systems | 1967-NG-025 | 100 ng/mL |
| Recombinant murine R-Spondin 1 (250 μg/mL stock solution) | R&D Systems | 3474-RS-050 | 500 ng/mL |
| Sorbitol | Nacalai Tesque | 32021-95 | 2% (w/v) |
| TE2000-S (inverted microscope) | Nikon | 24131 | |
| Time-lapse image microscope | Olympus | LCV100 | |
| TrypLE Express 1x | Gibco | 12605-010 | |
| ULVAC | ULVAC KIKO Inc. | 100073 | |
| Y-27632 | Fujifilm | 331752-47-7 | 10 μM |
References
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Seidlitz, T., et al. Human gastric cancer modelling using organoids. Gut. 68 (2), 207-217 (2019).
- Nikolaev, M., et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature. 585 (7826), 574-578 (2020).
- Artegiani, B., Clevers, H. Use and application of 3D-organoid technology. Human Molecular Genetics. 27, R99-R107 (2018).
- Lancaster, M. A., Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 345 (6194), 1247125 (2014).
- Dedhia, P. H., Bertaux-Skeirik, N., Zavros, Y., Spence, J. R. Organoid models of human gastrointestinal development and disease. Gastroenterology. 150 (5), 1098-1112 (2016).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Sato, T., et al. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature. 469 (7330), 415-418 (2011).
- Aronowitz, J. A., Lockhart, R. A., Hakakian, C. S. Mechanical versus enzymatic isolation of stromal vascular fraction cells from adipose tissue. Springerplus. 4 (1), 713 (2015).
- Takahashi, T. New trends and perspectives in the function of non-neuronal acetylcholine in crypt-villus organoids in mice. Methods in Molecular Biology. 1576, 145-155 (2019).
- Batlle, E., et al. β-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell. 111 (2), 251-263 (2002).
- Baghdadi, M. B., Kim, T. -. H. Analysis of mouse intestinal organoid culture with conditioned media isolated from mucosal enteric glial cells. STAR Protocols. 3 (2), 101351 (2022).
- Takahashi, T., et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS Journal. 281 (20), 4672-4690 (2014).
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nature Reviews Molecular Cell Biology. 15 (1), 19-33 (2014).
- Fordham, R. P., et al. Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell. 13 (6), 734-744 (2013).
- Miyoshi, H., et al. Wnt5a potentiates TGF-β signaling to promote colonic crypt regeneration after tissue injury. Science. 338 (6103), 108-113 (2012).
- Jung, P., et al. Isolation and in vitro expansion of human colonic stem cells. NatureMedicine. 17 (10), 1225-1227 (2011).
- Merlos-Suárez, A., et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell. 8 (5), 511-524 (2011).
- Mao, W., et al. EphB2 as a therapeutic antibody drug target for the treatment of colorectal cancer. Cancer Research. 64 (3), 781-788 (2004).
- Takahashi, T., et al. Muscarinic receptor M3 contributes to intestinal stem cell maintenance via EphB/ephrin-B signaling. Life Science Alliance. 4 (9), e202000962 (2021).
- Jung, P., et al. Isolation of human colon stem cells using surface expression of PTK7. Stem Cell Reports. 5 (6), 979-987 (2015).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Schulte, L., Hohwieler, M., Müller, M., Klaus, J. Intestinal organoids as a novel complementary model to dissect inflammatory bowel disease. Stem Cells International. 2019, 8010645 (2019).
- Puzan, M., Hosic, S., Ghio, C., Koppes, A. Enteric nervous system regulation of intestinal stem cell differentiation and epithelial monolayer function. Scientific Reports. 8 (1), 6313 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved