3D-культивирование органоидов из крипт кишечника мышей и одной стволовой клетки для исследования органоидов
In This Article
Summary
Мы описываем протокол для выделения мышиных тонких кишечных крипт и культивирования кишечных 3D-органоидов из крипт. Кроме того, мы описываем способ получения органоидов из одной стволовой клетки кишечника в отсутствие субэпителиальной клеточной ниши.
Abstract
В настоящее время органоидная культура представляет собой важный инструмент для исследований in vitro различных биологических аспектов и заболеваний в различных органах. Мышиные тонкие кишечные крипты могут образовывать органоиды, которые имитируют кишечный эпителий при культивировании во внеклеточном 3D-матриксе. Органоиды состоят из всех типов клеток, которые выполняют различные гомеостатические функции кишечника. К ним относятся клетки Панета, энтероэндокринные клетки, энтероциты, бокаловидные клетки и пучковые клетки. Хорошо охарактеризованные молекулы добавляют в культуральную среду для обогащения кишечных стволовых клеток (МСК), меченных богатыми лейцином повторами, содержащими рецептор 5, связанный с G-белком, и используются для дифференцировки по определенным линиям; эти молекулы включают эпидермальный фактор роста, ноггин (костный морфогенетический белок) и R-спондин 1. Кроме того, подробно описан протокол генерации органоидов из одного гепатоцеллюлярного рецептора B2 (EphB2)-положительного ISC, продуцирующего эритропоэтин. В этой статье описаны методы выделения тонких кишечных крипт и одного ISC из тканей и обеспечения эффективного установления органоидов.
Introduction
Кишечные органоиды, которые были впервые созданы в 2009 году, стали мощным инструментом in vitro для изучения биологии кишечника, учитывая их морфологическое и функциональное сходство со зрелыми тканями. В последнее время технологические достижения в области культивируемых органоидов, полученных из стволовых клеток взрослой ткани, позволили проводить долгосрочную культуру кишечных стволовых клеток (ISC) с потенциалом самообновления и дифференцировки. Эти органоиды широко используются для фундаментальных и трансляционных исследований по физиологии желудочно-кишечного тракта и патофизиологии 1,2,3,4,5,6. 3D-органоиды, разработанные группой Clevers, представляют собой мощный инструмент для изучения кишечного эпителия с улучшенной физиологической значимостью7. Поскольку кишечные органоиды получены из тканевых стволовых клеток и состоят из нескольких типов клеток, они повторяют функциональность кишечного эпителия. Следует отметить, что стволовая клетка, содержащая 5-положительный рецептор, связанный с G-белком (Lgr5+), также может генерировать 3D-органоиды без каких-либо клеток Панета или ниши ISC, такой как эпителиальная ниша или стромальная ниша7. Однако органоидообразующая способность односортных клеток Lgr5+ низка по сравнению с таковыми у дублетов клеток крипт и ISC-Paneth8.
Все большее число исследований показало, что методы инкубации этилендиаминтетрауксусной кислоты (ЭДТА) или диссоциации коллагеназы вызывают ослабление эпителия и высвобождение крипт. Поскольку ферментативная диссоциация может влиять на состояние клеток крипт, для диссоциации ткани обычно используется метод механической изоляции. Хотя механическое пищеварение является быстрым методом, этот метод может быть связан с непостоянным выходом крипт или плохой жизнеспособностью клеток9. Таким образом, обработка ЭДТА и механическая диссоциация могут быть объединены для получения лучших выходов крипты. Особенностью методики, показанной в данной статье, является использование энергичного встряхивания тканевых фрагментов после хелатирования ЭДТА10. Энергичное встряхивание позволяет эффективно изолировать крипты от комплексов крипты-ворсинок в тонком кишечнике. Степень ручного встряхивания определяет разделение. Таким образом, получение крипт из комплексов важно для экспериментаторов в этой области. Кроме того, надлежащая сноровка может свести загрязнение ворсинок к минимуму и увеличить количество склепов.
Следовательно, этот экспериментальный протокол, в котором используются органоиды тонкой кишки мышиного происхождения, может лучше изолировать крипты с физической силой после обработки ЭДТА для диссоциации. Известно, что паттерн экспрессии эритропоэтин-продуцирующего гепатоцеллюлярного рецептора В2 (EphB2) частично отражает среду крипты. Например, EphB2-положительные клетки организованы снизу вверх11. Флуоресцентно-активируемая сортировка клеток (FACS) проводилась на основе экспрессии EphB2, и полученные клетки были разделены на четыре группы: EphB2высокий, EphB2med, EphB2низкий и EphB2neg. Затем был продемонстрирован рост органоидов из односортныхвысоких клеток EphB2 у мышей дикого типа (WT).
Protocol
Все эксперименты на мышах были одобрены Комитетом по этике животных Suntory (APRV000561), и все животные содержались в соответствии с руководящими принципами комитета по уходу и использованию лабораторных животных. Использовался стандартный штамм WT Mus musculus (C57BL6/J). Использовались как самцы, так и самки мышей в возрасте от 10 до 20 недель. Мышей усыпляли с помощью удушья CO2 .
1. Изоляция тонкой кишки
- Иссекают тонкую кишку, включая двенадцатиперстную кишку и проксимальную половину тощей кишки, лабораторными ножницами.
- Перенесите ткань в чашку Петри и промойте тонкую кишку 5 мл холодного PBS-ABx (PBS + пенициллин-стрептомицин [1%] + гентамицин [0,5%]) в шприце объемом 5 мл, чтобы очистить содержимое просвета.
- Разрежьте ткань вдоль лабораторными ножницами и вручную промойте холодным PBS-ABx во время встряхивания.
ПРИМЕЧАНИЕ: Соскабливая ворсинки с помощью предметного стекла, можно уменьшить загрязнение ворсинок12. - Соберите кусочки сегмента кишечника размером примерно 5 мм х 5 мм с помощью лабораторных ножниц. Перенесите фрагменты в пробирку объемом 50 мл с помощью пинцета и добавьте 25 мл холодного PBS-ABx.
- Промойте фрагменты, перемешивая вперед и назад 10 раз с 25 мл холодного PBS-ABx, чтобы удалить содержимое кишечника в пробирке объемом 50 мл.
2. Изоляция крипты
- Инкубируйте кусочки в PBS-ABx, содержащем 2 мМ ЭДТА, в течение 30 минут на льду без встряхивания.
- Для легкого затвердевания внеклеточного матрикса (ECM) предварительно инкубируйте 24-луночную пластину в инкубаторе для культивирования тканей с температурой 37 °C.
- Аспирируйте раствор ЭДТА из системы клеточных культур с помощью вакуумного насоса, добавьте 25 мл свежего холодного PBS-ABx, а затем энергично встряхните кусочки вверх и вниз вручную 30x-40x, чтобы высвободить комплексы крипты-ворсинок.
ПРИМЕЧАНИЕ: Отделенные крипты и ворсинки могут быть проверены путем микроскопического наблюдения за каплей 25 мкл из суспензии при 4-кратном увеличении. - Затем один раз отфильтруйте суспензию через сетчатый фильтр 70 мкм.
- Центрифугу суспензию при 390 × г в течение 3 мин при 4 °С.
- Ресуспендировать гранулу крипты в 20 мл сорбитола DMEM (усовершенствованный DMEM/F12 + пенициллин-стрептомицин [1%] + гентамицин [0,5%] + эмбриональная бычья сыворотка [1%] + сорбит [2%]) с помощью пипетки и перенести суспензию крипты в две новые пробирки по 15 мл для разделения на два раствора по 10 мл для центрифуги на низкой скорости.
ПРИМЕЧАНИЕ: Большая масса ячеек и клетки/мусор могут быть разделены с помощью низкоскоростного центрифугирования. Большая клеточная масса находится в грануле, а клетки/мусор - в надосадочной жидкости. - Центрифугируют две суспензии крипт при 80 × г в течение 3 мин при 4 °C, а затем осторожно аспирируют надосадочную жидкость.
ПРИМЕЧАНИЕ: Поскольку образование гранул слабое, не аспирируйте слишком сильно. Оставьте по 2 мл надосадочной жидкости в каждой пробирке. - Снова добавьте 10 мл сорбита DMEM в каждую пробирку. Центрифугируют суспензию при 80 × г в течение 3 мин при 4 °С.
- После аспирации надосадочной жидкости, оставляя 2 мл надосадочной жидкости в каждой пробирке, добавляют 10 мл сорбита DMEM для ресуспендирования и центрифугируют суспензию крипты при 80 × г в течение последних 3 мин при 4 ° C.
- После аспирации надосадочной жидкости, оставляя по 2 мл надосадочной жидкости в каждой пробирке, добавьте 10 мл полного DMEM (усовершенствованный DMEM/F12 + пенициллин-стрептомицин [1%] + гентамицин [0,5%] + эмбриональная бычья сыворотка [1%]) для ресуспендирования гранулы пипеткой вверх и вниз и оставьте на 1 минуту.
ПРИМЕЧАНИЕ: Подождите 1 минуту, чтобы эффективно получить плавающие крипты. - Через 1 минуту соберите каждую суспензию объемом 10 мл, чтобы получить в общей сложности 20 мл, и один раз отфильтруйте с помощью ситечка для клеток 70 мкм для очистки крипт.
- Перед посевом по существу чистых крипт подсчитывают количество крипт в отфильтрованном полном DMEM, а затем центрифугу при 290 × г в течение 3 мин при 4 °C.
- Капните 25 мкл-капель в 6-сантиметровую посуду в трех точках. Подсчитайте количество крипт под микроскопом при 4-кратном увеличении и рассчитайте концентрацию крипт на каплю 25 мкл.
- Суспендировать 100 крипт с 40 мкл ECM на лунку. Пипетка вверх и вниз 5x-10x для получения однородной суспензии крипт в ECM, а затем посев в предварительно разогретую 24-луночную пластину с температурой 37 °C.
ПРИМЕЧАНИЕ: Всегда держите ECM на льду, чтобы избежать полимеризации. Используйте пипетку осторожно, чтобы избежать образования пузырьков воздуха в ECM. - Инкубируйте 24-луночную пластину в течение 15 минут в инкубаторе с температурой 37 °C, 5% CO2 для полимеризации ECM.
- Наконец, накройте ECM 500 мкл питательной среды, содержащей эпидермальный фактор роста мыши (EGF), рекомбинантный мышиный R-спондин 1 и рекомбинантный мышиный ноггин при комнатной температуре. Конечная концентрация материалов на лунку следующая: пенициллин-стрептомицин (1%), 50 ЕД/мл каждый; гентамицин (0,5%), 25 мкг/мл; EGF, 20 нг/мл; Ноггин, 100 нг/мл; R-спондин 1, 500 нг/мл; L-глютамин, 2 мМ.
- Запускайте культивирование крипты при 37 °C в инкубаторе с 5% CO2 .
ПРИМЕЧАНИЕ: Питательную среду для органоидов в 24-луночном планшете см. в таблице 1. - Выполняйте долгосрочную визуализацию в реальном времени для наблюдения за морфогенезом органоидов с помощью регистрирующего микроскопа с покадровой съемкой, оснащенного 20-кратным объективом, каждые 3 часа в течение 7 дней. Получение последовательных z-стекированных изображений с z-шагом 1 мкм (1 мкм x пять шагов).
- Меняйте носитель через день.
3. Сортировка клеток, активируемая флуоресценцией (FACS)
- Изолируйте крипты от мышей (см. раздел 2).
- Изолированные крипты обрабатывают 2 мл трипсина в течение 30 мин при 37 °C.
- Остановите реакцию с помощью 10 мл PBS, а затем пропустите через сетчатое фильтр ячейки 20 мкм.
- Центрифугируют раствор при 390 × г в течение 3 мин при 4 °C и ресуспендируют 100 мкл полного ДМЭМ.
- Добавьте антитело против EphB2 APC-конъюгированного (1/50) и инкубируйте в течение 30 минут на льду.
- Промойте клетки 3 раза PBS и, наконец, добавьте 7-аминоактиномицин D (7-AAD) (1/100).
- Отсортируйте окрашенные ячейки с помощью FACS.
- Отрегулируйте коэффициент масштабирования области и отсортируйте его по размеру ячейки (прямой разброс, FSC-A) и степени детализации (боковой разброс, SSC-A).
- Отсортируйте отрицательные и положительные клетки 7-AAD на жизнеспособность с помощью лазерной установки на длине волны 488 нм и мощности 50 мВ.
- Разграничьте ворота для сортировки клеток EphB2 с высоким (EphB2высокий), EphB2-средний (EphB2med), EphB2-низкий (EphB2низкий) и EphB2-отрицательный (EphB2neg) клетки с помощью лазера, установленного на длине волны 640 нм и мощности 100 мВ.
- Начните культивироватьEphB2 с высоким содержанием клеток при 37 ° C в инкубаторе с 5% CO2 .
4. Одноклеточные культивируемые органоиды
- Проведите метод выделения клеток в соответствии с градуированными поверхностными уровнямиEphB2 11, а затем получите четыре различные популяции (высокие, средние, низкие и отрицательные).
- Соберите, гранулируйте центрифугированием при 390 × г в течение 3 мин при 4 °C и поместите односортныевысокие ячейки EphB2 в ECM путем пипетирования с последующим посевом на 24-луночную пластину (100 синглетов/40 мкл ECM/лунка).
- Как и на этапе 2.14, дайте ECM полимеризоваться и покройте ECM питательной средой, содержащей ингибитор Rho-ассоциированной киназы (ROCK) (10 мкМ) в течение первых 2 дней для поддержаниявысокого уровня клеток EphB2.
ПРИМЕЧАНИЕ: Ингибитор ROCK эффективен против анойкиса. - Вручную осмотрите клетки с помощью инвертированного микроскопа с 40-кратным увеличением и наблюдайте жизнеспособные органоиды с образованием сфероидов и выступом крипты.
Representative Results
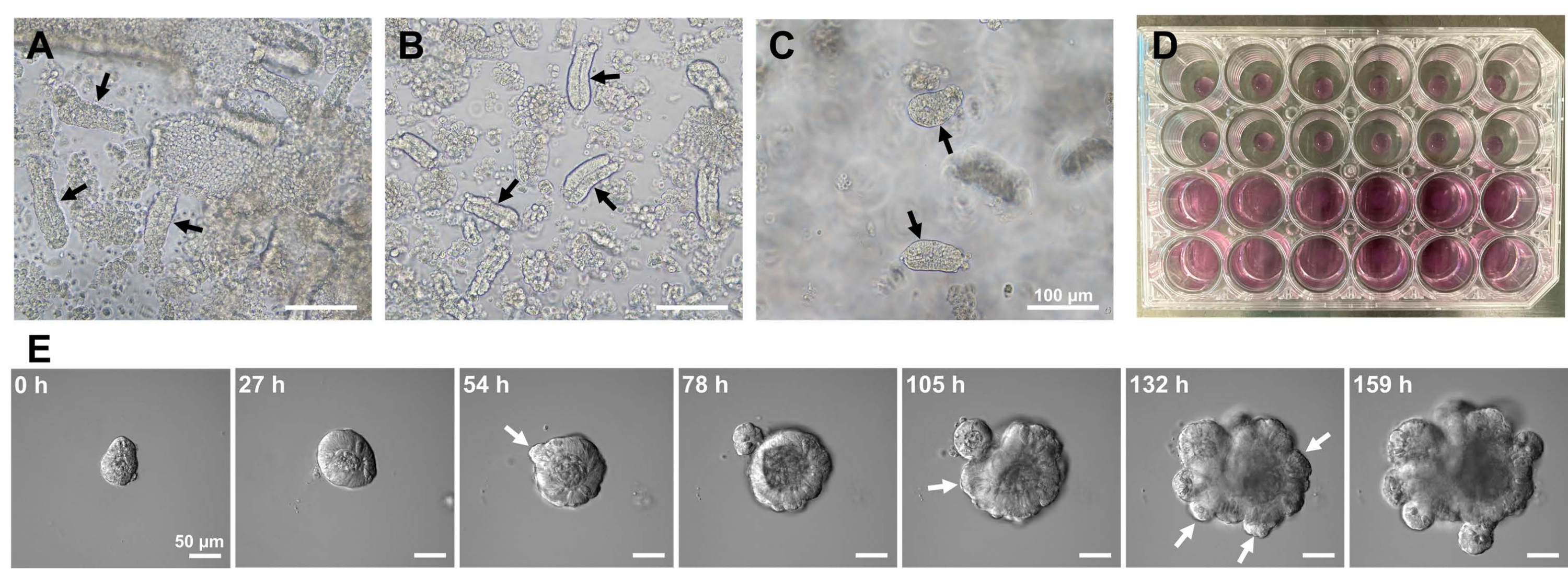
Для получения органоидов тонкого кишечника мыши можно использовать комбинацию лечения ЭДТА и метода механической изоляции для эффективного выделения крипт10,13. Результаты этого исследования показали, что почти все изолированные крипты были немедленно запечатаны и выглядели конусообразными после того, как их выдавили из эпителиальных ниш (рис. 1А). Чтобы свести к минимуму загрязнение ворсинок, полученную суспензию пропускали через сетчатый фильтр 70 мкм, а затем фильтрат центрифугировали. Поскольку некоторые крипты разрушаются во время фильтрации и суспензии, эти шаги следует выполнять осторожно. Результаты показали, что почти все крипты в конечной фракции были интегрированы и пригодны для использования в культуре (рис. 1B). Чтобы визуализировать все гальванические склепы по отдельности, было нанесено 100 склепов на скважину (рис. 1C). После добавления специфической среды для культивирования крипт (рис. 1D) развитие органоидов ежедневно контролировалось с помощью микроскопа. Кроме того, рост органоидов из крипт наблюдался с помощью покадровых изображений для наблюдения за их развитием (рис. 1E и дополнительное видео S1). Культивируемые склепы вели себя стереотипно. Внутренний просвет органоида был заполнен массой апоптотических клеток. Активная пролиферация и дифференциация ISC происходили в области крипты с почкованием (рис. 1E и дополнительное видео S1). Почкование сочеталось с миграцией и пролиферацией ISC и дифференцировкой клеток Панета. Дифференцированные клетки Панета всегда располагались в месте почкования (дополнительный рисунок S1). Поскольку было подтверждено, что органоиды стабильны в культуре с использованием инвертированного микроскопа при 10-кратном увеличении, этот метод может быть использован для изучения образования крипт в развивающейся тонкой кишке и определения способности к регенерации тканей и долгосрочной выживаемости ISC для производства новых эпителиальных клеток кишечника14,15,16.
Lgr5 определяется как маркер ISC, а мышиные клетки Lgr5+ образуют 3D-органоиды7. Однако, поскольку содержание белка LGR5 на клеточной поверхности низкое, а высокоаффинных антител против LGR5 не хватает, сложно эффективно изолировать мышиные ISC с помощью FACS. EphB2 ранее был идентифицирован как поверхностный маркер для очистки мышиных и человеческих ISC из тканей кишечника17,18. Паттерн экспрессии EphB2 увеличивает сложность, связанную с маркерами ISC. EphB2-положительные клетки организованы по всему пролиферативному компартменту, достигая пика в нижней части крипт, в то время как они уменьшаются в градиенте к вершине крипт11. Клетки Панета и клетки-предшественники также локализуются в крипте. Клетки Панета в основном экспрессируют EphB3, который необходим для их позиционирования, а клетки-предшественники над ними в крипте экспрессируют в основном EphB2. Таким образом, контаминация обоих типов клеток может произойти в ходе очистки ISC с использованием антитела против EphB2. Соответственно, следует оценивать экспрессию их маркерных генов и органоидообразующую способность клеток, выделенных с помощью EphB2 методом FACS.
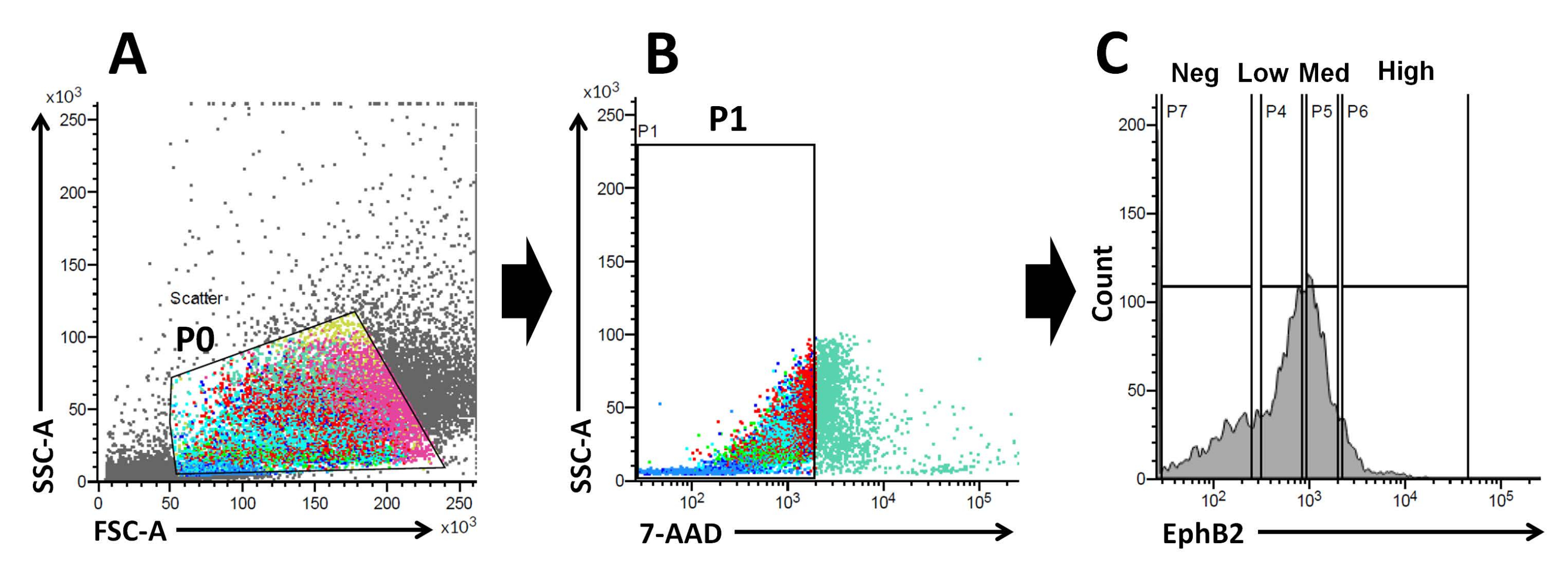
Основываясь на этих фактах, используя анализ FACS, клетки EphB2, меченные поверхностью, могут быть выделены из криптWT 19. Было исследовано, может ли экспрессия EphB2 различать четыре группы с экспрессией специфических маркеров, таких как ISC-специфические маркерные гены (Lgr5, Ascl2 и Olfm4) и гены-маркеры-предшественники клеток (Ki67, Myc и FoxM1). Этот эксперимент показал, чтовысокие клетки EphB2 были преимущественно ISC, в отличие отмедицинских клеток EphB220,21. Наконец, на основе метода выделения клеток полученные клетки были разделены на четыре группы (EphB2 высокий, EphB2медицинский, EphB2низкий и EphB2neg клетки) (рис. 2). Затем отдельные клетки, экспрессирующие высокие уровни EphB2, отсортированные FACS, культивировали для роста органоидов. Одинвысококлеточный EphB2 может быть самостоятельно применен для локализованного лечения и воссоздания самоорганизующихся крипт-ворсинчатых структур, напоминающих нормальную тонкую кишку (рис. 3). Однако клетки, полученные из других групп (EphB2med, EphB2low и EphB2neg), не генерируют органоиды20.
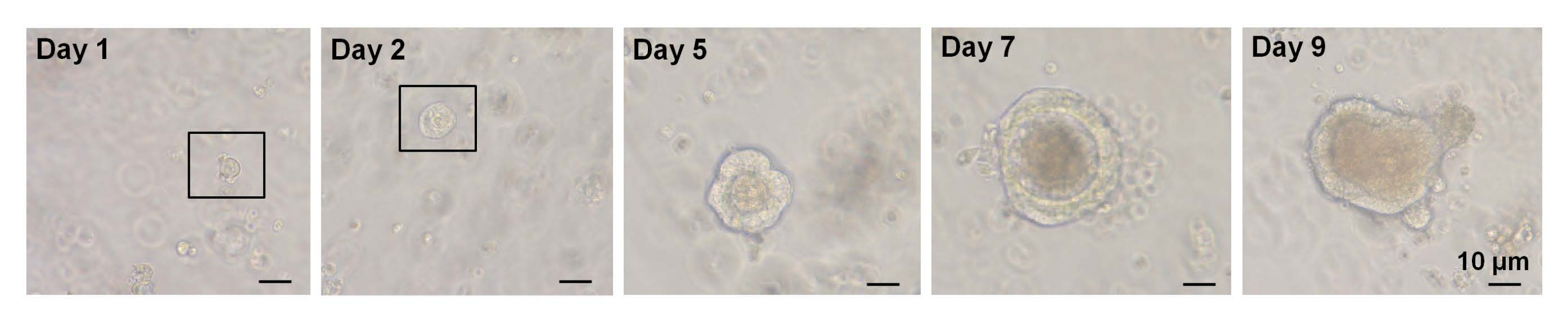
В предыдущем исследовании ~ 6% одинарно отсортированных Lgr5-GFPhi-клеток смогли инициировать крипт-ворсинчатые органоиды7. Однако оставшиеся клетки не смогли генерировать органоиды и погибли в течение первых 12 ч7. Авторы предположили, что это было результатом физического и/или биологического стресса, присущегопроцедуре изоляции7. Менее 6% роста органоидов также было получено из односортныхвысоких клеток EphB2 у мышей WT. К 5-му дню культуры образовались сфероидоподобные структуры (рис. 3). С 7-го по 9-й день происходила эвагинация пятен с образованием крипт (рис. 3). Важно отметить, что применение выбранного ингибитора ROCK к односортнымвысоким клеткам EphB2 уменьшало апоптоз, вызванный диссоциацией, и повышало эффективность роста органоидов.

Рисунок 1: Генерация органоидов тонкой кишки мыши. (A) Крипты, полученные комбинацией хелатирования ЭДТА и механической диссоциации. (B) Полученные очищенные крипты. (C) Крипты, встроенные во внеклеточный матрикс. (А-С) Черные стрелки обозначают склепы. (D) Трехмерная культура крипт и органоидов. (E) Репрезентативные изображения растущего органоида, полученного из склепа. Белые стрелки указывают на почку крипты. Масштабные линейки = (A-C) 100 мкм и (E) 50 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Стратегия стробирования проточной цитометрии для получения популяции EphB2-положительных (EphB2+) клеток у мышей дикого типа . (A) Прямые и боковые диаграммы рассеяния используются для разделения клеток в соответствии с их размером и зернистостью соответственно. (B) Флуоресцентное рассеяние используется для разделения жизнеспособных клеток в соответствии с интенсивностью флуоресценции клеток 7-AAD (PerCP). Были выбраны ворота для 7-AAD-отрицательной клеточной популяции. (C) Были выбраны ворота для популяций клеток EphB2 с высоким (EphB2 высоким), EphB2-средним (EphB2med), EphB2-низким (EphB2низким) и EphB2-отрицательным (EphB2neg) клеточными популяциями. Сокращения: FSC-A = площадь пика прямого рассеяния; SSC-A = площадь пика бокового рассеяния; 7-AAD = 7-аминоактиномицин D. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Временной ход односортногороста высококлеточного органоида EphB2 у мышей дикого типа. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Таблица 1: Питательная среда для 24-луночного планшета. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Дополнительное видео S1: Покадровые изображения растущего органоида. Масштабная линейка = 50 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S1: Репрезентативное изображение окрашивания антител к лизоциму в органоиде. Белыми стрелками обозначены ячейки Панета. Аббревиатура: DIC = дифференциальный интерференционно-контрастный микроскоп. Масштабная линейка = 10 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Discussion
В этом протоколе описывается метод последовательной изоляции сккриптов тонкой кишки и последующего культивирования 3D-органоидов. Для повышения скорости высвобождения крипты был разработан метод механической изоляции, включающий энергичное встряхивание после обработки ЭДТА. Состав среды отличается от исходного протокола Sato et al.7. Оригинальный носитель относительно дорогой. Таким образом, питательная среда и специализированная среда для органоидов тонкой кишки мышей, содержащие фармакологические ингибиторы, рекомбинантные факторы роста и/или кондиционированные среды, показаны в таблице 1. Wnt3A и N-ацетилцистеин не включены в питательную среду в этом протоколе. Поскольку клетки Панета экспрессируют Wnt3, клетки производят Wnt3 и поддерживают обслуживание ISC. Кроме того, при выделении крипты кондиционированная среда не используется. Органоидная модель является динамической и имеет клеточную и структурную гетерогенность (клетки Панета, энтероциты, бокаловидные клетки, энтероэндокринные клетки, пучковые клетки и МСК). Следовательно, эти органоиды могут быть использованы в масштабе для изучения фундаментальных вопросов биологии органоидов.
Градиент EphB2 поддерживает стволовость и пролиферацию ISC вдоль оси крипты-ворсинок в тонкой кишкевзрослого человека 18. Преимущество получения органоидов из одной клетки EphB2 по сравнению с изолированными криптами связано с пониманием биологии мышиных ISC, поскольку ISC играют ключевую роль в различных кишечных расстройствах человека. Одиночные EphB2 свысокой экспрессией ISC могут культивироваться с образованием органоидов аналогично развитию органоидов из одиночных Lgr5-экспрессирующих ISC. Наиболее важным шагом является точное разделение клеток на четыре группы (EphB2высокий, EphB2медицинский, EphB2низкий и EphB2neg) в соответствии с экспрессией EphB2 в криптах с использованием FACS. Графики прямого и бокового рассеяния (FSC и SSC) обычно используются для идентификации интересующих ячеек на основе их размера и детализации. FSC указывает размер ячейки, а SSC относится к сложности или детализации ячейки в затворе P0 (рис. 2A). В этой работе клетки, попавшие в определенные ворота (P0), были впоследствии проанализированы на жизнеспособность. Затем их жизнеспособность определяли по отрицательным и положительным популяциям сигналов флуоресценции 7-AAD. Граница между 7-AAD-отрицательными и -положительными клетками была строго определена так, чтобы получить отрицательные с минимальным положительным клеточным загрязнением. Вентили EphB2 были примерно установлены на основе градуированного выражения EphB2.
Чтобы подтвердить, что четыре группы были точно разделены, была проанализирована экспрессия мРНК выбранных генов. Уровни мРНК маркеров ISC высоки вклетках EphB2 с высоким уровнем 20. Кроме того, уровни мРНК маркеров, специфичных для клеток-предшественников, относительно высоки вмедицинских клетках EphB220. Тем не менее, эксэкспрессия EphB2 в клетках EphB2 с низким уровнем иEphB2 neg является низкой или отрицательной по сравнению с экспрессией EphB2с высоким уровнем имедицинскими клетками EphB2 20. Предыдущие меры должны быть приняты для обеспечения обогащениявысококлеточной популяции EphB2 перед нанесением покрытия. Однако рост органоидов менее чем на 6% изклеток с высоким содержанием EphB2 может быть обусловлен гибелью стволовых клеток во время процесса культивирования, а не энергичным встряхиванием во время выделения крипты. Было показано, что применение селективного ингибитора Rho-ассоциированной киназы (ROCK) к эмбриональным стволовым клеткам человека заметно уменьшает индуцированный диссоциацией апоптоз22. Таким образом, в качестве технического изменения стоит попробовать добавить ингибитор ROCK в более высокой концентрации и с более длительной инкубацией для повышения жизнеспособности.
Wnt3A-секретирующие клетки Paneth рядом с ISC обеспечивают существенную поддержку ISC8. Действительно, дублеты ячеек ISC-Paneth демонстрируют значительно повышенную органоидообразующую способность по сравнению с одиночными ISC8. Кроме того, было показано, что добавление Wnt3A в концентрации 100 нг/мл в течение первых 3 дней культивирования увеличивает органоидообразующую способность8. Таким образом, в качестве еще одного технического изменения добавление экзогенного Wnt3A может улучшить органоидообразующую способность одиночных EphB2с высокой экспрессией ISC.
По сравнению с подходами in vivo, органоиды могут быть легко использованы для генетических манипуляций, анализа фенотипов злокачественных новообразований и скрининга лекарств20,23. Комбинация хелатирования ЭДТА и метода механической изоляции является эффективной, воспроизводимой и эффективной по времени для создания органоидов тонкой кишки из крипт и может быть легко использована персоналом лаборатории без какого-либо передового опыта. Таким образом, добавление механической изоляции с энергичным встряхиванием после обработки ЭДТА может эффективно установить органоиды тонкой кишки мышей ex vivo и предоставить потенциальный инструмент для культивирования органоидов и моделирования заболеваний других эпителиальных тканей взрослого человека.
Эпителиальные клетки кишечника поляризованы и ориентированы апикальной стороной, направленной к просвету. Однако апикальная сторона, обращенная к просвету 3D-органоидов, находится внутри них. Таким образом, эта организация предотвращает доступ к апикальной стороне, что является проблемой при изучении воздействия компонентов просвета, таких как питательные вещества, микробы и метаболиты, на эпителиальные клетки. Чтобы обойти этот недостаток, была разработана культура органоидных клеток в виде2D-монослоев 24. С точки зрения будущих применений, будет использоваться культивирование монослоев органоидных клеток, поскольку это представляет собой наиболее эффективную и управляемую систему.
Disclosures
У авторов нет конфликтов интересов, о которых можно было бы заявить.
Acknowledgements
Эта работа была поддержана грантами на научные исследования (C) для T.T. (номера грантов JP17K07495 и JP20K06751). Мы благодарим профессора Минеко Кенгаку за использование оборудования для долгосрочной покадровой съемки (LCV100; Олимп).
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL Eppendorf tube | Eppendorf | 0030 125.215 | |
| 5 mL syringe | TERUMO | SS-05SZ | |
| 15 mL Falcon tube | Iwaki | 2325-015 | |
| 20 μm cell strainer | Sysmex | 04-004-2325 | |
| 24-well plate | Iwaki | 3820-024 | |
| 50 mL Falcon tube | Iwaki | 2345-050 | |
| 60 mm tissue culture dish | FALCON | 353002 | |
| 70 μm cell strainer | Falcon | 352350 | |
| 100 mm Petri dish | Iwaki | 3020-100 | |
| 7-AAD | BD Biosciences | 559925 | |
| Advanced DMEM/F12 | Gibco | 12634-010 | |
| Alexa Fluor 568 Goat Anti-Mouse IgG (H+L) | Invitrogen | A-11004 | |
| Anti-EphB2 APC-conjugated antibody | BD Biosciences | 564699 | |
| C57BL6/J mice | Japan SLC, Inc. | ||
| Clean bench | HITACHI | CCV-1306E | |
| Confocal laser scanning microscope | Olympus | FV3000 | |
| EDTA (0.5 mol/L) | Nacalai Tesque | 06894-14 | 2 mM |
| FACSMelody | BD Life Sciences-Biosciences | 661762 | |
| Fetal bovine serum | Sigma | 173012 | 1% (v/v) |
| Fiji (software) | https://fiji.sc/ | ||
| Gentamicin (10 mg/mL) | Nacalai Tesque | 16672-04 | 25 μg/mL |
| Hammacher laboratory scissor | SANSYO | 91-1538 | |
| Incubator | Panasonic | MCO-170-PJ | |
| Laboratory tweezer | AS-ONE | 7-164-04 | |
| L-Glutamine 200 mM | Gibco | 25030081 | 2 mM |
| Matrigel | BD Biosciences | 354230 | ECM for 3D organoids |
| Mouse Anti-Human Lysozyme | LSBio | LS-B8704-100 | |
| Murine EGF (20 μg/mL stock solution) | PeproTech | 315-09 | 20 ng/mL |
| PBS 1x | Gibco | 10010-023 | |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140-122 | 50 U/mL |
| Pipetman (10 μL, 20 μL, 200 μL, and 1,000 μL) | GILSON | 1-6855-12, -13, -15, and -16 | |
| Recombinant murine Noggin (20 μg/mL stock solution | R&D Systems | 1967-NG-025 | 100 ng/mL |
| Recombinant murine R-Spondin 1 (250 μg/mL stock solution) | R&D Systems | 3474-RS-050 | 500 ng/mL |
| Sorbitol | Nacalai Tesque | 32021-95 | 2% (w/v) |
| TE2000-S (inverted microscope) | Nikon | 24131 | |
| Time-lapse image microscope | Olympus | LCV100 | |
| TrypLE Express 1x | Gibco | 12605-010 | |
| ULVAC | ULVAC KIKO Inc. | 100073 | |
| Y-27632 | Fujifilm | 331752-47-7 | 10 μM |
References
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Seidlitz, T., et al. Human gastric cancer modelling using organoids. Gut. 68 (2), 207-217 (2019).
- Nikolaev, M., et al. Homeostatic mini-intestines through scaffold-guided organoid morphogenesis. Nature. 585 (7826), 574-578 (2020).
- Artegiani, B., Clevers, H. Use and application of 3D-organoid technology. Human Molecular Genetics. 27, R99-R107 (2018).
- Lancaster, M. A., Knoblich, J. A. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 345 (6194), 1247125 (2014).
- Dedhia, P. H., Bertaux-Skeirik, N., Zavros, Y., Spence, J. R. Organoid models of human gastrointestinal development and disease. Gastroenterology. 150 (5), 1098-1112 (2016).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Sato, T., et al. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature. 469 (7330), 415-418 (2011).
- Aronowitz, J. A., Lockhart, R. A., Hakakian, C. S. Mechanical versus enzymatic isolation of stromal vascular fraction cells from adipose tissue. Springerplus. 4 (1), 713 (2015).
- Takahashi, T. New trends and perspectives in the function of non-neuronal acetylcholine in crypt-villus organoids in mice. Methods in Molecular Biology. 1576, 145-155 (2019).
- Batlle, E., et al. β-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell. 111 (2), 251-263 (2002).
- Baghdadi, M. B., Kim, T. -. H. Analysis of mouse intestinal organoid culture with conditioned media isolated from mucosal enteric glial cells. STAR Protocols. 3 (2), 101351 (2022).
- Takahashi, T., et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS Journal. 281 (20), 4672-4690 (2014).
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nature Reviews Molecular Cell Biology. 15 (1), 19-33 (2014).
- Fordham, R. P., et al. Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell. 13 (6), 734-744 (2013).
- Miyoshi, H., et al. Wnt5a potentiates TGF-β signaling to promote colonic crypt regeneration after tissue injury. Science. 338 (6103), 108-113 (2012).
- Jung, P., et al. Isolation and in vitro expansion of human colonic stem cells. NatureMedicine. 17 (10), 1225-1227 (2011).
- Merlos-Suárez, A., et al. The intestinal stem cell signature identifies colorectal cancer stem cells and predicts disease relapse. Cell Stem Cell. 8 (5), 511-524 (2011).
- Mao, W., et al. EphB2 as a therapeutic antibody drug target for the treatment of colorectal cancer. Cancer Research. 64 (3), 781-788 (2004).
- Takahashi, T., et al. Muscarinic receptor M3 contributes to intestinal stem cell maintenance via EphB/ephrin-B signaling. Life Science Alliance. 4 (9), e202000962 (2021).
- Jung, P., et al. Isolation of human colon stem cells using surface expression of PTK7. Stem Cell Reports. 5 (6), 979-987 (2015).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Schulte, L., Hohwieler, M., Müller, M., Klaus, J. Intestinal organoids as a novel complementary model to dissect inflammatory bowel disease. Stem Cells International. 2019, 8010645 (2019).
- Puzan, M., Hosic, S., Ghio, C., Koppes, A. Enteric nervous system regulation of intestinal stem cell differentiation and epithelial monolayer function. Scientific Reports. 8 (1), 6313 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved