Расщепленная сетчатка как усовершенствованный плоский препарат для исследования нейронов внутреннего ядерного слоя сетчатки позвоночных
In This Article
Summary
В этой работе представлен альтернативный препарат плоской сетчатки, в котором удаление тел фоторецепторных клеток обеспечивает более быструю диффузию антител и улучшенный доступ к внутренним нейронам сетчатки с помощью пипетки для иммуногистохимии, гибридизации in situ и электрофизиологических экспериментов.
Abstract
Биполярные клетки и горизонтальные клетки сетчатки позвоночных являются первыми нейронами, которые обрабатывают визуальную информацию после того, как фотоны детектируются фоторецепторами. Они выполняют фундаментальные операции, такие как адаптация света, контрастная чувствительность, пространственная и цветовая оппоненция. Полное понимание точных схем и биохимических механизмов, которые управляют их поведением, будет способствовать развитию исследований в области визуальной нейробиологии и офтальмологической медицины. Тем не менее, современные препараты для исследования биполярных и горизонтальных клеток (целые срезы сетчатки и вертикальные срезы) ограничены в своей способности охватить анатомию и физиологию этих клеток. В данной работе мы представляем метод удаления телец фоторецепторных клеток из сетчатки живых плоских мышей, обеспечивающий расширенный доступ к биполярным и горизонтальным клеткам для эффективного пережатия пластырей и быстрого иммуномечения. Расщепленная сетчатка готовится путем зажатия изолированной сетчатки мыши между двумя кусочками нитроцеллюлозы, а затем осторожно отделяя их друг от друга. При разделении сетчатка расщепляется чуть выше внешнего плексиформного слоя с образованием двух кусочков нитроцеллюлозы, один из которых содержит тела фоторецепторных клеток, а другой — оставшуюся внутреннюю часть сетчатки. В отличие от вертикальных срезов сетчатки, препарат расщепленной сетчатки не разрывает дендритные отростки внутренних нейронов сетчатки, что позволяет регистрировать биполярные и горизонтальные клетки, которые интегрируют вклад сетей, связанных с щелевыми контактами, и широкопольных амакриновых клеток. Данная работа демонстрирует универсальность данного препарата для исследования горизонтальных и биполярных клеток в электрофизиологии, иммуногистохимии и экспериментах по гибридизации in situ .
Introduction
Сетчатка представляет собой тонкую нервную ткань, расположенную в задней части глаза, где свет перехватывается и преобразуется в электрохимический сигнал, который может быть интерпретирован мозгом. В задней части сетчатки фоторецепторы палочек и колбочек стимулируются светом, что снижает скорость тонического высвобождения нейромедиатора глутамата1. Первыми нейронами, которые испытывают и реагируют на это вызванное светом изменение концентрации глутамата, являются биполярные клетки (BC) и горизонтальные клетки (HC), чьи сомы находятся во внешней области внутреннего ядерного слоя (INL). Эти нейроны второго порядка выполняют первую стадию обработки сигналов в сетчатке и формируют важнейшие характеристики зрения, такие как адаптация к свету, контрастная чувствительность и пространственная/цветоваяпротивоположность. Несмотря на то, что эти функции приписываются БК и ГЦ, схемотехника и биохимические механизмы, лежащие в основе этих процессов, доконца не изучены. Поэтому развитие средств и методов изучения физиологии БК и ГК имеет первостепенное значение.
Вертикальные (поперечные) срезы сетчатки давно зарекомендовали себя как наиболее практичная модель для изучения БК и ГК; однако некоторые аспекты физиологии БК и ГЦ недоступны экспериментатору в рамках этой модели. Прямые записи от ГЦ или косвенные измерения их влияния на БК не отражают эндогенную связность сетчатки, поскольку латеральные отростки этих клеток разрываются во время среза. Цельные препараты сетчатки позволяют обойти эту проблему, сохраняя эти боковые отростки, но окружающие слои сетчатки создают проблему для доступа кэтим клеткам. Несмотря на то, что существует множество примеров иммуноокрашивания 5,6,7,8 и записи патч-клэмпов9 из нейронов INL в сетчатке всего маунта, существует возможность ускорить и упростить сбор этих данных. Ограничения, присущие поперечным сечениям, и проблемы, связанные со всей моделью монтировки, вдохновили на разработку этого альтернативного плоского препарирования сетчатки.
В следующей работе описывается протокол для легкого удаления фоторецепторного слоя с живой, плоской сетчатки для улучшения доступа к РМЖ и ГЦ для упрощения зажима пластырей и более быстрого и эффективного иммуномечения. Отслаивание двух кусков нитроцеллюлозной мембраны, прикрепленных к обеим сторонам изолированной сетчатки, разрывает ткань через аксоны фоторецепторов, оставляя расщепленную сетчатку, которая сохраняет внешний плексиформный слой (OPL) и все внутренние слои сетчатки. В то время как другие описывают протоколы механического разделения слоев сетчатки, эти методы либо плохо подходят для наложения заплаток и микроскопии, либо требуют утомительных манипуляций с тканями. Некоторые из этих методов требуют замороженной или лиофилизированной ткани для разделения слоев, что делает их несовместимыми с электрофизиологическими экспериментами10,11,12. Другие предназначены для живых тканей, но требуют 5-15 последовательных пилингов фильтровальной бумагой 4,11 или обработки трипсином 13 для удаления фоторецепторов. Описанный здесь метод является улучшением по сравнению со своими предшественниками, упрощая процедуру удаления фоторецепторов и расширяя репертуар последующих применений.
Protocol
Мышей давали воду и пищу вволю и поддерживали в 12-часовом цикле света/темноты. Мышей усыпляли путем воздействия изофлурана с последующим вывихом шейки матки. Все процедуры с животными соответствовали рекомендациям Национальных институтов здравоохранения и были одобрены Комитетом по уходу за животными и их использованию Орегонского университета здоровья и науки.
ПРИМЕЧАНИЕ: Энуклеация глаза, диссекция сетчатки и расщепление сетчатки должны выполняться как можно быстрее, чтобы сохранить здоровье живых тканей. Постарайтесь завершить вскрытие за < 4 минуты на каждый глаз. Эти три шага должны выполняться последовательно. Мыши дикого типа: Для экспериментов использовались взрослые (>3 месяца) самцы и самки мышей C57BL/6J. Для морфологии синапсов использовали мышей, экспрессирующих зеленый флуоресцентный белок (GFP) под промотором Pcp2 (Pcp2-cre/GFP)14 . Трансгенные мыши: Для горизонтальной визуализации клеток с помощью GFP во время иммуногистохимических или электрофизиологических экспериментов использовали тройную трансгенную мышь: vGATFLPo; vGlut2Cre; Аи80д. Штаммы vGATFlpo и vGluT2Cre представляют собой мышей, экспрессирующих Flpo или Cre-рекомбиназу после соответствующих промоторов. Мышь Ai80d является интерсекциональной репортерной мышью (CatCh/EYFP) и экспрессирует только Ca2+ проницаемый канальный родопсин (ChR2) в клетках, экспрессирующих рекомбиназы Cre и Flpo. Таким образом, тройная трансгенная мышь экспрессирует ChR2 только в клетках с историей экспрессии как VGAT, так и vGluT2.
1. Подготовка материалов для диссекции и расщепления сетчатки
- Подготовьте кусочки нитроцеллюлозной мембраны
ПРИМЕЧАНИЕ: Отделение расщепленной сетчатки от нитроцеллюлозной мембраны уменьшает фоновую флуоресценцию при микроскопии и упрощает запись с помощью патч-зажима. Удаление мембраны может быть выполнено до или после фиксации тканей. При фиксированном расщеплении сетчатки нет необходимости обрабатывать кусочки нитроцеллюлозной мембраны. При живом расщеплении сетчатки обработайте мембрану в соответствии с шагами 1.1.3 - 1.1.5, чтобы облегчить мягкое отделение от ткани.- Нарежьте 16 кусков (или более) нитроцеллюлозной мембраны на квадраты 5 мм х 5 мм. Экстра можно заготавливать навалом и хранить впрок.
- Отложите половину кусочков мембраны для последующего использования. Эти кусочки не будут обрабатываться блокирующим раствором.
- Инкубируйте оставшиеся кусочки в растворе ИГХ, не содержащем моющих средств (например, 3% лошадиной сыворотки + 0,025%NaN3 , разбавленный в PBS) в течение 10 минут при комнатной температуре, слегка встряхивая.
ВНИМАНИЕ: Используйте соответствующие СИЗ при работе с NaN3, так как он является сильнодействующим токсином. - Тщательно промойте кусочки мембраны путем инкубации в бикарбонатно-буферной среде Эймса в течение 10 минут при комнатной температуре, слегка встряхивая.
- Полностью высушите на воздухе заблокированные части мембраны (~20 мин). Маркируйте и храните кусочки мембраны при комнатной температуре, держа их отдельно от необработанных частей мембраны.
- Подготовьте медиа Эймса
- Приготовьте бикарбонатно-буферную среду Эймса и выдержите раствор при комнатной температуре при постоянном карбогенировании (95% О2 и 5%СО2).
2. Энуклеация глаз мыши
- Усыпите мышь любым доступным методом в соответствии с рекомендациями IACUC.
- Переверните мышь на одну сторону и двумя пальцами осторожно надавите на глазницу. Это приведет к тому, что глаз будет выпячиваться из черепа.
- Используя изогнутые ножницы для вскрытия, сделайте надрез под выпуклым глазом, чтобы перерезать зрительный нерв и отделить глаз от черепа.
- Зачерпните глаз ножницами и поместите его в чашку Петри, наполненную ледяной средой Эймса.
ПРИМЕЧАНИЕ: Для последующих применений, в которых ткань будет зафиксирована после расщепления, вместо среды Эймса можно использовать ледяную PBS. - Повторите шаги 2.1 - 2.4 для оставшегося глаза.
3. Диссекция сетчатки
- Используйте специальную стеклянную пипетку для переноса одного глаза в новую чашку Петри, содержащую свежую, ледяную среду Эймса.
ПРИМЕЧАНИЕ: Широкое отверстие специальной трансферной пипетки предотвращает случайное сжатие ткани, а использование стекла сводит к минимуму прилипание ткани к стенкам пипетки. Тем не менее, пластиковая трансферная пипетка с широким горлышком также приемлема, если экспериментатор уже владеет этим инструментом. - Используйте щипцы, чтобы стабилизировать глаз, прикрепив его дополнительную соединительную ткань ко дну чашки Петри. Затем проколите ушко вдоль линии ora serrata с помощью иглы 25G, чтобы создать точку входа для ножниц Vannas.
- Используйте ножницы Vannas, чтобы разрезать вдоль линии ora serrata до тех пор, пока роговица не освободится от остальной части глаза (дополнительный рисунок 1A). Извлеките линзу из наглазника с помощью щипцов (дополнительный рисунок 1B).
- С помощью специальной стеклянной пипетки перенесите наглазник на большой объем (≥100 мл) карбогенированного препарата Эймса и повторите шаги 3.1–3.3 с оставшимся глазом.
ПРИМЕЧАНИЕ: Наглазники помещаются в карбогенированный Эймс для поддержания здоровья тканей, пока выполняется вскрытие другого глаза. - Переложите один наглазник в чашку Петри, наполненную свежекарминированным Эймсом.
- Используя ножницы Ванны, сделайте небольшой надрез внутрь от края склеры, затем с помощью двух пар щипцов отделите склеру от сетчатки (дополнительный рисунок 1C). Избегайте захвата сетчатки щипцами. Вместо этого раздвиньте лоскуты склеры, созданные ножницами.
- С помощью ножниц Vannas перережьте зрительный нерв, соединяющий склеру и сетчатку (дополнительный рисунок 1D), затем осторожно подденьте сетчатку от склеры с помощью ножниц или щипцов, чтобы изолировать сетчатку. (Рисунок 1А).
ПРИМЕЧАНИЕ: Несмотря на то, что РПЭ обычно остается прикрепленным к наглазнику, никаких дополнительных действий для снятия РПЭ в случае, если он прикреплен к сетчатке, не требуется. На этом этапе края сетчатки могут быть дополнительно подрезаны скальпелем, чтобы предотвратить скручивание на этапе уплощения (рис. 1B). - С помощью скальпеля разрежьте сетчатку на половинки или четвертинки (рис. 1C), затем с помощью специальной пипетки для переноса верните кусочки в большой объем (≥ 100 мл) непрерывно карбонированной среды Эймса.
ПРИМЕЧАНИЕ: Выбор половинок или четвертин субъективен. Выберите оптимальный вариант для нужного применения. - Повторите шаги 3.5 - 3.8 для оставшегося глаза, прежде чем приступать к расщеплению сетчатки.
4. Расщепление сетчатки
- Выбросьте среду Эймса из чашек Петри и замените ее свежекарбонированной жидкостью Эймса.
ПРИМЕЧАНИЕ: Чтобы сохранить карбогенизацию на протяжении оставшейся части процедуры расщепления сетчатки, заменяйте среду в чашке Петри свежекарбонизированным материалом Эймса примерно каждые 5 минут. - С помощью специальной пипетки поместите кусочек сетчатки на предметное стекло (7,5 см x 5 см) ганглиозной стороной вверх, затем разровняйте его, удалив окружающую жидкость деликатной салфеткой (рисунок 1D). При необходимости аккуратно потяните края сетчатки тонким кончиком кисти под рассекающим микроскопом.
- С помощью щипцов опустите сухой кусочек нитроцеллюлозной мембраны размером 5 мм x 5 мм на сетчатку, заставляя ее прилипать к ганглионарной клетке (рис. 1E).
ПРИМЕЧАНИЕ: Если требуется удаление мембраны из живой ткани (например, для электрофизиологии), используйте для этого шага сухой кусок мембраны, обработанной сывороткой (см. шаги 1.1.3 - 1.1.5 для получения подробной информации). Это снижает прочность адгезии к слою ганглиозных клеток, облегчая удаление сетчатки из нитроцеллюлозы после расщепления. - Переверните сетчатку так, чтобы нитроцеллюлоза лежала на предметном стекле, и поместите сухой кусок мембраны размером 5 мм x 5 мм на фоторецепторную сторону сетчатки (рис. 1F).
- Прикоснитесь смоченным кончиком кисти к пространству между двумя мембранами и позвольте капиллярному действию всосать Эймса в бутерброд (Рисунок 1G). Это уменьшает сцепление мембран с сетчаткой и необходимо только в том случае, если сетчатка была чрезмерно высушена деликатной салфеткой.
ПРИМЕЧАНИЕ: Если сетчатка потеряла свой блестящий вид, она была чрезмерно высушена, и необходимо выполнить шаг 4.5. - Чтобы обеспечить равномерное прилипание, надавите на верхнюю мембрану влажной кистью снизу вниз (Рисунок 1H).
- Прикрепляя нижнюю мембрану к стеклу одной парой щипцов, медленным, устойчивым движением осторожно отклейте верхнюю мембрану второй парой щипцов. Это приведет к тому, что сетчатка расщепится чуть выше OPL (рис. 1I).
- Выбросьте верхнюю мембрану, содержащую фоторецепторы (рис. 1J, слева). Нижняя мембрана содержит внутреннюю сетчатку, далее называемую расщепленной сетчаткой (рис. 1J, справа).
- Немедленно верните расщепленную сетчатку в карбонизированную среду Эймса.
ПРИМЕЧАНИЕ: Для экспериментов на живых тканях сетчатке может быть полезен 15-30-минутный период восстановления у карбогенированного Эймса после расщепления.

Рисунок 1: Процедура расщепления сетчатки. (A) После энуклеации и подготовки наглазника в холодной среде PBS или среде Эймса изолируйте сетчатку мыши от наглазника и замените PBS карбонизированной средой Эймса комнатной температуры. (B) С помощью скальпеля обрежьте края сетчатки до тех пор, пока не останется областей с загибом внутрь (необязательно). (C) Разрежьте сетчатку на четвертинки или половинки с помощью скальпеля. (D) Поместите один кусочек сетчатки на предметное стекло (ганглиозной стороной вверх) с помощью специальной трансферной пипетки и удалите все излишки Эймса с помощью деликатной салфетки. Убедитесь, что полусухая сетчатка лежит ровно на стекле, прежде чем переходить к следующему шагу. Используйте смоченный Эймсом кончик кисти, чтобы аккуратно развернуть участки сетчатки, которые не являются плоскими. (E) С помощью щипцов поместите предварительно вырезанный кусок сухой нитроцеллюлозной мембраны (5 мм x 5 мм) на уплощенную сетчатку. (F) Переверните кусочек нитроцеллюлозы так, чтобы фоторецепторная сторона сетчатки была обращена вверх. Затем поместите еще один сухой кусочек мембраны на сетчатку. (G) Прикоснитесь влажным кончиком щетки к пространству между двумя мембранами и позвольте капиллярному действию втянуть Эймса в бутерброд. Это уменьшает сцепление мембран с сетчаткой и необходимо только в том случае, если сетчатка была чрезмерно высушена деликатной салфеткой. (H) Влажным кончиком кисти осторожно надавите вниз на центр зажатой сетчатки. (I) Используйте одну пару щипцов, чтобы приколоть нижнюю часть мембраны к предметному стеклу, в то время как другой парой щипцов осторожно отделите верхний кусок мембраны от нижнего. Внутренняя часть сетчатки (слева) остается на нижней мембране, в то время как фоторецепторы (справа) оттягиваются вместе с верхней мембраной. Панели (A), (B), (C), (D) и (J) были получены с помощью диссекционного микроскопа; масштабная линейка представляет собой приблизительно 1 мм; панели (E-I) были получены с помощью камеры смартфона без увеличения. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
5. Подготовка расщепленной сетчатки к иммунофлюоресцентным экспериментам
ПРИМЕЧАНИЕ: Расщепленная сетчатка будет прикреплена к нитроцеллюлозной мембране до шага 5.5. Выполните шаги 5.1, 5.2, 5.3 или 5.4, а не все четыре, так как они предназначены для разных экспериментов.
ВНИМАНИЕ: Используйте соответствующие СИЗ и будьте осторожны при обращении с параформальдегидом (фиксатором).
- Подготовка к флэтмаунтовой иммунофлюоресценции
- Инкубируйте расщепленную сетчатку в 4% параформальдегиде на льду в течение 30 минут, используя достаточное количество раствора, чтобы полностью покрыть сетчатку.
- Промыть расщепленную сетчатку 3 раза в 5-10 мл PBS комнатной температуры. Необязательная пауза: расщепленную сетчатку можно оставить в PBS при температуре 4 °C на срок до 24 часов.
- Препарат для иммунофлюоресценции с вертикальными срезами расщепленной сетчатки
- Инкубируйте расщепленную сетчатку в 4% параформальдегиде на льду в течение 30 минут, используя достаточное количество раствора, чтобы полностью покрыть сетчатку.
- Промыть расщепленную сетчатку 3 раза в 5-10 мл PBS комнатной температуры. Необязательная пауза: расщепленную сетчатку можно оставить в PBS при температуре 4 °C на срок до 24 часов.
- Не снимая мембраны, последовательно погружайте расщепленную сетчатку в 10%, 20% и 30% сахарозу при 4 °C на 1 ч каждый для криозащиты ткани.
- Поместите криозащищенные расщепленные сетчатки в компаунд с оптимальной температурой резки (O.C.T.) и храните их при -80 °C (до 6 месяцев) до криосекции.
- Извлеките встроенную расщепленную сетчатку при температуре -80 °C и с помощью криостата вырежьте участки толщиной 20 мкм. Установите срезы на предметные стекла микроскопа с электростатическим зарядом, дайте им высохнуть на воздухе, а затем храните их при температуре -20 °C до 6 месяцев.
- Подготовка к двойной флуоресцентной гибридизации in situ и иммуногистохимии
- Инкубируйте расщепленную сетчатку в 4% параформальдегиде на льду в течение 2 ч, используя достаточное количество раствора, чтобы полностью покрыть сетчатку.
- Промыть расщепленную сетчатку 3 раза в 5-10 мл PBS комнатной температуры. Необязательная пауза: расщепленную сетчатку можно оставить в PBS при температуре 4 °C на срок до 24 часов.
- Подготовка к электрофизиологии
- Подготовьте пластырные пипетки, вытягивая толстостенные пипетки из боросиликатного стекла с нитью накала с помощью съемника микропипеток. Используйте только пипетки с измеренным сопротивлением от 6 до 10 МОм.
- Обратно заполняют вытянутые пипетки внутренним раствором, содержащим (в мМ): 125 К-глюконат, 8 ККЛ, 5 ГЕПЭС, 1 MgCl2, 1CaCl2, 0,2 EGTA, 3 АТФ-Mg и 0,5 ГТФ-Na.
- Удаление расщепленной сетчатки с нитроцеллюлозной мембраны
- С помощью гидрофобной барьерной ручки подготовьте круглые лунки на предметном стекле микроскопа (~1 см в диаметре) и дайте им высохнуть на воздухе в течение 5-10 минут.
- Поместите расщепленные сетчатки в подготовленные углубления для гидрофобных барьерных ручек и добавьте достаточное количество PBS, чтобы полностью покрыть их.
- Под микроскопом просуньте щетинки тонкой кисти под края ткани и аккуратно приподнимите вверх. Таким образом, обработайте сетчатку по кругу, чтобы оторвать ее от мембраны.
- Используйте щипцы, чтобы удалить мембрану из-под плавающей части сетчатки.
- Осторожно отсосите оставшуюся PBS так, чтобы кусочек сетчатки остановился на предметном стекле микроскопа ганглионарной стороной вниз.
ПРИМЕЧАНИЕ: Следующие шаги не должны выполняться последовательно. Выберите подходящий протокол для желаемого применения (например, иммуноокрашивание или гибридизация с двойной флуоресценцией in situ [FISH] и иммуногистохимия [IHC] или электрофизиология).
6. Иммуноокрашивание
- Если вы еще не подготовлены, используйте гидрофобную барьерную ручку, чтобы создать круглые лунки на предметном стекле микроскопа (~1 см в диаметре) и дайте им высохнуть на воздухе в течение 5-10 минут. Все этапы инкубации и промывки будут выполняться в этих колодцах.
- Инкубируйте расщепленную сетчатку или вертикальные расщепленные участки сетчатки в инкубационном растворе антител (AIS: 3% лошадиная сыворотка, 0,5% Triton X-100, 0,025% NaN3 в PBS) в течение 30 минут при комнатной температуре.
- Инкубируют расщепленную сетчатку или вертикальные расщепленные участки сетчатки с первичными антителами, разведенными в AIS, в течение 1 ч при комнатной температуре.
ПРИМЕЧАНИЕ: Время инкубации первичных антител потребует оптимизации для различных белковых мишеней и антител. - Постирайте салфетку 3 раза при комнатной температуре PBS.
- Инкубируют ткань со вторичными антителами, разведенными в АИС, в течение 1 ч при комнатной температуре. Постирайте салфетку 3 раза при комнатной температуре PBS.
- Если требуется ядерное окрашивание, инкубируют ткань с DAPI, разведенным в PBS, в течение 30 с при комнатной температуре. Постирайте салфетку 1 раз при комнатной температуре PBS.
- Нанесите каплю скользящего монтажного носителя на каждый кусок ткани и установите стеклянный покровный лист.
- Нанесите лак для ногтей по краям покровного стекла, чтобы запечатать образец. Храните предметное стекло при температуре 4 °C.
7. Двойные FISH и IHC
- Выпекайте расщепленную сетчатку при температуре 40 °C в течение 30 минут в гибридизационной печи, чтобы усилить сцепление с предметным стеклом.
- Завершите протокол RNAscope FISH в соответствии с протоколом производителя со следующими исключениями и изменениями:
- Этап извлечения антигена не требуется. Используйте протеазу III с инкубационным временем 18 мин при комнатной температуре.
- Выполните все действия по промывке на горке в лунках, сделанных гидрофобным барьерным загоном.
- Инкубируют образцы в первичных антителах, разведенных (см. таблицу материалов) в PBS в течение 30 мин при 40 °C в гибридизационной печи. Промыть образцы 3 раза при комнатной температуре ПБС.
- Инкубируют образцы во вторичных антителах, разведенных (см. таблицу материалов) в PBS в течение 30 мин при 40 °C в гибридизационной печи. Промыть образцы 3 раза при комнатной температуре ПБС.
- Инкубируют образцы в 1x DAPI в течение 30 с при комнатной температуре. Промыть образцы 1 раз при комнатной температуре ПБС.
- Нанесите каплю монтажного носителя с защитой от выцветания на каждый кусок ткани и установите стеклянный защитный лист.
- Нанесите лак для ногтей по краям покровного стекла, чтобы запечатать образец. Храните предметное стекло при температуре 4 °C.
8. Электрофизиология
- После удаления нитроцеллюлозной мембраны переместите расщепленную сетчатку в записывающую камеру с патч-зажимом и аккуратно закрепите ее на месте платиновой арфой.
- На протяжении всего эксперимента непрерывно перфузируйте расщепленную сетчатку раствором Эймса, карбогенизированным 95% О2 и 5%СО2. Выдерживайте раствор при температуре 32-34 °C.
ПРИМЕЧАНИЕ: Во время эксперимента ткань может быть визуализирована с помощью градиентной контрастной микроскопии Додта. - При комнатном освещении выполните зажим напряжения всей ячейки для записи с нейронов INL.
- Во время записи имитируйте световые реакции с помощью микроклеточного инъекционного устройства для нанесения фармацевтических соединений или светодиода с длиной волны 470 нм для стимуляции каналродопсина (ChR2).
ПРИМЕЧАНИЕ: Интенсивность света можно измерить с помощью цифрового измерителя оптической мощности.
- Во время записи имитируйте световые реакции с помощью микроклеточного инъекционного устройства для нанесения фармацевтических соединений или светодиода с длиной волны 470 нм для стимуляции каналродопсина (ChR2).
9. Конфокальная микроскопия
- Для конфокальной иммунофлуоресценции делайте снимки с помощью конфокального микроскопа с помощью масляного иммерсионного объектива 40x/1,3 или 63x/1,40. Используйте FIJI для настройки яркости и контрастности, а также для создания Z-проекций из стеков изображений.
Representative Results
Расщепление сетчатки сохраняет клеммы фоторецепторов
Чтобы подтвердить, что расщепление сетчатки не повреждает дендриты нейронов второго порядка в OPL, вертикальные участки расщепленной сетчатки были окрашены антителами против белка синаптического везикула синаптофизина (зеленый) и протеинкиназы С альфа (PKCα; красный). Интенсивная полоса синаптофизиновой метки в верхней части расщепленной сетчатки указывает на то, что синаптические окончания фоторецепторов сохранены (рис. 2). Кроме того, окрашивание PKCα выявляет нормальную морфологию палочек биполярных клеток (эритроцитов). Ядра фоторецепторов не видны, что указывает на то, что сетчатка разделена между OPL и самым внутренним рядом тел фоторецепторных клеток (рис. 2).

Рисунок 2: Расщепленная сетчатка сохраняет фоторецепторные окончания. Флуоресцентные конфокальные микрофотографии, показывающие вертикальный поперечный срез сетчатки слюны, которая была криосекционирована (толщиной 20 мкм) после процедуры расщепления. Каждое изображение представляет собой максимальную проекцию конфокального z-стека. Срез был иммуномечен антителами против PKCα (вверху в центре) и синаптофизина (вверху справа) для визуализации эритроцитов и синаптических везикул соответственно. На объединенном изображении (внизу) показаны синаптические везикулы (зеленый), которые находятся в терминалях фоторецепторов, чуть выше апикальных отростков эритроцитов (красный) в OPL. Клеточные ядра помечены DAPI (синим цветом). В ОНЛ не видно ядер фоторецепторов. Сокращения: ONL = внешний ядерный слой; OPL = наружный плексиформный слой; INL = внутренний ядерный слой; IPL = внутренний плексиформный слой; GC = ганглиозные клетки. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Морфология синапсов в OPL сохраняется после расщепления сетчатки
При использовании мыши, экспрессирующей GFP в эритроцитах под промоторомpcp2 14, пре- и постсинаптические белки в OPL были иммуномечены для оценки целостности этого синаптического слоя после расщепления14. Несмотря на сдвиговые силы, происходящие через аксоны фоторецепторов, расщепление не нарушает морфологию фоторецептор-БК-синапсов в ОПЛ, так как наблюдается нормальное расположение дендритов эритроцитов, меченных для RGS11, и синаптических лент фоторецепторов, меченных для CtBP215 (рис. 3). Для каждого синаптического контакта между палочками и эритроцитами RGS11 можно увидеть в виде красных точек, лежащих в пределах подковообразной формы синаптических лент (зеленый). В последующем эксперименте антитело 16 к GPR179 было использовано для мечения постсинаптических дендритных кончиков16 ON-BC, а антитело против PSD-95 было использовано для мечения клемм фоторецепторов пресинаптической палочки (дополнительный рисунок 2). Эти результаты еще раз подтверждают стабильность OPL в препарате расщепленной сетчатки, поскольку показано, что дендриты эритроцитов тесно связаны со своим пресинаптическим партнером, палочкодержателями.

Рисунок 3: Морфология синапсов в OPL сохраняется после расщепления сетчатки. Конфокальные иммунофлуоресцентные изображения расщепленной сетчатки трансгенной мыши, экспрессирующей GFP в эритроцитах под промотором Pcp2. Уровни экспрессии GFP (синий) варьируются в зависимости от эритроцитов в сетчатке. После расщепления сетчатку фиксировали, а затем инкубировали с антителами против CtBP2 (зеленый) и RGS11 (красный) для мечения синаптических лент фоторецепторов и дендритных кончиков ON-BC соответственно. Каждая красно-зеленая пара представляет собой синаптический контакт между палочкой и ON-BC. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Расщепление сетчатки поддерживает жизнеспособность эритроцитов
Для оценки жизнеспособности нейронов внутренней сетчатки после расщепления использовали мембранонепроницаемый ближний инфракрасный ядерный краситель (MI-NIR), который позволяет идентифицировать мертвые клетки. После инкубации с MI-NIR расщепленные сетчатки фиксировали, а затем помечали анти-PKCα для идентификации эритроцитов. Конфокальные микрофотографии расщепленной сетчатки показывают региональную вариабельность жизнеспособности клеток в ткани, при этом в некоторых областях наблюдается более высокая скорость гибели клеток, чем в других. Эта вариабельность может быть результатом повреждения, нанесенного определенным областям сетчатки во время процедур рассечения, расщепления или обработки (рисунок 4). Учитывая, что клеточные тела эритроцитов находятся во внешней области INL, близко к месту расщепления, была оправдана тщательная оценка их жизнеспособности. Скудная колокализация PKCα и MI-NIR подтвердила, что большинство эритроцитов остаются жизнеспособными после расщепления сетчатки (рис. 4).

Рисунок 4: Палочковидные биполярные клетки жизнеспособны после расщепления сетчатки. Флуоресцентные конфокальные микрофотографии, показывающие область расщепленной сетчатки в плоской перспективе. После расщепления живую сетчатку инкубировали с красителем MI-NIR (красный) в течение 30 мин при 37 °C. Затем сетчатку фиксировали и иммунометировали антителами против PKCα для визуализации эритроцитов. В этой области сетчатки колокализация PKCα и MI-NIR встречается нечасто. MI-NIR локализуется с ядрами (синего цвета), которые не принадлежат эритроцитам. Сокращения: MI-NIR = мембранный непроницаемый NIR живой/мертвый цвет. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Расщепленная сетчатка поддается двойному FISH и IHC
Увеличивая время фиксации стандартного ВПХ, расщепленные сетчатки могут быть последовательно обработаны FISH и IHC для одновременного мечения мРНК и белков17,18. Эксперименты подтвердили, что 2-часовая фиксация в 4%-ном параформальдегиде дает надежное мечение мРНК, сохраняя при этом белковые эпитопы для связывания с антителами. FISH проводили на расщепленной сетчатке с последующей ИГХ для визуализации экспрессии субъединицы рецептора ГАМКА δ (GABRD; антисмысловые мРНК-зонды) в зависимости от положения эритроцитов (антител к PKCα) во внешнем INL (рис. 5A). Экспрессия мРНК GABRD встречается редко в эритроцитах (рис. 5A); однако транскрипт обильно экспрессируется амакриновыми клетками и ганглиозными клетками, о чем свидетельствует рисунок мечения на поперечных срезах интактной сетчатки (рис. 5B). Во внешнем INL (рис. 5A) мРНК GABRD распределена более равномерно по сравнению с внутренним INL (рис. 5C), где она сосредоточена в отдельных клетках. Антисмысловые зонды, нацеленные на другие субъединицы ГАМК-рецепторов, производят отчетливые паттерны мечения, демонстрирующие специфичность зондов (данные не показаны).

Рисунок 5: Двойные FISH и IHC в расщепленной сетчатке и интактной сетчатке. (A, C) Конфокальные микрофотографии плоской расщепленной сетчатки и (B) вертикальный срез интактной сетчатки. Изображения на рисунках (А) и (В) являются максимальными проекциями оптических сечений в верхней и нижней областях INL соответственно. Пунктирные прямоугольники на рисунке (B) представляют собой приблизительные границы, используемые для создания проекций, показанных на рисунках (A) и (C). Расщепленную сетчатку (А, С) фиксировали на 2 ч, затем помечали антисмысловыми мРНК-зондами против GABRD (красный). После этого расщепленную сетчатку окрашивали антителами против PKCα, чтобы пометить эритроциты (зеленым цветом). Канал PKCα был опущен в проекциях нижнего INL для наглядности. Интактную сетчатку в (В) фиксировали за 24 ч до секционирования. После этого фиксированная сетчатка была помечена антисмысловыми мРНК-зондами против GABRD (красный). Все образцы окрашивали DAPI (синий) в течение 20 с перед монтажом покровного стекла. Сокращения: INL = внутренний ядерный слой. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Расщепленная сетчатка хорошо подходит для регистрации электрофизиологии с помощью патч-зажимов от РМЖ и ГК
Чтобы наложить патч на сому BC или HC в традиционную цельную сетчатку, пипетка должна подходить либо со стороны ганглиозных клеток, либо со стороны фоторецепторов. Оба подхода требуют прохождения нескольких слоев сетчатки, чтобы добраться до INL, во время чего кончик пипетки часто засоряется мусором. При приготовлении вибратомного среза сомы BC и HC легко доступны, но их дендритные отростки могут быть разорваны, нарушая их боковые связи. Однако в расщепленной сетчатке клеточные тела эритроцитов и HC располагаются на поверхности ткани, обеспечивая значительно улучшенный доступ к патч-пипеткам, сохраняя при этом латеральную схему OPL.
На рисунке 6 показаны химически смоделированные световые реакции, зарегистрированные от БК в расщепленной сетчатке. Перфузионную среду Эймса дополняли L-AP4 (4 мкМ), агонистом mGluR III группы, для моделирования высвобождения глутамата фоторецепторами в темноте. Антагонист mGluR6, CPPG (600 мкМ, по Эймсу), наносили на дендриты патчированной клетки (удерживаемые при -60 мВ) для имитации световой вспышки путем ингибирования mGluR6. Клетки реагировали на затяжки CPPG двумя типами внутренних токов. На одном типе показан переходный ток, за которым следует плато (рис. 6А), аналогичный каноническим токам, вызванным светом, зарегистрированным от эритроцитов в срезах сетчатки19. Другой тип остается устойчивым на протяжении всего периода затяжки (рис. 6B), напоминая токи, зарегистрированные биполярными клетками ON колбочек (ON-CBC)19.
Отдельный эксперимент был проведен для таргетирования HC, типа клеток с широким дендритным полем, которое часто трудно сохранить в препаратах срезов. Мышиная линия, экспрессирующая канальный родопсин (ChR2) и GFP в HC, была использована для облегчения идентификации под флуоресцентным микроскопом. Во-первых, токи от HC регистрировались в ответ на серию шагов деполяризации (от -100 мВ до 50 мВ, размер шага = 15 мВ), на которые они отвечали внутренними токами, за которыми следовали внешние токи (рис. 6C). Затем эти клетки стимулировали коротким импульсом синего света (200 мс, 470 нм), производящим большие внутренние токи, управляемые ChR2 в двух клетках (рис. 6D).

Рисунок 6: Записи патч-зажимов от нейронов INL в расщепленной сетчатке. (A) Предполагаемые эритроциты и (B) CBC были зажаты напряжением при -60 мВ в перфузионной среде Эймса, содержащей L-AP4 (4 мкМ). Наложение CPPG (600 мкМ) на дендриты зажатых клеток вызывало внутренний ток, который был переходным в эритроцитах, но поддерживался в ОАК. Запись RBC в (A) представляет собой одну дорожку, тогда как запись CBC в (B) представляет собой среднее значение из 3 дорожек. (C) Запись патч-зажима из HC в vGATFLPo; vGlut2Cre; Мышь Ai80d. Красная линия показывает длительность светового импульса длиной 200 мс и длиной волны 470 нм, используемого для вызова большого внутреннего тока через ChR2. (D) Отклик инжекционного тока от HC, напряжение которого было зажато на -60 мВ, затем переходило в диапазон от -70 мВ до +35 мВ с интервалом 15 мВ и возвращалось к -60 мВ. На врезке показаны те же самые следы в окне 6 мс, окружающем начало шага напряжения. (E) Иммунофлуоресцентная микрофотография плоской расщепленной сетчатки, показывающая горизонтальные клетки, экспрессирующие GFP в vGATFLPo; vGlut2Cre; Мышь Ai80d. Масштабная линейка = 20 мкм. Электрофизиологические данные собирали с частотой дискретизации 20 кГц и фильтровали с помощью фильтра Бесселя на частоте 5 кГц. Затем данные были экспортированы, а автономная визуализация и анализ были выполнены с помощью Python 3. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
Расщепление сетчатки позволяет быстро исследовать анатомию INL и OPL
Внешняя лимитирующая мембрана сетчатки (ELM) и ONL образуют барьер толщиной ~90 мкм, который препятствует диффузии антител во внутреннюю сетчатку и создает неоптимальные условия иммуноокрашивания20,21,22. Таким образом, иммуномечение мишеней в OPL или INL с использованием обычной плоской сетчатки требует трудоемких протоколов окрашивания, которые часто требуют инкубации антителв течение 48-96 часов 5,6,7,8,20,22.
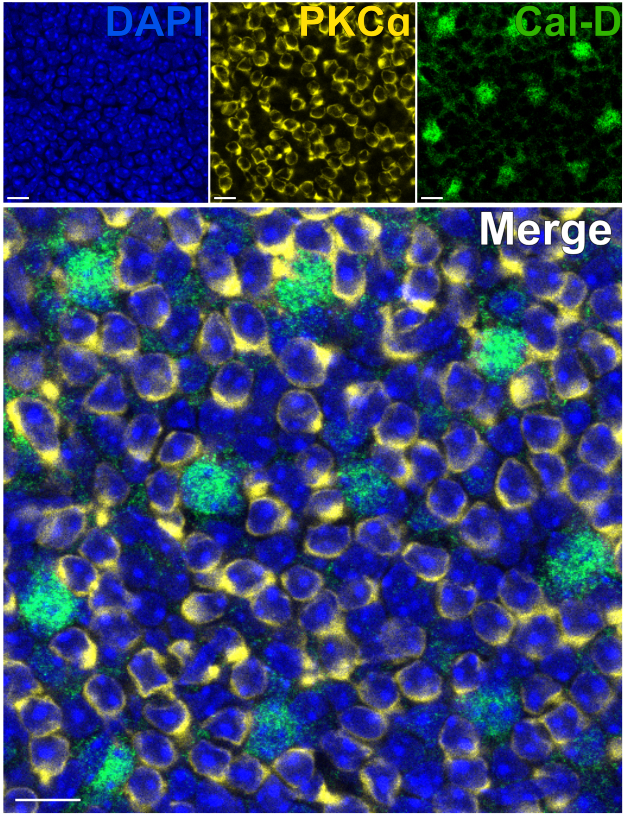
Удаление фоторецепторов позволяет быстро проникать антителами во внутренние нейроны сетчатки. В результате, мечение белков-мишеней внутри сетчатки может быть достигнуто всего за 1 ч с использованием первичных антител, конъюгированных красителем. Антитела против PKCα и Calbindin-D использовали для мечения эритроцитов и HC INL соответственно (рис. 7). В отличие от традиционных вертикальных срезов сетчатки, которые усекают латеральные отростки широкопольных нейронов, препарирование расщепленной сетчатки позволяет визуализировать полный дендритный арбор широкопольных клеток, таких как HC (рис. 6E, рис. 7).

Рисунок 7: Быстрое иммуномечение белков внутренней сетчатки в расщепленной сетчатке. Конфокальные иммунофлуоресцентные изображения расщепленной сетчатки с плоской точки зрения. Расщепленную сетчатку инкубировали с антителами против PKCα (желтый) и Calbindin-D (зеленый) в течение 1 ч при комнатной температуре для мечения ON-BCs и HCs соответственно. (A) Каждое одноканальное изображение представляет собой среднюю Z-проекцию, состоящую из четырех оптических секций: DAPI, Average z10-13; Кальбиндин-Д, средний z11-14; PKCα, Среднее z11-14. (Б) В объединенном изображении накладываются те же проекции. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок 1: Основные этапы диссекции сетчатки. Все изображения были сделаны камерой смартфона, установленной на окулярных линзах диссекционного микроскопа. (А) Изображение глаза мыши сверху вниз после удаления роговицы. (B) Изображение наглазника мыши сверху вниз после снятия объектива. (C) Делается небольшой разрез в склере на наглазнике мыши. Стрелками обозначены два лоскута склеры, которые вытягиваются щипцами в противоположных направлениях, чтобы начать отделять сетчатку от склеры. (D) После того, как склера была частично оттянута от сетчатки, между склерой и сетчаткой вставляются ножницы Ваннаса, и зрительный нерв перерезается, освобождая сетчатку. Красным пунктирным кругом показана головка зрительного нерва, а ножницы демонстрируют правильную траекторию резки (вставьте ножницы между склерой и сетчаткой). Изолированная сетчатка после склеры удаляется. Пожалуйста, нажмите здесь, чтобы скачать этот файл.
Дополнительный рисунок 2: Характеристика пре- и постсинаптических компонентов OPL в расщепленной сетчатке. Конфокальные иммунофлуоресцентные изображения с ОПЛ в расщепленной сетчатке. Расщепленную сетчатку инкубировали с антителами против GPR179 и PSD95 в течение 1 ч при комнатной температуре для мечения дендритными кончиками ON-BCs и терминалями палочковидных фоторецепторов соответственно. Левое и центральное изображения являются максимальными проекциями нескольких оптических секций; Эти же проекции накладываются на крайнее правое изображение. Видно, что GPR179 puncta на дендритных кончиках ON-BC тесно связаны с клеммами фоторецепторов палочек, демонстрируя интактные синаптические контакты внутри OPL. Масштабные линейки = 10 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 3: Поиск и устранение неисправностей: оценка качества расщепленной сетчатки. Флуоресцентные микрофотографии расщепленной сетчатки, окрашенной DAPI, для визуализации клеточных ядер. Клетки могут быть идентифицированы по диаметру и глубине ткани ядра. (А) Ядра фоторецепторов меньше, ярче и более поверхностны, тогда как (Б) Ядра БК крупнее, тусклее и глубже. (C) Изображение области с низким увеличением, в которой фоторецепторы были удалены не полностью. Ядра, которые появляются в фокусе, принадлежат BC, которые находятся глубже, чем ядра фоторецепторов по краям изображения, которые кажутся не в фокусе. Масштабные линейки для (A) и (B) = 20 мкм. Масштабная линейка для (C) = 50 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Discussion
После того, как фоторецепторы преобразуют поглощение фотонов в высвобождение нейротрансмиттеров, BC и HC являются первыми нейронами сетчатки, которые обрабатывают визуальный сигнал23. Несмотря на то, что важность этих нейронов хорошо известна, многие из их функций не до конца изучены или вообще не изучены. Во многих исследованиях физиологии РМЖ и ГК, вероятно, будет полезно использовать плоский препарат сетчатки, который улучшает доступ к нейронам INL, сохраняя при этом латеральную связь. Разработка метода расщепления сетчатки представляет собой попытку обеспечить простой протокол для получения высококачественных электрофизиологических записей и данных микроскопии от БК и ГК в плоской ориентации. Описанная здесь подготовка расщепленной сетчатки может быть выполнена примерно за 20 минут на мышь (10 минут на сетчатку) после изоляции сетчатки, без использования специализированного оборудования. Метод основан на существующих процедурах удаления фоторецепторов, но предлагает значительные улучшения в простоте, скорости и универсальности 4,10,11,12,13. В отличие от предыдущих методов разделения слоев сетчатки, расщепление сетчатки не требует замораживания, лиофилизации или многократного нанесения клея на сетчатку. С практикой почти все фоторецепторы могут быть удалены за один разрыв с помощью нитроцеллюлозной мембраны. Скорость и простота этого подхода позволяет свести к минимуму время, которое сетчатка проводит вне карбогенированного Эймса, обеспечивая высокую жизнеспособность клеток в течение длительных периодов времени; Расщепленная сетчатка может сохраняться в карбонизированной среде Эймса в течение нескольких часов после расщепления. В качестве свидетельства здоровья нейронов INL в этом препарате, окрашивание живых/мертвых клеток (рис. 4) и электрофизиология с заплатками (рис. 6) подтверждают жизнеспособность эритроцитов и ГЦ после расщепления.
Удаление фоторецепторного слоя в расщепленной сетчатке дает значительное преимущество при иммуномечении, резко сокращая время диффузии антител в INL. Маркировка первичными и вторичными антителами может быть завершена в течение 2 ч, что является существенным улучшением по сравнению с обычным плоским окрашиванием, которое может занять 72 часа или дольше в зависимости от целевого показателя 5,6,7,8,20,22. В результате данные микроскопии могут быть получены в тот же день, что и подготовка тканей, что значительно ускоряет темпы иммунофлуоресцентных экспериментов. Для облегчения отжига зондов мРНК эксперименты FISH обычно рекомендуют гораздо более длительное время фиксации (~24 ч), чем иммуномечение18. Тем не менее, эксперименты, представленные здесь, показывают, что фиксация в течение 2 ч по-прежнему дает исключительную маркировку FISH (рис. 5). Несмотря на увеличение времени фиксации с 30 мин до 2 ч, не было необходимости выполнять шаги по извлечению антигена для получения отличной иммунометки, но это может варьироваться в зависимости от антитела или антигена. Лечение протеазой в протоколе FISH может влиять на мечение антител, вероятно, из-за разрушения эпитопов-мишеней. Эту проблему удалось обойти с помощью поликлональных антител, нацеленных на несколько эпитопов, что снизило вероятность того, что разрушение эпитопов будет препятствовать иммуномечению. Кроме того, была использована умеренная терапия протеазой (ACD protease III) для предотвращения чрезмерного изменения эпитопов, обеспечивая при этом достаточное проникновение в ткани.
Иногда сетчатка вместо этого расщепляется через внешний ядерный слой (ONL), оставляя после себя слои фоторецепторных сом, в которых не видно INL-клеток. Чтобы предотвратить это, следует убедиться, что сетчатка полностью лежит на стекле, и что все остатки жидкости вокруг сетчатки удалены. Более сильное надавливание на нитроцеллюлозу кистью также может помочь предотвратить расщепление через ONL. Если мембрана становится слишком влажной или сетчатка складывается сама по себе, шансы на успешное расщепление значительно уменьшаются. Использование DAPI для окрашивания клеточных ядер полезно для оценки качества расщепления и для определения покрытия оставшихся фоторецепторов. Ядра фоторецепторов меньше, ярче и более поверхностны (дополнительный рисунок 3A), в то время как ядра BC крупнее, тусклее и глубже (дополнительный рисунок 3B). В некоторых случаях плоскость разрыва будет незначительно отличаться по части сетчатки, что приведет к образованию участков, в которых тела фоторецепторов клеток не были полностью удалены (дополнительный рисунок 3C). Для применения в микроскопии и электрофизиологии это не мешает сбору качественных данных из областей, где фоторецепторы были должным образом удалены; Большие открытые поля внутренней части сетчатки можно легко обнаружить при визуализации или записи с помощью пластырной пипетки. Если требуется более полное удаление фоторецепторов, второй разрыв может быть выполнен с помощью дополнительного куска нитроцеллюлозной мембраны, хотя 100% удаление фоторецепторов не гарантируется. Поэтому рекомендуется соблюдать осторожность при использовании расщепленной сетчатки в исследованиях экспрессии генов или протеомики, где остаточный фоторецепторный материал может повлиять на результаты. Для применения с одиночными клетками это беспокойство необоснованно, так как данные от фоторецепторов могут быть исключены из анализа.
Преимущества препарирования расщепленной сетчатки, пожалуй, наиболее заметны при электрофизиологических записях широкопольных интернейронов. В то время как традиционные вертикальные срезы прерывают обширные процессы широкопольных клеток, расщепление сетчатки оставляет OPL и IPL нетронутыми, позволяя захватывать входные данные от широкопольных клеток, таких как HCs 24, A17s25, TH ACs 26 и NOS-1 ACs27, которые в противном случае были бы упущены из виду на вертикальных срезах. Поэтому интерпретация результатов и сравнение с предыдущими данными, собранными на срезах сетчатки, требует тщательного обдумывания. Тем не менее, в экспериментах с использованием фармакологических имитаторов световой стимуляции эти результаты напоминают данные, полученные насрезах сетчатки. Экспрессируя ChR2 под клеточно-специфическими промоторами, можно стимулировать желаемую клеточную популяцию во время записи из БК в INL для изучения влияния желаемой клетки на вертикальный информационный путь. Запись непосредственно с более глубоких нейронов INL, таких как амакриновые клетки, также возможна в расщепленной сетчатке. Несмотря на то, что в этом случае пластырный электрод должен сначала пройти через более поверхностные нейроны INL, по сравнению с традиционным целым монтажным препарированием его путь преграждает значительно меньше тканей.
В дополнение к измерению влияния широкопольных клеток на другие нейроны, этот метод позволяет напрямую зажимать одноклеточные патчи от HC, дендриты которых образуют обширную щелевую связанную сеть в OPL28. Горизонтальные клетки посылают критическую обратную связь фоторецепторам, которые формируют передачу вертикальной информации через сетчатку. Однако, поскольку дендритные поля HC усечены в вертикальных срезах, данные регистрации отдельных клеток отсутствуют. В данной работе представлены анатомически и физиологически интактные ГЦ, от которых токи, вызванные ChR2, регистрируются в тройной трансгенной линии мышей (рис. 6 C-E). Вне стимуляции ChR2 расщепленная сетчатка может быть использована для изучения эндогенных токов HC и щелевой связи28. В то время как расщепленная сетчатка представляет собой удобную модель для изучения синаптических связей и активности нейронов, индуцированных химическим применением или стимуляцией ChR2, отсутствие фоторецепторов исключает любое прямое исследование естественных световых реакций или механизмов адаптации света.
Визуализация in situ на сетчатке в последние годы достигла замечательного прогресса. Тем не менее, большинство визуализирующих исследований ограничиваются слоем ганглиозных клеток в препаратах сетчатки29. Авторы предполагают, что отсутствие фоторецепторов в расщепленной сетчатке сделает ее идеальной моделью для визуализации кальция в реальном времени в OPL и INL. Помимо визуализации кальция, эта модель имеет большой потенциал для использования с генетически кодируемыми биосенсорами, такими как iGluSnFR 30,31, iGABASnFR32 и pHluorin33. В сочетании с препарированием расщепленной сетчатки эти мощные инструменты могут предложить эффективный подход к изучению синаптических взаимодействий и биофизических свойств БК и НС, которые способствуют обработке света в сетчатке.
Disclosures
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Acknowledgements
Эта работа была поддержана следующими грантами NIH: грант NIH R01EY031596 (C.M.); Грант NIH R01EY029985 (C.M.); Грант NIH P30EY010572 (C.M.); Грант NIH R01EY032564 (B.S.). Мы благодарим Тэмми Хейли за техническую поддержку в подготовке срезов сетчатки и д-ра Чарльза Аллена за щедрый вклад в зонды мРНК FISH, использованные в этой работе.
Materials
| Name | Company | Catalog Number | Comments |
| #1.5 glass coverslips | Fisherbrand | 12544E | |
| 2 pairs of Dumont #5 forceps | Ted Pella | 38125 | |
| 25 gauge needle | Becton Dickenson | 305122 | |
| 470 nm LED | THORLABS | M470L2 | |
| 5-306 curved scissors | Miltex | 5-306 | |
| 9" disposable pasteur pipetes | Fisherbrand | 13-678-20D | for constructing custom transfer pipette |
| Ai80d mouse | Jackson Laboratories | 25109 | RRID: IMSR_JAX:025109 |
| Ames Medium w/L-Glutamate | US Biological | A1372-25 | |
| amplifier control software | Molecular Devices | Clampex 10.3 software | |
| anti-calbindin D28K antibody | Invitrogen | PA-5 85669 | RRID: AB_2792808, host species = rabbit; 1:100 dilution |
| anti-CtBP2 antibody | BD Biosciences | 612044 | RRID: AB_399431, host species = mouse; 1:5000 dilution |
| anti-GPR179 antibody | NA | NA | gift from Kirill Martemyanov; Scripps Research Institute, Jupiter, FL; host species = sheep; 1:1000 dilution |
| anti-PKC alpha antibody | Sigma-Aldrich | P4334 | RRID: AB_477345, host species = rabbit; 1:5000 dilution |
| anti-PKC alpha antibody | Santa Cruz Biotechnology | sc8393 AF594 | RRID: AB_628142, host species = mouse; 1:1000 dilution |
| anti-PSD95 antibody | BD Transduction Laboratories | 610495 | RRID: AB_397862, host species = mouse; 1:1000 dilution |
| anti-RGS11 antibody | NA | NA | gift from Ted Wensel; Baylor College of Medicine, Houston, TX; host species = rabbit; between 1:1000 and 1:5000 dilution |
| anti-Synaptophysin P38 antibody | Sigma | S-S5768 | RRID: AB_477523, host species = mouse; 1:1000 dilution |
| Aquamount mounting media | Epredia | 13800 | slide mounting media |
| C57BL/6J mouse | Jackson Laboratories | 000664 | RRID: IMSR_JAX:000664 |
| carbogen tank | Matheson | NA | 95% O2 and 5% CO2 |
| custom transfer pipette | custom build | NA | Instructions: use scissors to cut off the tip of a plasitc transfer pipette at the point it begins to taper. Use pliers to safely break off the last 2-3 inches of a glass pasteur pipette. Fit the narrow end of the glass pasteur pipette into the wide tip of the plastic transfer pipete. Wrap parafilm around the joint of the two pieces to enhance the seal. |
| Digitical optical power meter | THORLABS | PM100D | |
| dissection microscope | Zeiss | Stemi 2000 | |
| electrophysiology amplifier | Molecular Devices | Axopatch 200B | |
| electrophysiology microscope | Olympus | OLYMPUS, BX50WI | Dodt gradient contrast microscopy |
| Fluoromount-G | SouthernBiotech | 0100-01 | |
| HC PL APO CS2 40x/1.3 | Leica | 506358 | |
| HC PL APO CS2 63x/1.40 | Leica | 15506350 | |
| Hybridization oven | Robbins Scientific | Model 1000 | for RNAscope protocol only |
| Immedge hydrophobic barrier pen | Vector Laboratories | H-4000 | |
| isoflurane | Piramal Critical Care | 66794-017-25 | |
| Kimwipe (delicate task wipe) | Kimtech Science | 34155 | |
| Leica HC PL APO CS2 40x/1.3 oil immersion objective | Leica | 506358 | |
| Leica HC PL APO CS2 63x/1.40 oil immersion objective | Leica | 15506350 | |
| Leica TCS SP8 X confocal microscope | Leica | discontinued | |
| medium 15 mm petri dish | Corning | 25060-60 | eyes are kept here during retina dissection |
| Merit 97-275 steel scissors | Merit | 97-275 | |
| Micropipette Puller | Sutter Instrument | p-97 | |
| Mm-Gabrd-C2 mRNA probe | ACD | 459481-C2 | |
| mouse euthanasia chamber | NA | NA | custom build; glass petri dish covering a small glass jar. |
| nitrocellulose membrane filters | GE Healthcare Life Sciences; Whatman | 7184-005 | 0.45 µm pore size |
| Picospritzer | General Valve Corporation | Picospritzer II | referred to in the text as microcellular injection unit |
| plastic transfer pipets | Fisherbrand | 13-711-7M | for constructing custom transfer pipette |
| Plastic tubing | Tygon | R-603 | for connection to carbogen tank |
| platinum harp | custom build | NA | for anchoring split retinas within the electrophysiology recording chamber. |
| size 0 paint brush | generic | NA | for flattening retina during splitting. |
| SlowFade Gold antifade reagent | Molecular Probes | S36937 | referred to in the text as anti-fade mounting media |
| small 10 mm petri dish | Falcon | 353001 | eyes are placed here following enucleation |
| small glass pane (7.5 cm x 5 cm) | generic | NA | isolatd retina pieces are placed onto this for the splitting procedure |
| Superfrost plus microscope slides | Fisherbrand | 12-550-15 | electrostatically-charged glass microscope slides |
| Thick-walled borosilicate glass pipettes with filament | Sutter Instrument | BF150-86-10HP | |
| Vannas Scissors; straight | Titan Medical | TMS121 | not brand specific; any comparable scissors will work |
| vGATFLPo mouse | Jackson Laboratories | 29591 | RRID: IMSR_JAX:029591 |
| vGlut2Cre mouse | Jackson Laboratories | 28863, 016963 | RRID: IMSR_JAX:028863, RRID: IMSR_JAX:016963 |
| Zombie NIR Fixable Viability Kit | BioLegend | 423105 | referred to in the text as MI-NIR |
References
- Morgans, C. W. Neurotransmitter release at ribbon synapses in the retina. Immunology & Cell Biology. 78 (4), 442-446 (2000).
- Euler, T., Haverkamp, S., Schubert, T., Baden, T. Retinal bipolar cells: elementary building blocks of vision. Nature Reviews Neuroscience. 15 (8), 507-519 (2014).
- Barnes, S., Grove, J. C. R., McHugh, C. F., Hirano, A. A., Brecha, N. C. Horizontal Cell Feedback to Cone Photoreceptors in Mammalian Retina: Novel Insights From the GABA-pH Hybrid Model. Frontiers in Cellular Neuroscience. 14, (2020).
- Walston, S. T., Chang, Y. C., Weiland, J. D., Chow, R. H. Method to remove photoreceptors from whole mount retina in vitro. Journal of Neurophysiology. 118 (5), 2763-2769 (2017).
- Stefanov, A., Novelli, E., Strettoi, E. Inner retinal preservation in the photoinducible I307N rhodopsin mutant mouse, a model of autosomal dominant retinitis pigmentosa. Journal of Comparative Neurology. 528 (9), 1502-1522 (2020).
- Matsuoka, R. L., Nguyen-Ba-Charvet, K. T., Parray, A., Badea, T. C., Chédotal, A., Kolodkin, A. L. Transmembrane semaphorin signaling controls laminar stratification in the mammalian retina. Nature. 470 (7333), 259-263 (2011).
- Matsuoka, R. L., et al. Guidance-Cue Control of Horizontal Cell Morphology, Lamination, and Synapse Formation in the Mammalian Outer Retina. Journal of Neuroscience. 32 (20), 6859-6868 (2012).
- Wässle, H., Puller, C., Müller, F., Haverkamp, S. Cone Contacts, Mosaics, and Territories of Bipolar Cells in the Mouse Retina. Journal of Neuroscience. 29 (1), 106-117 (2009).
- Thoreson, W. B., Dacey, D. M. Diverse Cell Types, Circuits, and Mechanisms for Color Vision in the Vertebrate Retina. Physiological Reviews. 99 (3), 1527-1573 (2019).
- Guido, M. E., et al. A simple method to obtain retinal cell preparations highly enriched in specific cell types. Suitability for lipid metabolism studies. Brain Research Protocols. 4 (2), 147-155 (1999).
- Rose, K., Walston, S. T., Chen, J. Separation of photoreceptor cell compartments in mouse retina for protein analysis. Molecular Neurodegeneration. 12 (1), 28 (2017).
- Todorova, V., et al. Retinal Layer Separation (ReLayS) method enables the molecular analysis of photoreceptor segments and cell bodies, as well as the inner retina. Scientific Reports. 12 (1), 20195 (2022).
- Shiosaka, S., Kiyama, H., Tohyama, M. A simple method for the separation of retinal sublayers from the entire retina with special reference to application for cell culture. Journal of Neuroscience Methods. 10 (3), 229-235 (1984).
- Ivanova, E., Hwang, G. S., Pan, Z. H. Characterization of transgenic mouse lines expressing Cre-recombinase in the retina. Neuroscience. 165 (1), 233-243 (2010).
- Sarria, I., Orlandi, C., McCall, M. A., Gregg, R. G., Martemyanov, K. A. Intermolecular Interaction between Anchoring Subunits Specify Subcellular Targeting and Function of RGS Proteins in Retina ON-Bipolar Neurons. The Journal of Neuroscience. 36 (10), 2915-2925 (2016).
- Orlandi, C., Cao, Y., Martemyanov, K. A. Orphan Receptor GPR179 Forms Macromolecular Complexes With Components of Metabotropic Signaling Cascade in Retina ON-Bipolar Neurons. Investigative Ophthalmology & Visual Science. 54 (10), 7153-7161 (2013).
- Dikshit, A., Zong, H., Anderson, C., Zhang, B., Ma, X. -. J. Simultaneous Visualization of RNA and Protein Expression in Tissue Using a Combined RNAscopeTM In Situ Hybridization and Immunofluorescence Protocol. Methods in Molecular Biology. 2148, 301-312 (2020).
- Wang, F., et al. RNAscope. The Journal of Molecular Diagnostics. 14 (1), 22-29 (2012).
- Morgans, C. W., et al. TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proceedings of the National Academy of Sciences of the United States of America. 106 (45), 19174-19178 (2009).
- Alessio, E., Zhang, D. Q. Immunostaining of whole-mount retinas with the CLARITY tissue clearing method. Investigative Ophthalmology & Visual Science. 61 (7), 5054 (2020).
- Ferguson, L. R., Dominguez, J. M., Balaiya, S., Grover, S., Chalam, K. V. Retinal Thickness Normative Data in Wild-Type Mice Using Customized Miniature SD-OCT. PLoS ONE. 8 (6), e67265 (2013).
- Ivanova, E., Toychiev, A. H., Yee, C. W., Sagdullaev, B. T. Optimized Protocol for Retinal Wholemount Preparation for Imaging and Immunohistochemistry. Journal of Visualized Experiments JoVE. (82), e51018 (2013).
- Kolb, H. Neurotransmitters in the Retina. Webvision: The Organization of the Retina and Visual System. , (1995).
- Chaya, T., et al. Versatile functional roles of horizontal cells in the retinal circuit. Scientific Reports. 7 (1), 5540 (2017).
- Egger, V., Diamond, J. S. A17 Amacrine Cells and Olfactory Granule Cells: Parallel Processors of Early Sensory Information. Frontiers in Cellular Neuroscience. 14, 600537 (2020).
- Dacey, D. M. The dopaminergic amacrine cell. The Journal of Comparative Neurology. 301 (3), 461-489 (1990).
- Park, S. J., et al. Connectomic analysis reveals an interneuron with an integral role in the retinal circuit for night vision. eLife. 9, 56077 (2020).
- Janssen-Bienhold, U., et al. Connexin57 is expressed in dendro-dendritic and axo-axonal gap junctions of mouse horizontal cells and its distribution is modulated by light. The Journal of Comparative Neurology. 513 (4), 363-374 (2009).
- Jain, V., et al. The functional organization of excitation and inhibition in the dendrites of mouse direction-selective ganglion cells. eLife. 9, 52949 (2020).
- Marvin, J. S., et al. Stability, affinity, and chromatic variants of the glutamate sensor iGluSnFR. Nature Methods. 15 (11), 936-939 (2018).
- Strauss, S., et al. Center-surround interactions underlie bipolar cell motion sensitivity in the mouse retina. Nature Communications. 13 (1), 5574 (2022).
- Marvin, J. S., et al. A genetically encoded fluorescent sensor for in vivo imaging of GABA. Nature Methods. 16 (8), 763-770 (2019).
- Beckwith-Cohen, B., Holzhausen, L. C., Wang, T. M., Rajappa, R., Kramer, R. H. Localizing Proton-Mediated Inhibitory Feedback at the Retinal Horizontal Cell-Cone Synapse with Genetically-Encoded pH Probes. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 39 (4), 651-662 (2019).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved