
Method Article
B Lenfosit Aktivasyonunu Incelemek Için Çekiş Kuvveti Mikroskobu
Bu Makalede
Özet
Burada B hücreleri üzerinde çekiş kuvveti mikroskobu deneyleri yapmak için kullanılan bir protokol salıyoruz. Yumuşak poliakrilamid jellerin hazırlanmasını ve işlevselleştirilmesinin yanı sıra mikroskopta veri toplama ve veri analizinin bir özetini anlatıyoruz.
Özet
Çekiş kuvveti mikroskobu (TFM), bir hücretarafından bir alt tabaka üzerinde üretilen kuvvetlerin ölçülmesini sağlar. Bu teknik, elastik bir substrat üzerinde çeken bir hücre tarafından üretilen deneysel olarak gözlemlenen bir yer değiştirme alanından çekiş kuvveti ölçümleri çıkartır. Burada, B hücre reseptörünün antijen etkileşimi ile aktive edildiğinde B hücreleri tarafından uygulanan kuvvet alanının mekansal ve zamansal yapısını araştırmak için TFM'yi uyarladık. Jel sertliği, boncuk yoğunluğu ve protein fonksiyonelizasyonu, hücre yüzey reseptörleri için özellikle ligandlara etkileşen ve özellikle yanıt veren nispeten küçük hücrelerin (~ 6 μm) incelenmesi için optimize edilmelidir.
Giriş
B hücreleri bağışıklık sisteminin antikor üreten hücreleridir. Adaptif immün yanıtı etkinleştirmek için, ilk olarak yerel bir biçimde antijen elde (yani, işlenmemiş) belirli bir reseptör aracılığıyla B hücre reseptörü (BCR)1. Bu süreç lenf nodül B hücre bölgesinde oluşur. Bazı antijenler lenfatik sıvılar yoluyla B hücresine ulaşabilse bile, çoğu antijen, özellikle yüksek molekül ağırlığına sahip (>70 kDa, lenfatik kanallar için sınır boyutudur) gerçekten de bir antijen sunucu hücre (APC) yüzeyinde kendi yerli formunda sunulur, genellikle bir subkapsüler sinüs makrofaj veya foliküler dendritik hücre, lektin veya Fcs (non-spesifik reseptör) ile. Bu hücre ile temas BCR APC ilişkili antijenler üzerinde kuvvet uygular bir bağışıklık sinaps oluşumuna yol açar. Bir antigenin BCR'ye bağlanması BCR sinyalini başlatır ve bu da kuvvet üreten mekanizmaları harekete geçirir. Bu kuvvetler BCR sinyalini yükseltmek için önemli olabilir, ama aynı zamanda B hücrelerinin antijeni ayıklayıp içselleştirmesi için de gereklidir.
Son çalışmalar BCR gerçekten mechanosensitive olduğunu göstermiştir2. Örneğin, daha sert yüzeyler gelişmiş BCRsinyal3ortaya çıkarır. Ayrıca, bağışıklık sinaps üretilen kuvvet antijen e afinitesini araştırmak ve böylelikle yakınlık ayrımcılıksağlamakiçin tek BCR çeker 4 . Bu nedenle, B hücrelerinin antijen sunumuna mekanik yanıtını araştırmak ve bu yanıtı, karışan reseptörlerin türü (IgG/IgM)5,yap- izyon molekülleri (integrin ligandlar) veya farmakolojik ve genetiği değiştirilmiş hücrelerde (yani BCR sinyalizasyonuveya sitoiskeletinin protein aşağı akımının susturulması) açısından incelemek ilginçtir 6 .
Bir hücrenin fizyolojik sertlik alt tabakasına tepkisini gözlemlemek için kullanılan basit bir yöntem ve aynı zamanda substrat üzerinde uygulanan çalışma kuvvetleri Çekiş Kuvveti Mikroskopisi (TFM). TFM, elastik bir substrat üzerinde çekerek hücre tarafından üretilen deplasman alanını gözlemlemekten oluşur. Başlangıçta jel deformasyonfaz-kontrastmikroskobu 7 tarafından elastomer kendisi kırışıklıkları ile gözlendi , ancak daha iyi çözünürlük için izin ve o zamandan beri standart8haline gelmiştir fidusikal belirteçler olarak floresan mikroboncukların eklenmesi . Bu yöntem yapışık hücreler, dokular ve hatta jellere gömülü organoidler tarafından uygulanan çekiş kuvvetini araştırmak için kullanılmıştır. TFM çeşitli varyasyonları dahil olmak üzere9 geliştirilmiştir, süper çözünürlüklü mikroskopi ile kombinasyon (yani, STED10 veya SRRF11), TIRF mikroskopisi için izin jel kırılma indeksi modifikasyonu12, nano baskılı desenler ilerle boncuk yerine13, ve düz yüzey yerine nanopillar kullanarak14. Bu varyasyonların tam bir incelemesi için Colin-York ve ark.15'ebakın.
Burada sunulan protokol, B hücrelerinin antijen kaplı bir alt tabaka üzerinde uyguladığı kuvvetleri ölçme prosedürünü tanımlar. Bu kuvvetler ligandlar (antijen) onları kümelemek ve daha sonra antijen sunan substrat onları ayıklamak için uygulanır. Standart TFM protokolünü fizyolojik antijen sunan yüzeylerin sertliğini, B hücrelerinin boyutunu ve ilgili kaplamasını taklit etmek için uyarladık. Bu protokol aynı anda birden fazla hücrenin incelenmesine olanak sağlar ve floresan mikroskopi teknikleri ve kimyasal tedaviler ile birlikte kullanılabilir. Ancak, optik cımbız16,moleküler gerilim probları17,18,biyomembran kuvvet probları19ve atomik kuvvet mikroskobu20 daha uygun teknikler olan tek molekül kuvvet ölçümleri, sonda amaçlamaz. Diğer tek hücre kuvveti ölçüm yöntemleriyle karşılaştırıldığında (örneğin, mikropipetler21 veyamikroplakalar 22)TFM, ~300 nm çözünürlüğe sahip sinapsta uygulanan kuvvetlerin tam bir haritasının yeniden yapılandırılmasına olanak sağlar. Bu, yüzeyde uygulanan kuvvetlerdeki spatio-temporal desenleri tanımlamak ve jel konfokal görüntüleme ile uyumlu olduğu için, bunları spesifik proteinlerin (örneğin, sitoiskelet ve sinyal proteinleri) işe alınmasıyla ilişkilendirmek için yararlıdır.
3D TFM mümkün olsa da, kullandığımız sertlik ve kurulum ile uyumlu değildir. 3D deformasyonlar çıkıntı kuvveti mikroskopisi gibi diğer daha karmaşık kurulumları ile ulaşılabilir (AFM hücrelerin plakalı bir deforme membran tarama)23,24 ve elastik rezonatör girişim stres mikroskopisi (ERISM, ışık için rezonans boşluğu olarak hareket eden bir jel ve birkaç nanometre doğruluğu ile substrat deformasyonları vurgulayarak)25. Bu teknikler çok umut verici olmasına rağmen, henüz B hücrelerinde kullanılmamıştır. Nanopillar14gibi Diğer TFM türleri, daha fazla tekrarlanabilir yüzeylere sahip olmak için kullanılabilir. Ancak, hücre sütunları içine nüfuz ettiği için bu geometri yumuşak hücrelere adapte edilmez ve bu da analizi zorlaştırır. Bu yaklaşım gerçekten t hücrelerinde hücrenin26sütunları etrafında yapılar inşa etme yeteneğini gözlemlemek için kullanılmıştır.
Sadeliği rağmen, Poliakrilamid jelleri kullanarak TFM birçok hücrenin eşzamanlı gözlem sağlar ve kolay ve ucuz bir tezgah ve epifloresan mikroskop ile donatılmış herhangi bir laboratuvarda uygulanabilir (biz konfokal / iplik disk tavsiye rağmen).
Bir APC fizyolojik sertlik taklit etmek için, ~ 500 Pa27 sertlik ile poliakrilamid jeller kullanılan ve antijenleri aktive ile jel fonksiyonel. Bu protokolde poliakrilamid jelin yüzeyini tavuk yumurtası lysozyme (HEL) ile işlevsel hale getirilmiştir. Bu antijen bağlama alanının angajman yoluyla BCR uyarılması tarafından oluşturulan kuvvetlerin ölçümü için izin verir. Bu antijenin ve MD4 farelerinin HEL spesifik B hücrelerinin kullanımı, antijen ligasyonuna yanıt olarak nispeten düzgün kuvvet oluşumunu sağlar28. Ancak, diğer moleküller (B6 fareler için anti-IgM gibi) jel üzerine aşılanmış olabilir, ancak bu durumlarda üretilen kuvvetler daha heterojen ve daha az yoğun olabilir. B hücreleri küçük hücreler (çap ~6 μm) olduğundan, boncuk sayısı maksimal ama yine de izlenebilir olarak optimize edilmiştir. Kendi substratları üzerinde ~ kPa kuvvetleri uygulamak büyük hücreler için, bir nispeten seyrek boncuk lar kullanarak tatmin edici sonuçlar elde edebilirsiniz veya basit parçacık görüntü velocimetry gerçekleştirerek (PIV) deformasyon alanı yeniden. Ancak, ~50 Pa kadar küçük stres uygulayan B lenfositler gibi küçük hücreler için, deformasyon alanını yeniden oluştururken istenilen doğruluğa ulaşmak için tek partikül izleme (parçacık izleme velocimetry, PTV) kullanımı gereklidir. Boncukları tek tek güvenilir bir şekilde takip edebilmek için objektif lensin büyütülmesi en az 60x ve 1.3 civarındaki sayısal diyafram açıklığı olmalıdır. Bu nedenle, jeller nispeten ince olmalıdır (<50 μm), aksi takdirde boncuklar hedefin çalışma mesafesi üzerinde olduğu gibi görünür değildir.
Ana protokol üç bölümden oluşur: jel hazırlama, jel fonksiyonelleştirme ve görüntüleme; iki bölüm daha isteğe bağlıdır ve floresan hücrelerin antijen ekstraksiyon nicelleştirme ve görüntüleme adamıştır.
Protokol
1. Jel hazırlama
- Jel desteğinin silanizasyonu
- 2 dakika uv lambaile coverslip veya cam dip petri kabını (jel desteği olarak kullanılacak) etkinleştirin (artık ozona maruz kalmamak için UV lambasına maruz kalmadan önce 30 s bekleyin).
- 5 dakika boyunca 200 μL aminopropyltrimethoxysilane (APTMS) kullanarak coverslip/cam alt çanak silanize. Bu jel kovalent bağlanması için destek hazırlayacaktır.
- Kapak/cam alt kabını ultra saf suyla iyice yıkayın.
- Vakum aspirasyon kullanarak kapak/cam alt çanak kuru.
- Jeli düzleştirmek için kullanılan 18mm coverslip'in hazırlanması
- Kapakları hazırlamak için önce seramik kapak tutucuya koyun. Daha sonra kapak tutucuyu küçük bir kabın içine (50 mL) koyun ve tamamen üzerini örttüldüğünde emin olmak için, kapaklar üzerinde silikonlaştırıcı reaktif (4 °C'de depolanır, yeniden kullanılabilir) dökün.
- Kabı alüminyum folyo ile kaplayın ve oda sıcaklığında 3 dk kuluçkaya yatırın. Beklerken, büyük bir kabı (500 mL) ultra saf suyla doldurun. Silikonlu reaktifte 3 dk kuluçkadan sonra, kapaklı kapak tutucuyu su kabına iletin.
- Kapakları ultra saf suyla iyice durulayın, iyice kurutun ve kağıt mendillerde saklayın. En iyi sonuçlar için hemen bir sonraki bölüme gidin.
- Jel polimerizasyonu
- 0,5 kPa'lık jeller için 75 μL ve %40 akrilamid %30 μL 2 bisacrylamid (crosslinker) ve 895 μL fosfat tamponlu salin (PBS) ile karıştırın. Bu premix 4 °C'de bir aya kadar saklanabilir.
- 0.5 kPa jel premix 167 μL için, bir banyo sonicator (50-100 W ve frekans 40 kHz gücü ile standart tezgah ultrasonik temizleyici) 5 dakika için boncuk, girdap ve sonicate% 1 (1,67 μL) ekleyin. Karışımı alüminyum folyo kullanarak ışıktan koruyun.
NOT: Başlatıcı (TEMED) eklenene kadar premix polimerize olmaz. - Polimerizasyonu katalize etmek için % 1 (1,67 μL) w/v amonyum persülfat (APS) ekleyin.
- Polimerizasyonu başlatmak için% 0.1 (0.2 μL) N,N,N′, N′-Tetrametiletiletilediamin (TEMED) ekleyin. Bir pipet ile karıştırın. APS ve TEMED eklendikten sonra, jel hızla polimerize sayılsa da jel döküme hızla devam edin.
- Jel döküm
- Pipet 9 μL jel karışımı her coverslip/cam alt çanak üzerine (merkezde damla, Şekil 1A)
- Silanized / hidrofobik kapak ve jel düzleştirmek yerleştirin(Şekil 1B). Forseps kullanarak, jelin dışarı sızmaya başlayana kadar kapağın tüm alanına(Şekil 1C)yayıldığını sağlamak için kapak kaymasına basın.
- Coverslip/cam alt kabı büyük bir Petri kabına ters çevirin ve jel yüzeyine doğru giden boncukları zorlamak için tezgaha dokunun(Şekil 1D).
- Alüminyum folyo ile kaplayın ve oda sıcaklığında nemli bir odada polimerize 1 saat bekletin (yani, buharlaşmayı önlemek için yemeğin üzerine ıslak bir doku koyun).
- 1 saat sonra, coverslip salınımını kolaylaştırmak için örneğe PBS ekleyin. Dikkatlice, bir iğne kullanarak coverslip çıkarın (farklı silanes ile kaplama jel, Şekil 1Ecoverslip kolay soyma izin vermelidir).
- Jeli PBS'de bırak.
NOT: Jeller artık 4 °C'de 5-7 gün süreyle PBS'de saklanabilir, ancak 48 saat içinde kullanılması tavsiye edilir.
2. Jel fonksiyonelleştirme
- 10 mM HEPES tamponunda 0.5 mg/mL'de sulfosuccinimidyl 6-(4'-azido-2'-nitrofenilamino)hekzanoat (Sulfo SANPAH) çözeltisi hazırlayın. Bu 4 °C'de alüminyum folyo ile bir haftaya kadar saklanabilir.
- PBS'yi jellerden aspire edin.
- Oda sıcaklığında jele 150 μL Sulfo SANPAH ekleyin(Şekil 1F).
- Sulfo SANPAH sitelerini fotoaktiletmek ve jel yüzeyine yapışmasını sağlamak için jeli UV tedavisine 2 dakika boyunca maruz bırakın.
- PBS ile üç kez yıkayın (Şekil 1G).
- Adımları 2.2-2.5'i yineleyin.
- Her jele 250 μL (100 μg/mL) ekleyin ve bir gecede 4 °C'de nemli bir odada kuluçkaya yatırın ve alüminyum folyo ile kaplanmış olarak bekletin(Şekil 1H).
- HEL antijenini çıkarın ve PBS ile üç kez yıkayın.
NOT: HEL hem antijen hem de yapışma molekülü olarak hareket eder. Reseptöre bağlanan diğer moleküller (örneğin, fare karşıtı IgM, Sığır Serum Albumin, Ovalbumin) veya integrin ligandlarla (örneğin, LFA1'e bağlanan ICAM1) ile karıştırılabilir. Gerekirse, antijen ekstraksiyon HEL floresan versiyonu ile görülebilir (bir protein etiketleme kiti ile molekül boyama elde, adım 4 bakınız). Toplu olarak belirli bir konsantrasyonun jel üzerinde camdakiyle aynı yüzey konsantrasyonunu ortaya çıkaramayabileceğini unutmayın: doğrudan karşılaştırma gerekiyorsa bunun ikincil boyama ile ölçülmesi gerekir.
3. Hücre yükleme ve görüntüleme
- Görüntülemeden önce, PBS'yi jellerden çıkarın ve 500 μL B hücreli ortam ekleyin (RPMI 1640, %10 dekomplemasyonlu fetal baldır serumu, %1 penisilin-streptomisin, %2 Sodyum Pirüuvat, 50uM Mercaptoetanol ve 1X Non Essential Amino Asitler) ekleyin ve RT'ye dengede vermelerini sağlar.
- Hücre hazırlama
- Birincil B hücrelerini negatif seçim protokolüne göre dalaktan arındırın (bkz. Malzemeler Tablosu). Tipik son B hücre verimi 1 x 107 hücre civarındadır. Bunu B hücreli ortamda 3 x 106 hücre/mL'ye yoğunlaştırın (RPMI-1640 %10 fetal buzağı serumu, %1 penisilin-streptomisin, %0.1 merkaptoetanol ve %2 sodyum piruvat ile desteklenmiştir).
- 4 °C'de 6 saate kadar hücreleri gerektiği gibi saklayın.
- Görüntü edinilmeden önce hücreleri 30 dakika boyunca 37 °C'de tutun.
- Görüntüleme
- Termal ve (muhtemelen) CO2 kontrolü ile bir konfokal mikroskop kullanın.
NOT: Konfokal veya dönen disk mikroskobu kullanılsa kullanılsın, analiz aşamasındaki boncukları rahatça izlemek için piksel boyutu <200 nm sağlayan nesnel/iğne deliği kullanmak önemlidir (örn. 60x, NA 1.3). Epifloresan mikroskopi de kullanılabilir, ancak gürültü oranı daha düşük sinyal sağlar ve bireysel boncuk izleme zor yapabilir. - Boncuk iki ana tabaka alt ve jel üst görünecektir. Jel düzlemine odaklan.
NOT: Güzel bir jel yıldızlı bir gökyüzü olarak görünür, boncuklar yaklaşık aynı düzlemde eşit olarak dağıtılan ile. - 5 s kare hızı ile 30 dk için satın alma programı (bu deney ihtiyaçlarına uyarlanabilir, örneğin, diğer renkler elde, z yığını, vb elde)
- Jelden ortam aspire, jel üzerinde yaklaşık 200 μL medya bırakarak. Mikroskop üzerinde jel konumlandırın ve boncuk yüzey tabakası ve jel üzerinde güzel bir çift alan bulmak.
- 80 μL hücre ekleyin (odaklanmayı korumak için jele dokunmaktan kaçının).
- Odak hala doğru olduğundan emin olun ve hücrelerin bölgede azalan görülebilir (iletilen ışık altında). Hücreler jel ulaşmadan önce edinimi başlatın.
- Jel, titreşimler veya odak kayması ile kazara temas durumunda, odağı ayarlayın.
NOT: Bu rahat jel bir görüntü toplamak için çok önemlidir ve bu jel üzerinde hücrelerin gelmeden önce alınan herhangi bir görüntü olabilir.
- Termal ve (muhtemelen) CO2 kontrolü ile bir konfokal mikroskop kullanın.
4. Floresan HEL ekstraksiyon deneyi
- Floresan HEL'yi bir floresan boyayı (Alexa 555 gibi boncuklardan farklı bir renkten) bağlayarak hazırlayın, Malzeme Tablosu'nabakın.
- Adım 2.7, floresan HEL ile konvansiyonel HEL değiştirin.
- Fotoğraf beyazlatma önlemek için düşük aydınlatma ayarları na veya düşük kare hızına (örneğin, dakikada 2 kare) görüntüler edinin.
- HEL ekstraksiyonu ölçmek için, formüle göre 0 çerçevenin yoğunluğu I(0)tarafından düzeltilen ve normalleştirilen her kare için hücre alanı üzerinde entegre edilen yoğunluğu hesapla:

NOT: Florofor ile konjuge antijen görünür değildir (muhtemelen jel yüzeyinde florofor söndürme nedeniyle), ancak jel üzerindeki varlığı bir anti-HEL ve floresan ikincil antikor ile doğrulanabilir. Bu florofor anti-HEL ile kaplı bir coverslip ile jel sıyırma ve ikincil bir floresan antikor ile ortaya (coverslip)6ile ayrılarak zaman florofan olduğunu doğrulanabilir . Çıkarılan antijenin sinyali çok sönüktür ve bazen boncukların sızdırılmasıyla maskelenir. Bir antijen ekstraksiyon sadece ilgileniyorsa, boncuk olmadan jel hazırlamak için tavsiye edilir (adım 1.3.2 ve 1.4.3 atlamak).
5. Floresan görüntüleme
- Yabani tip için yapılan (örneğin, Lifeact-GFP veya Myosin II GFP farelerinden) genetiği değiştirilmiş farelerin dalaklarından B hücresini arındırarak floresan B hücreleri elde edin.
- Floresan hücrelerin görüntülenmesi için, (mümkünse) bir su daldırma uzun mesafe 40x-100x amacı ile dönen bir disk mikroskobu kullanın.
- Beyazlatma önlemek için pozlama süresi ve kare hızı düşük tutun.
NOT: Z'deki nokta yayma fonksiyonu jelin varlığı ndan oldukça bozulmuştur, bu nedenle su daldırma amacı nı kullanmanızı öneririz. Su daldırma hedefleri ile canlı dik mikroskopi emisyon yolunda (küresel) hücre (ve hücre çekirdeği) varlığı ile indüklenen güçlü küresel sapmalar muzdarip.
6. Analiz
NOT: Veri analizi genel olarak sürüklenme için tüm yığını düzelterek, her karedeki boncukları bularak, bir referans çerçevesine göre hareketlerini izleyerek (hücrelerin yokluğunda alınan), yer değiştirme alanını enterpolasyona ederek ve Fourier transform29kullanarak stresi elde etmek için sorunu tersine çevirerek gerçekleştirilir. Bu amaçla, imagej makro ve MATLAB programlarının bir kombinasyonunu kullanarak çevrimiçi bir depodan indirilebilir30öneririz.
- Filmi ImageJ'de görüntü yığını olarak açın
- Makro "Crop_and_save.ijm" çalıştırın
- "Dikdörtgen" aracıyla ilgi çeken bölgeleri (YG) seçin ve 't' tuşunu kullanarak YG listesine ekleyin.
- Hücreyi kırpırken, en az 5-10 piksel hareketsiz boncuk içeren bir bölge eklediğinizden emin olun. Çözümlemeden sınırlara veya diğer hücrelere çok yakın olan hücreleri hariç tinaltın. Bittiğinde 'Tamam'a tıklayın.
- Makro hücrenin bir maskesi önerir: Bu tatmin edici ise "Tamam"ı tıklatın. Tatmin edici değilse, "Tamam Değil"e tıklayın ve ardından herhangi bir seçim aracıyla (örn. "Freehand" veya "Oval") kapalı bir bölgeyi el ile seçin ve "Devam Et"e tıklayın.
- MATLAB'ı açın ve "TFM_v1.m" çalıştırın.
- Gerekli parametreleri gir: özellikle görüntü özelliklerini (piksel boyutu, edinme zaman aralığı) ve jel özelliklerini (Genç modül E, Poisson oranı) kontrol edin.
- Başvuru görüntüsü varsayılan olarak ilk olarak ayarlanır. Gerekirse başka bir çerçeveye ayarlayın veya harici bir dosyayı yüklemek için "0" olarak ayarlayın.
- Yazılımın çıktılarını özgün dosyayla aynı dizinde bulun (açıklama için User_notice.pdf dosyaya bakın). Bu boncukbir ön parça içerir ("FILENAME.fig"), zaman içinde kontraktil enerji bir arsa ("FILENAME_energy.fig"), hücre (enerji, alan, anlar, vb) "FILENAME_finaltable.mat", deplasman ve kuvvet alanı içeren bir yapı, boncuk filmleri içeren bir yapı, deplasman alanı, stres ve enerji (herhangi bir avi okuyucu ile açılabilir).
NOT: Giriş parametrelerinde "Pencere boyutu" yer değiştirmenin enterpolasyona tabi olduğu penceredir, dolayısıyla gerilim ve yer değiştirme alanının nihai çözünürlüğüdür. Bu, birkaç (varsayılan olarak dört) piksel olarak ayarlanır. Boncuk olmayan bölgeleri enterpolasyona ederek çözünürlüğü yapay olarak artıracağı için bunu azaltmak tavsiye edilmez.
Sonuçlar
Hücrelerin büyüklüğü göz önüne alındığında, korenik teknikleri (parçacık görüntü velocimetry gibi) ile boncuk deplasman haritası ayıklamak algoritmaları genel olarak çok kesin değildir. Ancak gerekli çözünürlük derecesine bağlı olarak, bir kolayca ücretsiz bir Fiji / ImageJ eklentisi 31 ,32kullanarak nitel sonuçlar elde edebilirsiniz. Bu yaklaşım uyarıcı karşı uyarıcı koşulları karşılaştırmak için yeterli olmakla birlikte, ayrıntılı bir analiz için biz bir online depo dan indirilebilir yazılımımızı kullanmanızı öneririz30, tek tek boncuk izler ve bireysel boncuk deplasmanları interpolasyonu olarak belirli bir zaman noktasında deplasman alan haritası sağlar33. Bu noktada çeşitli nicelikler mümkündür. Örneğin (deplasman sadece jel yüzeyine stres teğet neden olduğunu varsayarak) yazılım da belirli bir deplasman haritası neden her noktada stres sağlar. Bu bir tür "ters çevirme sorunu"dur: belirli bir noktadaki yer değiştirme, diğer noktalara uygulanan tüm kuvvetlerin toplamına bağlıdır. "Inversiyon algoritması" substrat fiziksel parametreleri dikkate alır: sertlik (Genç modül) ve Poisson oranı. Doğrudan algoritmalar genellikle çok doğru ama hesaplama pahalıdır. Bizimki gibi Fourier dönüşümüne dayanan algoritmalar, fourier alanında temelde bir dekonvolution gerçekleştirir ve daha verimlidir ancak bazı hatalara yatkındır (esas olarak enterpolasyon adımı nedeniyle). Bu algoritmalar genellikle küçük yerel (ve potansiyel olarak artifactual) yer değiştirmeleri vurgulayan bir parametrenin tonuyla gerilim alanının hesaplanmasında çok alakalı olmasını gerektirir (Tikhonov düzenlileştirme parametresi8,29; İletişim penceresinde "Düzenlileştirme" değişkeni; burada genellikle 5 x 10-19'aeşit olarak ayarlanırız). Daha gelişmiş yorumlama ve analiz için, örneğin spatio-temporal korelasyonlar, lokal hareketler, floresan kanallarla korelasyonlar, bu alandaki uzmanlarla işbirliği yapmanızı öneririz. Hesaplama yöntemleri üzerinde bir inceleme için Schwarz ve ark.9bakın.
Yukarıda belirtildiği gibi, doğru boncuk görüntüleri bir "yıldızlı gökyüzü", parlak noktalar bir üniforma ve rasgele dağılımı gibi görünüyor(Şekil 2A). Boncuk sayısı çok düşükolduğunda(Şekil 2B)veya görüntü odak dışında olduğunda veri ve analiz güvenilir değildir (Şekil 2C). B hücreleri jel yüzeyine yerleştikten sonra, hücrelerin altındaki boncuklar jel üzerinde hücre tarafından uygulanan çekiş kuvveti nedeniyle hareket etmeye başlar. Boncukların izlenilememesi gereken çerçeveler atılmalıdır.
Bir kontrol olarak, genellikle substrat ile hücrenin ilk temas öncesinde bir "referans çerçevesi" karşılaştırarak boncuk hareketini göz ile gözlemlemek mümkündür. Şekil 3A'dayapılan tek parçacık takibinden (örneğin, Trackmate, Fiji 34)yaklaşık sonuçlar elde edilebilir. Analiz, referans görüntüsündeki boncukların ("FILENAME.fig") denetim olarak bölümlemesi sağlar.
Önerdiğimiz yazılımla, deplasman(Şekil 3B)ve gerilim alanı (her pikseldeki yerel gerilimin vektörü ve deplasman alanından ters çevrilme ile elde edilen her zaman noktası, Şekil 3C)elde edilebilir. Hücre nin alanına entegre edilmiş yer değiştirme ve kuvvet alanlarının skaler ürünü, hücrenin substrat üzerinde uyguladığı toplam çalışmayı sağlar(Şekil 4A). Bu hesaplama, protokolün 6.2.
İki biyolojik durumu (HEL'yi aktive edici olmayan substrat BSA'ya karşı veya vahşi tipe karşı nakavt) karşılaştırmak için ortalama eğriyi(Şekil 4B)veya daha da sentetik olarak, enerjinin bir platoya ulaştığı son zaman noktaları (20 dk) üzerindeki ortalama değeri hesaplamak yararlıdır(Şekil 4C). Kuvvetlerin mekansal bilgileri ilgili olduğunda her koşulun tek zaman noktalarını karşılaştırmak mümkündür(Şekil 4D). Daha derin analiz için Kumari ve ark.6'ya bakın.
Floresan antijen ekstraksiyon zaman atlamalı bir örnek Şekil 5Agösterilmektedir: sinaps floresan sinyalleri ilerleyici görünüm jel antijen ayrılması gösterilir. 15 hücrenin üzerindeki güven aralığı (ortalamanın standart hatası) ile ortalama çıkarma eğrisi Şekil 5B'degösterilmiştir.

Şekil 1: Jelin hazırlanmasının ve işlevselleştirilmesinin şematik gösterimi. Adımlar protokolde açıklanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: Farklı niteliklere sahip boncuk görüntülerinin üç örneği. (A) Gürültü oranına ve doğru yoğunlukta doğru sinyale sahip boncuk görüntüsü örneği. (B) Çok sayıda boncuk ve(C)odak düzleminden çıkan görüntülere örnekler. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Kuvvet alanını ayıklamak için görüntülerin işlenmesi. (A) Boncukların görüntüsüne örnek olarak (beyaz hücrenin anahat, iletim görüntüsünden çıkarılan), boncuk izleme zamanında t = 5 dk (kırmızı bindirme) ve yer değiştirme (oklar) zamana göre t = 0 dk (ölçek çubuğu 5 μm). (B) İnterpolated deplasman alanı (vektör titreme ve büyüklük haritası olarak temsil edilir, oklar deplasman [nm] ile orantılıdır; sağdaki renk çubuğuna bakın); alt: büyüklüğü daha yumuşak bir görüntü (bikübik bir fonksiyon ile enterpolasyon ile elde). (C) B panelindeki yer değiştirme alanından gelen gerilim alanı (vektör titremesi ve büyüklük haritası olarak gösterilir; oklar kesme gerilimi [Pa] ile orantılıdır; sağdaki renk çubuğuna bakın); alt: büyüklüğü daha yumuşak bir görüntü (bikübik bir fonksiyon ile enterpolasyon ile elde). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
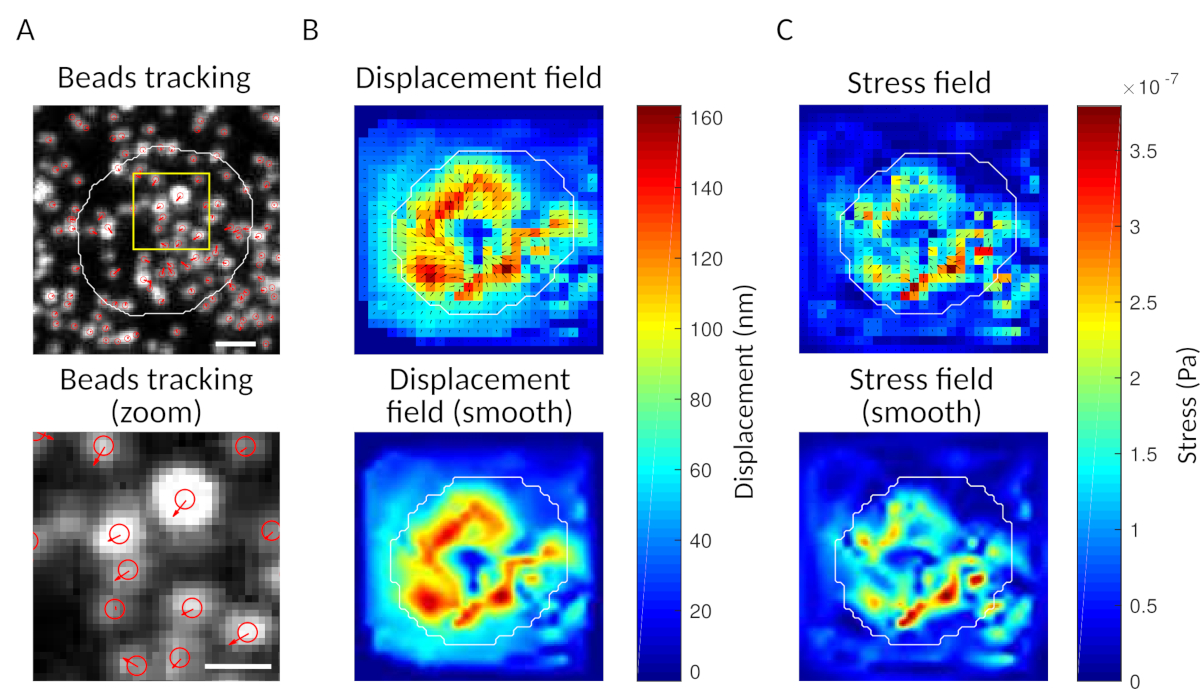{kind=link}

Şekil 4: Kuvvet ve yer değiştirme alanlarından çıkarılabilen bilgi örneği. (A) Tek bir hücre için enerjinin zamanında evrimine örnek: Bir plato fazı (gri renkle vurgulanır) yaklaşık 10 dakika sonra ortaya çıkar. (B) Hel (aktive edici) kaplamalı jel ve BSA (aktive olmayan) kaplı jel üzerinde 35 hücre (ortanca ± interquartile aralıkları gösterilmiştir, Mann-Whitney testi istatistiksel anlamlılık için kullanılmıştır) kaplı 65 hücre için göreceli plato düzeylerinin ortalama enerji eğrileri ve(C)karşılaştırılması. (D) HEL ve kontrol BSA durumu için stres hızlandırılmış renk haritaları; hem büyüklük hem de titreme çizimleri gösterilir. Bu görüntüler Kumari ve ark.6'danuyarlanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
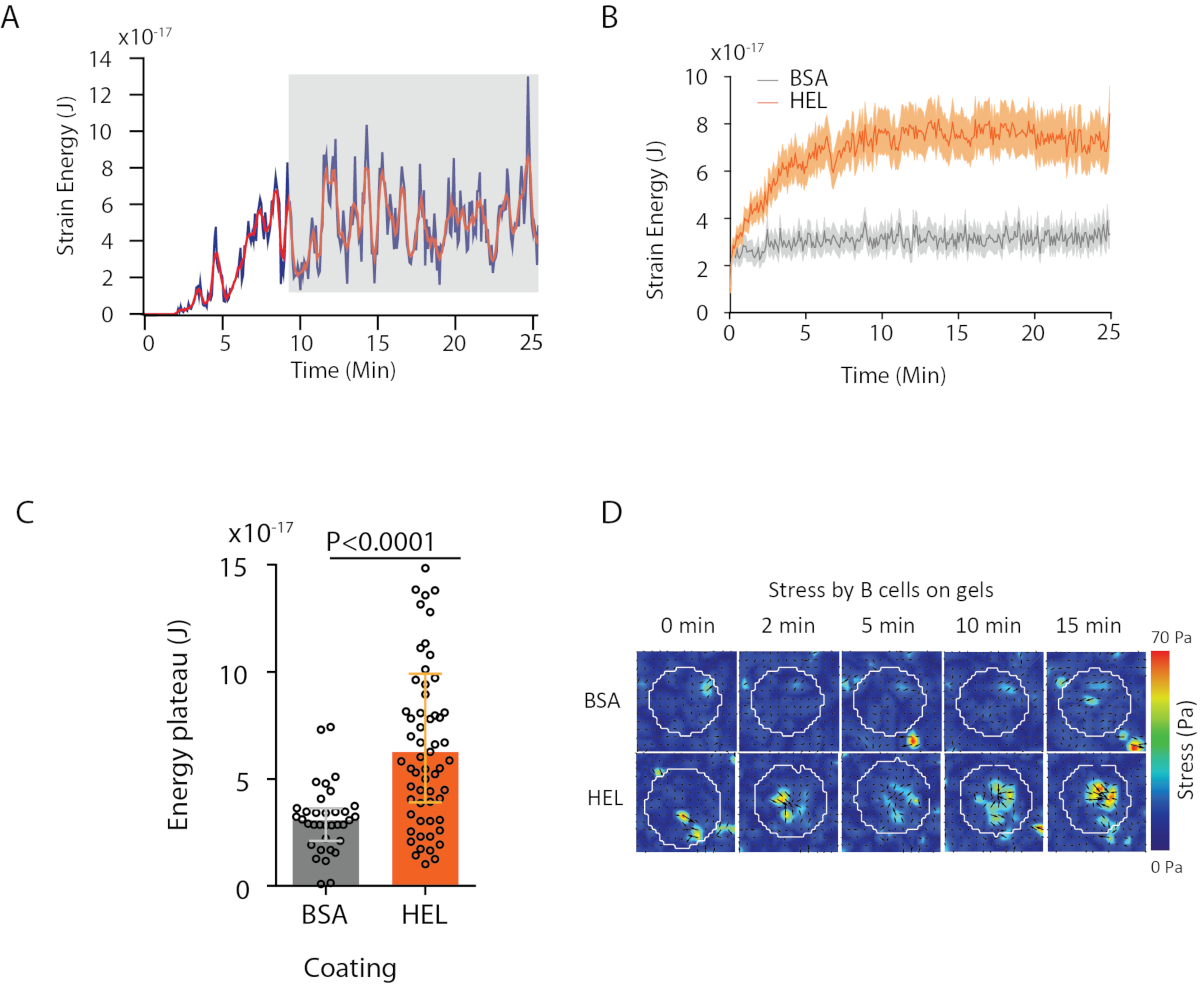{kind=link}

Şekil 5: Floresan antijenle yapılan deneylere örnek. (A) Floresan HEL çıkarma nın zaman atlamalı (aşağıda: maksimum yüzde, ölçek çubuğu = 3μm). (B) Zaman içinde antijen toplanması (Ortalama ± SEM, n = 15). Bu görüntüler Kumari ve ark.6'danuyarlanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Burada açıklanan TFM yöntemi, B hücrelerinin aktif mekanik yeteneklerinin sistematik olarak incelenmesine olanak sağlar. B hücreleri bağlamında, bu antijenayık ve içselleştirmek yeteneği ile ilgilidir. Diğer TFM yöntemleri ile karşılaştırıldığında, burada sunulan protokol basit ve oldukça tekrarlanabilir: sertlik, bir cam mikrosfer girintisi ile ölçülen ve Hertz modeli kullanılarak, arasında 400 ve 600 Pa. Benzer protokoller başarıyla B hücreleri için sadece kullanılmıştır35 ama aynı zamanda T hücreleriiçin 36. Nanopillars ile karşılaştırıldığında (Ayrıca T lenfositler için kullanılan37) düz homojen bir yüzey sağlar, bu nedenle jel etkileşimi esas olarak yüzeye teğet olarak kısıtlanmış olarak sonuçları yorumlamak daha kolaydır.
Tarif ettiğimiz protokol, B hücrelerinin antijen sunan yüzeyler üzerinde uyguladığı kuvvetlerin spatiotemporal dinamiklerine erişim sağlar. Mekansal düzeyde bu kuvvetlerin lokalizasyonu hakkında bilgi sağlar ve floresan mikroskobu ile birlikte, deneyci belirli moleküllerin varlığı ile yerel kuvvetleri ilişkilendirmek için sağlar (yani, sitoiskelet veya BCR sinyal kaskad bileşenleri). Zamansal düzeyde, zaman noktası başına bir değer sağlamak ve gürültüyü azaltmak için miktarları (toplam enerji veya toplam stres gibi) entegre etmek mümkündür. Bu zaman içinde çekiş kuvvetevrimi gözlem sağlar (büyüme ve plato) ve pulsatil desenlerin varlığı.
Analiz için kritik deneysel yönleri aşağıdaki gibi açıklanmıştır. (i) Hücre yoğunluğu: doğru bir analiz yapmak için, hücreler yeterince ayrılmalıdır. Bir hücrenin etrafında kendi boyutunda boş bir bölge varsa analiz edilebilir olduğunu düşünüyoruz. (ii) İletim görüntüsü: Deney sırasında analizde maske olarak kullanılmak üzere hücrelerin en az bir iletim görüntüsünün toplanması tavsiye edilir. (iii) Resimdeki boncuk sayısı: yalnızca sinapstaki boncuk sayısının 30 ile 200 arasında olduğu (yani 1-8 boncuk/μm²) görüntüleri analiz etmenizi öneririz. Düşük yoğunluklar yeterli harita deplasman rekonstrüksiyonuna izin vermez. Yüksek boncuk yoğunluğu tek parçacık takibini güvenilmez yapar. (iv) Deney sırasında boncuk sayısı sabit olmalıdır; ancak, görüntüleme koşullarında (özellikle birbirine çok yakın boncuklarda) küçük değişkenlik nedeniyle dalgalanmalar oluşabilir. Odak kayması, meydana geliyorsa, düzeltilmeli ve sorunlu çerçeveler atılmalıdır. (v) Jel kalitesi: çok fazla çatlak, boncuk dağılımında değişkenlik veya çok kalın jeller ile jeller atılmalıdır. (vi) Hücre tipine bağlı olarak, tekrarlanan maruziyetlerden sonra, geç zaman noktalarındaki hücreler (>300 kare) fototoksik etkilere maruz kalabilir. Bu verilerle karşılaştırıldığında bir "taban çizgisi" olarak hücrelerden yoksun bir maske üzerinde programı çalıştırmak için tavsiye edilir. Bu sadece deneysel koşullar nedeniyle gürültü seviyesinin bir büyüklük sağlar.
Klasik yapışmada çekiş kuvvetini ölçmek için kullanılan jeller odak yapışmasında meydana gelen süreçlerin araştırılmasına olanak sağlar (aktin akımları ve sinyal moleküllerinin işe alınması)—kuvvetlerin uygulandığı noktalar38,39. Ancak, sinapstaki kuvvetler odak yapışıklıkları ile uygulanmaz. B hücresi immün sinapslarında kuvvet oluşumunun spatiotemporal paterni yakın zamana kadar bu yöntem kullanılarak nicel olarak araştırılmamıştır. TFM kullanarak, biz ilk kez gözlenen, B hücre immün sinaps kuvvet desenleme, bizim son çalışmada sunulan6, lenfositler çalışmada cesaret verici bakış açıları açılması.
Özellikle, bu yöntem kuvvet hesaplaması için bir referans görüntü olarak jel üzerinde hücrelerin gelmeden önce çekilen bir görüntü kullanır. Her zamanki TFM protokolleri deney sonunda referans görüntü alarak öneririz, trypsin ile hücreleri ayırdıktan sonra; bu, deneycinin hücreler açısından zengin bir bölge aramasını sağlar. Bu da burada mümkün olsa da, tripsin antijen kaplı jel B hücrelerini ayırma oldukça verimsiz, bir kopma ve jel modifikasyon ve hareketleri riski (tüm veri seti kullanılamaz yapmak) için uzun beklemek gerekir daha yüksektir.
Burada sunulan yöntem esnektir ve diğer proteinlerin jel yüzeyine (örneğin, integrin ligandlar ve immünglobulinler test edilmiştir) ve hatta floresan antijene aşılanmasına olanak sağladığından, bağışıklık sinapstaki diğer sinyallerin etkisini incelemek için uygulanabilir (bkz. bölüm 4). Ayrıca, hücreler ilaç tedavisi ve lokal tedirginlikler için deneyci için erişilebilir kalır. Son olarak, yöntem de sabit hücreleri görüntüleme ile uyumludur. Bu gözlemler için jelin bir kapak kayması üzerinde kullanılması, hücreleri lekelemesi, kapak kaymasını kaydırak üzerine yapıştırması ve ancak daha sonra montaj ortamı ve başka bir kapak kayması eklemesi önerilir. Gözlem daha sonra jel aracılığıyla görüntünün bozulmasını önlemek için üst jel ile yapılacaktır.
Olası tuzaklar polimerizasyon ve kaplama jel değişkenliği vardır. Polimerizasyon problemleri esas olarak başlatıcı/katalizör kalitesinden kaynaklanmaktadır. Ayrıca, jel, özellikle montaj dan hemen sonra kullanılmaz, şişirmek olabilir. Bu sorun önemli ölçüde jel mekanik özelliklerini etkilemek için görünmüyor, ama etkili jel işe yaramaz hale, amaç için ulaşılmaz boncuk tabakası yapabilirsiniz. Bu sorun ortaya çıktığında her durum için ekstra jel hazırlamanızı öneririz. Kaplamada da belirli bir değişkenlik olabilir ve sulfo SANPAH'ın taze seyreltilmesi çok önemlidir.
Sonuç olarak, BCR ligand tarafından aktive edildiğinde immünolojik sinapsta B hücreleri tarafından uygulanan kuvvetleri ölçmek için basit, ucuz ve tekrarlanabilir bir yöntem tanımladık. Bu uygun reseptör ligand kullanımı ile diğer ligands ve lenfositler (bellek B hücreleri, T hücreleri, vb) diğer tür reaksiyon çalışması için adapte edilebilir.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Yazarlar eleştirel okuma için M. Bolger-Munro teşekkür ve Nikon Görüntüleme Center@CNRS-InstitutCurie ve PICT-IBiSA, Institut Curie, Paris, Fransa-BioImaging ulusal araştırma altyapısı üyesi, görüntü edinimi ve Curie Hayvan Tesisi destek için kabul. PP CNRS tarafından desteklendi. AK ve JP Paris Descartes Doktora bursu ve Ecole Doctorale FIRE-Program Bettencourt tarafından desteklendi. Bu proje PP (ANR-10-JCJC-1504-Immuphy) ve AMLD (ANR-PoLyBex-12-BSV3-0014-001, ERC-Kayışacemi-GA 243103) hibe ile finanse edilmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 3-aminopropyltrimethoxysilane (APTMS) | Sigma-Aldrich | 281778 | Store aliquoted, protected from humidity |
| 40% Acrylamide Solution | Biorad | 1610140 | |
| Alexa555 microscale protein labeling kit | Molecular Probes | A30007 | |
| Ammonium Persulfate (APS) | Sigma-Aldrich | A3678 | |
| B cell Isolation Kit, Mouse | Miltenyi Biotec | 130-090-862 | |
| B-mercaptoethanol | Gibco | 31350-010 | |
| 2% Bis Solution | Biorad | 161-0142 | |
| Bovine Serum Albumin (BSA) | Euromedex | 04-100-812-C | |
| Coverslip 18mm | VWR | 631-1580 | |
| Fetal calf serum | PAA | A15-151 | Decomplemented (40min @56°C) |
| Fluorodishes FD35 | World Precision Instruments, Inc | FD35100 | |
| Fluosphere: carboxylate-modified, 0.2um, dark red | Molecular Probes | F8807 | |
| Hen Egg Lysozyme | Sigma-Aldrich | L6876 | Stocked in aliquote 100mg/ml |
| MEM Non-Essential Amino Acids Solution (100X) | Thermofisher/Gibco | 11140035 | |
| N,N,N',N'-tetrametiletilendiammine (TEMED) | Euromedex | 50406-B | |
| PBS (Phosfate Buffer Saline) | Gibco | 10010-015 | |
| Penicillin–streptomycin | Gibco | 15140-010 | |
| RMPI 1640 – Glutamax I | Thermofisher | 61870-010 | |
| Sigmacote | Sigma-Aldrich | SL2 | |
| Sodium pyruvate | Gibco | 11360-039 | |
| sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino)hexanoate (Sulfo-SANPAH) | Thermo Scientific | 22589 |
Referanslar
- Yuseff, M. -. I., Pierobon, P., Reversat, A., Lennon-Duménil, A. -. M. How B cells capture, process and present antigens: a crucial role for cell polarity. Nature Reviews. Immunology. 13 (7), 475-486 (2013).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Shaheen, S., Wan, Z., et al. Substrate stiffness governs the initiation of B cell activation by the concerted signaling of PKCβ and focal adhesion kinase. eLife. 6, (2017).
- Natkanski, E., et al. B cells use mechanical energy to discriminate antigen affinities. Science. 340 (6140), 1587-1590 (2013).
- Wan, Z., Chen, X., et al. The activation of IgM- or isotype-switched IgG- and IgE-BCR exhibits distinct mechanical force sensitivity and threshold. eLife. 4, (2015).
- Kumari, A., Pineau, J., et al. Actomyosin-driven force patterning controls endocytosis at the immune synapse. Nature Communications. 10 (1), 2870 (2019).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Schwarz, U. S., Soiné, J. R. D. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica et Biophysica Acta. 1853 (11), 3095-3104 (2015).
- Colin-York, H., Shrestha, D., et al. Super-Resolved Traction Force Microscopy (STFM). Nano Letters. 16 (4), 2633-2638 (2016).
- Stubb, A., Laine, R. F., Guzmán, C., Henriques, R., Jacquemet, G., Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. BioRxiv. , (2019).
- Gutierrez, E., Tkachenko, E., et al. High refractive index silicone gels for simultaneous total internal reflection fluorescence and traction force microscopy of adherent cells. Plos One. 6 (9), 23807 (2011).
- Bergert, M., Lendenmann, T., et al. Confocal reference free traction force microscopy. Nature Communications. 7, 12814 (2016).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Colin-York, H., Fritzsche, M. The future of traction force microscopy. Current Opinion in Biomedical Engineering. 5, 1-5 (2018).
- Feng, Y., et al. Mechanosensing drives acuity of αβ T-cell recognition. Proceedings of the National Academy of Sciences of the United States of America. 114 (39), 8204-8213 (2017).
- Spillane, K. M., Tolar, P. DNA-Based Probes for Measuring Mechanical Forces in Cell-Cell Contacts: Application to B Cell Antigen Extraction from Immune Synapses. Methods in Molecular Biology. 1707, 69-80 (2018).
- Stabley, D. R., Jurchenko, C., Marshall, S. S., Salaita, K. S. Visualizing mechanical tension across membrane receptors with a fluorescent sensor. Nature Methods. 9 (1), 64-67 (2011).
- Merkel, R., Nassoy, P., Leung, A., Ritchie, K., Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 397 (6714), 50-53 (1999).
- Hinterdorfer, P., Dufrêne, Y. F. Detection and localization of single molecular recognition events using atomic force microscopy. Nature Methods. 3 (5), 347-355 (2006).
- Sawicka, A., Babataheri, A., et al. Micropipette force probe to quantify single-cell force generation: application to T-cell activation. Molecular Biology of the Cell. 28 (23), 3229-3239 (2017).
- Desprat, N., Guiroy, A., Asnacios, A. Microplates-based rheometer for a single living cell. Review of Scientific Instruments. 77 (5), 055111 (2006).
- Labernadie, A., Bouissou, A., et al. Protrusion force microscopy reveals oscillatory force generation and mechanosensing activity of human macrophage podosomes. Nature Communications. 5, 5343 (2014).
- Bouissou, A., Proag, A., et al. Protrusion force microscopy: A method to quantify forces developed by cell protrusions. Journal of Visualized Experiments. (136), 57636 (2018).
- Kronenberg, N. M., Liehm, P., et al. Long-term imaging of cellular forces with high precision by elastic resonator interference stress microscopy. Nature Cell Biology. 19 (7), 864-872 (2017).
- Basu, R., Whitlock, B. M., et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell. 165 (1), 100-110 (2016).
- Bufi, N., Saitakis, M., et al. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophysical Journal. 108 (9), 2181-2190 (2015).
- Goodnow, C. C., Crosbie, J., et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature. 334 (6184), 676-682 (1988).
- Butler, J. P., Tolić-Nørrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- . MBPPlab/TFM_v1: Software for Time dependent Traction Force Microscopy Available from: https://github.com/MBPPlab/TFM_v1 (2019)
- Tseng, Q., Duchemin-Pelletier, E., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1506-1511 (2012).
- . ImageJ plugins by Qingzong TSENG Available from: https://sites.google.com/site/qingzongtseng/ (2019)
- Plotnikov, S. V., Sabass, B., Schwarz, U. S., Waterman, C. M. High-resolution traction force microscopy. Methods in Cell Biology. 123, 367-394 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Wang, J., Lin, F., et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Science Signaling. 11 (542), (2018).
- Hui, K. L., Balagopalan, L., Samelson, L. E., Upadhyaya, A. Cytoskeletal forces during signaling activation in Jurkat T-cells. Molecular Biology of the Cell. 26 (4), 685-695 (2015).
- Bashour, K. T., Gondarenko, A., et al. CD28 and CD3 have complementary roles in T-cell traction forces. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2241-2246 (2014).
- Gardel, M. L., Sabass, B., Ji, L., Danuser, G., Schwarz, U. S., Waterman, C. M. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of Cell Biology. 183 (6), 999-1005 (2008).
- Stricker, J., Sabass, B., Schwarz, U. S., Gardel, M. L. Optimization of traction force microscopy for micron-sized focal adhesions. Journal of Physics. Condensed Matter. 22 (19), 194104 (2010).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır