
Method Article
Yara kaynaklı Poliploidizasyonu Incelemek için Bir Drosophila Modeli
Bu Makalede
Özet
Yara kaynaklı poliploidizasyon, hücrelerin hücre kaybını telafi etmek için bölmek yerine boyut olarak büyüdüğü korunmuş bir doku onarım stratejisidir. Burada epitel yara onarım ploidy ve genetik düzenleme ölçmek için bir model olarak meyve sinek nasıl kullanılacağı hakkında ayrıntılı bir protokoldür.
Özet
Poliploidi organizma sağlığı ve hastalığı üzerindeki etkisi hala tam olarak anlaşılamayan sık rastlanan bir olgudur. Bir hücre, endorepyon veya hücre füzyonu sonucu kromozomlarının diploid kopyasından daha fazlasını içeriyorsa poliploid olarak tanımlanır. Doku onarımında, yara kaynaklı poliploidizasyon (WIP) meyve sineklerinden omurgalılara kadar korunmuş bir iyileşme stratejisi olarak bulunmuştur. WIP onkojenik büyüme ve genotoksik strese direnç de dahil olmak üzere hücre çoğalması üzerinde çeşitli avantajları vardır. Zorluk, poliploid hücrelerin neden ortaya çıkıp bu eşsiz hücrelerin nasıl çalıştığını belirlemekolmuştur. Sağlanan poliploid hücrelerin bir delinme yarası sonra 2 gün içinde üretilen yetişkin meyve sinek epitel WIP çalışma için ayrıntılı bir protokoldür. D. melanogaster'in kapsamlı genetik araç kitinden yararlanarak, MYC de dahil olmak üzere WIP'yi başlatmak ve düzenlemek için gereken genler tespit edilmeye başlandı. Bu yöntemi kullanarak devam eden çalışmalar, cinsiyet, diyet ve yaş gibi diğer genetik ve fizyolojik değişkenlerin WIP'nin işlevini nasıl düzenlediğini ve etkilediğini ortaya çıkarabilir.
Giriş
Drosophila melanogaster epitel yara onarım hücresel ve moleküler mekanizmaları incelemek için çekici bir model sistemidir. Memelilerde olduğu gibi kullanılan doku onarım mekanizmaları hem dokuya hem de gelişim evresine bağlıdır. Scarless yara iyileşmesi meyve sinek embriyo oluşur nerede bir actomyosin "çanta dize" epitel önde gelen kenarında formları sorunsuz yakın yara sağlayan1,2. Larvalarda post-embriyonik yara iyileşmesi, pupa, ve yetişkin meyve sinekleri ekstrasellüler matriks remodeling sonuçları, melanin skar oluşumu, ve epitel hücre büyümesi3,4,5,6. Epitel hücreleri hücre füzyonu ve endocycle, mitoz,3,4,7,,8atlar eksik bir hücre döngüsü ile boyutu artış . Sonuç olarak, hücre kaybı hücre bölünmesi yerine poliploid hücre büyümesi ile telafi edilir. Yetişkin sinek hindgut, midgut, ve foliküler epitel de doku hasarısonrahücre kaybı telafi etmek için poliploid hücre büyümesi güveniyor 9,10,11.
Poliploidi bitki ve böceklerde organizmasal gelişimiyi iyi bilinen bir yönüdür, ancak son birkaç yıl içinde poliploidvertebras korunmuş bir doku onarım stratejisi olduğunu daha belirgin hale gelmiştir12. Kalbini yenileme kapasitesine sahip olan zebra balığı, hasarlı epikardiyum13'üniyileşmesi için poliploid hücre büyümesine güvenir. Poliploid de memeli karaciğer rejenerasyonu ve böbrek tübül epitel onarım akut yaralanma sonra katkıda14,15. Bu örneklerde poliploid hücreler endosikasyon yoluyla endosiklasyon yoluyla veya endomitoz isme ile üretilir, bu da sitokinezdeki bir blok nedeniyle binükleli hücre ile sonuçlanır12. Muamma neden poliploid hücrelerin yara onarımı sırasında ortaya çıkması ve poliploidin doku fonksiyonlarını nasıl etkilediğidir. Son çalışmalar poliploidbir şifa avantajı veya dezavantaj sunuyor olup olmadığı sorusuna yeni bir fikir sağlamıştır. Zebra balığı epikardiyum, poliploid yara iyileşme hızını artırdı13. D. melanogaster hindgut ve memeli karaciğer, poliploid onkojenik büyümeye karşı koruyucu olduğu bulunmuştur11,14. Yetişkin sinek epitel, son zamanlarda poliploid genotoksik stres varlığında yara onarımı sağlar bulundu16. Endoçoğalma DNA hasarına karşı dirençlidir, hücre çoğalması aksi takdirde tehlikeye olurdu yara iyileşmesi sağlayan17. Fare ve zebra balığı kalplerinde kardiyomiyositler için, ancak, poliploid iyileşme yavaşlar, gelişmiş yara oluşumu ile sonuçlanan18,19. Bu nedenle, organ ve / veya hücre tipine bağlı olarak, poliploid yararlı veya zararlı doku onarım stratejisi olabilir. D. melanogaster genetiğinin erişilebilirliği, yara kaynaklı poliploidizasyon (WIP) yanıtının analizi ile birleştiğinde, bu yara iyileşme stratejisini yönlendiren moleküler ve hücresel mekanizmaları niçin aydınlatılması için ideal bir model sistemi dir.
Burada, yetişkin D. melanogaster epitel WIP analiz için bir protokol salıyoruz. Meyve sineği yaralanması, diseksiyon, immünboyama, montaj, görüntüleme ve yeniden epitelizasyon, hücre füzyonu ve endorepsiyon (ploidi) analizi için talimatlar dahildir. Görüntüleme ve ploidi analizi, WIP'nin oluşup oluşmadığını test etmek için diğer modellere de uyarlanabilir. Bu nükleer DNA içeriğinde bir artış ile sık sık nükleer boyutu karşılık gelen bir artış olduğu unutulmamalıdır. Ancak, nükleer boyutu ploidy20karşılık gelen bir değişiklik yansıtmaz biyoloji birçok örnek vardır. Hücrelerin genellikle yayıldığı veya yara bölgesini kapsayacak şekilde uzanacağı bir yara ortamı bağlamında nükleer boyutu yorumlarken daha da dikkatli olunmalıdır. Bu nedenle, ploidi değişim tek kesin kanıtı bu yöntem (veya diğerleri, tüm genom dizileme gibi)21DNA içeriğini ölçmektir. Bu yöntem, yara onarımında poliploidin rolünü ve düzenlenmesini incelemek için bir model olarak erişkin D. melanogaster abdominal epitelin uygunluğunu arttırmaktadır.
Protokol
1. Yetişkin meyve sineklerinin evreleme ve yaralama
- Seçtiğiniz D. melanogaster suş seçim (yani, epi-Gal4 / UAS suşu, Malzeme Tablosubakınız).
NOT: Burada Gal4/UAS sistemi, uas'ın aşağı kodlanmış bir genin epitelyal spesifik gen ekspresyonunu (epi-Gal4) veya RNAi'yi etkinleştirmek için kullanılır. Bu çalışmada floresan membran proteini (UAS-Cd8.mRFP), mitotik indükleyici (UAS-fzrRNAi, UAS-stg) ve WIP inhibitörü (UAS-E2F1RNAi; UAS-RacDN). - 10-15 yeni eklenmiş dişi meyve iki şişe toplamak ve 3-5 gün eski kadar 25 °C taze gıda şişeleri üzerinde yaş. Bir şişe yaralanmamış kontrol olarak görev yapacak ve diğer şişe aşağıda açıklandığı gibi yaralı olacaktır. Dişi sinekler erkeklerle birlikte muhafaza edilmelidir (~5/vial).
- Sinekleri yaralamak için, her biri tek bir 0,10 mm paslanmaz çelik pimile birkaç pim tutucu monte edin. Pimin keskin ucunun dışarı yasolduğundan emin olun. İğneler sineği deldikten sonra kolayca bükülebilir veya talaşlanabilir ve kancalı veya hasarlı iğneler atılmalıdır.
- Bir stereomikroskop altında bir CO2-sinek ped üzerinde yaşlı dişi meyve sinekler anestezi ve bir boya fırçası kullanarak bir satır içine hizalamak. Güvenlik gözlüğü takmak, bir elinde pim tutucuyu tutarken diğer ini de forseps, ventral karın kısmı yukarı bakacak şekilde bir sineği konumlandırmak için forseps kullanın.
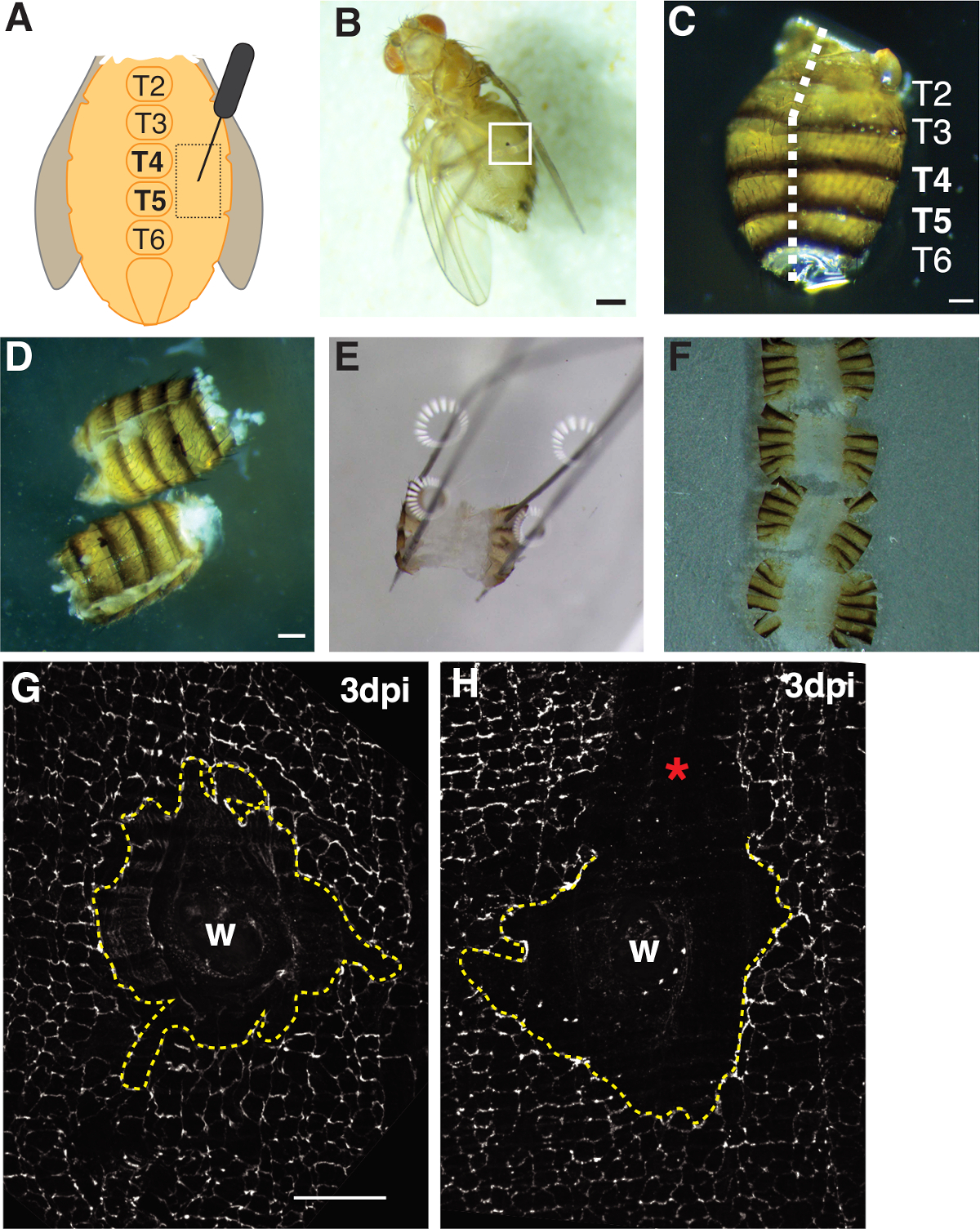
- Erişkin dişi, ventral orta hat sternitlerinin her iki tarafında tergit E4'ün epitel pleürit bölgesinde uçar (Şekil 1A). Bu ventral bölgeyi delmek, doku kenarlarının mekanik işleme ile yırtılacak olan diseksiyon bölgelerinden en uygun alanı sağlar.
- Yaralı sinekleri yiyecek şişesine ve yaşa, istenilen gün sonrası yaralanmaya (dpi) geri getirin. Epitelyal yara iyileşmesi 1 dpi ile başlar ve 3 dpi ile sona erer. Endorepyon zirveleri 2 dpi, EdU teşpiçin idealdir (bölüm 4, Şekil 2).
2. Uç karın diseksiyonu
NOT: Bu adımda ventral karın dokusuna diseksiyon araçları ile dokunmamak önemlidir, çünkü epitelin bütünlüğünü tehlikeye atacaktır.
- Diseksiyon için gerekli tüm malzemeleri edinin: Grace'in çözeltisi, forsepsleri, Vanna'nın Yay makasları, 0,10 mm pimleri, diseksiyon plakaları, 9 kuyu cam diseksiyonu, fiksatif solüsyon (%4 1x PBS), 1x PBS, mendil, pipet ler ve 30 μm için uçlar ve eldivenler (Bkz. Malzeme Tablosu).
- Sineklerin stereomikroskop altında CO2-flypad üzerinde yaralı sinekleri anestezi ve yara izi varlığını kontrol ederek başarıyla yaralandığını doğrulayın (yani, karında bir melanin noktası, Bkz. Şekil 1B). Başarılı bir şekilde yaralanmayan deney grubundan herhangi bir sinek atın.
- Diseksiyonu başlatmak için Grace'in çözümüyle 9 kuyulu cam diseksiyon kabından birini doldurun. Thorax'ın sırt tarafında yaralı bir dişi sineği kavramak ve Grace'in solüsyonunu içeren kuyuda sineği batırmak için bir çift forseps kullanın.
- Toraks bırakmadan karşı elinde forseps kullanarak, tergitE A6 altında dorsal mitikül delin ve meyve sinek arka ucundan cuticle çekin. İç organlar (yumurtalık ve bağırsak) genellikle bu adımda çıkacaktır. Değilse, yavaşça geri kalan organları sıkmak ve boş bir kuyuya atmak için forceps ile karın sırt tarafında itin.
- TergitE A2'nin üzerindeki toraks kavşağında forsepsleri kullanarak karının tamamını koparın ve karnı Grace'in çözeltisinin ~100 μL'sini içeren boş bir kuyuya aktarın.
- Tüm sinek karınları parçalanana kadar 2.3-2.5 adımlarını tekrarlayın.
- Grace'in çözeltisinin hacmini havuzlu, parçalanmış karınlarda 30°L'ye düşürün.
- Bir elinde forseps ile karın sırt tarafında konumlandırma ve daha sonra diğer el ile karın boşluğuna Vanna'nın bahar makas alt bıçak yerleştirerek karın açık. Karın tamamen açılana kadar dorsal orta hat boyunca kesin, hangi kadar üç kesim gerektirebilir(Şekil 1C, 1D).
- Karın montaj alanı başına dört adet 0,10 mm pimli kuru bir kesme plakası kurutun. Her 35 mm'lik kesme plakası yedi montaj alanına kadar sığabilir. Pipet 30 μL Grace'in çözeltisi her montaj alanında ve her damlacık için bir filetolu karın aktarın.
- Filetolu karınları dört dorsal köşedeki tabağa sabitle(Şekil 1E). Karın dokusunu yırtmadan veya aşırı germeden dokunun düz yattığından emin olun.
- Dokuyu onarmak için Grace'in çözeltisinin pipetini kapatın ve sabitlenmiş karına 30 μL düzeltme çözeltisi ekleyin.
DİkKAT: Paraformaldehit toksik olduğu için düzeltme çözeltisini kullanırken eldiven takın. - Tüm filetolu karınlar diseksiyon plakasına sabitlenene kadar 2.10-2.11 adımlarını tekrarlayın.
- Her kontrol ve deney grubunu işaretlemek için her yemeğin altına bir bant etiketi yerleştirin. Oda sıcaklığında (RT) 30-60 dk için düzeltme örnekleri.
- Her plakaya 1,5 mL 1x PBS boru takarak düzeltme çözeltisini yıkayın. Düzeltme çözeltisini ve plastikleri kurumsal yönergelere uygun sıvı veya kuru kimyasal atık kaplarına atın.
- 1x PBS'nin 1,5 mL'lik yıkama plakaları 2x'i yıkayın ve 1,5 mL 1x PBS ile kaplı sabit dokuyu kapaklı plastik bir kapta saklayın. Kabın altına bir tabaka nemli kağıt havlu ekleyin ve numuneleri 1 hafta boyunca diseksiyondan sonra immunostain'e hazır olana kadar 4 °C'de saklayın.
3. İmmünofluoresans
- Reaktifleri yeni hazırlayın (bkz: Malzeme Tablosu):tampon çözeltisini yıkayın (%0.3 Triton X 100, 1x PBS'de %0.3 BSA). Artık yıkama tamponu 4 °C'de kaydedilebilir ve 2 günlük boyama protokolü süresince kullanılabilir. Anti-FasIII (1:50Figure 2fare anti-Fasciclin-III) kullanarak tsay başına yeterli birincil antikor çözeltisi hazırlayın (1:50 fare anti-Fasciclin-III) ya anti-Grh (1:300 yakınlık saf tavşan anti-Grainyhead8)veya anti-RFP (1:1.000 tavşan anti-RFP) ile yıkama tamponu. Birincil antikor çözeltileri 4 °C'de kaydedilebilir ve sinyal önemli ölçüde azalana kadar birden çok kez yeniden kullanılabilir.
- 1x PBS'yi borulandırarak, 1,5 mL yıkama tamponu ekleyerek ve RT'de bir orbital shaker (80 rpm) üzerinde en az 30 dakika kuluçkaya yatarak dokuyu permeabilize edin.
- Yıkama tamponu ve leke dokusunu bir gecede 1,5 mL primer antikor solüsyonu ile çıkarın, 4 °C'de orbital shaker (80 rpm) ile kuluçkaya yatırın. Birincil antikor çözeltisini toplayın ve gelecekteki deneyler için 4 °C'de bir tüpte saklayın.
- Önce numuneyi 1x PBS ile hızlı bir şekilde durulayın ve ardından 1,5 mL yıkama tamponu yla 3x yıkayın. Her yıkama için, en az 30 dakika boyunca bir orbital shaker üzerinde RT örnekleri inkübat.
- Son yıkama sırasında, ikincil antikor solüsyonu hazırlamak: 1:1.000 eşek anti-tavşan Alexa 488 veya 568 ve 1:1.000 keçi anti-fare Alexa 488 veya 568 (veya seçim floroporores) yıkama tampon.
- Yıkama tamponunu ve leke dokularını 1,5 mL ikincil antikor solüsyonu ile çıkarın. Alüminyum folyo ile kapak örnekleri ve 3 saat için RT bir orbital shaker üzerinde inkübat. Alternatif olarak, numuneler bir gecede 4 °C'de bir orbital shaker üzerinde kuluçkaya yatırılabilir.
- Numuneleri önce ikincil antikor solüsyonu atarak yıkayın ve ardından numuneyi 1x PBS ile hızlı bir şekilde durulayın ve ardından 1,5 mL yıkama tamponu ile üç yıkama yıkayın. Her yıkama için, en az 30 dakika boyunca bir orbital shaker üzerinde RT örnekleri inkübat.
- DAPI'yi yıkama tamponunda 10 μg/mL'ye seyrelterek DAPI çözeltisini hazırlayın. Son yıkamadan sonra, 1,5 mL DAPI çözeltisi ile leke örnekleri 30 dakika boyunca RT'de kuluçkaya yatırılanın.
- DAPI çözeltisini atın ve numuneleri 1,5 mL 1x PBS'de 2 x durulayın. Lekeli dokuyu 1,5 mL 1x PBS karanlıkta, alüminyum folyo ile kaplı olarak 4 °C'de, kapaklı cam bir kaydırağa monte etmeye hazır olana kadar saklayın. Montaj adımı 1 hafta içinde yapılmalıdır.
4. Hücre döngüsü aktivitesi (EdU Suresi)
- Click-iT kitinden (Bkz. Malzeme Tablosu)10 mM EdU stok çözeltisi dH20'da EdU tozunu eriterek ve ~15 dk tamamen çözülene kadar karıştırarak hazırlayın. Stok çözeltisi aliquoted (tüp başına 250 μL) ve -80 °C'de saklanabilir.
- Yem ilk dH2O 5 mM EdU stok seyreltme tarafından EdU uçar. 0,5 mL'lik bir tüp kapağını kesin ve sineklik şişesinin altına yerleştirin. Kapağı yiyeceğe itin, böylece stabil olur.
- Sinekleri anesthetize ve şişe içine 3-5 günlük sinekler aktarın. Sineklere tek bir kenara dokunun, böylece hiçbiri kapağına yapışmaz.
- Pipet 75 μL maya-EdU çözeltisi kapağına. Sinekler her gün taze maya-EdU çözeltisi ile beslenmeli ve sineklerin şişenin dibine sıkışmamasını sağlamak için her gün bir kapaklı taze gıda şişesine aktarılmalıdır.
- Sinekaktarmak için, bir kap ile yeni bir şişe çevirmek, uyku sinekkoymak, bir tarafa sinek dokunun ve taze maya-EdU çözeltisi ekleyin.
- Üçüncü gün, sinekler zarar ve 2 dpi diseksiyon kadar maya-EdU beslemeye devam (Şekil 4A). Diseksiyon ve fiksasyon yöntemleri için protokol bölüm 2'ye bakın.
- Hazırlayın EdU boyama reaktifleri: yıkama tamponu (%0.3 Triton X 100, 1x PBS'de %0,3 BSA), permeabilizasyon tamponu (%0,5 Triton X 100 1x PBS), tamponengelleme (%3 BSA 1x PBS) ve EdU titre kitinden reaktifler (Bkz. Malzeme Tablosunabakınız), üretici tarafından yönlendirilen 1x reaksiyon tamponu ve 1x reaksiyon tamponu katkı maddesi dahil olmak üzere reaktifler hazırlayın.
- 1 saat boyunca numuneleri yıkayın, RT'de 1,5 mL yıkama tamponu karıştırın.
- 20 dakika boyunca 1,5 mL permeabilizasyon tamponu ve kuluçka numunesi ekleyin.
NOT: 500 μL/plaka hacmi ni kullanarak reaksiyon kokteyli çözeltisini eritin ve hazırlayın. - Örnekleri 1x PBS ile 1 x hızlı bir şekilde yıkayın ve ardından 1 mL engelleme tamponu ile 3 kat hızlı bir şekilde yıkayın.
- Pipet kalan tüm engelleme tamponunu kapatın ve plaka başına 500 μL reaksiyon kokteyl çözeltisi ekleyin. Dokuların tamamen kapalı olduğundan emin olmak için girdap plakaları. RT'de 1 saat boyunca karanlıkta bir çekmecede kuluçka.
- 1,5 mL engelleme tamponu ile numuneleri 1 kat hızlı yıkayın.
- 1:5.000'de 1.5 mL DAPI çözeltisi ile 30 dk yıkama tamponu ile leke.
- 2x'i 1x PBS ile hızlıca yıkayın, folyoya sarın ve numuneleri 3 gün içinde monte etmeye hazır olana kadar 4 °C'de karanlıkta saklayın.
5. Lekeli doku montajı
- Montaj için gerekli tüm malzemeleri edinin: cam kaydıraklar, cam kapaklar, açık oje, montaj ortamı, bir çift forseps ve mendil.
- Lekeli sinek dokusu monte etmek için, stereomikroskop altında forceps kullanarak kesme plakasından karınları unpin. Dorsal yanlarından forcep'lerle dokuyu hafifçe tutarak, ventral alana forceps ile dokunmamaya özen tutarak, bir cam kapak üzerine montaj ortamının ~30 μL'sine doku aktarın.
- Stereomikroskop altında karın dokusunu yönlendirin, böylece iç kapak kaymaya doğru bakar (yani, dış kütikül/kıllar yukarı bakacak şekilde). Forceps kullanarak medya damlacık kenarına odaklı karın çekin. Yüzey gerilimi dokuyu düz tutmaya yardımcı olacaktır (Şekil 1F).
NOT: Bu aşamada bir sütun veya satırda karın düzenlemek için görüntüleme için yararlıdır. - Cam bir slaytetiketle (örn. kontrol veya deneysel) ve kaydırağı yavaşça kapak kaymasına yaklaştırarak kapak kaymasını alın. Fazla montaj ortamını kaldırmak için kaydırağı ters çevirin ve bir silme ile hafifçe lekeleyin.
- Coverslip'in kenarlarını açık oje ile kapatın ve kalan tüm deney grupları için tekrarlayın. Slaytları 4 °C'de bir slayt kutusunda görüntülenmeye hazır olana kadar saklayın.
6. Görüntüleme ve işleme
- Melanin yarasını ilk olarak bir konfokal mikroskop(Şekil 1B),bir nokta tarayıcı sı veya yapılandırılmış bir aydınlatma (ApoTome) ile 40x yağ veya kuru bir amaç ile bularak sinek karın yara alanını görüntüleyin.
- Sinyalin doygunluk altında olduğundan emin olmak için her kanaldaki pozlamayı kontrol edin. Görüntüleme ayarları en parlak örnek grubuna dayanmalıdır. DAPI kanalının DNA içeriğini doğru bir şekilde ölçmek için doğrusal aralıkta kalması gerektiğinden, ploidi analizi için bu özellikle önemlidir.
- Dilimler arasında en iyi 0,50 m'lik en uygun mesafeye sahip üç kanalda da tam bir z-stack görüntüsü alın. Yakalanan görüntüleri kaydedin ve dosyayı görüntü çözümleme programı Fiji'de (ImageJ olarak da bilinir) açın.
- Her görüntü için, tüm kanallar için dilimler seçeneğini kullanarak bir z yığını projeksiyonu oluşturun.
- Çekirdeklerin görüntüler arasında yatay olarak dizilmesini sağlamak için görüntüleri gerektiği gibi döndürün(Şekil 3A ve Şekil 4E).
- Tüm görüntüleri, yara bölgesinin veya yaralanmamış kontrol merkezinin etrafında ortalanmış 300 μm x 300 m'lik dikdörtgen bir seçkiye kazıyın. Dikdörtgen çizerek ve Düzenleme'yi seçerek alanı tanımlayın | Seçim | Belirtin. Tüm görüntülerin analiz edilmesi için aynı boyuttakutunun kullanıldığından emin olmak için ölçeklenmiş birimlerin mikronlar halinde olduğundan emin olun.
7. Endoreplik (ploidy) analizi
- Fiji'yi kullanarak Grh kanal penceresini seçin ve görüntüyü çoğaltın. Ardından maske oluşturmak için eşik aracını kullanın. Çekirdeklerin büyük ölçüde küçülmesine neden olmadan arka planı en aza indirmek için üst çubuğu kaydırarak eşiği el ile ayarlayın (Şekil 4D).
- Grh kanalındaki herhangi bir çekirdek eşik görüntüsüne dokunuyorsa, çekirdekler arasında bir çizgi çizmek için arka planla aynı renkte paintbrush aracını (2 piksel genişliği) kullanın. Son maskeyi oluşturmak için tamamlandığında 1x uygulamak için tıklatın.
- Parçacıkları Analiz etme işlevini kullanarak bir ilgi bölgesi (ROI) haritası oluşturun: arka plan dahil olmadan çekirdeklerin çoğunu yakalamak için boyutu 5 μm-60 m'ye ayarlayın.
- YG yöneticisindegerektiği gibi YG haritasını el ile ayarlayın. Çekirdek olmayan seçimleri silin ve serbest el seçim aracıyla çekirdeğin ana hatlarını çizerek ve YG yöneticisine ekleyerek tanımlanmayan listeye çekirdek ekleyin (Şekil 4D).
- DAPI kanalını seçin ve Grh kanalında oluşturulan yg haritasını DAPI kanalına uygulamak için YG yöneticisinde Tümünü Göster'i tıklatın.
- E-posta çekirdeği anahattının nonepithelyal çekirdeklerle (örn. kas veya yağ daki çekirdekler) yg-iş görene haritasından çakıştığı seçimleri silin. Grainyhead sadece epitel çekirdeklerini lekelerken, DAPI tüm çekirdekleri lekeler. Özetlenen her seçimin yalnızca bir çekirdek içerdiğini kontrol edin ve birden fazla çekirdeğe sahip seçimleri silin veya düzenleyin. Düzenlenen YG listesini kaydedin.
- Fiji'deki analiz araçlarını kullanarak Yatırım Getirisi haritasındaki her epitel çekirdeğinin alanını ve tümleşik yoğunluğunu ölçün. Değerleri bir elektronik tablo programına dışa aktarın.
- Dairesel seçim aracını kullanarak ortalama görüntü arka planını ölçün. DAPI görüntüsünün farklı alanlarında ki çekirdeklerle çakışmayan üç daire çizin. Arka plan görüntüsü parlaklığını oluşturmak için bir elektronik tablo programına her bir dairenin alanını ve tümleşik yoğunluğunu ekleyin.
- Her arka plan tümleşik yoğunluk değerini karşılık gelen alana bölerek her görüntü için birim alan başına ortalama arka plan hesaplayarak başlayın. Daha sonra birim alan başına ortalama arka plan elde etmek için görüntü için alan başına üç entegre yoğunluk ölçümleri ortalama.
- Ardından, her DAPI çekirdeğinin toplam arka planını birim alan başına ortalama arka plan ile çarparak hesaplayın. Ölçülen her çekirdek için normalleştirilmiş DAPI yoğunluğu, ölçülen tümleşik yoğunluğundan her çekirdeğin toplam arka planı çıkarılarak hesaplanabilir.
- Yaralanmamış epitel kontrolündeki normalleştirilmiş DAPI yoğunluk değerlerinin ortalamasını ala. Yaralanmamış epitel çekirdekleri daha önce 2C ploidi değerine sahip olduğu hesaplanmıştır ve deneysel koşullardan epitel çekirdekleri ploidi hesaplanması için bir referans olarak hizmet verebilir8.
- Her çekirdeğin normalleştirilmiş DAPI yoğunluğunu referans yaralanmamış epitel kontrolünden (2C) normalleştirilmiş değere bölerek her çekirdeğin ploidini hesaplayın ve sonra değeri normalleştirilmiş ploidi (C değeri) eşitlemek için 2 ile çarpın:
(Nükleer Entegre Yoğunluk - Arka Plan Nükleer Entegre Yoğunluk)/Ortalama Nükleer Entegre Yoğunluk (yaralanmamış epitel çekirdeği = 2C) x 2 = epitel nükleer ploidi (C) - Bir nokta çizimi olarak ploidy değerleri ile grafik çekirdekleri, histogram veya buna göre bir çubuk grafiğine gruplandı (yani, 2C [0.6-2.9C], 4C [3.0-5.9C], 8C [6.0-12.9C], 16C [13.0-24.9C], ve >32C [>25.0C] (Şekil 3F).
Sonuçlar
D. melanogaster'in yara kaynaklı poliploidizasyonu (WIP) incelemek için model olarak kullanması için ayrıntılı bir protokol sağlanmaktadır. Bu yara iyileştirici modeli WIP memeli ve diğer sinek modelleri üzerinde birçok avantaj sağlar. Poliploid, böcek pimi ile mekanik bir delinme ile kolayca indüklendi ve kısa bir süre içinde poliploid hücreler üretildi (2-3 dpi)(Şekil 1A, 1B)4. En büyük sorun epitel herhangi bir tedirginlik olmadan bozulmamış karın dokusunun diseksiyon olduğunu. D. melanogaster epitel kolayca yanlışlıkla çarptı veya keskin diseksiyon araçları ile çizik. Bu nedenle, bu protokolün adımları kullanım ve analiz önce uygulanmalıdır.
İlk olarak, yaralanma ventral kadın karın ile sınırlıydı, hangi görüntüleme için ideal büyük, düz opak doku alanı sağlar. Delinme yaraları, ventral orta hat sternitlerinin her iki tarafında bulunan ve tergit (T) segmentleri T4-T5(Şekil 1A-C)arasında hedeflenen pleürepitelin içinde meydana geldi. Bu yara yerleşimi diseksiyon tarafından kesintiye uğramayan geniş bir görünür alan sağlar. Zorlu adımlar karın yay makas kesme ve sabitleme adımları(Şekil 1D, 1E)içerir. Yay kesme adımı, karınlar doku hareketini azaltmak için Grace'in çözeltisinin (~30 μL) azaltılmış hacminde kesildiğinde en iyi şekilde işe yaradı. Diseksiyon plakası üzerinde açık pimlemek için karın sırt kapaklarında yeterli alan sağlamak için dorsal orta hat boyunca iyi merkezli bir kesim gerekliydi(Şekil 1C). Karın hafifçe aşırı güç olmadan dört köşeye sabitlenmelidir (Şekil 1E). Çok sert bir iğne itme karın dokusu nu bozar ve hatta diseksiyon plaka içine doku itebilir. Bu durumda, doku atılmalıdır. Karın dokusu sabitlendikten sonra, immünororesans boyama tamamlanana ve karınlar görüntüleme için cam bir kapak kaymasıüzerine monte edilene kadar diseksiyon plakası üzerinde kalmıştır (Şekil 1F).
Yara iyileşmesi endorepsiyon ve hücrefüzyonu4 bağlıdır oluşturmak için sürekli bir epitel levha gerektirir,16. Hücre-hücre bağlantılarını etiketleyen septat bağlantı proteini FasIII, hazırlık sırasında herhangi bir işlem tedirginliği oluşup oluşmadığına dair bir gösterge sağlamıştır(Şekil 1G, 1H). Yara bölgesini tedirgin eden büyük çizikler (lekesiz alan) olan karınlar atılmalı ve daha fazla analiz için kullanılmamalıdır (Şekil 1H).
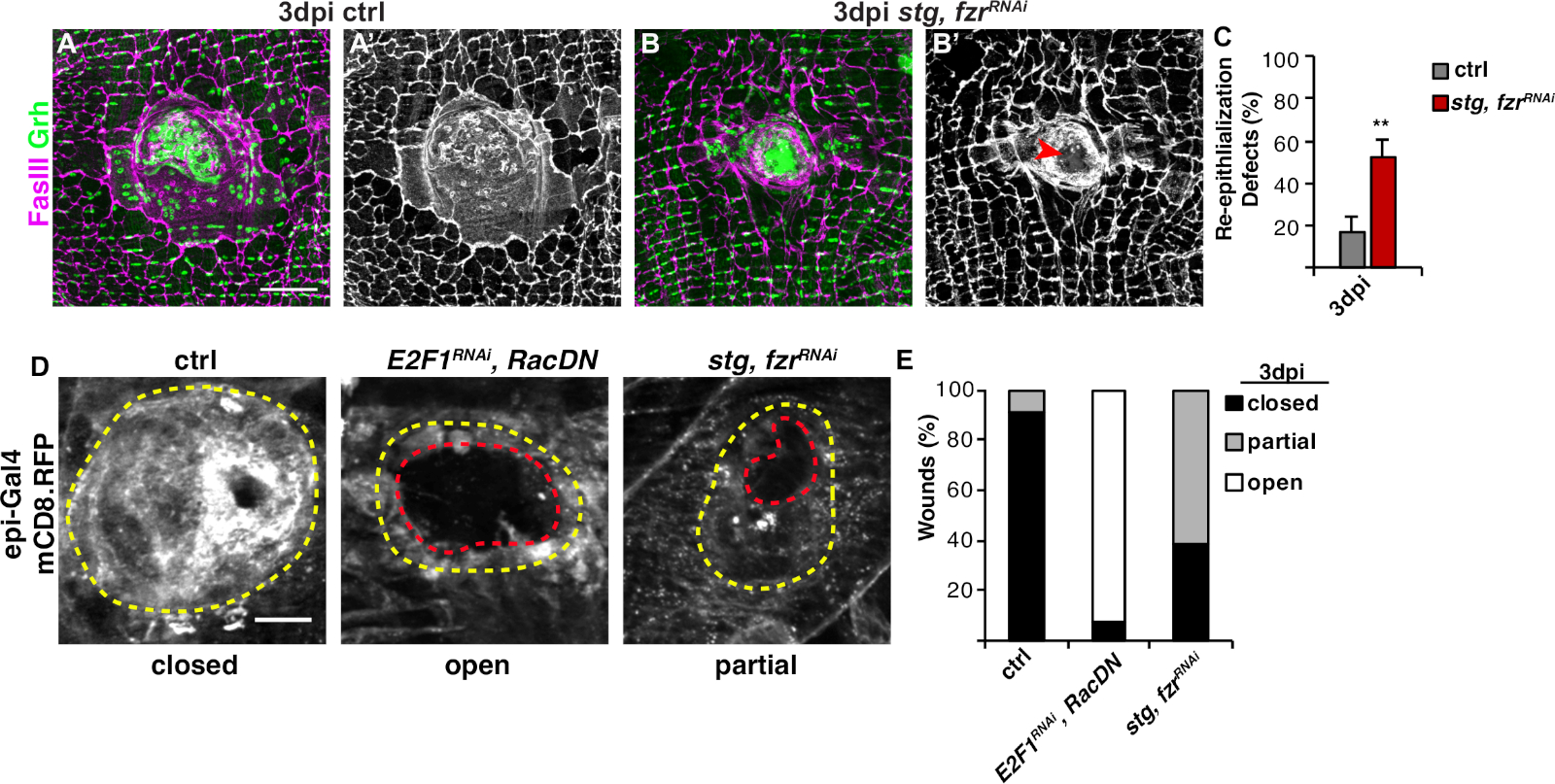
Bir sonraki adım WIP herhangi bir hata için bozulmamış örnekleri analiz etmek oldu. Bu protokol, WIP yanıtının farklı yönlerini algılamak için farklı tahliller içerir(Şekil 2). Merkezi, büyük, çok çekirdekli bir hücre yara yarasını kapladığında yara onarımı tamamlanmıştır(Şekil 3A). Burada hücre füzyonu FasIII/Grh için boyama ve FasIII özetlenen alan4'tekapsanan Grh+ epitel çekirdeklerinin sayısının ölçülmesi ile saptanmıştır. Epitel tabakasında >10 μm boşluklar gözlendiğinde yara kapanması veya yeniden epitelizasyon da kusurlar tespit edilmiştir(Şekil 3B, kırmızı ok). Bu durum, örneğin, WIP stg ifadesi ile mitotik döngüsünün aktivasyonu tarafından inhibe edildiğinde, fzrRNAi, son zamanlarda rapor16. Bu genetik durumda, yaraların %52'si yara kabuğunun üzerinde sürekli epitel tabakası oluşturamamıştır(Şekil 3B, 3C).
Bu modelde yara onarımını ölçmek için bir diğer yöntem de UAS-mCD8-ChRFP4 'in epi-Gal4 ekspresyonu ile epitel zarını görselleştirmektir (Şekil 2, Şekil 3D). Kontrolde, epitel yaraların% 91 tamamen 3 dpi tarafından kapalı, ancak endorepyonu engelleyerek WIP inhibe (E2f1RNAi) ve hücre füzyonu (RacDN) aynı anda, daha önce bildirildiği gibi, epitel yaraların% 92 tamamen açık kalmasına neden(Şekil 3D, 3E)8,16. Stg ekspresyonu ile mitotik hücre döngüsünün aktivasyonu, fzrRNAi de epitel yarası kapatma defekti ile sonuçlandı. Ancak epitel hücre zarının görselleştirilmesiyle re-epitelizasyon defektinin boyutu belirlenebilir. WIP mutantı (E2f1RNAi, RacDN) sinek yaraları stg daha açık, fzrRNAi yaralar(Şekil 3D, kesik kırmızı anahat)16. Bu membran yara iyileşme satozu yara onarım defekti ölçüde daha fazla bilgi sağladı. Sonuç olarak, yeniden epitelizasyon kusurları tamamen açık, kısmen kapalı (yani >10 μm boşluklar) veya tamamen kapalı olarak gruplandırılabilir(Şekil 3D, 3E).
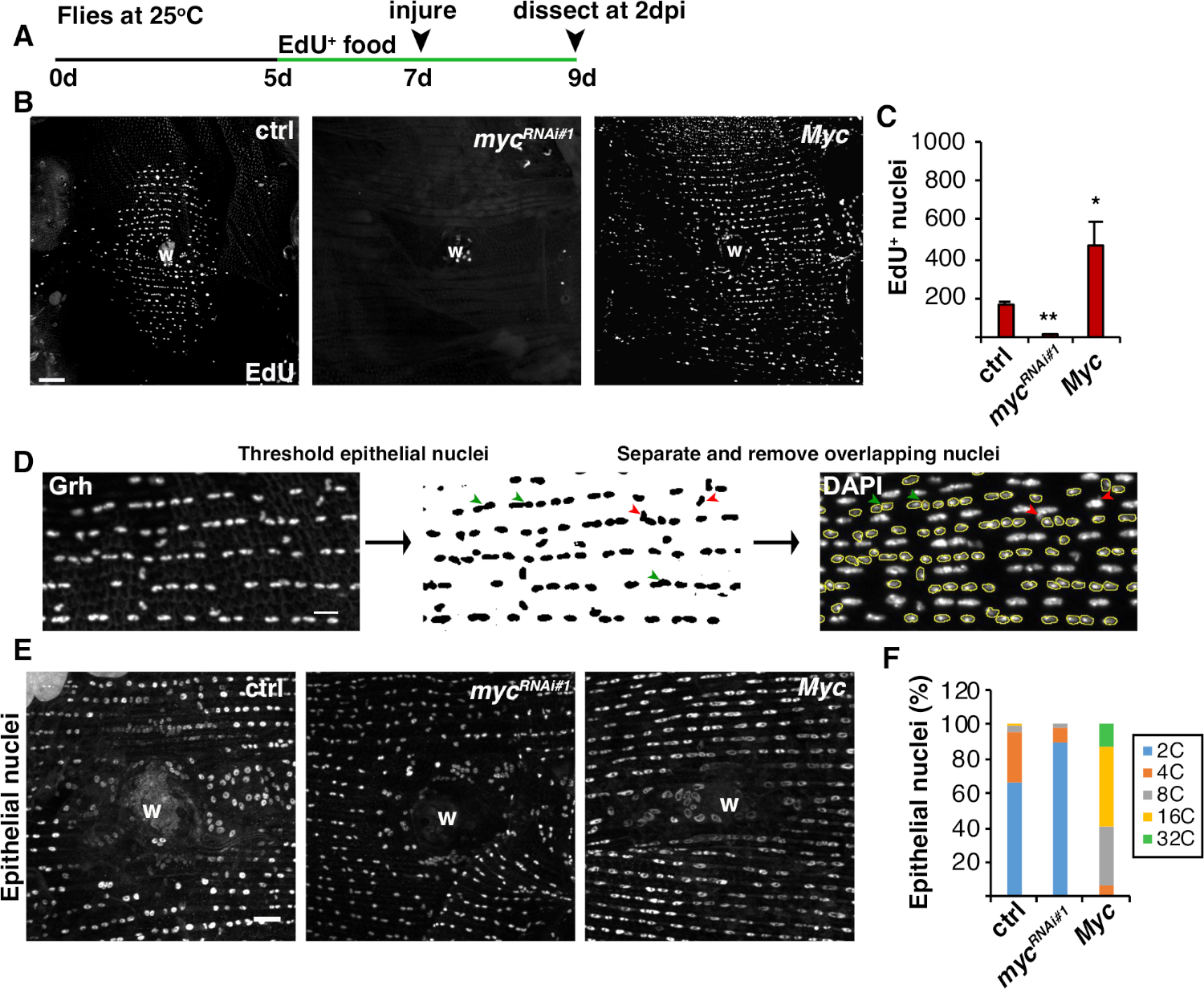
Hücre füzyonuna ek olarak, epitel hücreleri endorepyon, nükleer DNA içeriğini iki katına eksik bir hücre döngüsü ile boyutu büyür. Endorepsiyon, hem hücre döngüsü aktivitesi hem de direkt nükleer DNA ploidi ölçümleri ile titreşti (Şekil 2 ve Şekil 4). Burada, hücre döngüsü aktivitesi timidin analog, EdU (Şekil 4A, 4B)dahil edilerek tespit edilmiştir. D. melanogaster epitel hücreleri endocycle girmek için bulundu, Bir müdahale M faz ı olmadan S ve G fazları arasında salınım eksik bir hücre döngüsü4,12. Erişkin D. melanogaster diyeti yaralanmadan önce EdU+ gıda ile desteklendi ve sinekler 2 dpi diseksiyona kadar EdU+ diyette muhafaza edildi(Şekil 4A). EdU daha sonra üreticinin Click-iT protokolü kullanılarak algılandı. Bu EdU tonu, bir yaraya yanıt olarak S fazına girmek için nerede, ne zaman ve kaç çekirdeğin tetiklenerek tetikleniyi belirlemek için kullanılmıştır. Gal4/ UAS sistemini kullanarak, son zamanlarda myc epitelyal spesifik ekspresyonu ya blok olabilir bulundu(mycRNAi) veya şiddetlendirmek (Myc aşırı ekspresyon) S fazgirmek için epitel hücrelerinin yetkinliğini. Sonuç olarak, Myc postmitotik hücrelerde endoreplik indüklemek için yeterli olduğu gösterilmiştir, yaralanma olmadan bile16,22.
Daha sonra, epitel ploidi doğrudan nükleer DNA içeriği ölçerek belirlendi. Epitel çekirdekleri epitelyal spesifik belirteç için immünororesans boyama ile tespit edildi, Grh (Şekil 4D). Fiji görüntüleme yazılımında epitel çekirdekleri sistematik olarak tanımlandı ve Grh nükleer lekesi kullanılarak eşiklendi. Çekirdekler daha sonra ayrıldı ve ROI yığınları DAPI görüntü(Şekil 4D, yeşil ok) toplamı üzerine kaplanmış. Seçili çekirdeklerin Entegre Yoğunluğu ölçülmeden önce çakışan çekirdekler el ile silindi(Şekil 4D, kırmızı oklar). Bu yarı otomatik yöntem bir dağılım ve yaralanmamış ve onarılmış sinek abdominal epitel boyunca en çekirdeklerin ploidy ölçmek için izin verir8. Yakın zamanda bildirildiği gibi, yarayı çevreleyen epitel çekirdekleri 3 dpi 'de 3C'den fazla DNA içeriği olan%44poliploid çekirdekten oluşuyordu ( Şekil 4E, 4F)16. EdU sonuçlarından beklendiği gibi, myc'nin yıkılması endorepyonda önemli bir blota yol açtı, epitel çekirdeklerinin sadece %9'u poliploid olduğu için, Myc'in aşırı ekspresyonu yara bölgesi nin etrafında %100 poliploid epitel çekirdeği ile sonuçlandı (Şekil 4F)16. Epitel nükleer boyutu da görünür ya azaltılmış veya genişlemiş çekirdekleri mevcut ile myc ifade etkilenmiştir. Ancak, nükleer alan ploidi ve fizyolojik etkileri doğru bir ölçü değildir, hücre germe gibi faktörler de nükleer DNA içeriğini etkilemeden nükleer boyutu etkileyebilir çünkü20.

Şekil 1: Erişkin meyve sinek karın yaralanma, diseksiyon, ve doku montaj. (A) Erişkin karın yarası tadına göre diyagram. Tergite 4 (T4) de karın her iki tarafında sinekler yaralanmalıdır. (B) Yetişkin dişi meyve yara iyileşmesi (beyaz kutu) oluşan melanin kabuğu ile 3 dpi sinek. Ölçek çubuğu = 50 μm. (C) Diseksiyonlu yetişkin karın, dorsal görünüm, tergitler etiketli. Karınlar dorsal tarafın orta çizgisinden aşağı fileto edildi (beyaz kesikli çizgi). Ölçek çubuğu = 50 μm. (D) Sabitleme den önce parçalanmış ve filetolu yetişkin karınları. Ölçek çubuğu = 50 μm. (E) Bir diseksiyon plakası üzerinde sabitlenmiş yetişkin karın. Dorsal tarafta karın dört köşesinin her birine bir iğne yerleştirildi. Doku hafifçe açıldı ama gerildi değil, yırtılma önlemek için. (F) Yetişkin karınlar monte edilmiş ve cam kaydırağa doğru doğru bakan karın iç coverslip ve cuticle bakan bir cam kapak üzerine yerleştirilir. (G) Hiçbir işleme pertürbasyon ve merkezi bir syncytium (kesik sarı çizgi) ile bozulmamış yara alanı FasIII boyama. Ölçek çubuğu = 50 μm. (H) Senkronu bozan lekesiz FasIII bölgesine sahip çizik bir yara alanının (*) görüntüsü. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: WIP analizi iş akışı. Akış şeması, bu çalışmada açıklanan üç tahlilve wip yanıtını algılamak ve ölçmek için üst üste gelen ve farklı adımları betimlener. EdU titreci hücre döngüsü aktivitesini ölçer (mavi kutular), ploidi ve yeniden epitelizasyon Grh/FasIII immünboyama (yeşil kutular) tarafından tespit edilir ve membran RFP ekspresyonu epitel yara kapatma (pembe kutular) ölçüde ölçümü sağlar. Ortak adımlar gri kutularve D. melanogaster suşu genotipler yukarıda listelenmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: WIP sırasında yeniden epitelizasyonu algılama yöntemleri. WIP genetik olarak inhibe edildiğinde yeniden epitelizasyon tedirgin edildi. Kontrol immünofloresan görüntüleri (A) ve stg, fzrRNAi (B) at 3 dpi. Epitel çekirdekleri ve septate kavşakları sırasıyla Grh (yeşil) ve FasIII (macenta) ile boyandı. (A' ve B') FasIII boyama tek başına yeniden epitelizasyon stg (kırmızı ok) bozulmuş olduğunu gösterdi, fzrRNAi epitel. Ölçek çubuğu = 50 μm. (C) Yeniden epitelizasyon kusurlarının sayısallaştırılması (%) at 3 dpi (gri): kontrol (n = 8), stg, fzrRNAi (n = 6). Hata çubukları standart hatayı gösterir; istatistiksel anlamlılık Öğrencinin T-testi, **P < 0.01 ile ölçüldü. Yara onarımı sırasında yeniden epitelizasyon da epi-Gal4, UAS-mCD8-RFP kullanılarak bir membran bağlantılı RFP ekspresyonu ile tespit edilebilir. (D) Kontrol İmmünofloresan görüntüleri, E2F1RNAi, RacDN, ve stg, fzrRNAi at 3 dpi. Ölçek çubuğu = 20 μm. Yara kabuğu (sarı anahat) ve açık epitel yarası alanı (kırmızı anahat). (E) Ctrl (n = 11), E2F1RNAi, RacDN (n = 13) ve stg, fzrRNAi (n = 13) içinde yara kapanmasının sayısallaştırılması. Grendler ve ark.16'danuyarlanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: WIP sırasında endoreplikasyonu algılama yöntemleri. (A) EdU suresinin zaman çizelgesi: erişkin Drosophila yaralanmadan 2 gün önce her gün 5 mM maya-EdU ile 75 μL beslendi ve 2 dpi'ye kadar devam etti. (B) 2 dpi epi-Gal4/ UAS sistemi ile ifade edilen sinek suşlarında EdU etiketinin immünororesan görüntüleri. Yara kabuğu (W). Ölçek çubuğu = 50 μm. (C) Ortalama edU+ epitel çekirdeği sayısı 2 dpi: ctrl (n = 37), mycRNAi#1 (n = 10) ve Myc (n = 8). Hata çubukları standart hatayı gösterir; istatistiksel anlamlılık Öğrencinin T-testi, *P < 0.05, **P < 0.01 ile ölçüldü. (D) Epitel nükleer ploidinin saptanması ve ölçülmesi şeması. Epitel çekirdekleri Fiji'deki anti-Grh lekesi ile tanımlanmış ve eşiklenmiştir. Üst üste binen epitel çekirdekleri ayrılmış (yeşil ok uçları) veya kaldırıldı (kırmızı ok uçları) nonepithelial çekirdekleri tarafından overlaid eğer. İlgili DAPI lekeli çekirdek görüntüsünün entegre yoğunluğu ve nükleer alanı ölçüldü. (E) Epitel nükleer boyutu (Grh) 3 dpi myc ifade ile değiştirildi. (F) Epitel nükleer ploidi (%) 3 dpi: ctrl (n = 4), mycRNAi#1 (n = 6) ve Myc (n = 3). Grendler ve ark.16'danuyarlanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Sunulan nasıl diseksiyon ve yetişkin D. melanogaster abdominal epitel nasıl genlerin yara onarımı sırasında re-epitelizasyon ve endoreplikasyon değiştirerek WIP düzenleyen çalışma için kullanmak için ayrıntılı birprotokoldür 16. Bu yöntemi kullanarak, proto-onkogen Myc son WIP önemli bir düzenleyici olarak tespit edildi. Myc epitel hücreleri için endoreplicate sonrası yaralanma için gereklidir ve yetişkin sinek epitel ve aksesuar bezleri hem de endocycle için quiescent epitel hücreleri için yeterlidir16,22. Ayrıca stg ifadesi ile bir mitotik hücre döngüsü ne epitel hücreleri geçiş bulundu, fzrRNAi yara onarım zararlıdır. Bu yöntemi kullanarak devam eden çalışmalar, WIP sırasında yeniden epitelizasyon ve endorepsiyonasyonu düzenlemek için gerekli diğer genleri belirleyecek, poliploidin nasıl düzenlendiği ne kadar benzerlik ve farklılıkları ortaya koyarak çeşitli dokularda işlevlerini ortaya çıkaracaktır.
Bu model ve yöntem, mekanik bir delinme ile poliploid kolay indüksiyon ve poliploid hücrelerin gün içinde üretilen gerçeği de dahil olmak üzere benzersiz avantajlar sunuyoruz4. Doku diseksiyonu ve hazırlık protokolleri larva diseksiyon teknikleri dayanmaktadır23, ama yetişkin sinek karın daha sert ve bu nedenle kolayca tedirgin. Sonuç olarak, bu protokol WIP çalışması için bozulmamış bir doku izole etmek için uygulama ve hassasiyet gerektirir. Bir kez diseksiyon, ancak, epitel açıkça görülebilir ve kolayca görüntülenmiş, yara iyileşme sürecinin bir anlık verim. Bu yöntem yetişkin sinek epitel organizasyonu, hücre ve senkron boyutu ve hücrelerin ve bireysel çekirdeklerin ploidy bilgi zenginliği sağlar. Canlı görüntüleme henüz bozulmamış meyve sineği içinde opak cuticle nedeniyle mümkün olmasa da, bu protokol şu anda mevcut ex vivo kültür koşulları D. melanogaster kısa vadeli canlı görüntüleme çalışmaları gerçekleştirmek için kullanılan dahil adapte edilebilir24.
Gelecekte, bu model ilgi diğer hücre türlerinde Gal4/UAS sistemi ile gen ekspresyonu düzenleyerek hücre-hücre crosstalk ve WIP diğer hücre türlerinin katkısını incelemek için ideal olacaktır. Benzer sorular da genetik ve mutant arka planlar çeşitli kullanılarak cevaplanabilir. Diseksiyon yetişkin sinek karın kolayca yağ vücut ve oenositler, lateral kas lifleri, duyusal nöronlar, trakea ve makrofaj benzeri hemositler de dahil olmak üzere, bu yöntem kullanılarak kolayca görselleştirilebilir hücre tipleri çeşitli içerir. Buna ek olarak, bu model araştırmacılar nasıl fizyolojik değişkenler WIP etkisi araştırmak için izin verecektir, seks de dahil olmak üzere, diyet, enfeksiyon, yaş, ve çevresel stres. Protokol, yetişkin dişi sineği daha büyük boyutu nedeniyle kullanırken, WIP erkek meyve sineğinde de görülür (Gjelsvik ve Losick, yayınlanmamış). Poliploid hücrelerin yaşlanma ve yaşla ilişkili hastalık sırasında ortaya çıkan bulunmuştur memeli karaciğer, beyin, göz, ve kalp12. Meyve sinek modeli, insan hastalığına bağlı genler yüksek oranda korunduğu için araştırmacıların fizyolojik ve hastalık bağlamında poliploidizasyonu incelemelerini sağlayacaktır.
Açıklamalar
Hiçbiri.
Teşekkürler
Boston Koleji'nde, dr Eric Folker'a laboratuarının kamerasını ve görüntüleme için stereoskop mikroskop kurulumunu ve Boston College Imaging Core'daki Bret Judson'ı altyapı ve destek için kullandığı için teşekkür ederiz. Bloomington Drosophila Stock Center (NIH P40OD018537), Vienna Drosophila Resource Center ve Harvard Tıp Fakültesi'ndeki TRiP Center (NIH/NIGMS R01-GM084947) bu çalışmada kullanılan transgenik stokları temin ettiği için de sinek topluluk kaynaklarına teşekkür ederiz. Fare FasIII antikor NIH NICHD tarafından desteklenen Ve Iowa Üniversitesi, Biyoloji Bölümü, Iowa City, IA sürdürülen Gelişimsel Çalışmalar Hybridoma Bankası elde edildi. Bu yayında bildirilen araştırma, R35GM124691 Ödülü altında Ulusal Sağlık Enstitüleri Ulusal Tıp Bilimleri Enstitüsü tarafından desteklenmiştir. İçerik sadece yazarların sorumluluğundadır ve Ulusal Sağlık Enstitüleri'nin resmi görüşlerini temsil etmek zorunda değildir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 35 mm Petri dishes | Fisher Scientific | FB0875713 | For creating plates to dissect in |
| 50 mL Conical Centrifuge Tubes | Fisher Scientific | 14-432-22 | For preparing staining reagents in |
| AxioImager M2 with Apotome | Zeiss | NA | For imaging samples |
| Blowgun mini | Genesee Scientific | 54-104M | For anethesizing D. melanogaster strains |
| Bovine Serum Albumin, 30% | Sigma | A7284-500ML | For immuostaining |
| Carbon dioxide tank | various distributors | N/A | For anethesizing D. melanogaster strains |
| Click-iT EdU 594 Kit | Thermofisher | C10339 | For EdU assay |
| Coverslips | Thermofisher | 3406 | For mounting |
| DAPI | Sigma | D9542-10MG | For immuostaining |
| Dissecting Plates (use Sylgard 184 Sil Elastic Kit) | Ellsworth Adhesives | 184SIL | For creating plates to dissect in. Mix epoxy as directed, let dry overnight |
| Donkey anti-Rabbit IgG (H+L) Secondary Antibody, Alexa Fluor 488 conjugate | Thermofisher | A21206 | For secondary immuostaining |
| Donkey anti-Rabbit IgG (H+L) Secondary Antibody, Alexa Fluor 568 conjugate | Thermofisher | A10042 | For secondary immuostaining |
| Drosophila tubing and fittings | Genesee Scientific | 59-124C, 59-123, 59-140 | For anethesizing D. melanogaster strains |
| Dumont #5 Forceps | Fine Science Tools | 11252-20 | For dissecting |
| epi-Gal4 | Bloomington Drosophila Stock Center (b) | b38793 | Losick et al. Current Biology, 2013 |
| epi-Gal4, UAS-mCD8.RFP | Bloomington Drosophila Stock Center (b) | b38793, b27392 | Losick et al. Current Biology, 2013 |
| Excel | Microsoft | For performing ploidy calculations | |
| Fiji/ImageJ (image analysis software) | NIH | https://imagej.nih.gov/ij | For image analysis |
| Fly food | Archon Scientific | N/A | Corn Syrup/Soy food |
| Flystuff Flypad | Genesee Scientific | 59-114 | For anethesizing D. melanogaster strains |
| Glass dissecting dish | Fisher Scientific | 13-748B | For performing dissections in |
| Glass slides | Fisher Scientific | 12-518-104C | For mounting |
| Goat anti-Mouse IgG (H+L) Secondary Antibody, Alexa Fluor 488 conjugate | Thermofisher | A11001 | For secondary immuostaining |
| Goat anti-Mouse IgG (H+L) Secondary Antibody, Alexa Fluor 568 conjugate | Thermofisher | A11031 | For secondary immuostaining |
| Grace's Insect Medium, unsupplemented | Thermofisher | 11595030 | For dissecting in |
| Insect pins | Fine Science Tools | 26002-10 | For wounding and pinning fly abdomens flat |
| Mouse anti-Fasciclin III (Drosophila) Primary Antibody | Developmental Studies Hybridoma Bank | 7G10 | For immunostaining epithelial cell-cell junctions |
| Mouting media | Vector Laboratories | H-1000 | Anti-fade mounting media to prevent photo bleaching during imaging |
| Nail polish | Electron Microscopy Sciences | 72180 | For sealing slides |
| Ortibal shaker | Fisher Scientific | 02-217-988 | For immuostaining |
| Phosphate Buffered Saline, PH 7.4 | Sigma | P3813-10PAK | For staining |
| Pin holders | Fine Science Tools | 91606-07 | For wounding |
| Rabbit anti-Grainyhead Primary Antibody | N/A | N/A | For immunostaining epithelial nuclei. Protocol to make antibody can be found (Ref. #4 and 8) |
| Rabbit anti-RFP Primary Antibody | MBL | PM005 | For immunostaining mCD8-RFP fly epithelium |
| Stereomicroscope | Olympus | SZ51 | For dissecting and mounting fly tissue |
| Triton X-100 | Sigma | 10789704001 | For immuostaining |
| UAS-E2F RNAi, UAS-RacDN | VDRC (v) and Bloomington Drosophila Stock Center (b) | v108837, b6292 | Losick et al. Current Biology, 2013 |
| UAS-fzr RNAi, UAS-Stg | VDRC (v) and Bloomington Drosophila Stock Center (b) | v25550, b56562 | Grendler et al. Development, 2019 |
| UAS-Myc | Bloomington Drosophila Stock Center (b) | b9674 | Grendler et al. Development, 2019 |
| UAS-myc RNAi | Bloomington Drosophila Stock Center (b) | b36123 | Grendler et al. Development, 2019 |
| Vannas Spring Scissors | Fine Science Tools | 15000-00 | For dissecting |
Referanslar
- Abreu-Blanco, M. T., Verboon, J. M., Liu, R., Watts, J. J., Parkhurst, S. M. Drosophila embryos close epithelial wounds using a combination of cellular protrusions and an actomyosin purse string. Journal of Cell Science. 125 (Pt 24), 5984-5997 (2012).
- Wood, W., et al. Wound healing recapitulates morphogenesis in Drosophila embryos. Nature Cell Biology. 4 (11), 907-912 (2002).
- Galko, M. J., Krasnow, M. A. Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biology. 2 (8), E239 (2004).
- Losick, V. P., Fox, D. T., Spradling, A. C. Polyploidization and cell fusion contribute to wound healing in the adult Drosophila epithelium. Current Biology. 23 (22), 2224-2232 (2013).
- Wang, Y., et al. Integrin Adhesions Suppress Syncytium Formation in the Drosophila Larval Epidermis. Current Biology. 25 (17), 2215-2227 (2015).
- Ramos-Lewis, W., LaFever, K. S., Page-McCaw, A. A scar-like lesion is apparent in basement membrane after wound repair in vivo. Matrix Biology. 74, 101-120 (2018).
- Lee, C. W., Kwon, Y. C., Lee, Y., Park, M. Y., Choe, K. M. cdc37 is essential for JNK pathway activation and wound closure in Drosophila. Molecular Biology of the Cell. 30 (21), 2651-2658 (2019).
- Losick, V. P., Jun, A. S., Spradling, A. C. Wound-Induced Polyploidization: Regulation by Hippo and JNK Signaling and Conservation in Mammals. PLoS One. 11 (3), e0151251 (2016).
- Tamori, Y., Deng, W. M. Tissue repair through cell competition and compensatory cellular hypertrophy in postmitotic epithelia. Developmental Cell. 25 (4), 350-363 (2013).
- Xiang, J., et al. EGFR-dependent TOR-independent endocycles support Drosophila gut epithelial regeneration. Nature Communications. 8, 15125 (2017).
- Cohen, E., Allen, S. R., Sawyer, J. K., Fox, D. T. Fizzy-Related dictates A cell cycle switch during organ repair and tissue growth responses in the Drosophila hindgut. Elife. 7, e38327 (2018).
- Gjelsvik, K. J., Besen-McNally, R., Losick, V. P. Solving the Polyploid Mystery in Health and Disease. Trends in genetics: TIG. 35 (1), 6-14 (2019).
- Cao, J., et al. Tension Creates an Endoreplication Wavefront that Leads Regeneration of Epicardial Tissue. Developmental Cell. 42 (6), 600-615 (2017).
- Zhang, S., et al. The Polyploid State Plays a Tumor-Suppressive Role in the Liver. Developmental Cell. 44 (4), 447-459 (2018).
- Lazzeri, E., et al. Endocycle-related tubular cell hypertrophy and progenitor proliferation recover renal function after acute kidney injury. Nature Communications. 9 (1), 1344 (2018).
- Grendler, J., Lowgren, S., Mills, M., Losick, V. P. Wound-induced polyploidization is driven by Myc and supports tissue repair in the presence of DNA damage. Development. 146 (15), 173005 (2019).
- Mehrotra, S., Maqbool, S. B., Kolpakas, A., Murnen, K., Calvi, B. R. Endocycling cells do not apoptose in response to DNA rereplication genotoxic stress. Genes & Development. 22 (22), 3158-3171 (2008).
- Patterson, M., et al. Frequency of mononuclear diploid cardiomyocytes underlies natural variation in heart regeneration. Nature Genetics. 49 (9), 1346-1353 (2017).
- Gonzalez-Rosa, J. M., et al. Myocardial Polyploidization Creates a Barrier to Heart Regeneration in Zebrafish. Developmental Cell. 44 (4), 433-446 (2018).
- Webster, M., Witkin, K. L., Cohen-Fix, O. Sizing up the nucleus: nuclear shape, size and nuclear-envelope assembly. Journal of Cell Science. 122 (Pt 10), 1477-1486 (2009).
- Knouse, K. A., Lopez, K. E., Bachofner, M., Amon, A. Chromosome Segregation Fidelity in Epithelia Requires Tissue Architecture. Cell. 175 (1), 200-211 (2018).
- Box, A. M., et al. Endocycles support tissue growth and regeneration of the adult Drosophila accessory gland. bioRxiv. , 719013 (2019).
- Karim, M. R., Moore, A. W. Morphological analysis of Drosophila larval peripheral sensory neuron dendrites and axons using genetic mosaics. Journal of Visualized Experiments. (57), e3111 (2011).
- Dai, W., Montell, D. J. Live Imaging of Border Cell Migration in Drosophila. Methods in Molecular Biology. 1407, 153-168 (2016).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır