
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Kafatası ve Deri Temizleme ile Uyanık Zebra Balığı Larvaları ve Yavrularında Tam Aktif Beyin Dokusunun In vivo Görüntülemesi
Bu Makalede
Özet
Burada zebra balığı embriyonik beynini larva ve çocuk evrelerine kadar in vivo olarak görüntüleyen bir yöntem sunuyoruz. Elektrofizyolojik yaklaşımlardan uyarlanan bu mikroinvaziv prosedür, olgun nöronun hücresel ve hücre altı ayrıntılarına erişim sağlar ve beyin fonksiyonu ve ilaç müdahalesini karakterize etmek için optogenetik ve nörofarmakolojik çalışmalarla birleştirilebilir.
Özet
Beyin gelişimi ve olgunlaşma sırasında meydana gelen kısa ömürlü değişiklikleri anlamak, hücresel ve hücre altı çözünürlükte uzay ve zamanda ayrıntılı yüksek çözünürlüklü görüntüleme gerektirir. Moleküler ve görüntüleme teknolojilerindeki gelişmeler, şeffaf zebra balığı embriyosunda beyin gelişiminin hücresel ve moleküler mekanizmaları hakkında çok sayıda ayrıntılı içgörü elde etmemizi sağladı. Son zamanlarda, döllenmeden birkaç hafta sonra daha sonraki larva aşamalarında meydana gelen nöronal bağlantının iyileştirilmesi süreçleri, örneğin sosyal davranış, karar verme veya motivasyon odaklı davranışların kontrolü, araştırma odağına taşınmıştır. Bu aşamalarda, zebra balığı derisinin pigmentasyonu beyin dokusuna ışık penetrasyonunu engeller ve embriyonik aşamalar için çözümler, örneğin pigmentasyonun farmakolojik inhibisyonu artık mümkün değildir.
Bu nedenle, uyanık zebra balıklarının beynine mikroskopi erişimi için elektrofizyolojik yaklaşımlardan elde edilen minimal invaziv bir cerrahi çözüm sağlanır. Teleostlarda, cilt ve yumuşak kafatası kıkırdağı, bu katmanların mikro soyulmasıyla, alttaki nöronların ve aksonal yolların hasar görmeden açığa çıkarılmasıyla dikkatlice çıkarılabilir. Bu, sinaptik yapılar ve moleküler içerikleri de dahil olmak üzere nöronal morfolojinin kaydedilip Ca2+ geçici veya hücre içi taşıma olayları gibi fizyolojik değişikliklerin gözlemlenmesine izin verir. Ayrıca bu süreçlerin farmakolojik inhibisyon veya optogenetik manipülasyon ile sorgulanması mümkündür. Bu beyin maruziyeti yaklaşımı, nöronlardaki yapısal ve fizyolojik değişikliklerin yanı sıra bu olayların canlı beyin dokusunda dakika veya saat aralığında korelasyonu ve birbirine bağımlılıkları hakkında bilgi sağlar. Teknik, şimdiye kadar test edilen en son gelişim aşaması olan döllenmeden 30 güne kadar zebra balığı larvalarının in vivo beyin görüntülemesi için uygundur. Bu nedenle, sinaptik arıtma ve ölçekleme, aksonal ve dendritik taşıma, sitoskeletal kargonun sinaptik hedeflemesi veya yerel aktiviteye bağımlı ifade gibi önemli sorulara erişim sağlar. Bu nedenle, bu montaj ve görüntüleme yaklaşımı için geniş bir kullanım beklenebilir.
Giriş
Son yıllarda, zebra balığı (Danio rerio) embriyonik ve larva gelişimsel çalışmalar için en popüler omurgalı model organizmalardan biri olarak evrimleşmiştir. Zebra balığı dişilerinin büyük doğurganlığı, embriyonun hızlı ex utero gelişimi ve erken embriyonik gelişim aşamalarında şeffaflığı ile birleştiğinde, zebra balığını gelişimsel soruları ele almak için güçlü bir model organizma haline getiren sadece birkaç temel faktördür1. Moleküler genetik teknolojilerdeki gelişmeler, yüksek çözünürlüklü in vivo görüntüleme çalışmaları ile birlikte, gelişimsel süreçlerin altında kalan hücre biyolojik mekanizmalarının ele alınmasına izin verilir2. Özellikle, nöronal farklılaşma, fizyoloji, bağlantı ve işlev alanında zebra balığı, moleküler dinamiklerin, beyin fonksiyonlarının ve organizma davranışlarının benzeri görülmemiş ayrıntılarla etkileşimine ışık tutmaktadır.
Bununla birlikte, bu çalışmaların çoğu, sinir sistemi dokusunun şeffaflığı giderek kaybolduğundan, gelişimin ilk haftasında embriyonik ve erken larva aşamaları ile sınırlıdır. Bu aşamalarda, kafatası farklılaşması ve pigmentasyon ile korunan yüksek çözünürlüklü mikroskopi yaklaşımları ile beyin dokusunun erişimi engellenir3.
Bu nedenle, nöronal farklılaşma, olgunlaşma ve nöronal bağlantının iyileştirilmesi veya sinaptik ölçekleme gibi plastisite ile ilgili temel soruların incelenmesi zordur. Bu hücresel süreçler, örneğin sosyal davranış, karar verme veya motivasyona dayalı davranış gibi hücresel mekanizmaları tanımlamak için önemlidir, zebra balığının birkaç haftalık larvalar üzerinde yaptığı araştırmaların son zamanlarda davranışsal çalışmalara dayanan temel bulgulara katkıda bulunduğu alanlar4.
Zebra balığı larvalarında pigmentasyonu birkaç hafta boyunca inhibe etmek için farmakolojik yaklaşımlar zar zor uygulanabilir veya hatta zararlı etkilere neden olabilir5,6,7,8. Casper9 veya kristal10gibi spesifik pigmentasyon kusurlarına sahip çift veya üçlü mutant suşları, çok değerli araçlar haline gelmiştir, ancak üremede zahmetlidir, az sayıda yavru sağlar ve aşırı akraba evliliği nedeniyle genetik malformasyonların birikmesi tehlikesi oluşturur.
Burada, herhangi bir zebra balığı türü için geçerli olan alternatif olarak minimal bir invaziv prosedür sağlanır. Bu prosedür, canlı ve uyanık zebra balığı larvalarında nöronal aktiviteyi kaydetmek için elektrofizyolojik çalışmalardan uyarlanmıştır. Teleostlarda, deri ve yumuşak kafatası kıkırdağı, beyin vaskülatıyla sıkıca iç içe olmadıkları için bu katmanların mikro soyulmasıyla dikkatlice çıkarılabilir. Bu, nöronlar ve aksonal yollar içeren beyin dokusunun hasar görmeden açığa çıkıp sinaptik yapılar ve moleküler içerikleri de dahil olmak üzere nöronal morfolojinin kaydedillenmesine izin verir, bu da Ca2+ geçicileri veya hücre içi taşıma olayları gibi fizyolojik değişikliklerin birkaç saate kadar gözlemini içerir. Ayrıca, tanımlayıcı nitelemelerin ötesinde, beyin dokusuna doğrudan erişim, nörofarmakolojik madde yönetimi ve optogenetik yaklaşımlar yoluyla olgun nöronal fonksiyonların sorgulanmasını sağlar. Bu nedenle, bu beyin maruziyet stratejisi kullanılarak yavru zebra balığı beyninde gerçek yapı fonksiyon ilişkileri ortaya çıkarılabilir.
Protokol
Burada açıklanan tüm hayvan çalışmaları yasal düzenlemelere uygundur (AB Direktifi 2010/63). Balıkların bakımı ve elleçlenimi yerel yetkililer ve Technische Universität Braunschweig hayvan refahı temsilcisi tarafından onaylanmıştır.
1. Yapay serebro omurilik sıvısının (ACSF), düşük erime agarose ve keskin cam iğnelerin hazırlanması
- Listelenen kimyasalları damıtılmış suda aşağıdaki konsantrasyonlarda eriterek ACSF'yi hazırlayın. 134 mM NaCl (58,44 g/mol), 2,9 mM KCl (74,55 g/mol), 2,1 mM CaCl2 (110,99 g/mol), 1,9 mM 2 mM MgCl2 6x H2O (203.3 g/mol), 10 mM HEPES (238.31 g/mol) ve 10 mM d-Glikoz (180.16 g/mol).
NOT: MgCl2,CaCl2ve KCl için, 1 M stok çözeltileri tuzdan arındırılan steril suda hazırlanır ve daha sonra taze ACSF hazırlamak için 4 °C'de saklanır. Glikoz, HEPES ve NaCl, taze ACSF çözeltisinde katı bileşikler olarak çözülür. Kimyasalları çözmek için üreticinin talimatlarına uyun. - ACSF'nin pH'ını 10 M NaOH ile 7,8'e ayarlayın. ACSF'nin hazırlanması, beyin omurilik sıvısının yerini alması ve nöronların tamamen işlevsel olması için gerekli fizyolojik koşulları koruduğu için kimyasalların hassas bir şekilde ölçülmesini ve pH'ın ince ayarını gerektirir, aksi takdirde beyin yanlış beslenmesine ve nöronal ölüme neden olabilir.
- Yeni hazırlanan ACSF'i en fazla 4 hafta boyunca 4 °C'de saklayın. Çalışma koşulları için, 25-28 ° C'de gün / deney ve ön ısıtma için gerekli ACSF hacmini aliquot (ve isteğe bağlı olarak oksijenlendir, adım 2.5)
NOT: Yeni hazırlanmış ASCF 1 gün boyunca iyidir. Birkaç gün boyunca kullanmayı planlıyorsanız, ACSF'nin steril filtrelenmesi gerekir. - Larvaların daha sonra anestezisi için, damıtılmış suda 50 mM'lik bir d-Tubocurarine stok çözeltisi hazırlayın ve çözeltiyi -20 °C'de 100 μL aliquots olarak dondurucuda ihtiyaç duyulana kadar saklayın.
- Balıkları gömmek için, 50 mL ACSF'de 1,25 g LM-agarose(Malzeme Masası)eriterek% 2,5 düşük erime (LM) agarose hazırlayın ve agarose tamamen çözünene kadar kaynatın.
NOT: Alternatif olarak, deneysel kuruluma bağlı olarak daha yüksek veya daha düşük LM-agarose konsantrasyonları kullanılabilir. Bununla birlikte, agarose çok yumuşaksa, kafatasını açarken balığı yerinde tutamaz. - Katılaşmayı önlemek için agarose'u 37 ° C su banyosunda saklayın ve bu sıcaklık gömme sırasında larvalara da zarar vermez. Haşlanmış agarose su banyosunda 37 ° C'ye kadar soğutulduktan sonra, günün 10 μM çalışma konsantrasyonuna ulaşması için gereken aliquoted agarose'a gerekli miktarda d-Tubocurarine ekleyin. İleride kullanmak için, kirlenmeyi önlemek için sol agazozları 4 °C'de saklayın.
- Aşağıdaki ayarlara sahip bir mikropipette çekme kullanarak cam kılcal damarlardan keskin ve ince cam iğneler hazırlayın(Ek Şekil 1).
- Puller I, Kılcal Tip 1: Isı 1: 65.8; Isı 2: 55.1; 2 adımlı çekme
Puller II, Kılcal tip 2: Isı = 700; Fil = 4; Vel = 55; Del = 130; Pul = 55; 1 adımlı çekme.
NOT: Üniteler burada kullanılan her çektirici ve cam kılcal damar için sırasıyla özeldir (bkz. Malzeme Tablosu). Cam iğneleri hazırlamak için diğer kılcal damarlar ve çektiriciler de kullanılabilir. Ancak cam iğneler kafatası ile temas ederken kırılabilecekleri kadar ince olmamalıdır. Kılcal damar: uzunluk: 100 mm (4 inç); OD: 1,5 mm; Kimlik: 0.84 mm; filament: Evet
- Puller I, Kılcal Tip 1: Isı 1: 65.8; Isı 2: 55.1; 2 adımlı çekme
2. Larvaların anestezisi ve gömme hazırlıkları
- Deneye başlarken, plastik bir Pasteur pipet ile ihtiyaç duyulan hayvanları, Danieau (hala Danieau ile bir Petri kabında tutulan larvalar için) veya balık tesisinden su (7 dpf'den eski olan ve balık tesisinde tutulan larvalar için) ile dolu 90 mm çapında bir Petri kabına aktarın.
- 2 haftadan eski balıkları pipetlerken, pipet açıklığının balıkları aktarırken yaralanmayı önleyebilecek kadar büyük olduğundan emin olun. Bir ağ kullanmayın, çünkü özellikle genç larvalara fiziksel olarak zarar verir.
- Gıdaya ücretsiz erişim ve larvaların maksimum sağlık durumunu sağlamak ve stresi azaltmak için Petri kabında tutulan larvaların büyüklüğüne uygun rotifera veya artemia nauplii ekleyin.
- Gömme için, seçilen larvaları ACSF ile dolu 35 mm çapında bir Petri kabına aktarın. 10 μM'lik bir çalışma konsantrasyonuna / etkili doza ulaşmak için gerekli d-Tubocurarine hacmini ekleyin ve larvalar tamamen hareketsiz hale gelene kadar birkaç dakika bekleyin11.
NOT: Balık büyüdüğünde veya daha hızlı bir tam anestezi gerektiğinde (5 dk'nın altında), d-Tubocurarine konsantrasyonunu artırmak mümkündür (fareler için LD50 intravenöz olarak 0,13 mg /kg'dır). Curare ile aynı etkiye sahip olan ve beyni tamamen aktif tutan α-bungarotoxin (çalışma konsantrasyonu: 1 mg / mL) gibi farklı bir anestezik kullanmak damümkündür 13. İlgi konusu için tamamen aktif bir beyin gerekli değilse, ölümcül olmayan bir dozda (%0.02) Tricaine de larvaları tamamen uyuşturmak için bir seçenektir. Bununla birlikte, Tricaine sodyum kanallarını bloke eder, böylece beyin aktivitesini bozar14. - 35 mm çapındaki Petri kabının kapağını alarak montaj odasını hazırlayın, kapağı ters çevirin ve kapağın altına kare cam bir kapak kapağı (24 x 24 mm) yerleştirin. Bu adımların şematik açıklaması için Şekil 1'e (üst bölüm) bakın. Camın daha pürüzsüz yüzeyi, kafatası açma prosedürü sırasında larvaları içeren agarose bloğunun kaymasını önler.
- Uygun bir şişede (örneğin, 50 mL tüp, beher, Schott şişesi vb.) gün için gereken ACSF miktarını aliquot ve karbojen (%5 CO2, %95O2)ile oksijene alın. Beynin bütünlüğünü sağlamak için sadece morfoloji (örneğin floresan desenleri) ACSF gerekliyse ve hücrelerin ozmolarite etkilerinden olumsuz etkilenmediği gibi ACSF'nin oksijenlenmesine de gerek yoktur. Bu adımın sadece görüntüleme için tam beyin aktivitesi gerekli olduğunda gerçekleştirilmesi gerekir.
NOT: Ortamın optimum oksijen doygunluğu için karbojen tüpünün ucuna bir hava taşı ekleyin. Yeterince yüksek bir oksijen seviyesini garanti etmek için, aynı görüntüleme odasına gömülü larvaların sayısına ve yaşına bağlı olarak, görüntüleme odalarındaki ACSF'yi her 20-60 dakikada bir taze oksijenli ACSF ile değiştirmek gerekir (örneğin, her saat tek bir gömülü larva ACSF değişimi için yeterlidir. Paralel olarak gömülü 14 dpf'den daha eski altı larva için, her 20 dakikada bir ACSF alışverişi gereklidir) bu nedenle planlanan deneye göre gerekli miktarda oksijen doymuş ACSF planlayın.
3. Larvaların gömülmesi
- Tamamen uyuşturuldulanmış larvaları plastik bir Pasteur pipetle (adım 2.4) hazırlanan montaj odasına aktarın. Daha sonra, LM-agarose'un seyreltilmesini önlemek için fazla ortamı dikkatlice çıkarın. Aşağıdaki adımların tümü, yeterli büyütmeye sahip bir stereo mikroskop altında gerçekleştirilmelidir.
NOT: Montaj odasını eğmek ortamı tamamen çıkarmaya yardımcı olabilir. - Hayvanları kurumaya karşı korumak ve gereksiz stresi azaltmak için larvaların üzerine yeterince büyük bir LM-agarose damlası ekleyerek (larvaların büyüklüğüne bağlı olarak yaklaşık 1 mL) hemen bir sonraki adıma geçin.
- Agarose katılaştırılmadan önce larvaları pozisyona getirin. Larvaların sırt kısmının yukarı doğru yönlendirildiğinden emin olun. Ayrıca, larvaları agarose yüzeyine mümkün olduğunca yakın gömdökeğizden emin olun.
NOT: Aynı anda katıştırmak için planlanan larvaların büyüklüğüne ve sayısına bağlı olarak, agarose konsantrasyonunun ayarlanması mümkündür. Örneğin, 30 dpf eski 1-3 larva için% 1.8-% 2 LM-agarose konsantrasyonu önerilir. 7 dpf eski olan 1-4 larva için, % 2.5 LM-agarose kullanmak en uygun olanıdır, oysa 5-8 larva için% 2 daha uygundur. Tamamen aktif bir beyin gerekiyorsa, larvaları çalıştırmak için gereken süreyi azaltmak için aynı anda sadece üç balık gömmeniz önerilir. Genel olarak, larvalar yaşlandıkça veya aynı anda daha fazla larva gömülmesi planlandıkça daha düşük konsantrasyonların (% 1.8-% 2) kullanılması önerilir. - Görüntüler ters bir mikroskop kullanılarak kaydedilecekse, larvaları içeren agarose bloğunu küçük bir küboid şeklinde kırpın. Bu, larvaların daha sonra görüntüleme odasına aktarılması için önemlidir. Dik bir mikroskop kullanıyorsanız, bu tür bir kırpma gerekli değildir, çünkü montaj odası görüntüleme odası olarak da kullanılabilir. Şekil 1'de (üst bölüm), bu adımların şematik bir açıklamasını bulabilirsiniz.
4. Beyni açığa çıkarmak
NOT: Aşağıdaki adımların tümü larvaları gereksiz yere yaralamamak için büyük bir dikkatle yapılmalıdır. Deney için tamamen aktif bir beyin gerekiyorsa, geçen her saniye, balık hala agarose'a tamamen monte edilmişken ve oksijenli ACSF olmadan açık bir kafatasına sahipken, beynin oksijen eksikliğinden muzdarip olacağını ve ayrıca kuruyacağını unutmayın. Oksijen eksikliğinin etkileri daha da dramatik hale gelecektir, gömülü larvalar ne kadar yaşlı olursa. Bu nedenle iğne ile mekanik beyin hasarı uyandırmamak için ameliyatın sadece mümkün olan en kısa sürede değil, maksimum hassasiyetle yapılması önemlidir. Eğitildiğinde, 4.2-4.4 adımları balık başına 30 s'den fazla almamalıdır.
- Agarose katılaşır sağlamlaştırılmaz ameliyata başlayın. İlk olarak, kafaya ücretsiz erişim ve net bir çalışma alanı elde etmek için ilgi çekici beyin bölgesinin üzerindeki tüm fazla agaroseyu kesin. Başın sırt kısmı zaten agarose'dan dışarı yapışıyorsa, bu adımı atlayın.
- İlgi alanına bağlı olarak, ameliyatla başlamak için bir yer seçin. Cam iğneyi alın ve deriden küçük bir kesi yapın, ancak dokuya çok derinlemesine nüfuz etmeden. Bu, kaplama cildini soymak için başlangıç noktası olacaktır.
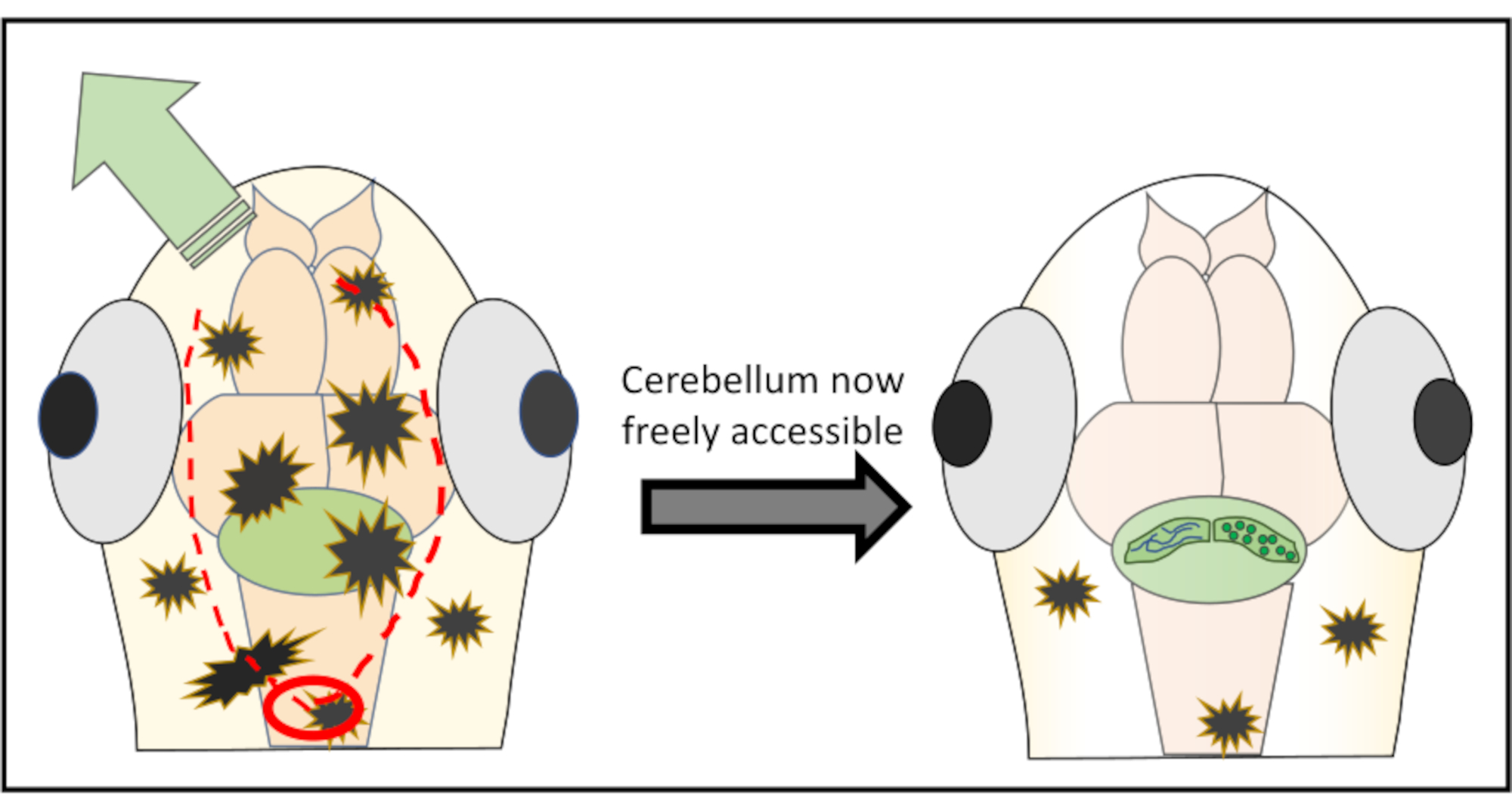
NOT: En iyi sonuçlar için, önemli yapılara zarar verme riskini azaltmak için asla doğrudan ilgi bölgesinin üzerinde başlamayın. Gerekirse, arka beyine arka plan başlatmak ve oradan cildin istenmeyen bölgesi soyulana kadar ileriye doğru çalışmak bile mümkündür. - İğneyi yüzeyin hemen altına zar zor hareket ettirerek çıkarmayı amaçlayan cilt kısmı boyunca çok küçük kesiklerle devam edin. Çoğu zaman beynin etrafında tamamen hareket etmek ve daire benzeri bir deri ve kafatası parçasını kesmek gerekli değildir, aksine sadece kafa boyunca iki kesi yapın ve sonra cildi bir veya diğer tarafa itin. Şekil 2, beyinciklere ücretsiz erişim elde etmek için en uygun kesme stratejisinin şematik bir temsilini göstermektedir.
NOT: Bu mikro cerrahi hassas bir prosedürdür ve büyük olasılıkla alttaki beyne zarar vermeden cildi mükemmel bir şekilde çıkarmak için biraz eğitime ihtiyaç duyacaktır. Ayrıca, ilgi çekici beyin bölgesi için en uygun kesme stratejisini bulmanız ve deney süresi boyunca onunla kalmanız önerilir. - Cildi tüm gömülü larvalardan çıkardıktan hemen sonra, istenmeyen cilt parçacıklarını ve kanı sel ve beyni tamamen aktif tutmak ve kurumasını önlemek için agarozun üzerine (oksijenli) ACSF dökerek devam edin.
NOT: Deney için sağlıklı bir beyin gerekiyorsa, aynı anda en fazla üç balık için gitmeniz önerilir. - Dik mikroskop kullanıyorsanız, doğrudan görüntüleme ile başlayın.
- Ters mikroskop kullanırken, küboid agarose bloğunun altına küçük bir spatula kaydırın (adım 3.4).
- Görüntüleme odasının altına küçük bir damla LM-agarose ekleyin (örneğin, cam tabanlı tabak) ve larvaları içeren agarose bloğunu hemen 180 ° spatula ile çevirin ve sıvı agarose damlası tutkal görevi görürken, görüntüleme odasının dibine hafifçe itin.
- Agarose katılaştığında, görüntüleme odasını (oksijenli) ACSF ile doldurun, ardından görüntülemeye başlayın. Şematik açıklama için Şekil 1'e (alt bölüm) bakın.
- Deney için tam beyin aktivitesi gerektiğinde, her zaman görüntüleme odasındaki ACSF'nin yeterince yüksek bir oksijen seviyesine sahip olduğundan emin olun. Bunu sağlamak için, ortamı mümkün olduğunda her 20-60 dakikada bir taze oksijenli ACSF ile dikkatlice değiştirin (balığın sayısına ve boyutuna, görüntüleme odasının büyüklüğüne ve yüzeyine ve görüntüleme süresine bağlı olarak).

Şekil 1: In vivo görüntüleme için açık kafatası zebra balıklarının adım adım hazırlanması için şematik prosedür. Farklı adımlar için çalışma yönergeleri grafiğin kendisinde bulunabilir. Florian Hetsch tarafından tasarlanan ve Paul Schramm tarafından uyarlanan grafik. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: İlgi çekici beyin bölgesinin üzerindeki deri ve kafatası parçalarını çıkarmak için yapılan mikro cerrahinin ayrıntılı şematik gösterimi. Kırmızı daire, ilk kesimin yapılması gereken noktayı işaretler. Kırmızı noktalı çizgi, beyinciklere zarar vermeden serbest erişim elde etmek için iğne ile birlikte kesmek için en uygun yolu açıklar. Yeşil ok, aşırı cilt ve kafatası parçalarının kolayca itilebileceği yönü işaretler. Tüm işlem sırasında beyin dokusuna asla nüfuz etmeyin. Cildi başarıyla soyduktan sonra, ilgi çekici beyin bölgesine (burada, beyincik) her türlü yüksek çözünürlüklü in vivo görüntüleme için serbestçe erişilebilir olacaktır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Sonuçlar
Şekil 3A,C transgenik çizginin 14 dpf larvasını göster Tg[-7.5Ca8:GFP]bz12[15] Kafatası hala sağlamken. Kaplama derisindeki pigment hücreleri başın her yerine dağılmıştır ve ilgi çekici bölgedeki floresan sinyaline müdahale etmektedir (burada, beyincik). Bu durumdaki larva ile beynin yüksek çözünürlüklü görüntülerini elde etmek mümkün değildir. ?...
Tartışmalar
Sunulan yöntem, beyin izolasyonuna veya zebra balığı larvalarının in vivo ortamlarında yüksek çözünürlüklü nöron görüntülerini kaydetmek için pigmentasyonu engelleyen farmasötiklerle tedavisine alternatif bir yaklaşım sağlar. Bu yöntemle kaydedilen görüntülerin kalitesi, ekilmiş beyinlerden gelen görüntülerle karşılaştırılabilir, ancak doğal koşullar altında.
Ayrıca, floresan yoğunluğunda bir kayıp önlenir, çünkü fiksatiflerle tedaviye ge...
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Özellikle timo Fritsch'e mükemmel hayvan bakımı için ve Hermann Döring, Mohamed Elsaey, Sol Pose-Méndez, Jakob von Trotha, Komali Valishetti ve Barbara Winter'a yardımsever destekleri için teşekkür ederiz. Köster laboratuvarının diğer tüm üyelerine de geri bildirimleri için minnettarız. Proje kısmen Alman Araştırma Vakfı (DFG, KO1949/7-2) 241961032 (RWK'ya) ve Bundesministerium für Bildung und Forschung (BMBF; Era-Net NEURON II CIPRESS projesi 01EW1520 to JCM) kabul edilmektedir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Calcium chloride | Roth | A119.1 | |
| Confocal Laser scanning microscope | Leica | TCS SP8 | |
| d-Glucose | Sigma | G8270-1KG | |
| d-Tubocurare | Sigma-Aldrich | T2379-100MG | |
| Glass Capillary type 1 | WPI | 1B150F-4 | |
| Glass Capillary type 2 | Harvard Apparatus | GC100F-10 | |
| Glass Coverslip | deltalab | D102424 | |
| HEPES | Roth | 9105.4 | |
| Hoechst 33342 | Invitrogen (Thermo Fischer) | H3570 | |
| Imaging chamber | Ibidi | 81156 | |
| Potassium chloride | Normapur | 26764298 | |
| LM-Agarose | Condalab | 8050.55 | |
| Magnesium chloride (Hexahydrate) | Roth | A537.4 | |
| Microscope Camera | Leica | DFC9000 GTC | |
| Needle-Puller type 1 | NARISHIGE | Model PC-10 | |
| Needle-Puller type 2 | Sutter Instruments | Model P-2000 | |
| Pasteur-Pipettes 3ml | A.Hartenstein | 20170718 | |
| Sodium chloride | Roth | P029.2 | |
| Sodium hydroxide | Normapur | 28244262 | |
| Tricain | Sigma-Aldrich | E10521-50G | |
| Waterbath | Phoenix Instrument | WB-12 | |
| 35 mm petri dish | Sarstedt | 833900 | |
| 90 mm petri dish | Sarstedt | 821473001 |
Referanslar
- Embryology. Zebrafish Development Available from: https://embryology.med.unsw.edu.au/embryology/index.php/Zebrafish_Development (2020)
- Sassen, W. A., Köster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. Dove Medical Press. 2015 (5), 151-163 (2015).
- Singh, A. P., Nüsslein-Volhard, C. Zebrafish stripes as a model for vertebrate colour pattern formation. Current Biology. 25 (2), 81-92 (2015).
- Kalueff, A. V., et al. Time to recognize zebrafish 'affective' behavior. Brill: Behaviour. 149 (10-12), 1019-1036 (2012).
- Karlsson, J., von Hofsten, J., Olsson, P. -. E. Generating transparent zebrafish: a refined method to improve detection of gene expression during embryonic development. Marine Biotechnology. 3, 522-527 (2001).
- Bohnsack, B. L., Gallina, D., Kahana, A. Phenothiourea sensitizes zebrafish cranial neural crest and extraocular muscle development to changes in retinoic acid and IGF signaling. PloS One. 6, 22991 (2011).
- Elsalini, O. A., Rohr, K. B. Phenylthiourea disrupts thyroid function in developing zebrafish. Development Genes and Evolution. 212, 593-598 (2003).
- Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. C. F., Segner, H. Thyroid disruption in zebrafish (Danio rerio) larvae: Different molecular response patterns lead to impaired eye development and visual functions. Aquatic Toxicology. 172, 44-55 (2016).
- White, R., et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell. 2, 183-189 (2008).
- Antinucci, P., Hindges, R. A crystal-clear zebrafish for in vivo imaging. Scientific Reports. 6, 29490 (2016).
- Burr, S. A., Leung, Y. L. Curare (d-Tubocurarine). Encyclopedia of Toxicology (3rd Edition). , 1088-1089 (2014).
- Gesler, H. M., Hoppe, J. 3,6-bis(3-diethylaminopropoxy) pyridazine bismethiodide, a long-acting neuromuscular blocking agent. The Journal of Pharmacology and Experimental Therapeutics. 118 (4), 395-406 (1956).
- Furman, B. . Alpha Bungarotxin. Reference Module in Biomedical Sciences. , (2018).
- Attili, S., Hughes, S. M. Anaesthetic tricaine acts preferentially on neural voltage-gated sodium channels and fails to block directly evoked muscle contraction. PLoS One. 9 (8), 103751 (2014).
- Namikawa, K., et al. Modeling neurodegenerative spinocerebellar ataxia type 13 in zebrafish using a Purkinje neuron specific tunable coexpression system. Journal of Neuroscience. 39 (20), 3948-3969 (2019).
- Hennig, M. Theoretical models of synaptic short term plasticity. Frontiers in Computational Neuroscience. 7 (45), (2013).
- Wang, Y., et al. Moesin1 and Ve-cadherin are required in endothelial cells during in vivo tubulogenesis. Development. 137, 3119-3128 (2010).
- Hobro, A., Smith, N. An evaluation of fixation methods: Spatial and compositional cellular changes observed by Raman imaging. Vibrational Spectroscopy. 91, 31-45 (2017).
- Knogler, L. D., Kist, A. M., Portugues, R. Motor context dominates output from purkinje cell functional regions during reflexive visuomotor behaviours. eLife. 8, 42138 (2019).
- Hsieh, J., Ulrich, B., Issa, F. A., Wan, J., Papazian, D. M. Rapid development of Purkinje cell excitability, functional cerebellar circuit, and afferent sensory input to cerebellum in zebrafish. Frontier in Neural Circuits. 8 (147), (2014).
- Scalise, K., Shimizu, T., Hibi, M., Sawtell, N. B. Responses of cerebellar Purkinje cells during fictive optomotor behavior in larval zebrafish. Journal of Neurophysiology. 116 (5), 2067-2080 (2016).
- Harmon, T. C., Magaram, U., McLean, D. L., Raman, I. M. Distinct responses of Purkinje neurons and roles of simple spikes during associative motor learning in larval zebrafish. eLife. 6, 22537 (2017).
- Zehendner, C. M., et al. Moderate hypoxia followed by reoxygenation results in blood-brain barrier breakdown via oxidative stress-dependent tight-junction protein disruption. PLoS One. 8 (12), 82823 (2013).
- Dhabhar, F. S. The short-term stress response - mother nature's mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Frontiers of Neuroendocrinology. 49, 175-192 (2018).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır