
Method Article
Genle Manipüle Edilmiş Fare Modelleri Geliştirmek için Embriyonik Kök Hücrelerde CRISPR / Cas9 Aracılı Yüksek Verimli Gen Hedefleme
Bu Makalede
Özet
Burada, embriyonik kök hücreler kullanarak, özellikle büyük DNA knock-in (KI) için genetiği değiştirilmiş fare modelleri geliştirmek için bir protokol sunuyoruz. Bu protokol, CRISPR / Cas9 genom düzenlemesi kullanılarak ayarlanır ve geleneksel homolog rekombinasyon aracılı doğrusallaştırılmış DNA hedefleme yöntemine kıyasla önemli ölçüde iyileştirilmiş KI verimliliği ile sonuçlanır.
Özet
CRISPR / Cas9 sistemi, döllenmiş zigotlar kullanarak doğrudan genom düzenleme ile genetiği değiştirilmiş farelerin geliştirilmesini mümkün kılmıştır. Bununla birlikte, küçük indel mutasyonunu indükleyerek gen nakavt fareleri geliştirmedeki etkinlik yeterli olsa da, büyük boyutlu DNA knock-in (KI) yapmak için embriyo genom düzenlemesinin etkinliği hala düşüktür. Bu nedenle, embriyolardaki doğrudan KI yönteminin aksine, embriyonik kök hücreler (ESC'ler) kullanılarak gen hedefleme ve ardından kimera fareleri geliştirmek için embriyo enjeksiyonu hala çeşitli avantajlara sahiptir (örneğin, in vitro yüksek verim hedefleme, çoklu alel manipülasyonu ve Cre ve floks gen manipülasyonu kısa sürede gerçekleştirilebilir). Ek olarak, BALB / c gibi in vitro olarak ele alınması zor embriyolara sahip suşlar da ESC hedeflemesi için kullanılabilir. Bu protokol, CRISPR / Cas9 aracılı genom düzenlemesini uygulayarak ESC'lerde büyük boyutlu DNA (birkaç kb) KI için optimize edilmiş yöntemi açıklar ve ardından gen manipüle edilmiş fare modelleri geliştirmek için kimera fareleri üretimi yapılır.
Giriş
Genetiği değiştirilmiş fareler üretmek ve fenotiplerini analiz etmek, spesifik gen fonksiyonlarını in vivo olarak ayrıntılı olarak anlamamızı sağlar. Yaşam bilimleri alanında gen modifiye hayvan modelleri kullanılarak çok sayıda önemli bulgu ortaya çıkarılmıştır. Ayrıca, CRISPR / Cas91 kullanan genom düzenleme teknolojisinin raporundan bu yana, genetiği değiştirilmiş fareler kullanılarak yapılan araştırmalar hızla birçok laboratuvara yayılmıştır 2,3. Fare zigotlarının CRISPR / Cas9 tarafından genom düzenlemesi, indel mutasyon odaklı gen nakavtı4, tek nükleotid replasmanı veya tek sarmallı oligonükleotidler (ssODN'ler) kullanılarak kısa peptit etiketi yerleştirme gibi kısa DNA modifikasyonu geliştirmek için kabul edilebilir bir verimlilik elde etmiştir5. Öte yandan, genom düzenleme yoluyla zigotlara büyük DNA fragmanlarının KI'sı, kısa boyutlu DNA modifikasyonu 6,7'ye kıyasla düşük verimlilikte kalmaktadır. Ek olarak, immünoloji gibi spesifik araştırma alanları için önemli bir suş olan BALB / c gibi fare suşlarını, zigot tabanlı genom düzenlemesi için kullanmak zordur, çünkü preimplantasyon embriyoları in vitro manipülasyona duyarlıdır.
Genetiği değiştirilmiş fare modelleri geliştirmenin bir başka yolu, embriyonik kök hücre (ESC) hedefleme tekniğini kullanmak ve ardından hala geleneksel bir yöntem olarak rutin olarak kullanılan kimera 8,9,10 üretmek için preimplantasyon embriyoya ESC enjeksiyonunu kullanmaktır. Geleneksel ESC hedefleme yöntemlerinde doğru KI-ESC klonları elde etmek için elde edilme oranı çok yüksek olmasa da, ESC hedefleme, özellikle uzun DNA KI için zigot genom düzenlemesine kıyasla bazı avantajlar sunmaktadır. Örneğin, zigot genomuna uzun DNA fragmanlarının (> birkaç kb) KI etkinliği daha az belirgindir 6,7 ve hayvan deneylerinin mevcut perspektifinde istenmeyen bir KI fare çizgisi bile geliştirmek için birçok zigota ihtiyaç vardır. Zigot genom düzenlemesinin aksine, ESC'lere uzun DNA hedeflemesi ve ardından kimera üretimi, zigot genom düzenlemesinden önemli ölçüde daha az embriyoya ihtiyaç duyar. Ayrıca, BALB / c'den preimplantasyon embriyoları in vitro manipülasyona duyarlı olsa da, ESC'leri diğer yetkili 129 veya F1 arka plan ESC'leri gibi in vitro 11'de muhafaza edilebilir ve işlenebilir, bu nedenle kimera üretimleri için geçerlidir. Bununla birlikte, bir hedefleme vektörü pozitif veya negatif seleksiyon için 5' ve 3' homolog kollar ve ilaca dirençli gen kasetleri içermesine rağmen, ESC'lerin geleneksel KI etkinliği genellikle yetersizdir, çünkü rastgele genomik entegrasyonun yüksek frekansı 8,10, Bu nedenle, kesin ESC hedefleme verimliliğine sahip gelişmiş bir yöntem gereklidir. Son zamanlarda, geleneksel hedefleme yöntemlerinden daha yüksek KI verimliliği elde etmek için CRISPR / Cas9 tabanlı genom düzenleme kullanan ayarlanmış bir ESC KI yöntemi bildirdik11. Burada tarif ettiğimiz yöntem, uzun DNA (> birkaç ila 10 kb) KI'yı ESC'lere ilaç seçimi olmadan rutin işler için kabul edilebilir verimlilikle sağlayan bu prosedüre dayanmaktadır; Böylece, vektör yapım prosedürü çok daha kolay olacak ve daha kısa bir süreye ihtiyaç duyacaktır veya hücre kültürü periyodu da önemli ölçüde kısalacaktır.
Protokol
Tüm fare deneyleri, Tokyo Üniversitesi (onay numarası PA17-63) ve Osaka Üniversitesi (onay numarası Biken-AP-H30-01) Kurumsal Hayvan Bakımı ve Kullanımı Komitesi tarafından onaylanmış ve kılavuzlarına ve ARRIVE kılavuzlarına (https://arriveguidelines.org) göre gerçekleştirilmiştir.
1. Vektör yapısını hedefleme
- Bir KI kasetini (CreERT burada, PCR için şablon DNA orijinal olarak ticari bir gen sentez şirketindendir) ve PCR ile 5 'veya 3' homoloji kollarının yaklaşık 1 kb'lik bir parçasını yükseltin. DNA klonlaması için (adım 1.3), her PCR primerinin 5' ucuna 15-mer örtüşme dizileri ekleyin. PCR DNA fragmanlarını agaroz jel elektroforezi ile saflaştırın, ardından bir DNA saflaştırma kiti kullanarak DNA fragmanının ekstraksiyonunu yapın.
- Klonlama için çoklu klonlama bölgesinde bir yerde benzersiz bir şekilde sindirilen uygun bir kısıtlama enzimi (leri) kullanarak omurga plazmidini (örneğin, pUC19 veya pBS) doğrusallaştırın. Ardından, sindirilmiş plazmidi bir DNA saflaştırma kiti kullanarak saflaştırın.
- Her PCR fragmanını (örneğin, 5'-homoloji kolu, 3'-homoloji kolu) ve KI dizisini, üreticinin talimatlarına göre bir DNA klonlama kiti kullanarak sindirilmiş plazmidin içine aynı anda klonlayın.
- Yetkin hücreleri, bir su banyosunda 1 dakika boyunca 42 ° C'de ısıtarak inşa edilmiş plazmidi (adım 1.3) kullanarak dönüştürün, ardından ampisilin veya kanamisin gibi uygun antibiyotik konsantrasyonunu içeren bir LB plakasına tohumlayın. Dirençli klon seçimleri için onları gece boyunca kültürleyin.
- 200 μL pipet ucu kullanarak birkaç (dört ila sekiz) ayrı koloni toplayın ve uçları uygun antibiyotikler (100 μg / mL ampisilin veya 20 μL / mL kanamisin) içeren 3 mL sıvı LB içine ve gece boyunca 37 ° C'de sallama ile kültüre aktarın.
- Plazmidi ertesi gün üreticinin talimatlarına göre endotoksin içermeyen dereceli ticari plazmid saflaştırma kiti kullanarak saflaştırın. Plazmiddeki klonlanmış diziyi Sanger dizilimi ile onaylayın. Plazmid konsantrasyonunu nükleaz içermeyen su kullanarak 1 μg DNA / μL'de ayarlayın. Plazmidi gen hedefleme vektörü olarak kullanın.
2. ESC için besleyici hücreler olarak fare embriyonik fibroblastının (MEF) hazırlanması
- 8-10 haftalık hamile ICR dişi fareleri (14.5 gün postkoitum) servikal çıkık ile feda edin. Sterilizasyon için% 70 (v / v) etanol kullanarak karnı iyice silin, orbital makas ve forseps kullanarak karnı kesin ve fetüs içeren uterusu kurtarın.
- Rahimi, 100 U/mL penisilin ve 100 μg/mL streptomisin içeren 10 mL fosfat sığır serumu (PBS) içeren 100 mm'lik bir kaba koyun. Plasentaları ve ekstraembriyonik dokuları orbital makas ve forseps kullanarak fetüslerden çıkarın. Orbital makas kullanarak fetüsleri kıyın ve kıyılmış fetal hücreleri içeren PBS çözeltisini 50 mL'lik konik bir tüpe aktarın.
- 4 °C'de 5 dakika boyunca 280 x g'de santrifüj yapın ve süpernatantı aspirasyonla atın. 10 mL% 0.25 (w / v) tripsin-EDTA çözeltisi ekleyin, pipetleme ile iyice karıştırın ve daha sonra tripsin sindirimi için bir su banyosu kullanarak 37 ° C'de 10 dakika boyunca inkübe edin.
- Enzimatik reaksiyonu durdurmak için 20 mL MEF ortamı (% 10 (v / v) fetal sığır serumu, 100 U / mL penisilin ve 100 μg / mL streptomisin içeren DMEM) ekleyin, nazik inversiyonla iyice karıştırın ve 4 ° C'de 5 dakika boyunca 280 x g'de santrifüj yapın. Süpernatantı aspirasyonla atın, 10 mL MEF ortamı ekleyin ve pipetleme ile iyice karıştırın.
- Hücre süspansiyonunu birkaç yeni 100 mm'lik tabakta tohumlayın (100 mm'lik bir tabak için 10 mL MEF ortamına sahip fetüs sayısıyla aynı sayıda yemek hazırlayın). Bağlanmamış hücreleri çıkarmak ve bağlı MEF'in büyümesine izin vermek için tohumlama sonrası ortamı 4 ila 5 saat arasında değiştirin. Tripsin kullanarak ~% 80 akıcılığa ulaştıklarında hücreleri geçirin.
- Çoğalan MEF'i içeren kültür kaplarını, tohumlamadan yaklaşık 12-15 gün sonra, bir X-ışını ışınlama cihazında MEF ortamına yerleştirin. Üreticinin talimatına göre hücre döngüsünü durdurmak için MEF'i 50 Gy X-ışını (toplam) ile maruz bırakın.
- 37 ° C'de 5 dakika boyunca 100 mm'lik bir çanak için 2 mL% 0.25 tripsin-EDTA ekleyerek ışınlanmış MEF'i tripsinize edin, ardından tripsin sindirimini durdurmak için 4 mL MEF ortamı ekleyin ve hücre süspansiyonunu yeni bir 50 mL tüpte toplayın.
- MEF'i 4 ° C'de 5 dakika boyunca 280 x g'de santrifüjleme yaparak taze bir MEF ortamı ile yıkayın, tripan mavisi boyamalı bir hücre sayacı kullanarak hücre sayısını sayın ve bunları hücre dondurucu bir ortamda 1,6 x 106 hücre / tüpte dondurun. Kullanıma kadar saklayın; hücreler birkaç yıl boyunca sıvı azotta stabil bir şekilde depolanabilir.
3. Cas9-RNP-DNA karışımı hazırlama
- TracrRNA ve crisprRNA'yı seyreltme tamponunda (her RNA konsantrasyonu 200 μM'de) pipetleme ile çözün. TracrRNA çözeltisini, crisprRNA çözeltisini ve seyreltme tamponunu (2: 2: 5'te hacim oranı), 10 dakika boyunca 95 ° C'de bir termal döngü kullanarak her RNA'yı hafifçe vurarak ve tavlayarak ve ardından sıcaklık 4 ° C'ye ulaşana kadar 1 ° C / dak basma döngüleri ile karıştırın.
NOT: gRNA dizisi CRISPOR, http://crispor.tefor.net kullanılarak tasarlanmıştır. - Cas9-RNP kompleksini oluşturmak için tavlanmış RNA'ları, 10 μg / μL Cas9 nükleazını ve elektroporasyon tamponunu 20 dakika boyunca 37 ° C'de 5: 3: 2 hacim oranında karıştırılarak inkübe edin. Rutin çalışmalarda, bir lokus genom düzenlemesi için 1.25 μL tavlanmış RNA, 0.75 μL Cas9 nükleaz ve elektroporasyon tamponunun 0.5 μL'sini karıştırın ve inkübe edin.
- Aşağıdaki malzemeleri 1,5 mL'lik bir tüpte karıştırın: 10 μL elektroporasyon tamponu, 1 μL Cas9-RNP kompleksi (adım 3.2) ve 1 μL dairesel hedefleme vektörü (1 μg / μL, adım 1.6). Cas9-RNP-DNA karışımının toplam miktarı 12 μL'dir. Cas9-RNP-DNA karışımını elektroporasyona kadar buz üzerinde tutun.
NOT: KI'dan sonra gRNA'nın yeniden bölünmesini önlemek için, gRNA'yı, gRNA tanıma dizisinin KI donörünün entegrasyonu ile bölündüğü bir konumda tasarlayın. Eğer gRNA böyle bir yerde tasarlanamazsa, amino asit dizisinin değişmediği birkaç nükleotid sessiz mutasyon, KI donörünün plazmidine dahil edilir.
4. ESC'lerin gen hedeflemesi
- Jelatin kaplı hücre kültürü kaplarını hazırlamak için, 60 mm'lik bir tabak için 2 mL'de% 0.1 (w / v) jelatin çözeltisi, 24 delikli bir tabak için 500 μL, 96 delikli bir plaka için 100 μL ekleyin ve nemli bir inkübatörde 2 saat boyunca inkübe edin. Jelatin çözeltisini çıkarın, aynı miktarda PBS ile iki kez yıkayın ve kullanana kadar oda sıcaklığında saklayın.
- 1 dakika boyunca 37 ° C'lik bir su banyosu kullanarak mitotik olarak inaktive edilmiş dondurulmuş MEF stoğunu (adım 2.2) çözün ve MEF'i ESC tohumlamasından 1 gün önce jelatin kaplı 60 mm'lik bir tabağa (iki 60 mm tabak için 1.6 x 106 hücre içeren bir dondurulmuş MEF stok tüpü, adım 2.2) yerleştirin.
- Dondurulmuş bir ESC stok tüpünü (2 x 10 5 ESC / tüpte saklanır; hücre sayımı bir hücre sayacı kullanılarak gerçekleştirildi) 1 dakika boyunca 37 ° C'lik bir su banyosu kullanarak çözün ve önceden tohumlanmış MEF içeren 60 mm'lik bir kaba 1 x 105 ESC yerleştirin (adım 4.1).
- 4 mL ESC kültür ortamında kültür (ESCM, %15 (v/v) fetal sığır serumu ile DMEM bazlı modifiye ortam, 2 mM L-glutamin substratı, 100 U/mL penisilin, 100 μg/mL streptomisin, 0.1 mM 2-merkaptoetanol, lösemi inhibitör faktör ve t2i (0.2 μM PD0325901 ve 3 μM CHIR99021) ile ESC11'in pluripotensini koruyan 37 °C'de %5 CO2 ile ESC %50 ila %70 yakınlığa ulaşana kadar.
NOT: Üç farklı ESC hattı kullanıyoruz: JM8. B6N'den A3, B6-129 F1'den V6.5 ve şirket içinde BALB/c ESC geliştirildi. Bu makaledeki aynı KI protokolleri tüm ESC hatları için kullanılmaktadır. - Kültür ESC'sini 4 mL PBS ile yıkayın, ardından sindirim için 37 ° C'de 5 dakika boyunca 800 μL% 0.25 tripsin-EDTA çözeltisi ile muamele edin. Tripsini inaktive etmek için 1 mL ESCM ekleyin ve pipetleme ile ESC'leri tek hücrelere ayırın.
- ESC içeren çözeltiyi 280 x g'de 5 dakika boyunca santrifüj edin, süpernatanı atın, ESC'leri taze 1 mL ESCM'de yeniden askıya alın ve tripan mavisi boyamalı bir hücre sayacı kullanarak hücre sayısını sayın. 1 x 10 5 ESC'yi yeni bir1,5 mL tüpe aktarın ve 500 x g, 4 °C'de santrifüjleme yoluyla PBS ile üç kez yıkayın.
- ESC peletini 12 μL Cas9-RNP-DNA karışımında (adım 3.3) yeniden askıya alın ve köpürmeyi önlemek için nazik pipetleme ile iyice karıştırın. Elektroporasyon için yeniden askıya alınmış ESC'yi servis edin. Elektroporasyon sistemi, Cas9-RNP-DNA transdüksiyonu için 1.400 V ve 30 ms'de tek bir darbe kullanır (Şekil 1).
- Elektroporated ESC'leri, mitotik olarak inaktive edilmiş MEF ile 4 mL ESCM içeren 60 mm'lik bir kapta kültürleyin (bölüm 4.2). Ortamı her gün değiştirin.
- ESC'leri elektroporasyondan 3 ila 5 gün sonra 4 mL PBS ile yıkayın, ardından sindirim için nemli bir inkübatörde 5 dakika boyunca 37 ° C'de 800 μL% 0.25 tripsin-EDTA ve kültür ile muamele edin. Tripsin sindirimini durdurmak için 2 mL ESCM ekleyin ve hücre karışımını 5 dakika boyunca 280 x g'de santrifüj edin.
- Süper natantı atın, ESC'leri 1 mL taze ESCM'de yeniden askıya alın ve tripan mavisi boyamalı bir hücre sayacı kullanarak hücre konsantrasyonunu sayın. ESC'leri, ESCM ve besleyici MEF içeren 60 mm'lik çanak başına 1 x 103 ESC'de geçirin. Koloni toplanana kadar ortamı her gün değiştirin.
NOT: ESC'nin toplanmadan önce bir kez geçmesinin nedeni, genom düzenlemesinin ESC bölünmesinden sonra gerçekleşmeye devam edebilmesiydi, bu nedenle ilk koloni birçok durumda her zaman klon değildir. Bu nedenle, koloni toplamadan önce ESC'nin ilk geçişi, tek klonları almak için önemli bir adımdır.
5. Hedeflenen ESC'lerin PCR genotiplemesi
- ESC kültür kabından ESCM'yi ilk geçişten 5 ila 7 gün sonra aspire edin (adım 4.9) ve 4 mL PBS ekleyin. Stereomikroskop altında 20 μL'lik pipet kullanarak 5 μL PBS'li tek ESC kolonilerini toplayın (30x ila 40x büyütme). Bireysel kolonileri, 15 μL% 0.25 tripsin-EDTA çözeltisi içeren yuvarlak tabanlı 96 kuyucuklu bir plakanın kuyucuğuna yerleştirin.
- 96 kuyucuklu plakayı, 48 ayrı koloni toplanana kadar buz üzerinde tutun. 96 delikli plakayı 37 °C nemli% 5 CO2 inkübatöründe 5 dakika boyunca inkübe edin, ardından tripsin sindirimini durdurmak ve ESC kolonilerini pipetleme yoluyla tek hücrelere ayırmak için 80 μL ESCM ekleyin.
NOT: Bu yöntemin KI verimliliği genellikle% 10 -% 50 olduğundan, rutin olarak 48 klon alırız ve önce 24'ünü kullanarak genotipleme yaparız. Bu noktada birden fazla KI klonu elde edilemezse, kalan 24 klonu kullanarak KI'yı daha fazla tararız. - PCR genotiplemesi için kuyucuk başına 50 μL ESCM içeren jelatin kaplı besleyici içermeyen 96 delikli bir plakaya 40 μL ESC süspansiyonunu (adım 5) aktarın. Dondurulmuş ESC stoğunu yapmak için kalan 60 μL ESC süspansiyonunu, 500 μL ESCM ve besleyici MEF içeren jelatin kaplı 24 delikli bir plakadaki bir kuyuya aktarın.
- 24 delikli plakada (adım 5.3) kültürlenmiş bireysel ESC klonlarını% 60 ila% 80 akıcılığa ulaşana kadar stoklayın. Yukarıda belirtildiği gibi tripsinizasyon ile bireysel ESC klonlarını kurtarın, hücreleri yeni bir 1.5 mL tüpte toplayın ve 5 dakika boyunca 280 x g'de santrifüj yapın. Süper natantı atın, hücreleri 500 μL hücre dondurma ortamında yeniden askıya alın ve -80 ° C derin dondurucu kullanarak dondurun.
- PCR genotiplemesi için, kültür ESC, ESC% 90'dan fazla akıcıya ulaşana kadar 96 kuyucuklu besleyicisiz plakada (adım 5.3) klonlanır. ESCM'yi her bir kuyucuktan aspirasyonla çıkarın ve 100 μL PBS ile iki kez yıkayın.
- PBS'yi aspire edin ve proteinaz K içeren 100 μL lizis tamponu ekleyin, iyice karıştırın ve ESC lizatını yeni bir 1.5 mL tüpe aktarın. ESC lizatını 65 °C'de bir ısı odası kullanarak en az 1 saat ısıtın. Geleneksel bir fenol-kloroform DNA saflaştırma yöntemi ve ardından etanol çökeltmesi kullanarak genomik DNA'yı çıkarın ve saflaştırın.
- Çökeltilmiş DNA'yı 20 μL DNaz içermeyen suda çözün ve bir spektrofotometre kullanarak DNA'nın saflığını ve konsantrasyonunu belirleyin. Hedef bölgeyi yükseltmek için lokusa özgü primer setleri kullanarak genomik PCR gerçekleştirin. Ardından, diziyi kontrol edin ve istediğiniz KI dizilerine sahip ESC klonlarını seçin.
6. Sekiz hücreli aşama embriyonun hazırlanması ve ESC'lerin mikroenjeksiyonu
- Yetişkin ICR kadınlarına intraperitoneal olarak 5 IU hamile kısrak serum gonadotropin (PMSG) enjekte edin. 48 saat sonra, aynı dişilere 5 IU insan koryonik gonadotropin (hCG) enjekte edin, ardından hormonla tedavi edilen kadınları ICR erkekleriyle eşleştirin. Ertesi gün vajinadaki çiftleşme tıkacını kontrol edin (embriyonik gün 0.5, ED0.5).
NOT: Kimerizmi kat renk oranlarına göre değerlendirmek için, siyah veya agouti kaplama rengi arka planına sahip bir ESC için alıcı olarak ICR suşundan blastosist veya albino arka plana sahip bir ESC için alıcı olarak C57BL / 6 suşundan blastosist kullanın. - Yumurta kanalını ED1.5'te toplayın ve HEPES tamponlu orta (FHM) damlalarına yerleştirin. İki hücreli veya dört hücreli aşama embriyolarını, infundibuluma yerleştirilen bir yıkama iğnesi kullanarak yumurta kanalını FHM ile yıkayarak geri kazanın.
- Toplanan embriyoları bir ağız pipeti kullanarak birkaç taze FHM damlasına taşıyarak yıkayın. 50 μL KSOM'daki embriyoları, 37 ° C'de mineral yağ ile kaplı 35 mm'lik bir hücre kültürü kabına, sekiz hücreli veya morula aşamasına (ED2.5 ) ulaşılana kadar 1 gün boyunca% 5 CO 2 inkübatöre bırakın.
- Embriyolar bir sonraki toplama gününde kullanılmazsa, CARD protokolüne (http://card.medic.kumamoto-u.ac.jp/card/english/sigen/manual/ebvitri.html) göre vitrifikasyon yoluyla dondurun ve kullanıma kadar sıvı azotta saklayın.
- Bir çektirme ve mikroforge kullanarak embriyoları (80-100 μm dış çap) ve ESC enjeksiyonunu (yaklaşık 13 μm çapında) tutmak için cam pipetler hazırlayın.
- FHM içeren bir tutma pipetini, mikromanipülatörün sol tarafına monte edilmiş ve bir mikroenjektöre bağlı kılcal tutucuya entegre edin. Bir enjeksiyon pipetini başka bir mikro enjektöre bağlayın ve sağ tarafa yerleştirin. İki pipeti mikroskop görüş alanında düz bir şekilde hizalayın.
- FHM ortamında %12 (w/v) polivinilpirolidon (PVP) ve 5 μL FHM damla 5 μL damlayı 60 mm'lik bir kabın aynı kapağına yan yana dağıtın ve damlaları mineral yağ ile örtün. Embriyoları 5 μL FHM damlasına aktarın ve embriyo içeren FHM damlasına ESC süspansiyonunun 1-5 μL'sini ekleyin.
NOT: Damla hazırlığından önce ESC ve MEF süspansiyonunu 60 mm kültür kabındaki 4 mL ESM'de 30 dakika bekletin. MEF'ler tabana ESC'lerden daha hızlı yapıştığından, bu 30 dakikalık inkübasyon, süpernatanttaki bağlanmamış ES hücrelerinin konsantrasyonuna izin verir. - Enjeksiyon pipetinin içini PVP ile yıkayın, ardından embriyoları ve ESC'leri içeren damlaya geçin.
- Enjeksiyon pipetinde hücre bölünmesini (altı hücre) yeni bitirmiş üç ayrı ECS çiftini alın. Sıkıştırmayı tamamlamış sekiz hücreli veya morula evreli bir embriyoyu, tutma pipeti ile aspirasyonla tutun. Piezoelektrik darbeli zona pellucida'da bir delik açın (yoğunluk: 3; hız: 3). Zona pellucida'nın içindeki altı hücreli ESC'yi dışarı atın ve pipeti embriyolardan çekin.
- ESC enjekte edilen embriyoları bir ağız pipeti kullanarak birkaç KSOM damlası ile hafifçe yıkayın ve embriyolar blastosist aşamasına gelene kadar% 5 CO2 olan 37 ° C'de mineral yağ ile kaplı yeni bir 50 μL KSOM damlasında inkübe edin.
- ESC enjekte edilen 10 blastosisti, psödohamile dişi farelerin her bir uterus boynuzuna aktarın (2.5 gün postkoitum, kafa başına 20 blastosist).
- Taşıyıcı anne doğum yaptıktan sonra, en yüksek kimerizm oranına sahip en az iki erkek kimera seçin (ceket rengiyle doğrulandığı gibi% 70 veya daha yüksek) ve ardından F1 heterozigot KI elde etmek için uygun suş dişileriyle çiftleşin.
- İstenilen genotip (ler) e veya araştırmaya uygun genetik geçmişlere sahip fareler elde etmek için bireysel KI farelerini uygun ortaklarla yetiştirin.
Sonuçlar
ESC'deki spesifik genleri ve ardından önceki makalemiz11'e göre gen manipüle edilmiş fare üretimini geliştirmek için kimera üretimini hedefledik. ESC genotiplemesi (bölüm 4'te açıklanmıştır) rutin olarak PCR ile primerler kullanılarak gerçekleştirilir. Primerler, homoloji kollarının dışındaki genomik diziler ve KI DNA fragmanındaki spesifik diziler üzerinde tasarlanmıştır (Şekil 2A). Bu durumda, vahşi tip alel yükseltilmez, oysa belirli bir boyuttaki bir PCR amplikonu yalnızca hedeflenen eksojen DNA hedef lokusta KI olduğunda tespit edilir. Temsili genotipleme PCR sonuçları Şekil 2B'de gösterilmiştir. Bu durumda 22 klondan dokuzu (%40.9) KI'ya özgü bir bant gösterdi. Şekil 2'de gösterilen sonuç da dahil olmak üzere üç temsili hedefleme sonucu Tablo 1'de gösterilmiştir. Bu sonuçlar, burada gösterilen yöntemin, herhangi bir ilaç seçimi olmaksızın gen KI için etkili ve tekrarlanabilir olduğunu göstermektedir.
ESC bir enjeksiyon pipetinde toplanır, daha sonra zona pellucida'da bir piezoelektrik artı kullanılarak bir delik açılır ve ESC embriyonik hücreler arasında serbest bırakılır (Şekil 3). Teknik olarak, bir ESC'yi sekiz hücreli veya morula evreli bir embriyoya enjekte etmek için kullanılan bu protokol, birçok fare tesisinde yaygın olarak kullanılan bir blastosiste ESC enjeksiyonu protokolüne benzer. Temsili kimerik fareler Şekil 4'te gösterilmiştir. Kimerizmin kat rengi değerlendirmesi için, ICR suşundan (albino, beyaz saç) elde edilen embriyo B6 veya B6-129 F1 ESC alıcısı olarak kullanıldı ve B6 embriyoları BALB/c ESC'lerin alıcısı olarak kullanıldı (Şekil 4).

Şekil 1: Bir ESC genomunun spesifik lokusuna CRISPR / Cas9 ribonükleoprotein (RNP) aracılı dairesel plazmid entegrasyonunun bir şeması. Elektroporasyon, Cas9-RNP ile ESC'lere hedefleme vektörü olarak dairesel bir plazmid sunar. Kısaltmalar: GOI = ilgilenilen gen, HA = homoloji kolu. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
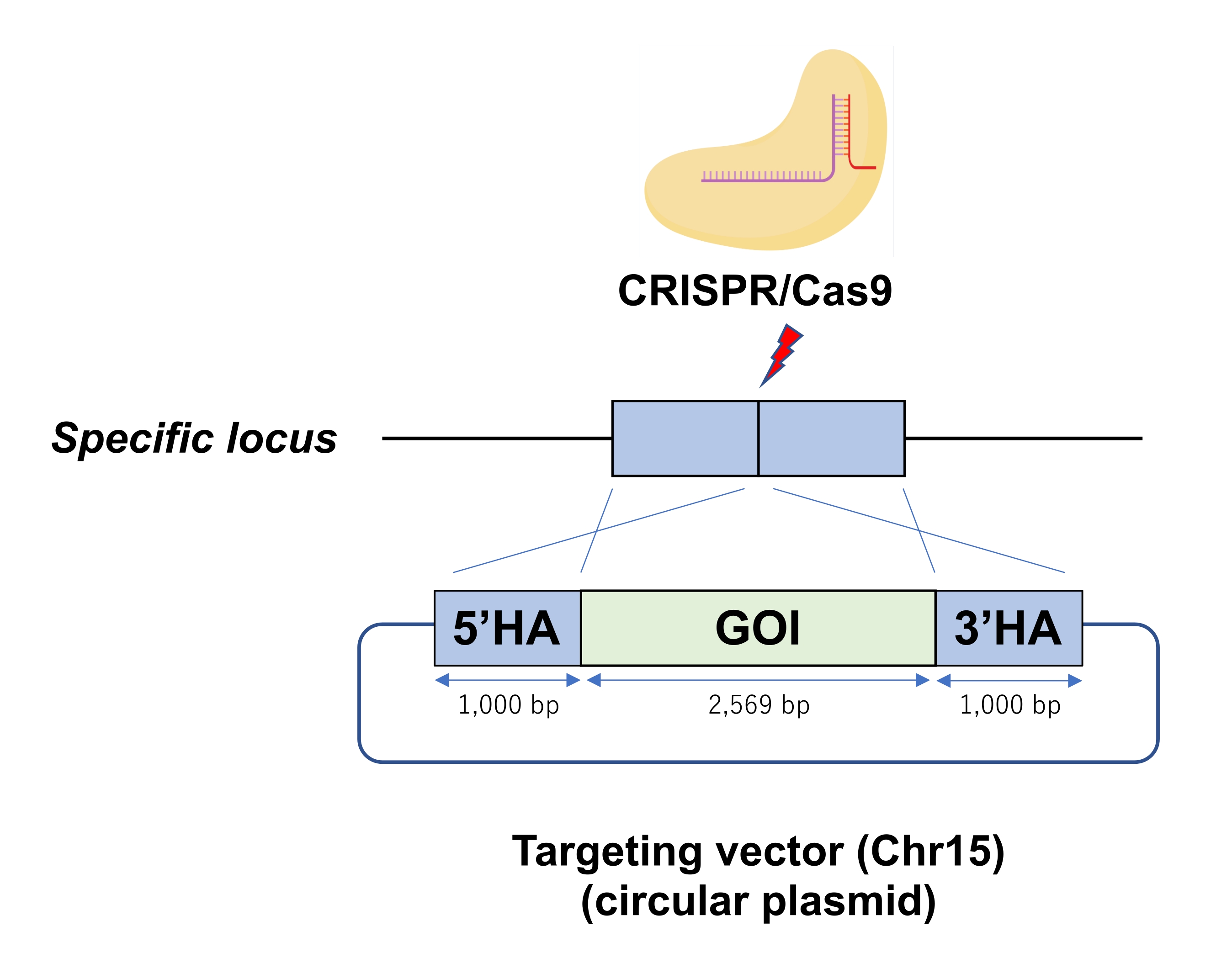{kind=link}

Şekil 2: Hedeflenen ESC klonlarının KI taraması için genomik PCR analizleri. (A) KI-spesifik PCR primerleri, homoloji kollarının dışındaki genomik diziler (ileri) ve KI DNA fragmanındaki spesifik diziler (ters) üzerinde tasarlanmıştır. (B) Temsili genotipleme PCR sonuçları. Negatif kontrol olarak vahşi tipte bir genom kullanıldı. Kısaltmalar: GOI = ilgilenilen gen, HA = homoloji kolu. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Sekiz hücreli embriyo enjeksiyonunun temsili görüntüleri . (A) Mikroenjeksiyon için, üç çift ESC (altı hücre, ok ucu) alınır. (B) Zona pellucida'da bir piezoelektrik darbe ile bir delik açılır ve ESC'ler her blastomer arasında dışarı atılır. Gösterilen ölçek çubuğu 50 μm'dir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Kimera farelerinin temsili görüntüleri . (A,B) ESC, C57BL/6N'den türetilmiştir (JM8. A3, Agouti saç; A) veya B6-129 F1 (Agouti saç; B) ICR (albino, beyaz saç) embriyolarına enjekte edilir, ardından embriyolar anne vekillerine transfer edilir. (C) BALB / C'den (albino, beyaz saç) elde edilen ESC, C57BL / 6J (siyah saç) embriyolarına enjekte edilir ve ardından embriyolar anne vekillere transfer edilir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Proje Kimliği | Koromozom | 5'HA uzunluğu (bp) | 3'HA uzunluğu (bp) | Kesici uç boyutu (bp) | Analiz edilen klonların sayısı | KI klonlarının sayısı | Verimlilik (%) | Açıklamalar |
| R26-CC* | Chr6 | 965 | 1006 | 5321 | 23 | 2 | 8.7% | - |
| R4-03* | Chr8 | 1000 | 997 | 3070 | 22 | 6 | 27.3% | - |
| P4-01* | Chr15 Serisi | 1000 | 1000 | 2569 | 22 | 9 | 40.9% | Şekil 2'de gösterilmiştir |
Tablo 1: ESC'deki üç bağımsız genomik lokusta KI verimlilikleri. *Bu projeler bugüne kadar yayınlanmamıştır. Bu genlerin adı gelecekte bağımsız el yazmalarında açıklanacaktır.
Tartışmalar
ESC'lerin gen hedeflemesi ve ardından kimera üretimi, gen manipüle edilmiş farelerin geliştirilmesi için geleneksel olarak kullanılmıştır. Bununla birlikte, hedefleme vektörü pozitif veya negatif ilaç seçimi gen kasetleri ile uzun (> normalde birkaç kb) homoloji kolları içermesine rağmen, gen knock-in etkinliği düşük kalmaktadır. Protokolümüz, rutin işler için kabul edilebilir bir verimlilikte Cas9-RNP aracılı genom düzenlemesi eşliğinde bir hedefleme vektörü olarak herhangi bir ilaç seçim kaseti içermeyen dairesel bir plazmid kullanarak uzun eksojen DNA'nın ayarlanmış bir ESC KI yöntemini tanıttı. Bu nedenle, bu protokol, geleneksel ESC hedeflemesine kıyasla genetiği değiştirilmiş kimerik farelerin üretiminde geçen süreyi önemli ölçüde azaltmaya yardımcı olabilir.
Bu protokolde, genomda bölgeye özgü çift iplikçikli kırılmaları indüklemek için CRISPR / Cas9-RNP kullandık ve hedefleme vektörü olarak doğrusallaştırılmış bir vektör yerine dairesel bir plazmid kullanıldı. Doğrusallaştırılmış plazmidler, genomik entegrasyon 8,9,10'un artan etkinliği nedeniyle geleneksel olarak gen KI için bir hedefleme vektörü olarak kullanılır. Bununla birlikte, vektör homolog kollar 9 içermesine rağmen, birçok genomik entegrasyon spesifikdeğildir. Öte yandan, dairesel plazmidler, ESC12 veya fibroblastlar13'ün genomuna entegre edilmesi zor özellikleri nedeniyle vektörleri hedeflemek için nadiren kullanılır. Bu nedenle, dairesel bir plazmidin CRISPR / Cas9 aracılı genom düzenlemesi eşliğinde bir hedefleme vektörü olarak uygulanmasının, spesifik olmayan rastgele entegrasyonu en aza indirmesi, ancak ESC genomuna bölgeye özgü entegrasyonu en üst düzeye çıkarması mümkün olacaktır. CRISPR / Cas9 tarafından çift iplikçik kırılmasının indüksiyon verimliliğinin oldukça önemli olduğu dikkat çekici olacaktır14, bu nedenle yüksek verimlilikte çift iplikçik kırılmasına neden olan gRNA'nın kullanılması gerekli olacaktır. KI verimliliği çok düşük olduğunda gRNA'nın yeniden tasarlanması düşünülmelidir. Şekil 2'de gösterilen durumda, ESC klonlarının% 40.9'u KI'ya özgü bir bant göstermiştir. Aslında, enstitünün çekirdek laboratuvarı olarak, burada açıklanan yöntemle ESC'leri kullanarak rutin olarak gen hedefleme yapıyoruz ve gen lokusuna bağlı olarak bazı varyasyonlar olmasına rağmen, Cre, CreERT veya floresan muhabirleri gibi 1-2 kb dizileri için% 10-50'lik KI verimlilikleri elde ettik. Bu yöntemi kullanarak başarılı bir şekilde KI'ya sahip olduğumuz en uzun DNA dizileri, Rosa26 lokusuna yaklaşık 11.2 kb'dir ve verimlilik% 12.2'dir (analiz edilen 41 koloniden beş KI kolonisi).
Bu protokolün ESC mikroenjeksiyonu için alıcı olarak sekiz hücreli veya morula evre embriyoları kullandığı, ancak blastosist evre embriyolarını kullanmadığı da not edilecektir. Bu yazıda karşılaştırma deneyleri yapmamış olsak da, birkaç rapor ESC'lerin sekiz hücreli veya morula evreli embriyolara enjeksiyonunun, blastosistlere ESC enjeksiyonuna kıyasla kimerik yavrulara ESC katkısının verimliliğini önemli ölçüde artırdığını göstermiştir15,16,17,18 . Gerçekten de, C57BL / 6, B6-129 F1 ve BALB / c dahil olmak üzere çeşitli ESC hatlarının ESC enjeksiyonları, genellikle bazı yavrularda yüksek kat renginde kimera gelişimine neden olmuştur, ancak hepsinde değil (bkz. Şekil 4). Yöntemin sınırlaması, preimplantasyon embriyosuna enjekte edilen ESC'lerin her zaman bir kimera 19,20'deki germ hücrelerine katkıda bulunamamasıdır. ESC'lerin germline iletimi, her ESC klonu19'un kalitesine bağlı olacaktır. Bu nedenle, kararlı kimerik fare üretimi için yedek olarak birden fazla ESC klon hattı geliştirmek avantajlı olacaktır. Sonuç olarak, burada sunulan protokoller, ilaca dirençli herhangi bir kaset olmadan sadece her bir homoloji kolunu ve KI için ilgi çekici geni içeren basit hedefleme vektörleri kullanmaktadır. Bu nedenle, vektör yapısı çok daha kolay olacak ve kültür süresi geleneksel ESC hedefleme tekniğine kıyasla kısaltılabilir. Bu, yaşam bilimlerinde gelecekteki analizler için çeşitli genetiği değiştirilmiş farelerin hızlı ve kolay bir şekilde üretilmesine yardımcı olacaktır.
Açıklamalar
Yazarların ifşa etmek için rakip çıkarları yoktur.
Teşekkürler
Osaka Üniversitesi'nden Saki Nishioka'ya, Biyoteknoloji Araştırma ve Geliştirme (kar amacı gütmeyen kuruluş) ve Tokyo Üniversitesi Tıp Bilimleri Enstitüsü'nden Mio Kikuchi ve Reiko Sakamoto'ya mükemmel teknik yardımları için teşekkür ederiz. Bu çalışma şu kişiler tarafından desteklenmiştir: Eğitim, Kültür, Spor, Bilim ve Teknoloji Bakanlığı (MEXT)/Japonya Bilimi Geliştirme Derneği (JSPS) KAKENHI MI (JP19H05750, JP21H05033) ve MO (20H03162); Evrimsel Bilim ve Teknoloji için Temel Araştırma (CREST), Japonya Bilim ve Teknoloji Ajansı (JST) MI'ye hibe (JPMJCR21N1); Eunice Kennedy Shriver Ulusal Çocuk Sağlığı ve İnsani Gelişme Enstitüsü MI'ye (R01HD088412); Bill & Melinda Gates Vakfı'ndan MI'ye (Grand Challenges Explorations hibesi INV-001902); ve Mikrobiyal Hastalıklar Araştırma Enstitüsü, Osaka Üniversitesi'nden MI'ye ve MO'ya Ortak Araştırma Projesi için Hibe.
Malzemeler
| Name | Company | Catalog Number | Comments |
| BALB/c ESC | - | - | ESC developed from BALB/c strain |
| Bambanker | Nippon Genetics | CS-02-001 | Cell-freezeing medium. Section 2.6 and elsewhere |
| Cas9 Nuclease V3 | IDT | 1081059 | Section 3.2 and elsewhere. |
| CHIR99021 | FUJIFILM Wako | 038-23101 | Section 4.3 |
| CreERT gene fragment | GeneWiz | Section 1.1. | |
| CRISPR-Cas9 crisprRNA | IDT | crisprRNA. Section 3.1 and elsewhere. | |
| CRISPR-Cas9 tracrRNA | IDT | 1072534 | tracrRNA. Section 3.1 and elsewhere. |
| DMEM | Nacalai | 08458-45 | MEF medium. Section 2.3 and elsewhere |
| Duplex buffer | IDT | 1072534 | RNA dilution buffer. Section 3.1 and elsewhere. |
| FastGene Gel/PCR Extraction Kit | Nippon Genetics | FG-91302 | Section 1.1 and 1.2. |
| GlutaMax | Thermo Fisher | 35-050-061 | L-glutatime substrate |
| hCG | ASKA Animal Health | Section 6.1. | |
| In-Fusion HD Cloning Kit | Clontech | 639648 | DNA cloning kit. Section 1.3 and elsewhere |
| JM8.A3 ESC | EuMMCR | - | ESC developed from C57BL/6N strain |
| Knock-out DMEM | Thermo Fisher | 10829018 | Section 4.3 and elsewhere, DMEM-based modified commercial medium. |
| KSOM | Merck | MR-121-D | Section 6.3 and 6.9. |
| Leukemia inhibitory factor | FUJIFILM Wako | 125-05603 | Section 4.3. No unit concentration data is supplied by the provider. Used 1,000-fold dilution in this protocol. |
| Neon Electroporation system | Thermo Fisher | MPK5000 | Section 3.2, 4.5 and elsewhere. The system containes electroporation buffer as well used in section 3.2. |
| NucleoSpin Plasmid Transfection-grade | Takara | U0490B | Section 1.6. |
| PD0325901 | FUJIFILM Wako | 162-25291 | Section 4.3 |
| PMSG | ASKA Animal Health | Section 6.1. | |
| Tail lysis buffer | Nacalai | 06169-95 | Section 5.5. |
| Trypsin-EDTA | Nacalai | 32777-15 | Section 2.2 and elsewhere |
| V6.5 ESC | - | - | ESC developed from B6J-129 F1 strain |
| X-ray irradiation device | Hitachi | MBR-1618R-BE | Section 2.6. |
Referanslar
- Jinek, M., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Wang, H., et al. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell. 153 (4), 910-918 (2013).
- Yang, H., et al. One-step generation of mice carrying reporter and conditional alleles by CRISPR/Cas-mediated genome engineering. Cell. 154 (6), 1370-1379 (2013).
- Oji, A., et al. CRISPR/Cas9 mediated genome editing in ES cells and its application for chimeric analysis in mice. Scientific Reports. 6, 31666(2016).
- Kaneko, T., Mashimo, T. Simple genome editing of rodent intact embryos by electroporation. PLoS One. 10 (11), 0142755(2015).
- Chen, F., et al. High-frequency genome editing using ssDNA oligonucleotides with zinc-finger nucleases. Nature Methods. 8 (9), 753-755 (2011).
- Hashimoto, M., Yamashita, Y., Takemoto, T. Electroporation of Cas9 protein/sgRNA into early pronuclear zygotes generates non-mosaic mutants in the mouse. Developmental Biology. 418 (1), 1-9 (2016).
- Mansour, S. L., Thomas, K. R., Capecchi, M. R. Disruption of the proto-oncogene int-2 in mouse embryo-derived stem cells: a general strategy for targeting mutations to non-selectable genes. Nature. 336 (6197), 348-352 (1988).
- Johnson, R. S., et al. Targeting of nonexpressed genes in embryonic stem cells via homologous recombination. Science. 245 (4923), 1234-1236 (1989).
- Hasty, P., Ramires-Solis, R., Krumlauf, R., Bradley, A. Introduction of a subtle mutation into the Hox-2.6 locus in embryonic stem cells. Nature. 350 (6315), 243-246 (1991).
- Ozawa, M., Emori, C., Ikawa, M. Gene targeting in mouse embryonic stem cells via CRISPR/Cas9 ribonucleoprotein (RNP) mediated genome editing. Genome Editing in Animals - Methods and Protocols 2nd edition. Hatada, I. , (2022).
- Yagi, M., et al. Derivation of ground-state female ES cells maintaining gamete-derived DNA methylation. Nature. 548 (7666), 224-227 (2017).
- Gassmann, M., Donoho, G., Berg, P. Maintenance of an extrachromosomal plasmid vector in mouse embryonic stem cells. Proceedings of National Academy of Sciences. 92 (5), 1292-1296 (1995).
- Okita, K., Nakagawa, M., Hyenjong, H., Ichisaka, T., Yamanaka, S. Generation of mouse induced pluripotent stem cells without viral vectors. Science. 322 (5903), 949-953 (2008).
- Poueymirou, W. T., et al. F0 generation mice fully derived from gene-targeted embryonic stem cells allowing immediate phenotypic analyses. Nature Biotechnology. 25 (1), 91-99 (2007).
- Hu, M., et al. Efficient production of chimeric mice from embryonic stem cells injected into 4- to 8-cell and blastocyst embryos. Journal of Animal Science and Biotechnology. 4 (1), 1-7 (2013).
- Guo, J., et al. Contribution of Mouse Embryonic Stem Cells and Induced Pluripotent Stem Cells to Chimeras through Injection and Coculture of Embryos. Stem Cells International. 2014, 409021(2014).
- Bodai, Z., Bishop, A. L., Gantz, V. M., Komor, A. C. Targeting double-strand break indel byproducts with secondary guide RNAs improves Cas9 HDR-mediated genome editing efficiencies. Nature Communications. 13 (1), 2351(2022).
- Kobayashi, T., Goto, T., Oikawa, M., Sanbo, M., Yoshida, F., Terada, R., Niizeki, N., Kajitani, N., Kazuki, K., Kazuki, Y., Hochi, S., Nakauchi, H., Surani, A., Hirabayashi, M. Blastocyst complementation using Prdm14-deficient rats enables efficient germline transmission and generation of functional mouse spermatids in rats. Nature Communications. 12 (1), 1328(2021).
- Miura, K., Matoba, S., Hirose, M., Ogura, A. Generation of chimeric mice with spermatozoa fully derived from embryonic stem cells using a triple-target CRISPR method for Nanos3. Biology of Reproduction. 104 (1), 223-233 (2021).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır