
Method Article
通过的嗅球钙监控录像的变化温度引起的神经元电活动
* 这些作者具有相同的贡献
摘要
Here we describe a protocol for measuring and analyzing temperature responses in the olfactory bulb of Xenopus laevis. Olfactory receptor neurons and mitral cells are differentially stained, after which calcium changes are recorded, reflecting a sensitivity of some neural networks in the bulb to temperature drops induced at the nose.
摘要
The olfactory system, specialized in the detection, integration and processing of chemical molecules is likely the most thoroughly studied sensory system. However, there is piling evidence that olfaction is not solely limited to chemical sensitivity, but also includes temperature sensitivity. Premetamorphic Xenopus laevis are translucent animals, with protruding nasal cavities deprived of the cribriform plate separating the nose and the olfactory bulb. These characteristics make them well suited for studying olfaction, and particularly thermosensitivity. The present article describes the complete procedure for measuring temperature responses in the olfactory bulb of X. laevis larvae. Firstly, the electroporation of olfactory receptor neurons (ORNs) is performed with spectrally distinct dyes loaded into the nasal cavities in order to stain their axon terminals in the bulbar neuropil. The differential staining between left and right receptor neurons serves to identify the γ-glomerulus as the only structure innervated by contralateral presynaptic afferents. Secondly, the electroporation is combined with focal bolus loading in the olfactory bulb in order to stain mitral cells and their dendrites. The 3D brain volume is then scanned under line-illumination microscopy for the acquisition of fast calcium imaging data while small temperature drops are induced at the olfactory epithelium. Lastly, the post-acquisition analysis allows the morphological reconstruction of the thermosensitive network comprising the γ-glomerulus and its innervating mitral cells, based on specific temperature-induced Ca2+ traces. Using chemical odorants as stimuli in addition to temperature jumps enables the comparison between thermosensitive and chemosensitive networks in the olfactory bulb.
引言
Over the last years, temperature sensitivity has no longer been described as a somesthetic sense only, but also as a physiological function relevant for the olfactory system. In rodents, the main olfactory bulb receives input from the Grueneberg ganglion (GG), an organ in the nasal cavity, consisting of thermosensitive neurons. GG neurons respond to cool temperatures1 as well as to chemical stimuli, and their chemosensitivity is modulated by temperature fluctuations2. These observations suggest that the olfactory bulb may integrate chemical and temperature information collected at the nose. In order to explore this hypothesis, we present here a set of experiments enabling the detection of temperature responses in the olfactory bulb of non-transgenic animals, using the Xenopus laevis larva as a model. The organization of the olfactory system in these animals closely resembles that of mammals. The olfactory receptor neurons of premetamorphic X. laevis terminate in tufts, and make synaptic contacts with the dendrites of second-order neurons, the mitral cells. Pre- and postsynaptic fibers intermingle and form skein-like neuropil structures called glomeruli3. The abundant synapses of the glomerular layer represent the first processing center of olfactory information. Mitral cells further integrate the sensory input and convey it to higher olfactory areas.
We have developed a protocol combining electroporation of olfactory receptor neurons (ORNs) with calcium-sensitive and non-sensitive dyes followed by bolus loading of the postsynaptic network of glomeruli and mitral cells. The staining by electroporation of two spectrally distinct dyes loaded in the nasal cavities serves to single out the γ-glomerulus3 through its bilateral innervation by ORNs from both olfactory epithelia. Thus, the location of the γ-glomerulus is identified prior to further measurements. Subsequently, bolus loading4 with Fluo-8 acetoxymethyl (Fluo-8 AM) is carried out in a volume comprising the γ-glomerulus. Imaging calcium changes with fast confocal microscopy allows the visualization of temperature responses in the 3D neuropil surrounding the γ-glomerulus, a unique temperature-sensitive glomerulus in this system5. Mitral cells innervating this specific structure can also be identified by their Ca2+ signals responsive to induced temperature drops. Next, activity correlation imaging6 uses the specific Ca2+ traces of these cells to reveal the dendritic morphology of thermosensitive mitral cells. Alternating repeated applications of cold Ringer solution and chemical odorants in one measurement can be used to visualize the mitral cell networks for odor and temperature processing surrounding the γ-glomerulus and identify potential overlaps. To unambiguously assign the responses to either the chemical or the temperature stimulus, we constantly monitor temperature at the olfactory epithelium.
研究方案
与非洲爪蟾蝌蚪所有实验均根据道德哥廷根大学委员会在动物实验核准的准则进行。
1.电穿孔
- 根据新科普和法贝尔7选择阶段49-54的动物。
- 确保记录装置由一个大的工作距离并能够在至少2 Hz的频率施加20伏的电压脉冲20毫秒电穿孔装置的立体的。通过两个铂电极用200μm的粗径施加电压脉冲,以便它们可以被插入到蝌蚪'鼻孔,而不会造成损坏。
- 制备葡聚糖共轭染料的晶体( 例如,钙绿10 kDa的葡聚糖,或的Alexa Fluor 647 10 kDa的葡聚糖)由5毫克的通常输送量溶解于约100μl的蒸馏水,并让小液滴2微升的封口膜的板材干燥。
注:晶体不到一天的时间晾干,可在-18℃,之后在冰箱中储存期限超过一年的。 - 通过将它们在含有三卡因甲磺酸3%1-2分钟,直到它到达其特征在于手术麻醉状态自来水麻醉动物心脏速率,所有运动的损失和缺乏对机械刺激的反应减少。
- 沉浸在麻醉的动物在纯净的自来水10秒。
- 广场上的凝胶垫的动物,并通过将其周围的针而不伤害它解决它。
注意:不要通过其皮肤戳针固定的动物。 - 轻轻擦干周围用纸巾鼻孔的区域。
- 将动物的体视显微镜下,专注于鼻孔。
- 使用镊子挑大小相匹配的鼻孔孔的染料晶体,将其放置在鼻腔中的一个,等到它完全溶解,WHICH需要不到1分钟。如果晶体较小,加入2或3到每个空腔,直到它们产生一个非透光高浓缩溶液。
- 放置在阴极上的蝌蚪的皮肤和阳极插入鼻孔中的一个。
注:此特定设置适用于1.3步提到的染料。用于与不同极性的染料,染色结果可通过将阴极插入鼻孔得到改善。如果不能确定染料的极性,两个电极可以同时在鼻孔(每鼻孔一个)放置,并单电压脉冲之间交替的极性。 - 应用的20 V和20毫秒的六个脉冲大约0.5秒刺激间隔的。
注意:小气泡应该提供的电极与皮肤和鼻孔的溶液接触的电穿孔中出现在鼻孔电极周围。如果没有气泡可见检查连接电缆,确保电穿孔德维奇e的传送所需的电压脉冲。 - 重复第二鼻孔的过程(1.9-1.11)。
- 转移电穿孔蝌蚪成在室温下充满自来水的烧杯中。等待5到10分钟,直到动物苏醒并恢复游泳。给它并让它恢复为至少1天,在此期间染料沿着嗅神经至嗅球输送。
- 在使用1-7天电穿孔动物电的最佳成像效果如下。给至少24小时的时间前成像染料运输和恢复。
2.整装准备
- 麻醉动物在含有三卡因甲磺酸3%,直到所有的动作都停止,它不再响应机械刺激自来水。
- 转移蝌蚪到凝胶垫在立体显微镜下,并通过经由皮肤上的FO的每一侧戳针紧紧固定它rebrain。
- 通过切断其脊髓牺牲蝌蚪。
- 使用手术刀解剖出含有鼻腔,嗅觉神经和嗅球( 图1A)的组织块。使第一个切口靠近左边的鼻孔,不接触和移动刀片切割着旁边的左嗅神经和灯泡,直到端脑,间脑的边框。这样做右侧。通过使最后的切割后的端脑隔离来自神经系统的其他部分的制备方法。
- 蝌蚪体内的处置,并仔细翻转组织块上下颠倒使大脑制备的腹侧面向上。
- 通过插入嗅觉神经之间的两针针再次组织块。
- 放的青蛙Ringer溶液(98毫摩尔NaCl,2mM的氯化钾的下降,1毫氯化钙 ,2mM的MgCl 2的,5mM葡萄糖,5mM的丙酮酸钠,10毫米的HEPES,调节至pH 7.8,osmola对组织230 mOsmol / L)的RITY。
- 通过用细剪刀三切口取出覆盖在轴突分拣区脑膜和嗅球。从组织块的后缘开始,沿左嗅球caudorostrally紧密切直到入口点的嗅觉神经进入灯泡(轴突分拣区)。
- 重复步骤2.8右半球。
- 提高与钳脑膜和使所述第三切口,垂直于以往在轴突分拣区域。
注意:嗅球的腹侧是现在用于成像和丸药加载访问。 - 为了提高透射光图像的图像质量,重复背侧的过程。这些额外的步骤可以方便的微量为丸药装载的导航,但不是绝对必要的。
- 将样品转移到充满青蛙林格氏用于进一步处理和成像的记录室中。让小号URE该腹侧朝上,并与尼龙纤维的网固定样品跨越在小的铂帧。
- 对于刺激的一个更简单的应用放置在鼻腔的尼龙纤维中的一个的顶部。
3.丸装载
- 在含有的Pluronic F-127的20%的二甲亚砜20微升(DMSO)中溶解钙AM染料( 例如,的Fluo-8 AM)的50微克(重量/体积),以制备调幅染料的储备溶液。在1-2微升体积小等分冻结股票的解决方案。该原液为至少半年稳定的,但避免反复冻融。
- 使用微量拉马拉移液器5-8MΩ的电阻和1-2微米的尖端直径。
- 溶解在青蛙Ringer溶液的染料制备的储备溶液在250-500微米的浓度。
- 添加MK571达到500μM的浓度以阻止多药耐药转运。
- 使用一个细长枪头填充10微升该溶液的微量和轻弹微量除去所有气泡。
- 安装在吸液管保持器的微并确保压力可以用注射器或充气药物喷射装置是手动施加并监视用压力计所施加的压力。
- 如果可能,使用一个显微镜的设置有能力来激励注入染料,使得从微量移液器尖端流出可以可视化和调整。理想情况下,使用共焦成像设置。使用正置显微镜用浸泡的目的,这样的组织可以与微管抵达。
- 放置在显微镜下,整个安装制备,遵循嗅神经直到灯泡和聚焦到在嗅球感兴趣的区域。
- 灌注通过记录室新鲜林格氏溶液,以增加制剂的可行性和洗出的染料泄漏从微量移液器尖端。
- 降低吸移管到嗅球的表面上,与尖端指向成制剂的延髓方向。
- 应用一个小而恒定正压力(〜25百帕)到微量,以防止移液管的堵塞,并轻轻将其插入组织中。
- 一旦吸管违反外组织层,将它在rostro手背方向到僧帽细胞层。 (请注意,通过电穿孔嗅觉受体神经元的一个计数器染色有助于调节微量的立场。)理想的是,放置在移液管尖端在50-100微米从期望的记录部位的距离。
- 染色温度敏感性γ-肾小球,从γ-肾小球的突触前神经纤维的位置定位的区域约50μm吻侧。
- 在100-200百帕的范围内施加正压。视的大小调整压力强度微量移液器尖端。通过留意下当施加首次压力明亮光照明可见轻微组织运动确认从吸管的流出。
- 保持约10分钟的恒定压力,而微量残留在组织中。如果可能的话,请在此期间对小区负载的神经纤维在荧光灯的照明,如加载成功会导致可见单元胞体染色随着时间的推移不断增加强度。
注:通常不成功装载的原因是吸管堵塞是由于进入尖端或聚集染料集群组织。有时可以通过对录音室的下表面轻轻打破其尖端营救吸管。然而,枪头不应超过几微米。通过快速装载过程中施加压力较低的补偿更大的吸管开口。 - 装载的10分钟后,所施加的压力减小到零,并检查染色。
注意:染色区域的大小显著变化,并取决于几个参数,如注射染料和喷射站点的位置的量。一个良好的染色占地面积约100微米×100微米的区域。 - 如果污渍区域过小或没有覆盖所需的录制现场,重复步骤3.10-3.13。如果它尚未堵塞使用相同的微量为下一次注射。
- 在开始任何实验,让染率和脱酯化之前等待最后一次注射后至少30分钟。继续在任何时候都新鲜林格液灌注到录音室。
4.测量设置
- 确保该测量设置由具有足够的速度共焦显微镜来记录三维卷。选择每堆至少1 Hz的采集速率。
注:适用设置包括例如线照明显微镜扫描样品行明智的,而不是由对点oint。一个简单的实现这种设置的先前已描述6。其它选项旋转盘显微镜。如果没有这样的设置是可用的,它仍然是可能的与正常点扫描设置来测量较小的区域。 - 设置测量参数,使得足够大,以适应肾小球大小的体积可以被覆盖。
注意:记录卷的典型厚度是在其中应至少5层所覆盖的20微米的范围内。 - 检查所有漂白突触前测量。调整激光功率,直到所记录的图像的平均荧光强度不随记录的时间过程下降。
- 限制对突触后侧的记录,以避免漂白尽可能测量时间和面积,虽然测量超过20-30秒漂白是可能发生。
5.气味应用和温度实验
- 将25毫升新鲜林格氏溶液(预先保存在4℃),在50毫升的管中。把试管冰桶。
- 通过插入温度计的清洁探头插入管监视冷却林格氏的温度。等到温度低于1℃,开始实验前。
- 以10μM的浓度制备50毫升溶解在林格氏溶液L-组氨酸。
- 使该室中的水流量保持的刺激溶液的释放期间是恒定的和不间断的使用漏斗涂布器8或允许刺激递送伴随的灌注林格氏类似的应用的系统。漏斗以这样的方式定位,该远端出口是从嗅觉上皮小于1mm距离。
- 放置连接到数字温度计接近上皮和漏斗涂布器的出口的镍铬 - 镍温度传感器。接线温度计输出端口到一个计算机来记录和视觉DIS玩反映温度波动小的电压变化。
- 在实验开始前,另一个热传感器连接到一个标准温度计以建立电压 - 温度比例因子。
- 调查在该记录室中的浴温度,并确保其不超过22℃。
- 启动图像采集,并通过一个电子移液管顺序地应用200-400微升冷林格氏,L-组氨酸和室温林格氏(20-22℃)的20-30秒的刺激间隔。为了更好地控制刺激应用,释放的刺激与由所选择的成像设置发送到吸管如果可能的触发信号。重复再现的结果应用协议。
- 就拿套较长的录音与几轮的刺激,因为他们是最好的与活动相关成像成像后分析。
注:拍摄时间和切片生存能力之间的折中被发现。的约 2分钟覆盖6刺激应用测量提供对网络的良好重建足够的活性。
6.图像处理使用活动相关成像(ACI)
- 突触前录制的图像处理
- 与由冷林格溶液和组氨酸刺激的录音,可以通过不同的反应条件(参见Kludt 等人 5)区分从相邻组氨酸敏感肾小球温度敏感性γ-神经纤维。
- 创建在从脑制剂,其中ORNs已电穿孔用两种不同的染料以可视化的γ-肾小球的双边神经支配的录音轴向方向上的最大强度投影。
- 在突触后录音中使用活动相关成像的图像处理(ACI)
- 有足够的时间点(超过10个选择的录音0帧)和活动,像样的数目(至少两个事件)。
注意:活动可以是自发通过刺激多个应用程序在相同的记录过程中,揭示单个僧帽细胞的过程或引起以显示由嗅觉刺激激活了蜂窝网络。 - 选择的录音,其中所测量的结构在记录的时间过程移至不超过2-3个像素。
注:运动过多录音可通过应用Kludt 等人的转变校正程序被救出5。 - 检查是否录音从漂白苦。如果整个图像的平均强度下降在记录的时间过程,漂白校正是必要的,否则跳过下两个步骤。
- 通过进行线性回归计算每个像素的时间跟踪的线性趋势。
- 从每个像素减去的结果分别以消除线性的直流电阻漂白的吨。
注意:可以通过宝和席尔德9描述中使用的替代配件勒。 - 有一步一步的指导的活动相关成像(ACI)由Junek 等 人一起下载现成利用MATLAB脚本。6
注:按照Junek 等 6作为替代描述使用提供的MATLAB脚本的步骤。 - 从下载容器用于数据评价系统的MATLAB的路径移动文件'aci.m','matVis.m'和'aci_roiSelector.m'。
- 装载作为组织为[X,Y,Z,T] -矩阵与x和y所指的横向尺寸,Z,轴向方向和t的时间过程,以MATLAB的用户的工作区的可变在步骤5.8中获取的原始数据。
- 从MATLAB命令行调用"阿哲"。
- 在用户界面(UI)其中出现时,选择"准备数据",然后选择包含的数据,并在其中将结果保存在目录中的变量。
注:对于UI的完整说明请参阅手册伴随阿哲脚本。 - 通过测量Z-层由UI移动对应的滑块可以得到显示的变化映射的概述滚动。
- 输入感兴趣区域(ROI)到用户界面的区域的大小。要僧帽细胞胞体调节的ROI以跨越在轴向方向在横向大约10微米和5微米。对于肾小球,为20μm横向和10μm轴向略微较高的值是适当的。
注意:ROI大小必须输入作为像素数,这取决于选择用于记录的像素缩放。 - 用鼠标中键点击到CENTE选择包含周围僧帽细胞的每个可见SOMA的γ-肾小球和附加区域的投资回报率细胞/肾小球河
- 关闭触发相关的计算映射所有参考曲线的主界面。结果自动保存并显示出来。
- 有足够的时间点(超过10个选择的录音0帧)和活动,像样的数目(至少两个事件)。
结果
嗅觉受体神经元(ORNs)电穿孔与通过主动的轴突运输葡聚糖分子的顺行标记结合的Alexa Fluor染料或钙指标的实现。而前者的染料提供的感觉神经元和其轴突在玻壳肾小球层的支化鲜艳染色,后者允许神经元活性的这些细胞中的测定( 图1和2)。首先,热敏γ-肾小球其神经支配模式的位置,并通过电穿孔的Alexa Fluor 647葡聚糖和Alexa Fluor 546的葡聚糖在左和右的嗅觉上皮,分别( 图1A)可视化。手术后二十四小时,在鼻孔的ORNs,两个嗅觉神经和在两个半球肾小球荧光显微镜下也清晰可见。不同的肾小球簇是identifia由它们各自的位置BLE,特别是小簇包含γ-肾小球( 图1B)。对侧嗅纤维少数通过对侧嗅球跑了,越过了前连合和同侧γ-肾小球( 图2A)终止。
为了记录该γ-肾小球的突触前纤维的钙反应,钙绿葡聚糖电穿孔在ORNs,按照同样的程序。负温度跳跃被通过的冰冷林格氏溶液中的控制释放在鼻孔诱导(0-1℃)。包括γ-肾小球三维体积快线照明共聚焦显微镜下成像。的-1℃下ΔTs随足以触发该γ-肾小球冷的反应和其传入,识别为在ΔF/˚F 钙痕迹可逆峰( 图2B </ STRONG>,C)。
进一步的实验步骤进行了测量通过它们的枝状的树突末梢连接到γ-肾小球的僧帽细胞冷诱导的活性。的Alexa 647后这些突触后纤维和周围的神经纤维进行了有效的钙敏感染料的Fluo-8的AM丸药装载染色,进行几天葡聚糖已电穿孔进入ORNs( 图3A,B)。僧帽细胞充满了的Fluo-8 AM,以及嗅上皮被刺激根据以下范例两次:冷林格,组氨酸(10μM)和室温林格氏,随后施加。两个参考痕迹从感兴趣的区域取入记录有体积,一个专响应温度下降,另外一个,只到组氨酸。活动相关成像(ACI)6计算基于所选择的参考迹线具有高对比度可视化对应要么温度或组氨酸反应性的 Ca 2+信号( 图3C,D) 的突触后网络的树枝状形态。最后,热敏和化学敏感的地图是颜色编码和覆盖在彼此的顶部,示出了如何温度和化学信息是从单个肾小球传送到嗅觉二阶神经元( 图3E)。在共享嗅觉网络这两种类型的信息的整合和处理的描述,请参见Kludt 等 5

图1:ORN电穿孔和丸装载概述 ( 一 )X的两个鼻腔嗅觉受体神经元。 蟾幼虫与Alexa染料或calciu电耦合到葡聚糖分子间敏感的染料。荧光指标顺行传送,直到终端轴突分枝。电穿孔后24小时,在这两个半球肾小球层表现出荧光染色。嗅球在一个半球的细胞组织的(B)的示意图。肾小球层跨越集群灯泡:内侧,小,中间和横向集群。嗅觉信息从该受体的神经元传输经由肾小球兴奋性突触二尖瓣细胞。 Periglomerular细胞和粒细胞是抑制性神经元调制嗅觉处理和编码。所述γ-肾小球(青色)很容易被确定为小神经纤维,其中同侧(红色)和对侧(橙色)嗅觉纤维合并。 (℃)丸加载是在γ-肾小球的附近实现以染色突触后神经纤维主要由二尖瓣细胞及其dendri的抽动树在小球层大大分支。 请点击此处查看该图的放大版本。
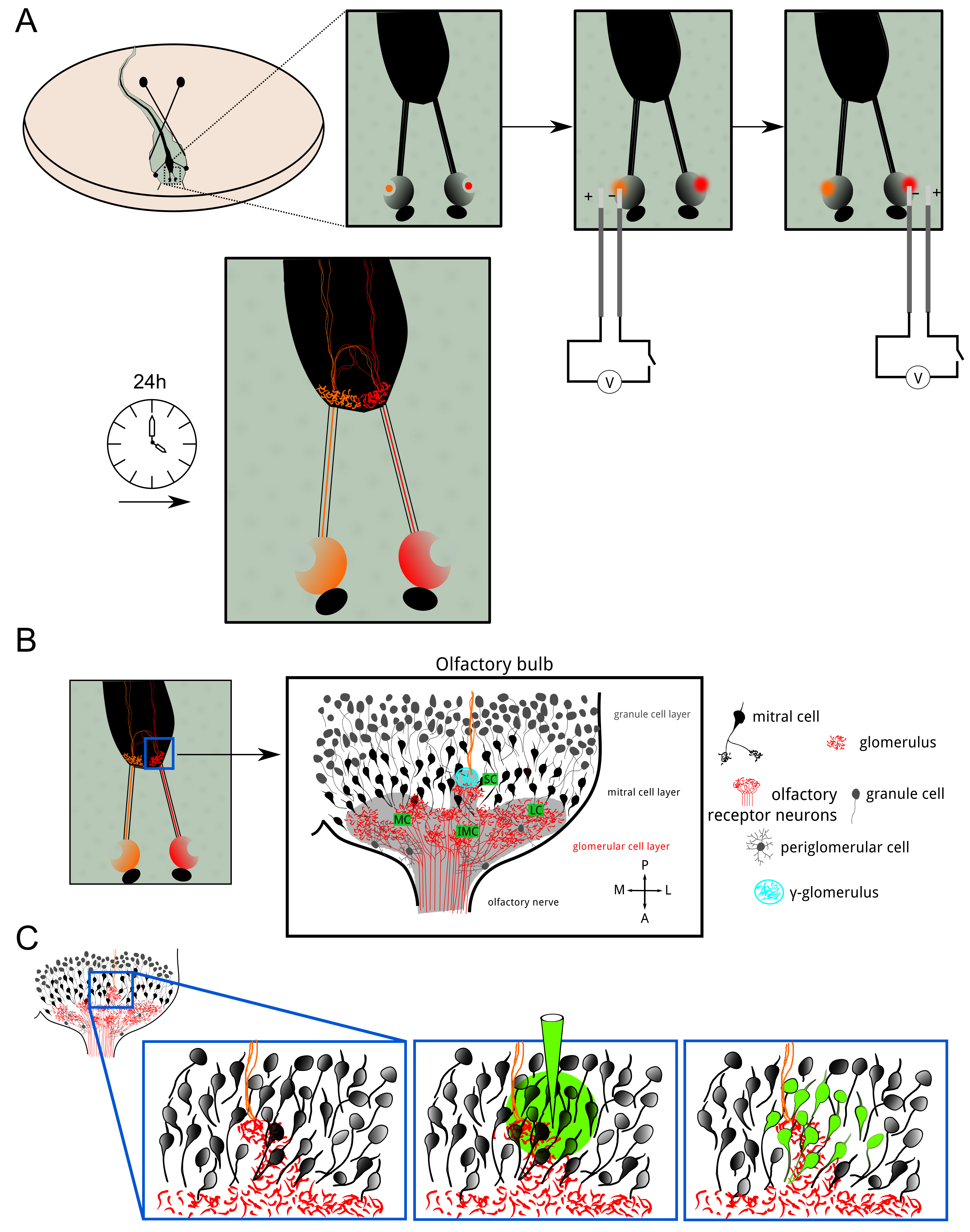{kind=link}

图2:ORNs的电穿孔揭示结构和功能连通性 (A)双边同侧(绿色)γ-肾小球的神经支配和对侧(红色)的嗅觉受体神经元(ORNs)。比例尺= 50微米。 (B)中与钙敏感染料钙绿葡聚糖电穿孔后嗅球。所述intermedial(IMC),内侧(MC)和小簇(SC)是可见的。该图像是100微米厚的测量容积的最大投影。比例尺= 50微米。 ( 三)小簇的特写图。所记录的卷(12微米)是在最大投射表示。图像描绘在灰色和彩色编码的ΔF/˚F地图作为覆盖基底荧光水平。刺激的最大响应冷林格氏液绘制。该γ-肾小球反应强烈,而两个相邻的肾小球保持沉默。插图显示了ΔF/˚F跟踪对应于感兴趣的指示的区域中的γ-肾小球。蓝色条代表了刺激的应用程序。比例尺= 20微米。 请点击此处查看该图的放大版本。
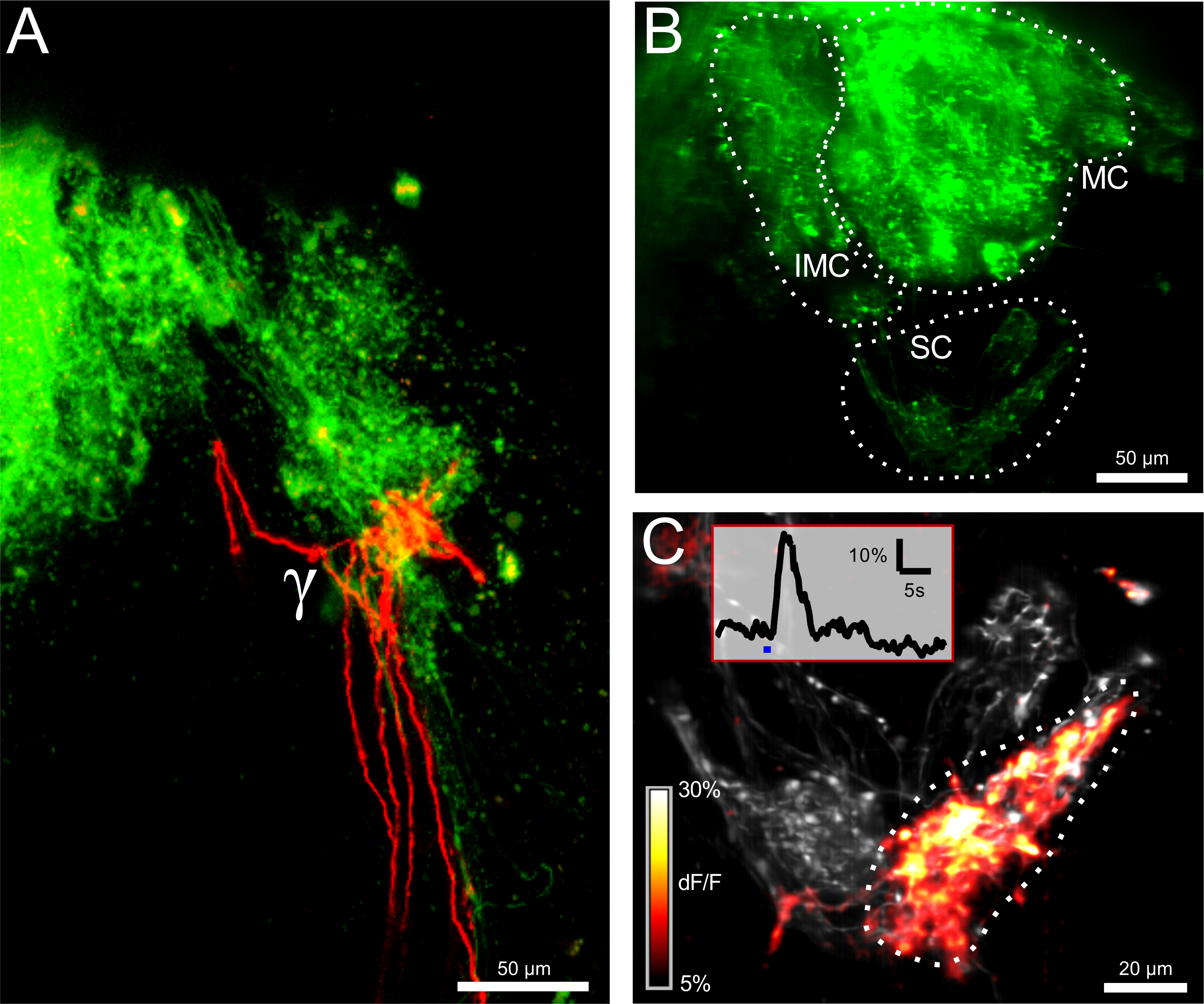{kind=link}

图3:ORNs小簇在端接的丸装载和ACI解离热敏和化疗敏感网络 (A)的轴突通过电穿孔与非卡尔奇染色嗯敏感的染料Alexa的647葡聚糖。虚线勾勒出γ-肾小球。在推注装载与钙敏感染料的Fluo-8点以后的第二测量信道的相同区域的(B)的图像作为在(A)中。有些僧帽细胞胞体是可见的,但对比是有限的。以下的箭头,两个响应迹线进行作图,它被用于活动相关成像(ACI)。蓝,迹线下面的红色和黑条描绘冷林格,组氨酸(10μM)和分别室温林格氏作为阴性对照,应用的开始。两个钙离子的痕迹,从所测量体积的兴趣不同区域取。 ( 三 )突出的地区主要是回应冷林格跟踪(B)的ACI结果。 ( 四 )突出的地区主要是响应组氨酸跟踪(B)的ACI结果。两个ACI地图(E)叠加。僧帽细胞响应为histidine和支配肾小球(红色)来自热敏僧帽细胞和γ-肾小球(青色),很容易分辨。这个数字的所有图像都是28微米厚的体积的最大强度的预测。比例尺= 20微米。 请点击此处查看该图的放大版本。
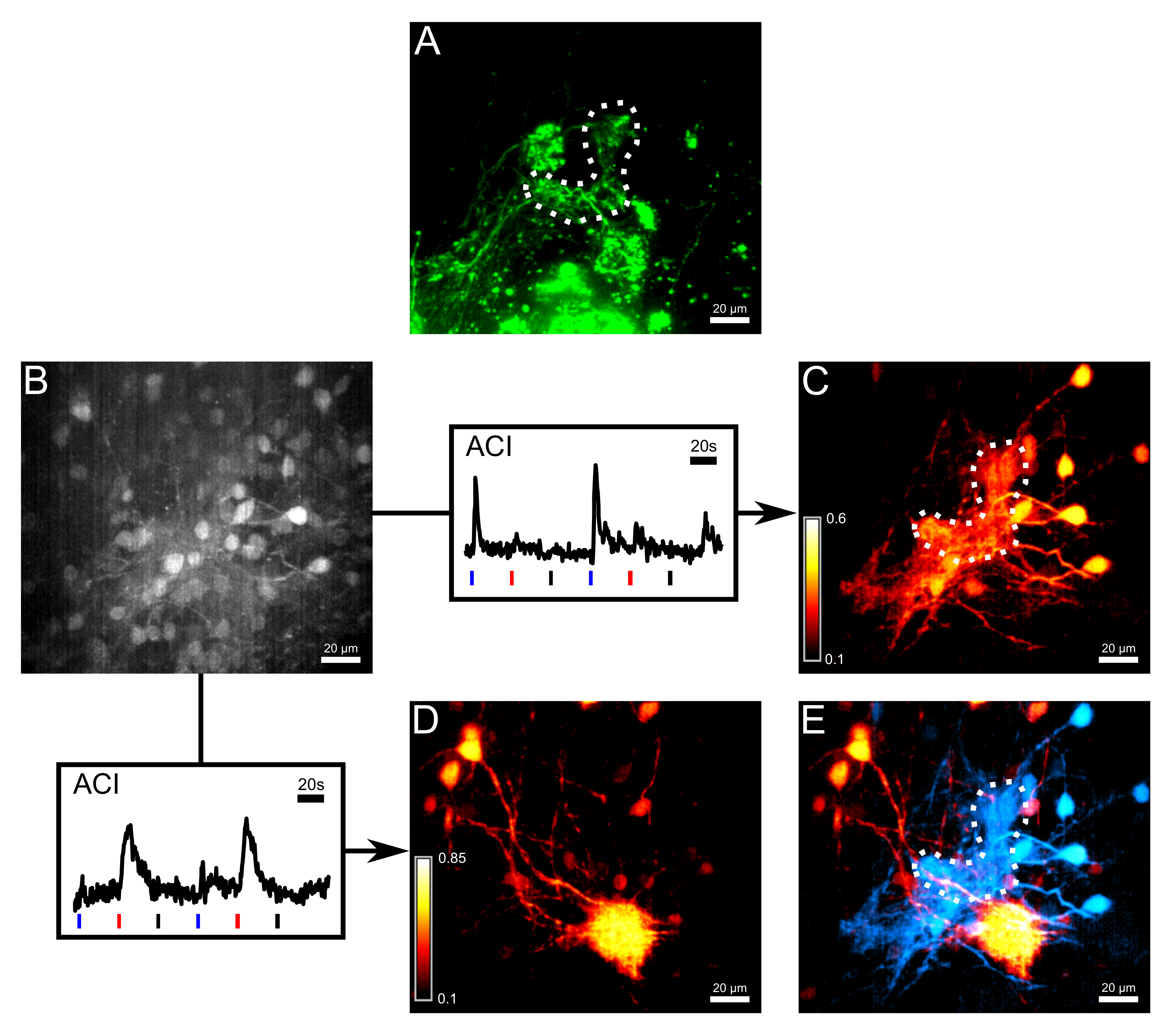{kind=link}
讨论
本文所提出的方法的目标是在记录温度处理在爪蟾的嗅球蟾蝌蚪。协议污渍第一和嗅球二阶神经元和提供了其中的嗅觉系统仍然主要完好样品制备。因此,温度敏感型γ-肾小球的活化可以监测和其化学 - 敏感相邻肾小球比较。此肾小球的独特双边支配由细胞电穿孔用频谱不同的染料显现。此外,大丸剂负荷允许僧帽细胞跨越嗅球内大量的染色。神经元网络处理温度诱导信号是通过服用钙测量反复刺激应用程序,随后与分析相关的活动成像数据显示。
该协议突出了两个先进的染色PROCE的既定程序,两者都需要谨慎操作和实践,以达到令人满意的和可重复的结果。在电穿孔动物的任何伤害已被避免,在电极定位成鼻孔时尤其如此。理想情况下,应该发生的嗅觉上皮细胞没有任何联系。请注意,动物是电穿孔手术后仍然生活和他们的恢复时间必须考虑在内。如果染色剩一个圆形电穿孔,它可以发生取决于类型中使用的染料的后过弱,其强度可通过在鼻孔提高染料浓度得到提高。由于右旋糖酐耦合分子通过多种机制,包括缓慢的轴突运输(在1-2毫米/10天速度)和被动扩散运输,另一种选择是牺牲动物前等待电后48小时。另外,电可以恢复一天后重复。
jove_content">丸装载是因为染料的量的一个关键步骤进入僧帽细胞是困难的调节和取决于各种参数,如该应用程序的枪头大小和位置。共聚焦荧光显微镜下监测过程证明是有用调节染料应用的持续时间,从而产生一个横穿制剂类似染色的结果。此外,先前电蝌蚪应该使用通过识别小簇的位置(包括γ-肾小球),以确定用于染料应用的最佳位置,最在测量过程中的关键步骤是,以避免样品的两个移位和漂白,移位可以通过仔细定位显微镜下林格氏流是可以避免的。至于限制所关注的区域的漂白,测量时间应减少到基本。与钙敏感染料丸药装载染色仅提供了非常相比之下有限,因为健康细胞通常患有低钙血症,从而表现出微弱的基础荧光。申请活动相关成像绕过基于类似的钙信号的活动和亮点结构产生反差此限制。这个采集后分析方法计算的兴趣(基准迹线)的选定区域中的钙信号和在3D体积的每个单独的像素之间的相关性因子。因此,极力得到的结果取决于所选择的参考曲线的活动模式。如果主要焦点是形象化僧帽细胞神经支配的图案,从自发的神经元活动导出的参考信号是优选的,并且选择最活跃僧帽细胞会产生最好的结果。为揭示僧帽细胞的化疗或热敏感的网络中,只包含要么组氨酸或冷林格反应参考曲线应选择。整个肾小球邻的选择ř僧帽细胞胞体作为关注可能并不总是提供一个明确的参考曲线,特别是如果响应于两个不同的刺激的结构躺在彼此的顶部区域。在这种情况下,常常是有用选择肾小球或细胞体作为感兴趣区域的较小的区域。
在过去的几十年中,电穿孔已经被描述为一种有效的方法来染色单个或多个小区11,12。此处它被用于特异性标记嗅觉受体神经元。葡聚糖共轭分子,得到最高的效率,而对于非钙敏感性染料,选择范围很广,包括通常在荧光显微镜13中使用的完整光谱。然而,这是成功电穿孔到嗅觉受体神经元钙敏感染料是在限定于钙绿葡聚糖的瞬间,而且如果仍的Fluo-4的葡聚糖市售的。此外,录音主要针对superficia仅嗅球的腹面上升层中,由于快速的测量技术的穿透深度是有限的。双光子成像可以部分地克服这一限制,但通常缺乏速度,并进一步限制可选择钙敏感染料的量。
我们在这里所描述的协议在嗅球测定温度引起的活性。脑中神经毡被扫描成一个三维体积来可视化涉及温度的嗅觉处理复杂的蜂窝网络。测定温度引起的活性嗅球已经非常最近报道5和需要特别定制的程序组合不同的技术。以上提出的技术的主要资产是数百细胞的三维成像是在大多数嗅觉系统保持完好的准备。这些优点放高要求的染色技术以及大脑p赔偿和成像。例如,细胞电穿孔和丸药加载命中大量细胞在嗅觉上皮和球,从而使完全蜂窝网络的可视化。此外,通过推注装载代替遗传编码荧光团的化学指标的递送使得能够在潜在的较大的组的物种的测量。像带AM染料浴温育其他替代,其中严重损坏嗅球片主要工作,留下完整的组织的只有几百微米。相比之下,在我们的协议中使用的整个装载制备确保例如使γ肾小球的双边支配保持不变,因此,录音采取在一个仍然有效的系统。最后,成像本身由线照明显微镜允许采集3D体积的完成。线照明显微镜是提供尽可能高的采集速率6共聚焦技术之一</ SUP>所必需的覆盖嗅球的很大一部分。可以使用较慢的采集系统,但有缺点,即所记录的卷的大小必须减小。近年来,其他方法进行快速图像获取已被开发,并且可以作为替代品使用14,15。然而,线照明显微镜仍然获得足够两者速度和分辨率的最简单的方法之一。这里如下一些信息作为选择适当的成像设置的指导方针。由于钙成像是从厚脑制剂中完成,设置应该提供体面共焦和目标应具有1.0或更高的数值孔径。对于一个基准点,用线照明显微镜拍摄的记录对应于所用的标准激光扫描显微镜具有0.5-1通风单元的针孔大小的图像。快速的采集速度是可取的。用厚度为20μm的体积覆盖至少5升艾尔斯,观点的100微米×100微米的横向场并为0.5μm或更小的像素尺寸应在每堆1 Hz的最小速度进行扫描。减少共焦可以提高计数的光子的数量,从而允许更快的收购如果有必要,但现在的记录更失焦光的缺点。然而,由于这样的方法增加了光学片的厚度,它实际上可以方便的ACI 6的应用后树突通过不同的z平面上的跟踪。
必要的工具,广泛地研究高温处理嗅球网络在此呈现。温度引起的活性是通过钙敏感的染料和两到达和来自γ-肾小球出发信号记录在第一和第二阶的神经元。此外,在何种程度上个别僧帽细胞处理化学和温度信息可以被评估。由于准备LEAVES嗅球完好,在嗅觉处理双边支配的作用,可以进一步研究。该过程也揭示是否以及如何热电偶和chemoinformation重叠嗅网5编码有用。最后,上述技术不限于在嗅球的温度响应的研究,但可应用于嗅觉系统的更一般的评价,特别是在大的三维体积蜂窝处理网络。丸药装载和活动相关成像是有力工具观察比较几十神经元的活性,这使得它们适用于不同的大脑的网络16。
披露声明
The authors have nothing to disclose.
致谢
This project was funded by the DFG Excellence Cluster 171, the Center for Nanoscale Microscopy and Molecular Physiology of the Brain, the Bernstein Center for Computational Neuroscience and the ENC-Network, an Erasmus Mundus Joint Doctoral Program. The authors thank Stephan Junek, Mihai Alevra and Guobin Bao for providing MATLAB codes and custom-written programs for image evaluation and data analysis.
材料
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Sodium chloride | Merck Millipore | 1064040500 | |
| Potassium chloride | Merck Millipore | 1049360250 | |
| Calcium chloride dihydrate | Merck Millipore | 1023820250 | |
| Magnesium chloride hexahydrate | Merck Millipore | 1058330250 | |
| D(+)-Glucose | Merck Millipore | 1083371000 | |
| Sodium pyruvate | Sigma-Aldrich | P2256 | |
| HEPES | Merck Millipore | 1101100250 | |
| Calcium Green 10 kDa Dextran | Thermo Fisher Scientific | C-3713 | |
| Dextran, Alexa Fluor 647 | Thermo Fisher Scientific | D-22914 | |
| Dextran, Alexa Fluor 546 | Thermo Fisher Scientific | D-22911 | |
| Fluo-8 AM | TEFlabs | 203 | |
| MK571 | Alexis Biochemicals | 340-021-M005 | |
| MS-222 | Sigma-Aldrich | E10521 | |
| Pluronic acid F-127 | Sigma-Aldrich | P2443 | powder |
| L-Histidine monohydrochloride monohydrate | Sigma-Aldrich | 53370 | |
| DMSO | Merck Millipore | 1029522500 | |
| Equipment | |||
| Electronic pipette | BrandTech | HandyStep Electronic Repeating Pipette | |
| NiCr-Ni thermocouple | Greisinger Elektronik | GTF 300 | |
| Micropipette puller | Narishige | Model PC-10 | two-step puller |
| Funnel applicator | (Custom-made) | ||
| Line-illumination microscope | (Custom-made) | otherwise, a commercially available spinning disk microscope | |
| Objective W Plan-Apochromat 63X/1.0 | Zeiss | 441470-9900-000 | |
| Objective W Plan-Apochromat 40X/1.0 DIC | Zeiss | 441452-9900-000 | |
| Software | |||
| MATLAB | The MathWorks | from R2010b upwards |
参考文献
- Mamasuew, K., Breer, H., Fleischer, J. Grueneberg ganglion neurons respond to cool ambient temperatures. Eur J Neurosci. 28 (9), 1775-1785 (2008).
- Brechbühl, J., Moine, F., Broillet, M. -C. Mouse Grueneberg ganglion neurons share molecular and functional features with C. elegans amphid neurons. Front. Behav. Neurosci. 7 (193), (2013).
- Gaudin, A., Gascuel, J. 3D atlas describing the ontogenic evolution of the primary olfactory projections in the olfactory bulb of Xenopus laevis. J Comp Neurol. 489 (4), 403-424 (2005).
- Stosiek, C., Garaschuk, O., Holthoff, K., Konnerth, A. In vivo two-photon calcium imaging of neuronal networks. Proc Natl Acad Sci U S A. 100 (12), 7319-7324 (2003).
- Kludt, E., Okom, C., Brinkmann, A., Schild, D. Integrating temperature with odor processing in the olfactory bulb. J Neurosci. 35 (20), 7892-7902 (2015).
- Junek, S., Chen, T. W., Alevra, M., Schild, D. Activity correlation imaging: visualizing function and structure of neuronal populations. Biophys J. 96 (9), 3801-3809 (2009).
- Nieuwkoop, P. D., Faber, J. Normal Table of Xenopus laevis. , Garland Publishing Inc. New York. (1994).
- Schild, D. A computer-controlled device for the application of odors to aquatic animals. J Electrophysiol Tec. 12 (2), 71-79 (1985).
- Bao, G., Schild, D. Fast and Accurate Fitting and Filtering of Noisy Exponentials in Legendre Space. PLoSONE. 9 (3), (2014).
- Terasaki, M., Schmidek, A., Galbraith, J. A., Gallant, P. E., Reese, T. S. Transport of cytoskeletal elements in the squid giant axon. Proc Natl Acad Sci U S A. 92 (25), 11500-11503 (1995).
- Haas, K., Sin, W. C., Javaherian, A., Li, Z., Cline, H. T. Single-Cell Electroporationfor Gene Transfer In Vivo. Neuron. 29 (3), 583-591 (2001).
- Haas, K., Jensen, K., Sin, W. C., Foa, L., Cline, H. T. Targeted electroporation in Xenopus tadpoles in vivo-from single cells to the entire brain. Differentiation. 70 (4-5), 148-154 (2002).
- Paredes, R. M., Etzler, J. C., Watts, L. T., Lechleiter, J. D. Chemical Calcium Indicators. Methods. 46 (3), 143-151 (2008).
- Salome, R., et al. Ultrafast random-access scanning in two-photon microscopy using acousto-optic deflectors. J Neurosci Methods. 154 (1), 161-174 (2006).
- Keller, P. J., Ahrens, M. B., Freeman, J. Light-sheet imaging for systems neuroscience. Nature Methods. 12 (1), 27-29 (2015).
- Hjorth, J. J., et al. Detection of silent cells, synchronization and modulatory activity in developing cellular networks. Dev Neurobiol. , (2015).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。