
Method Article
转录分析的新生RNA FISH
摘要
Trophoblast giant cells (TGCs) play a key role in the placenta to ensure a healthy pregnancy. We present a protocol for assessing the transcriptional status of genes in TGCs by nascent fluorescent in situ hybridization on cryostat sections of post-implantation embryos or short-term cultures of embryonic day 7 ectoplacental cones.
摘要
胎盘从一个胚外谱系,滋养外胚层派生的。在围植入鼠胚泡,壁画滋养外胚层细胞分化为初级滋养层巨细胞(TGCs),而极性滋养外胚层覆盖的内细胞团继续增殖后分化成次级TGCs。 TGCs在发展胎盘关键作用,并且是一个成功的妊娠是必不可少的。植入后发育过程中特定基因的转录调控的研究能给见解TGCs发展。从胚胎在7-7.5天妊娠(E7-7.5)的ectoplacental锥体(EPC),由该极性滋养外胚层衍生的细胞,分化成次级TGCs 1。 TGCs 可原位研究,对在E7胚胎冷冻切片虽然TGCs的数目是在该阶段非常低。分析次级TGCs的一个替代方法是使用单个的EPCs的短期培养物从E7胚胎秒。我们提出了一个技术研究的在体内和在使用原位杂交(RNA FISH)荧光可视化新生转录物的单细胞水平体外兴趣基因的转录状态。该技术提供了基因表达的直接读出,使的TGCs的染色体状态,这是大endoreplicating细胞的评估。事实上,TGCs终末分化的关键特征是它们退出细胞周期并经过多轮的endoreplication.This方法可以适用于检测从常染色体和/或性染色体表达的任何基因的表达,并可以提供重要的信息纳入发育机制以及胎盘疾病。
引言
哺乳动物滋养层巨细胞(TGCs)构成孕产妇和胚胎组织之间的屏障。他们调解注入和孕的侵入子宫和在发展中发挥关键作用的胎盘的形成。他们生产的多种生长因子(细胞因子)和催乳素/胎盘生乳素的家人和类固醇激素,必要时作胚胎生长和生存的激素。 TGCs是大的,单核和多倍体细胞与细胞周期,endocycle,即由交替S期和G2阶段。事实上,TGCs被endoreplicating细胞,能够进行多轮DNA合成,无任何分部2。为了研究基因在体内的转录状态,在TGCs相比其他胚胎和胚外细胞类型,新生RNA FISH可以在体内对在特定植入后阶段冷冻切片3进行。 TGCs上由于兵卫部分容易辨认可以记录ř大尺寸和它们的转录基因活性,但它们在早期植入后的阶段数是低的。在E7胚胎切片TGCs的缺乏,促使我们进行胚胎滋养外胚层组织的短期文化,以研究滋养外胚层发育过程中基因表达的调节,以获得差异化的TGCs。此外,为了保持生理越好,建立的细胞系即滋养层干细胞(TS)并不总是合适的调查发育机制代次级TGCs提供有价值的工具来研究由于异常基因与TGCs缺陷有关的病理的怀孕调节小鼠。
在ectoplacental锥(EPC)的滋养层细胞是次要TGCs 4的前体。培养的内皮祖细胞向次级TGCs的自发分化先前已报道5。然而,在对比主TGCs,在二级TGC differen研究tiation仍然有限,presumablydue到分离不含任何母亲或胚胎组织的EPC外植体的困难。我们适于这些方法中,为了对从个别胚在E7,其中TGCs很少,但可以从EPC前体产生的发育植入后阶段衍生的次级TGCs执行的RNA原位杂交。 RNA原位杂交分析核初级转录从未在二次TGCs单个细胞水平的水平已经完成。这允许转录的精确分析,并用来显示TGCs在植入后的阶段3的后生不稳定。
哺乳动物表观遗传学的一个典型例子,在X染色体失活(XCI)在听说实验室6进行了研究。在这个过程中女性的2 X染色体中的一个被失活。非编码Xist基因转录物涂覆X染色体从它所在雌性细胞中表达,并触发大多数基因的沉默。利用RNA FISH,的X连锁的基因新生转录物可被调查作为可以Xist基因 RNA的失活的X染色体(十一)上的积累。我们在这里描述对植入后胚胎的部分和EPC的短期培养物进行的RNA原位杂交的过程。该协议改编自那些被用来研究在XCI女性鉴别胚胎干细胞和植入前胚胎7-11。我们提供XCI的例子在雌性胚体内 ,以及在体外 TGCs。
研究方案
按照核准的体制照顾动物进行动物的程序和使用的居里研究所(CEEA-IC)协议(C 75-05-18)委员会。这项工作也得到了来自法国教育部高等教育和研究的批准为使用转基因生物(协议编号5549CA-I)的下进行的。
1.冰冻切片的制备
- 从自然排卵F1的C57BL / 6×DBA / 2J小鼠中谢伊等人 ,12描述收集胚胎。在妊娠(E7)的第7天,颈椎脱臼牺牲8-12周龄鼠标。收集整个孕即蜕膜12。
- 隔离E7 conceptuses,如乳木果等人所述,12。将它们放置在PBS含60毫米的培养皿。
- 冻结E7鼠标孕的冰冻切片。
- 准备适应铝箔井1cm的高度('自制'使用玻璃巴斯德点子等)。沉积在该小容器的底部组织冷冻介质的下降。存款孕正确的方向(即纵向段的孕应在其水平方向保持)。
- 填写好包含与组织冷冻介质孕。然后浸泡在液体N 2块几秒钟,直到它变成白色-为了让它慢慢地冻结在上面的蒸气液态氮镊子将其挂起。接着,转移块成冷冻小瓶中,并存储在-80℃下(可存储数月)。
- 冷冻切片之前,将含有在-20℃的孕在低温恒温器30分钟将冷冻块。执行8微米厚的冷冻切片。幻灯片上的存款4部分,以实现有效的附件。放置节接近足以容纳一个18×18 平方毫米盖玻片下。
- 检查的质量切片(完整和无划痕)和胚胎(纵向部分)用立体显微镜的取向,并选择合适的用于进一步分析的部分。尽可能快地将它们存放在罐子科普林,进行RNA原位杂交(参见3.3.2节)。
2.继发TGCs的制备
- 隔离胎体(见1.1和1.2)。
- 解剖E7 Ectoplacental锥(EPC)
- 利用体视显微镜和含有无菌PBS的培养皿。解剖钳蜕膜。皮尔斯样品和开放的镊子撕蜕膜双方分开。
- 掏出胚胎。在剪刀状闭合动作罚款钳(杜蒙第5号)的使用技巧分离从胚胎适当的EPC。使用特别小心,以获得一个非常干净的样品,从母体组织以及来自绒毛膜和卵黄囊分离。在PBS洗出EPC外植体。
- 随着一点点的勺子,个别剖分EPC转移到4 WEL含在培养基中的无菌盖玻片升盘。
- 从内皮祖细胞短期培养派生TGCs。
- 制备含有盖玻片4孔板中。浸渍在乙醇中,接着火焰干燥灭菌直径为12毫米的圆形盖玻片。
- 0.5毫升EPC介质添加到每个孔中(EPC培养基:RPMI 1640中补充有15%FCS,0.1mM的2-巯基乙醇和抗生素)。
- 存款单,解剖,内皮祖细胞在盖玻片在井的中心与培养基一起。用细镊子适用于玻璃盖玻片的EPC。 Culturefor在37℃下3-5天,5% 的 CO 2。个别外植体形成了利差扁平TGCs( 图2A)的单层的产物。
3. RNA FISH
注:协议是基于那些绍梅等人,8和博动和听见13为胚胎干细胞中所述(胚胎干细胞)。
- 先进的准备工作股票方案的比
- 制备3M乙酸钠缓冲液pH 5.2。
- 制备含有40%2×杂交缓冲液(重量/体积)葡聚糖硫酸钠,20×BSA,400mM的氧钒核糖核苷复合物(VRC)在4倍盐水柠檬酸钠(SSC)。
- 制备含90%(V / V)甘油,0.1%的封固剂(重量/体积)对苯二胺,在PBS pH为9。
- 新鲜烹制的解决方案
- 制备由3%新制备的多聚甲醛(PFA)的PBS的固定液。制备含有0.5%的Triton-X-100的PBS透化溶液补充了2mM VRC。通过混合50%甲酰胺(FA)和2×SSC准备洗涤缓冲液和pH调节至7.2-7.4。
- 制备的DNA为1微克/毫升的DAPI(4',6-二脒基-2-苯基二盐酸盐)在2×SSC的反染色溶液。
- 固定和渗透在鱼的制备
- 对盖玻片细胞,洗涤细胞在1X PBS,持续5分钟在,固定在盖玻片,或胚胎细胞的部分幻灯片上,在固定液10分钟,在RT(3%PFA)解决方案。
- 冲洗细胞三次用1×PBS。通透在冰冰冷通透的解决方案5分钟的细胞。洗涤细胞,用70%的乙醇三次。在4孔板商店盖玻片和幻灯片在70%的乙醇-4-滑动运输箱在-20℃。
注:盖玻片和幻灯片可存放在-20℃几个月。板和含有它们的盒子必须用石蜡膜密封,以避免乙醇蒸发。
- DNA探针标记
- 通过使用荧光核苷酸缺口平移标记的DNA探针。按照制造商的说明( 见表1)8。
- 对于50微升反应混合物,加1-2质粒微克,细菌人工染色体(BAC)或多重置换扩增(MDA)(DNA扩增;参见3.4.4节)的DNA 17.5微升水,2.5微升0.2毫米SR-,SG-,Cy5的-dUTP的,10微升的10mM各dNTP混合物(dGTP,的dATP,dCTP),5微升的10mM dTTP的,5微升10×缺口翻译缓冲液和8微升切口平移酶。
- 在黑暗中孵育16小时在15℃。灭活通过在-20℃冷冻该反应。商店探针在-20℃个月。
- MDA
注:对于BAC DNA,古典制备后得到的DNA的量是低的,因此人们可通过使用MDA使用DNA扩增的步骤标记之前。使用商品化试剂盒,并按照制造商的说明( 见表1)。 - 混合0.5微升BAC DNA与9.5微升样品缓冲液。加热在95℃下3分钟。立即淬火冰上;留在冰上10分钟。准备反应缓冲液/酶混合物:(9.5微升+ 0.5微升酶混合物),并保持在冰上。
- 混合10微升DNA混合+ 10微升反应缓冲液/酶混合物。孵育在30℃下至少20小时。灭活通过加热样品酶10分钟在65#176℃。酷样品4℃-20℃存放之前。
- 通过消化检查MDA。加入1μlMDA,2微升10X缓冲液,2微升Hind III和15微升H 2 O.在37℃孵育过夜。在0.8-1%琼脂糖凝胶运行缓慢,以确认准确的DNA扩增。高分子量的明确的DNA片段应在胶上观察到。
- 探针制备
- 使用每个盖玻片或幻灯片0.1或1微克探头。
注:加婴儿床-1 DNA的2-5微克,如果竞争需要例如,大多数探头,因为它们可以包含否则交叉杂交的重复序列,增加的背景。- 对于沉淀,添加5微克鲑鱼精子DNA,1/10体积的3M乙酸钠pH为5.2和3体积乙醇。自旋16,000 xg离心4℃25分钟。用70%乙醇洗涤颗粒和5分钟再次离心。
- 干粒料用于在集中器/速度真空2分钟。悬浮在100%的形式酰胺半杂交所需的体积( 如 2.5微升的盖玻片或7微升胚胎的部分幻灯片)。放置在37℃30分钟,在恒温摇动。在75℃下变性7分钟。
- 淬火冰,或者如果竞争需要直接放于37℃30-60分钟。混合用等体积的2×杂交液探针溶液。
- 使用每个盖玻片或幻灯片0.1或1微克探头。
- 杂交和洗涤
- 脱水;盖玻片井,和幻灯片在科普林氏缸,通过连续洗涤1X中的80%,1个95%和2x 100%乙醇每次5分钟。干盖玻片和幻灯片完全。
- 对于杂交,应用5微升探针杂交组合(见3.5)到滑动并降低盖玻片,面对进杂交组合的细胞。对于滑动部分,适用于14微升探针杂交组合(见3.5),并与一个18×18 平方毫米盖玻片。
- 在潮湿的地方室玻片(纸巾浸泡在50%FA / 2X SSC),并在37℃孵育过夜,在黑暗中。
- 杂交后洗涤:
- 加入1毫升50%FA / 2X SSC到盖玻片上的幻灯片松开;从幻灯片小心取出盖玻片(注意不要刮去细胞)和高达其放置单元侧到含有50%的FA / 2×SSC中4孔板中。上滑动的部分,除去盖玻片以类似的方式,并放置在滑动中含有50%的FA / 2×SSC中科普林缸。
- 在42℃或44℃下进行3次洗涤,每次7分钟,预热50%甲酸/ 2×SSC中盖玻片或幻灯片,分别。
- 用预温2X SSC进行3次,每次5分钟,在42℃或44℃的盖玻片或幻灯片,分别。
- 染液通过洗涤核在2×SSC用1μg/ ml的DAPI在RT 3分钟。用2×SSC冲洗两次。
- 盖玻片和幻灯片的安装
- 从培养TGCs小盖玻片,适用5&#181,L在幻灯片上安装网上平台。放置盖玻片细胞侧朝下放在滴的顶部。
- 对于部分滑道,适用15微升安装在幻灯片中。放置在下降顶部的22×22 平方毫米的盖玻片。
- 避免气泡。擦去多余的安装解决方案。密封盖玻片指甲油少量。如果可能的话,图像在-20℃立即或储存长达滑动到几个月。
4.显微镜和分析
- 在0.3微米获得3D顺序Z轴的图像使用荧光显微镜(63X物镜),并执行使用ImageJ的软件14的三维图像栈的分析。
结果
TGCs可以一经DAPI染色E7部分被识别,由于其在孕本地化和它们的大尺寸。这在图1A中所示的一个纵向截面。 RNA的鱼为了研究X染色体失活在这个胚外谱系这种胚胎切片上进行。
几个幻灯片或盖玻片可以同时被处理。通过使用不同的染料来标记不同的基因的RNA,可以在相同的核检测不同初级转录。至少2个探头可以混合在一起,例如Xist基因耦合到SG(绿色信号),并且ATRX耦合到SR(红色信号)。阴TGC的一个例子示于图1B,而其他2谱系被表示,胚胎正确(E)和内脏内胚层(VE)。一个TGC核显示Xist的与RNA覆盖在X CHROMosome被灭活(十一),而其他的X染色体哪个是活动的(XA)显示了一些针点。 X连锁基因ATRX初级转录是从因endoreplication几个副本活动的X染色体(不Xist基因饰)表示。这说明单等位基因表达比如,一个X染色体上的ATRX灭活。
因为如在图2B所示TGCs在大小异质的,只有最大的那些被记录。三个探针,也可以使用如在图2C中示出了用于辅助TGCs。在这种情况下,Xist基因 -SG(绿),两个X连锁基因:G6PD -SR(红色)和Huwe1 -Cy(品红色)分别在同一核同时进行分析。虽然G6PD是monoallelically表示如下所示(之前在二级TGCs 3所示)Huwe1是biallelically表示展示其公关operty逃脱XCI。 Endoreplication,有几个精确定位,即,新生的成绩单副本,也可见一斑。

从E7女性胚胎部分TGCs图1.初级转录表达。(A)用DAPI的E7孕的纵切面。胚胎和胚外组织中由母亲组织,蜕膜包围。不同谱系的定位是建立在DAPI染色5X目标完成。 TGCs是由于其大尺寸标识。 CH,绒毛膜,E,胚胎,EPC,ectoplacental锥,VE,内脏内胚层,TGC,滋养层巨细胞。比例尺= 100微米。绿色(B)在(63X物镜)盒装入区的高倍RNA鱼。ATRX初级转录红色和Xist基因 RNA是可视化的3系= E,VE,TGC。钪啤酒吧= 10微米。 体内 TGC哪里ATRX被monoallelically表达(XA,箭头)和无声的Xist的涂覆的X染色体上(十一)的实施例;看到了几个Xa的信号ATRX是由于endoreplication。ATRX = BAC克隆RP23-260I15。 请点击此处查看该图的放大版本。
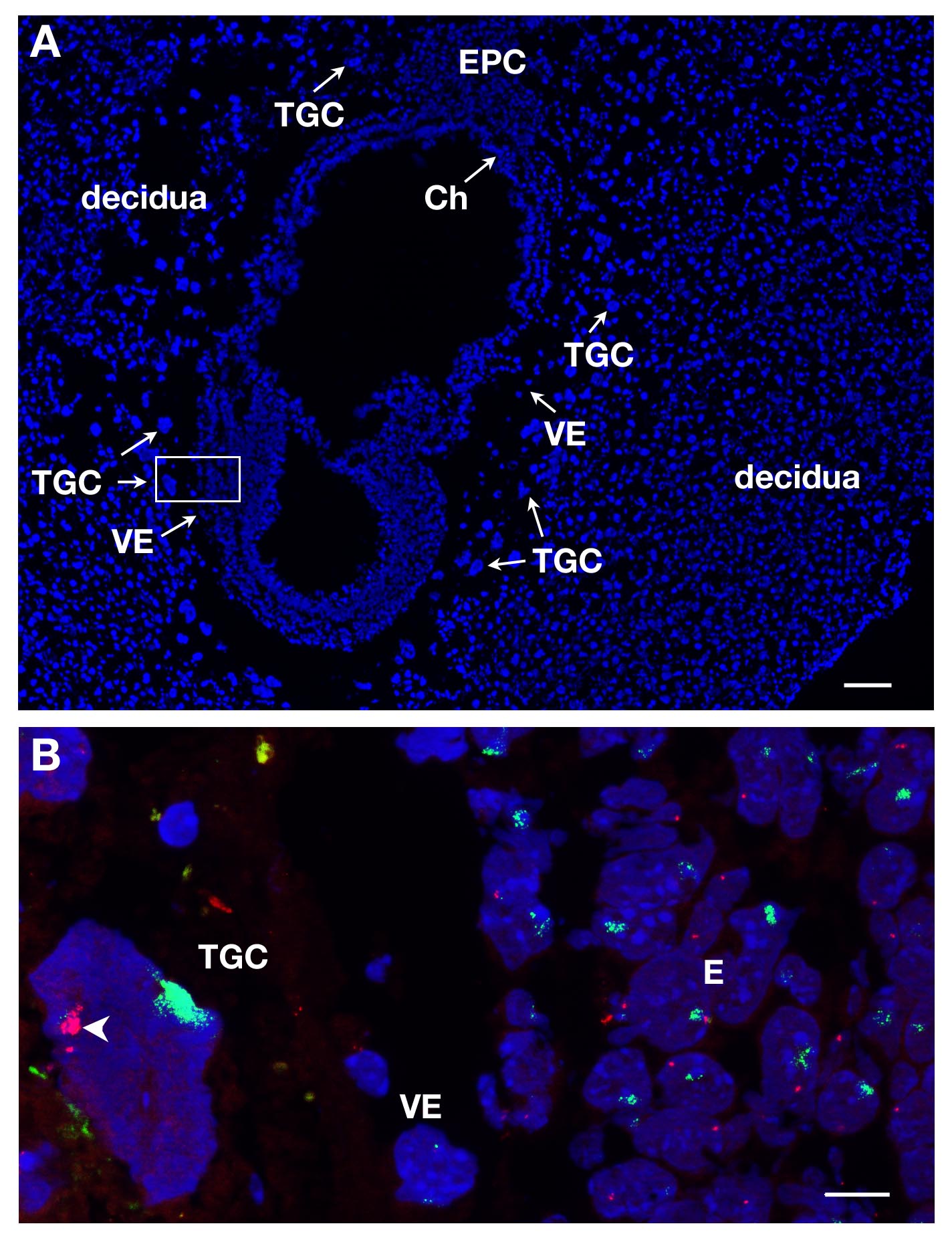{kind=link}

从EPC派生二次TGCs图2.初级转录表达。(A)成长辅助TGCs(X5目标)的一般看法。比例尺= 100微米。 (B)星号表示根据自己的体型(10X物镜)例如TGCs的。比例尺= 60微米。二级女性TGC RNA鱼使用3个探头的(C)示例(DOMA Xist的在绿色,覆盖失活X染色体:西安),显示G6PD和Huwe1初级转录的endoreplication(几针点,在红色和品红色)。虽然G6PD是monoallelically表示,Huwe1表现在这个核心等位基因Huwe1 = BAC克隆RP24-157H12; G6PD = BAC克隆RP23-13D21。夕=箭头; XA =箭头(63X的目标)。比例尺= 10微米。 请点击此处查看该图的放大版本。
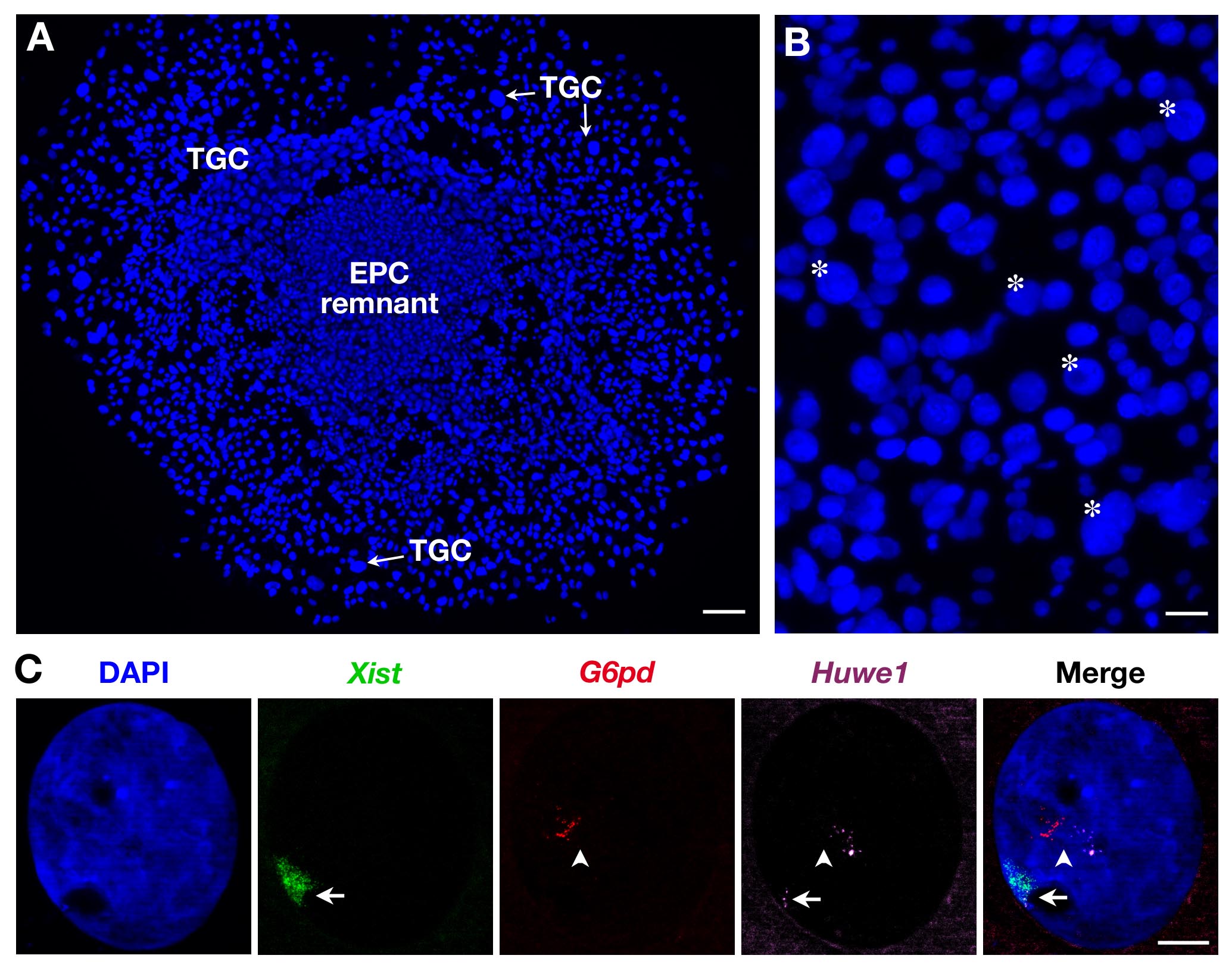{kind=link}
讨论
新生RNA FISH表示用于在不同发育阶段的胚胎组织转录活性的单细胞分析的简单和灵敏的方法。这种方法的功率是根据形态学标准在任何特定阶段,以确定不同的胚细胞系的能力。然而,这也需要最小的背景荧光的存在。任何这样的背景呈现不同胚胎区域和细胞类型具有挑战性的识别。为了保证最小的背景,也有这个协议的两个关键步骤。第一是冷冻切片的质量和第二个是RNA鱼,这取决于基因表达的水平和探针的质量的效率(信噪比)。对于后者,探针因此一直在培养的细胞上测试盖玻片(胚胎干细胞或体细胞),以上的部分的使用之前。
在这个协议中,我们提供日所需的E.技术由两种不同类型的样品制备的(冷冻切片和胚胎外植体的原代培养物)上TGCs进行RNA的FISH分析。这样的单细胞分析使得能够在不同的植入后阶段的基因表达的动态变化进行评估。
在我们的实验室中使用,我们描述生成次级TGCs(在E7-7.5植入后级)的方法,研究其在植入后级构成TGCs的胚外谱系的X染色体失活模式。在啮齿类动物外胚胎发育取决于TGCs的分化。事实上,TGCs是胎盘和胚胎由此发展至关重要。在TGC分化缺陷导致胚胎死亡(综述见参考15)。使用RNA FISH TGCs的转录活性允许我们小鼠发展3中表现出这样的细胞类型的一个不寻常的X染色体失活状态。
这里提出,以获得和研究二次TGCs的方法可用于研究分子途径中的重要胚外组织他们的一部分的展开,无论是在正常和突变的小鼠。这些方法可以适用于研究在其它哺乳动物TGC发展。我们的分析涉及使用野生型小鼠胚胎的允许我们评估在体内 TGCs并在从EPC植衍生体外 TGCs各种基因的转录活性。该方法可以扩展到转基因小鼠和/或通过在培养基中加入抑制剂分子的分析。我们已经成功地使用在其他类型TGCs的RNA原位杂交的方法中,诸如出现在小鼠发育,E3.0囊胚的较早阶段主TGCs,它可以是4-5天期间开发的生长,在单独培养的ICM受到大的初级TGCs 16,17包围。针对不同的新生成绩单基因以及Xist基因可以被可视化,并使用相同的RNA原位杂交的方法3量化。
合并的免疫荧光和RNA FISH也可以在冷冻切片在体外培养TGCs 3进行为好。这表明,该方法是相当健壮,因为初级转录物是容易退化且有无RNA酶的化合物的绝对要求。中频/ RNA的鱼提供关于转录水平,细胞定位和蛋白的表达,在一个给定的小区同时附加信息。虽然,石蜡包埋的组织切片先前已用于检测在人类肿瘤新生RNA,同时保持组织形态18中 ,在我们手中,冷冻切片更适合用于保存既通过免疫荧光中抗体检测所需的新生RNA和表位。
除了RNA FISH,以下IMmunofluorescence,染色体DNA FISH也可以使用标记有荧光染料,类似于那些用于RNA FISH 3探针次级TGCs进行。荧光探针可以是质粒/ F粘粒或BAC的,标记有荧光dUTPs如这里使用的,以及在绍梅等人 ,8说明。备选地,荧光标记的寡核苷酸可用于19。由于DNA的FISH不需要链特异性,寡核苷酸探针可以被设计为靶向或两条互补链中的靶区域。支链的DNA探针,其中信号放大是由两个连续的轮具体和放大器探针实现也可以用来增强的FISH信号20的信噪比。可替代地,RNA探针如核糖核酸探针可以使用尽管在我们的基于DNA的探针保证信号质量,特异性和易用性之间的更好折衷。
最后,3D图像acquisiti上是为了获得在这些大的细胞所需的空间信息必不可少的,并且可以使用各种荧光显微镜,如Apotome显微镜或表面荧光显微镜与去卷积如Deltavision(GEH)来执行,或其它显微镜合适成像组织切片,大(> 20微米)TGCs。应当注意的是,对于单拷贝DNA基因座,通过新生RNA FISH或DNA FISH检测的点状信号可能无法被容易地通过共聚焦显微镜检测。
总之,我们在这里描述的方法应是有益的既为TGS的中一个发展方面的详细的分析,也可在其他疾病的情况。所涉及的TGC发育和功能在啮齿类动物中的许多基因啮齿动物和人类中,如转录因子,蛋白酶和细胞粘附分子21之间是保守的。鼠标TGCs是调节胎盘发育的研究基因细胞模型因此先给见解人胎盘疾病。此外,由于这样的事实,这些细胞endoreplicating并且由于一些癌细胞接合endocycle方案中,除了基因扩增过程中,我们描述了方法也应在探索机制导致基因组的癌细胞的不稳定性所需的单细胞调查有帮助。
披露声明
The authors have nothing to disclose.
致谢
我们感谢苏菲Gournet与插图的帮助下,朱莉绍梅阅读手稿,动物住房设施和单位的成像平台。这项工作得到了支持程序«INVESTISSEMENTS D'艾文莉»下由法国政府发起,并与参考ANR-10的LabX-0044和ANR-10 IDEX-0001-02 PSL,该EpiGeneSys FP7没有用ANR实现。 257082网络卓越为EH,一个ERC高级研究者奖没有。 250367和欧盟FP7 SYBOSS不授予。 242129到EH作者要感谢居里研究所,法国生物成像(ANR-10-INSB-04)的成员的遗传与发育生物学系(UMR3215 / U934)的细胞和组织成像平台与光的帮助显微镜。
材料
| Name | Company | Catalog Number | Comments |
| Stereomicroscope | Nikon | SMZ 1500 | |
| Stereomicroscope | Zeiss | Stemi SV6 | |
| Scissors Pascheff-Wolff | Moria | MC19 | |
| Dumont #5 forceps | Roth | PK78.1 | |
| 4-well tissue culture dishes | Nunc | 176740 | |
| 60 mm Petri dishes Falcon | Dutsher, France | 353004 | |
| 100 mm Petri dishes Falcon | Dutsher, France | 353003 | |
| Coverslips 18 mm x 18 mm | VWR | 631-1331 | |
| Coverslips 22 mm x 22 mm | VWR | 631-0125 | |
| 12 mm glass round coverslips | Harvard apparatus | 64-0712 | |
| Slides Superfrost plus | VWR | 631-9483 | |
| 4-slide Transport box Lockmailer | Dutsher, France | 40684 | |
| Cryotubes 1.8 ml Corning | Fisher Science | 10418571 | |
| Glass Coplin staining jars | Fischer Scientific | W1561L | |
| TissueTek O.C.T compound | VWR | 4583 | |
| RPMI 1640 medium | Invitrogen | 61870 | |
| Phosphate buffered saline (PBS) | Sigma-Aldrich | D1408 | 10x is used |
| Water | Sigma-Aldrich | W3500 | |
| Paraformaldehyde | Panreac Quimica, Spain | 141451 | 3% in PBS |
| Triton-X-100 | Euromedex | 2000-A | 0.5% final |
| Vanadyl ribonucleoside complex (VRC) | New England Biolabs, USA | S1402S | |
| Sodium dextran sulfate | Sigma-Aldrich | D8906 | |
| Bovine serum albumin (BSA) | New England Biolabs, USA | B9001S | |
| Formamide | Sigma-Aldrich | 47671-1L-F | aliquots kept at -20 °C |
| Illustra TempliPhi Kit Construct (Kit MDA) | Dutsher, France | 25-6400-80 | |
| Nick translation kit | Abbott, USA | 07J00-001 | |
| 20x SSC buffer concentrate | Sigma-Aldrich | S6639 | |
| Spectrum green dUTP | Abbott, USA | 02N32-050 | |
| Spectrum red dUTP | Abbott, USA | 02N34-050 | |
| Cy-5 dUTP | Dutsher, France | PA55022 | |
| Mouse Cot-1 DNA | Invitrogen | 18440016 | |
| DNA, MB grade | Invitrogen | Roche | DNA from fish sperm |
| 4′,6-diamidino-2-phenylindole dihydrochloride | Sigma-Aldrich | D9564 | DAPI |
| Glycerol | Sigma-Aldrich | G9012 | |
| p-phenylenediamine | Sigma-Aldrich | 695106 | |
| Centrifuge 5417R | Eppendorf, Germany | molecular biology grade | |
| Eppendorf concentrator plus | Eppendorf | ||
| Eppendorf Thermomixer comfort | Eppendorf | ||
| Liquiport Liquid pump | KNF Neuberger, Trenton, USA | ||
| Shake'N'Bake Hybridization oven | Boekel Scientific, USA | ||
| Cryostat | Leica | CM3050 |
参考文献
- Cross, J. C. Genetic insights into trophoblast differentiation and placental morphogenesis. Sem. Cell Dev. Biol. 11, 105-113 (2000).
- Zybina, E. V., Zybina, T. G. Polytene chromosomes in mammalian cells. Int. Rev. Cytol. 165, 53-119 (1996).
- Corbel, C., Diabangouaya, P., Gendrel, A. -. V., Chow, J. C., Heard, E. Unusual chromatin status and organization of the inactive X chromosome in murine trophoblast giant cells. Development. 140, 861-887 (2013).
- Rossant, J., Tamura-Lis, W. Effect of culture conditions on diploid to giant-cell transformation in postimplantation mouse trophoblast. J. Embryol. Exp. Morphol. 62, 217-227 (1981).
- El-Hashash, A. H., Kimber, S. J. Trophoblast differentiation in vitro: establishment and characterization of a serum-free culture model for murine secondary trophoblast giant cells. Reproduction. 128, 53-71 (2004).
- Chow, J. C., Heard, E. X inactivation and the complexities of silencing a sex chromosome. Curr Opin Cell Biol. 3, 359-366 (2009).
- Chaumeil, J., Okamoto, I., Heard, E. X-chromosome inactivation in mouse embryonic stem cells: analysis of histone modifications and transcriptional activity using immunofluorescence and FISH. Methods Enzymol. 376, 405-419 (2004).
- Chaumeil, J., Augui, S., Chow, J. C., Heard, E. Combined immunofluorescence, RNA fluorescent in situ hybridization, and DNA fluorescent in situ hybridization to study chromatin changes, transcriptional activity, nuclear organization, and X-chromosome inactivation. Methods Mol. Biol. 463, 297-308 (2008).
- Okamoto, I., Otto, A. P., Allis, C. D., Reinberg, D., Heard, E. Epigenetic dynamics of imprinted XCI during early mouse development. Science. 303, 644-664 (2004).
- Okamoto, I., Arnaud, D., Le Baccon, P., Otte, A. P., Disteche, C. M., Avner, P., Heard, E. Evidence for de novo imprinted X-chromosome inactivation independent of meiotic inactivation in mice. Nature. 438, 369-373 (2005).
- Patrat, C., Okamoto, I., Diabangouaya, P., Vialon, V., Le Baccon, P., Chow, J., Heard, E. Dynamic changes in paternal X-chromosome activity during imprinted X-chromosome inactivation in mice. Proc. Natl. Acad. Sci. 106, 5198-5203 (2009).
- Geijsen Shea, K., N, Dissection of 6.5 dpc mouse embryos. J. Vis. Exp. (2), e160 (2007).
- Pollex, T., Piolot, T., Heard, E. Live-cell imaging combined with immunofluorescence, RNA, or DNA FISH to study the nuclear dynamics and expression of the X-inactivation center. Methods Mol. Biol. 1042, 13-31 (2013).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods. 9, 671-675 (2012).
- Hemberger, M. IFPA award in placentology lecture - characteristics and significance of trophoblast giant cells. Placenta. 29, 4-9 (2008).
- Carney, E. W., Prideaux, V., Lye, S. J., Rossant, J. Progressive expression of trophoblast-specific genes during formation of mouse trophoblast giant cells in vitro. Mol. Reprod. Dev. 34, 357-368 (1993).
- Shin, J., et al. Maternal Rnf12/RLIM is required for imprinted X-chromosome inactivation in mice. Nature. 467, 977-981 (2010).
- Capodieci, P., Donovan, M., Buchinsky, H., Jeffers, Y., Cordon-Cardo, C., Gerald, W., Edelson, J., Shenoy, S. M., Singer, R. H. Gene expression profiling in single cells within tissue. Nat Methods. 9, 663-665 (2005).
- Beliveau, B. J., Joyce, E. F., Apostolopoulos, N., Yilmaz, F., Fonseka, C. Y., McCole, R. B., Chang, Y., Li, J. B., Senaratne, T. N., Williams, B. R., et al. Versatile design and synthesis platform for visualizing genomes with Oligopaint FISH probes. Proc. Natl. Acad. Sci. 109, 21301-21306 (2012).
- Kenny, D., Shen, L., Kolberg, J. A. Detection of viral infection and gene expression in clinical tissue specimens using branched DNA (bDNA) in situ hybridization. J. Histochem. Cytochem. 50, 1219-1227 (2002).
- Cross, J. C., Baczyk, D., Hemberger, M., Hugues, M., Simmons, D. G., Yamamoto, H., Kingdom, J. C. Genes, development and evolution of the placenta. Placenta. 24, 123-130 (2003).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。