
Method Article
在生物安全2级环境中制备假型颗粒研究高致病性冠状病毒
摘要
在这里, 我们提出了一个协议, 以产生伪型粒子在 bsl-2 设置纳入高致病性病毒中东呼吸综合征和严重急性呼吸综合征冠状病毒的尖峰蛋白。这些伪类型粒子包含一个荧光素酶报告基因, 允许病毒进入目标宿主细胞的定量。
摘要
该协议的目的是产生冠状病毒 (cov) 尖峰 (s) 融合蛋白假型粒子与小鼠白血病病毒 (mlv) 核心和荧光素酶记者, 使用一个简单的转染程序广泛可用的 hek-293t 细胞系。一旦从生成细胞中形成和释放, 这些伪病毒体就会整合荧光素酶报告基因。由于它们表面只含有异源冠状病毒尖峰蛋白, 因此在进入步骤中, 这些粒子的行为就像它们的原生冠状病毒对应物。因此, 它们是研究病毒进入宿主细胞的本地病毒的优秀代孕者。在成功进入和感染目标细胞后, 荧光素酶记者被整合到宿主细胞基因组中并表达。使用简单的荧光素酶检测, 被转移的细胞可以很容易地量化。该程序的一个重要优点是, 它可以在生物安全 2级 (bsl-2) 设施中进行, 而不是在处理中东呼吸综合征冠状病毒 (中东呼吸综合征冠状病毒) 等高致病性冠状病毒时所需的 bsl-3 设施。严重急性呼吸综合征冠状病毒 (sars-cov)。另一个好处来自于它的多功能性, 因为它可以应用于属于所有三类病毒融合蛋白的包膜蛋白, 如 i 类流感血凝素 (ha) 和埃博拉病毒糖蛋白 (gp), ii 类 semliki 森林病毒 e1蛋白, 或 iii 类泡状口炎病毒 g 糖蛋白。该方法的一个局限性是, 它只能重述由所研究的包膜蛋白介导的病毒进入步骤。对于研究其他病毒生命周期步骤, 需要其他方法。这些伪型粒子可用于研究宿主细胞易感性和取向, 并测试病毒进入抑制剂对解剖所使用的病毒进入途径的影响的许多应用的例子。
引言
宿主细胞进入是病毒感染生命周期的初始步骤。对于包膜病毒, 这包括结合到单个宿主细胞受体或几个受体, 然后是病毒和细胞膜的融合。这些基本功能是由病毒包膜糖蛋白1,2来实现的。冠状病毒包膜糖蛋白被称为尖峰 (s) 蛋白, 是 i 类病毒融合蛋白2,3,4, 5,6的成员。研究病毒包膜糖蛋白对于了解特定病毒的许多重要特征至关重要, 例如: 生命周期开始、宿主和细胞取向、物种间传播、病毒间传播以及宿主细胞进入途径。病毒伪型粒子, 也被称为伪病毒, 是强大的工具, 使我们能够很容易地研究病毒融合蛋白的功能。假型颗粒或伪病毒是嵌合病毒, 由一个替代病毒核组成, 表面有异源病毒包络蛋白。该协议的主要目的是展示如何获得冠状病毒尖峰伪型粒子, 这些粒子是基于小鼠白血病病毒 (mlv) 的核心, 并包含荧光素酶报告基因。以高致病性严重急性呼吸综合征 (sars) 和中东呼吸综合征 (mers) 冠状病毒为例, 提出了用尖峰蛋白产生假型颗粒的方法。该协议描述了所涉及的转染过程, 如何感染易感靶细胞, 以及通过荧光素酶检测的传染性定量。
由于伪病毒的进入步骤在其表面受冠状病毒 s 的控制, 它们进入细胞的方式与本地对应细胞相似。因此, 他们是功能性传染性检测的优秀代孕者。伪型粒子通常来自亲本模型病毒, 如逆转录酶病毒 (mlv7、8、9、10、11、12、13 ),14,15,16,17,18,19,20,21,22和慢病毒人体免疫机能丧失病毒--艾滋病毒23、24、25、26、27、28、29,30,31,32,33,34,35) 或横纹肌病毒 (泡状口腔炎病毒-vsv 36,39,40,41,42, 43,44,45,46,47). 当用于伪分型时, 父母病毒的基因组被修改以去除基本基因, 使其在完成完整的复制周期方面有缺陷。此功能允许它们用于中间生物安全设施 (bsl-2), 与使用需要更高生物安全设施的高致病性原生病毒 (bsl-3、bsl-4 等不太容易获得) 相比, 这是一个重要的优势。进行病毒进入研究。在这里, 风险3病原体 sars-cov 和 mers-cov 的 s 蛋白被用作病毒包膜蛋白被纳入 mlv 伪型粒子的例子, 产生 sars-cov s 和 mers-cov s 伪病毒 (分别为 sars-spp 和 mers-spp)。这些伪病毒已成功地用于研究这些病毒的进入事件48,49, 50,51.另一个优点是, 这里描述的技术并不局限于伪型冠状病毒 s 蛋白: 它是非常灵活的, 可以用来纳入所有三类病毒融合蛋白的代表。这方面的例子包括流感血凝素 (ha, i 类)52、埃博拉病毒糖蛋白 (gp i 类)、半病毒 semliki 森林病毒 (sfv, ii 类) 的 e1 蛋白和 vsv 糖蛋白 (g, iii 类) 53。此外, 一种以上的病毒糖蛋白可以共同结合成一个伪型粒子, 如在流感 ha-和 na-假型颗粒51的情况下。
根据 bartosch 等人的工作, 该协议描述了 mlv 伪型粒子的生成与三种质粒共同转染策略使用广泛可用和高度可转换的 hek-293t 细胞系54. 第一个质粒编码 mlv 核心基因的口子和pol , 但缺乏 mlv 包络 encodes基因。第二个质粒是一个转移载体, 编码萤火虫荧光素酶报告基因, mlv rna 包装信号, 以及 5 '-和 3 '-侧翼 mlv 长终端重复 (ltr) 区域。第三个质粒编码感兴趣的融合蛋白, 在这种情况下, 无论是 sars-cov s 或中东呼吸综合征冠状岩合酶 s 蛋白。在使用转染试剂共同转染这三个质粒时, 病毒 rna 和蛋白质在转染细胞内表达, 允许产生伪型粒子。由于 mlv 被用作伪病毒主干, 这发生在质膜上: 含有荧光素酶基因报告和包装信号的 rna 被封装到新生的颗粒中, 这些颗粒也包含血浆膜表达的冠状病毒尖峰蛋白质。从细胞中发芽的颗粒表面含有冠状病毒 s 蛋白, 并被收获用于传染性检测。由于伪型粒子含有冠状病毒 s 蛋白, 而不是 mlv 包膜蛋白, 当用于感染细胞时, 它们的行为就像它们的本地冠状病毒对应的进入步骤。然后, 含有荧光素酶报告和侧翼 ltr 的病毒 rna 在细胞内释放, 而逆转录病毒聚合酶活性使其逆转录酶转化为 dna 并整合到宿主细胞基因组中。然后, 用简单的荧光素酶活性测定方法对病毒伪型颗粒在受感染细胞中的传染性进行定量。由于整合到宿主细胞基因组中的 dna 序列只包含荧光素酶基因, 而没有任何 mlv 或冠状病毒蛋白编码基因, 因此它们的使用本质上比具有复制能力的原生病毒更安全。
除了是更安全的代孕药物和高度适应性, 允许纳入各种包膜糖蛋白, 这里描述的伪类型粒子也是高度通用的, 可以用来研究病毒进入的许多方面。这包括但不限于: 检测宿主细胞对病毒感染的易感性, 分析包住病毒使用的细胞进入途径, 研究药理抑制剂和药物筛选的效果, 进行中和抗体检测, 描述宿主细胞进入的包膜病毒, 不能培养, 并产生病毒载体的基因传递, 稳定的细胞表达的兴趣基因, 或基因沉默。
研究方案
1. 用于假型颗粒生产的细胞播种
请注意:在生物安全柜中执行此步骤。
- 通过标准的细胞培养技术, 获得一个80–90% 融合75厘米2气瓶的 hek-293t17 细胞通过完整的 dulbecco 的修饰鹰的培养基 (dmem-c), 其中含有 10% (volvol) 胎牛血清 (fbs), 10 mm 4-(2-羟乙基)-1-哌嗪磺酸 (hepes)、100 uu/ml 青霉素和100μgml 链霉素。准备 DMEM-T 介质进行转染 (其成分与 DMEM-T 相同, 但不含抗生素)。
- 用10毫升的预加热 (37°c) 杜尔贝科的磷酸盐缓冲盐水 (dpbs) 清洗细胞两次。
请注意:小心地处理 hek29t17 细胞, 因为它们很容易分离。 - 在37°c 预热时, 用1毫升0.25% 的胰蛋白酶溶液吸收上清液和分离细胞。将细胞的烧瓶放在 37°c, 5% co 2 孵化器 3-5分钟或直到细胞开始分离。
请注意:避免用胰蛋白酶孵育细胞超过 5分钟, 因为这通常会导致细胞聚集。 - 通过添加4毫升的 dmem-c 培养基和计数细胞来停用胰蛋白酶, 并使用细胞计数幻灯片和光学显微镜。
请注意:为了避免需要计数过多的细胞, 可能需要事先采取额外的稀释步骤。在计算胰蛋白酶化细胞的实际细胞密度时, 请记住考虑到这种稀释。 - 用 dmem-c 将细胞稀释至 5x 10 5细胞。
- 种子6孔组织培养板与2毫升的细胞溶液每口井, 并轻轻地来回和侧向移动板均匀分布细胞, 避免圆周运动。
请注意:这是关键的一步。均匀分布的细胞将确保细胞不会聚集在井的中心。反过来, 这将确保良好的转染效率和伪型颗粒生产。 - 在37°c、5% co2细胞培养孵化器中孵育板 (16–18小时)。
2. 三粒质粒共转染
请注意:在生物安全柜中执行此步骤。
- 在倒置光学显微镜下观察细胞, 以检查细胞的形态和密度。
请注意:理想情况下, 细胞密度应在40–60% 的融合范围内。至关重要的是, 细胞既不是太汇出 (80-90% 融合), 也不是太稀疏分布 (20-30% 融合) 在每口井。细胞密度为40-60% 的融合将确保良好的伪型颗粒的产生。 -
质粒混合
- 根据表 1所示6井板的一口井的数量, 计算每个包络糖蛋白的质粒混合物。如果转染几口井, 并包括一口额外的井, 以避免混合耗尽, 则会增加数量。
请注意:除了 sars-cov s 和 mers-cov s 编码质粒外, 还包括空矢量控制, 用于产生缺乏病毒包膜糖蛋白 (env 粒子) 的负控制粒子, 以及一个正对照糖蛋白, 如囊泡口腔炎病毒 (vsv) g 糖蛋白, 已知能有力地感染非常广泛的细胞 (vsv-gpp)。可应要求提供质粒。 - 将计算出的质粒体积和减少的血清细胞培养培养基 (见材料表) 混合在微离心管中。
- 根据表 1所示6井板的一口井的数量, 计算每个包络糖蛋白的质粒混合物。如果转染几口井, 并包括一口额外的井, 以避免混合耗尽, 则会增加数量。
-
基于脂质的转染试剂混合物(见材料表)
- 根据表 2所示的一口井的数量计算转染试剂混合物的体积 (1:3 转染比, 根据需要乘以数量)。包括额外的井, 以避免耗尽转染试剂的混合物。
- 将计算出的脂基转染试剂 (每口3毫升) 和减少的血清细胞培养培养基 (每井 47μl) 混合在微离心管中, 确保将转染试剂添加到减少的血清细胞培养培养基中, 而不是以其他方式添加转染试剂周围。
- 在室温下分别将两种混合物 (一口井: 50μl 的质粒混合物和50μl 的脂质转染试剂混合物) 培养5分钟。
- 将转染试剂混合物的内容添加到质粒中, 比例为 1:1 (每种混合物为1口井: 50μl)
- 对产生的混合物进行彻底的上下移液。
- 在室温下将混合物孵化至少20分钟。
- 吸收细胞的乏培养基。
- 轻轻加入1毫升预热 (37°c) 减少血清细胞培养培养基。
- 在每口井中加入滴度100μl 转染混合物。
请注意:当 hek-293t 井容易分离时, 在将转染混合物添加到井时要小心。 - 将细胞孵育在 37°c, 5% co2 细胞培养孵化器中, 孵育4-6小时。
- 在不含抗生素的预热 (37°c) dmm-t 培养基中, 每口井添加1毫升
请注意:这是关键的一步。转染试剂可提高细胞通透性, 提高对抗生素的敏感性。为保证良好的转染效率和伪型颗粒的产生, 避免使用含有抗生素的细胞培养基是非常重要的 - 在 37°c, 5% co2 细胞培养孵化器中孵育细胞48小时。
3. 伪分型颗粒集合
请注意:在生物安全柜中执行此步骤。
- 观察倒置光镜下的细胞形态和一般情况。还要检查介质的颜色, 这些介质应该是浅的粉红/略带橙色。
请注意:这是一个重要的步骤。如果有太多的细胞死亡与转染或中等颜色变成橙色黄色 (酸性 ph), 这通常会与较低的传染性假型颗粒的产量 - 将转染细胞的上清液转移到50毫升锥形离心管中。
- 以 290 x g 离心管 7分钟 , 清除细胞碎片。
- 滤清器通过0.45μm 孔径的无菌过滤器澄清上清液。
- 在低温中制造小体积的伪型病毒溶液 (例如, 500μl 或 1 ml)。
- 存放在-80°c。
请注意:协议可以在这里暂停。假型颗粒在-80°c 下稳定数月, 但解冻后, 避免重新冻结, 因为它们会失去传染性
4. 易感细胞的假型颗粒感染
请注意:在生物安全柜中执行此步骤。
-
24孔板易感细胞的细胞播种
- 通过标准细胞培养技术获得80-90% 的75厘米2易感细胞的融合: sars-cov 伪型粒子 (sars-spp) 的 vero-e6 细胞和中东呼吸综合征 cov s 伪型粒子 (mers-spp) 的 huh-7 细胞。
请注意:为了确认所产生的伪型粒子是否具有传染性, 重要的是要仔细选择合适的易感细胞系进行伪病毒感染检测。使用许可性差的细胞会导致低传染性。 - 用预热 (37°c) dpbs 的10毫升清洗细胞两次。
- 在37°c 预热时, 用1毫升0.25% 的胰蛋白酶溶液吸收上清液和分离细胞。将细胞的烧瓶放在 37°c, 5% co 2 孵化器 3-5分钟或直到细胞开始分离。
请注意:避免用胰蛋白酶孵育细胞超过 5分钟, 因为这通常会导致细胞聚集。huh-7 细胞对这种效应特别敏感。 - 通过添加 dmem-c 培养基和使用细胞计数幻灯片和光学显微镜计数细胞来停用胰蛋白酶。
- 用 dmem-c 将细胞稀释至 5x 10 5细胞。
- 24孔板的种子井, 每口井有0.5 毫升的细胞溶液, 并轻轻来回和侧向移动板均匀分布细胞, 避免圆周运动
请注意:这是关键的一步。均匀分布的细胞将确保细胞不会聚集在井的中心。反过来, 这将确保良好的传染性检测。对于每个伪类型粒子 (sars-spp, mers-spp) 和条件, 为三个实验复制准备三口井。包括未受感染的井 (n. i.)、空向量 env 粒子和正对照粒子, 如 vsv-gpp - 在37°c、5% co2细胞培养孵化器中孵育板 (16-18小时)
- 通过标准细胞培养技术获得80-90% 的75厘米2易感细胞的融合: sars-cov 伪型粒子 (sars-spp) 的 vero-e6 细胞和中东呼吸综合征 cov s 伪型粒子 (mers-spp) 的 huh-7 细胞。
-
伪型粒子感染
- 在光学显微镜下观察细胞, 并在视觉上证实有一个混合地毯的细胞。
- 将伪型病毒的冷冻剂带到冰上解冻。
- 用 0.5 ml 预热 (37°c) dpbs 清洗细胞三次
请注意:这是关键的一步。在感染前没有适当冲洗的细胞通常会导致不良的传染性读数 - 吸收细胞的上清液。
- 具有200μl 解冻的伪型颗粒溶液的接种细胞。
- 在37°c、5% co2细胞培养孵化器中孵育细胞1-2小时。
- 加入300μl 的预热 (37°c) dmem-c 介质。
- 在37°c、5% co2细胞培养孵化器中孵育细胞72小时。
5. 通过荧光素酶检测读数进行的传染性定量
请注意:在生物安全柜中执行初始步骤。
- 解冻荧光素底物 (储存在-80°c) 和5倍荧光素酶裂解缓冲液 (储存在-20°c), 直到达到室温。
- 用无菌水稀释荧光素酶分析裂解缓冲液1x。
- 吸收假型颗粒感染的细胞的吸血上清液。
- 在每口井中加入100μl 的1x 荧光素酶分析裂解缓冲液。
- 将板材放在摇杆上, 在室温下摇椅孵育 15分钟 (从此时开始, 板材可在生物安全柜外处理)。
- 在每口井中加入20μl 的荧光素基板, 为每口井准备微离心管。
- 打开计时器。
- 通过将10μl 的裂解液转移到一个含有20μl 的荧光素基质的管中, 一次很好地测量一个荧光素酶活性。
- 轻轻滑动管, 混合内装物, 但避免更换管壁上的液体。
- 将管子放入设备并合上盖子。
- 使用电光计测量管的发光值。
- 记录相对光单位的测量值。
- 重复步骤 5.8–5.12, 直到对所有油井进行分析。
请注意:有了相应的设备, 如制版读数计, 这个过程可以自动执行。检测将需要缩放到板格式 (例如, 96 孔板格式)。
6. 数据分析
-
相对荧光素酶单位平均值和标准偏差的计算与绘制
- 使用图形绘图软件计算荧光素酶测定平均值和标准偏差的实验和生物复制。
- 将数据绘制为带有标准偏差的条形图。
请注意:在对数据进行统计分析时, 请确保在数据集中至少包括三个生物复制。
结果
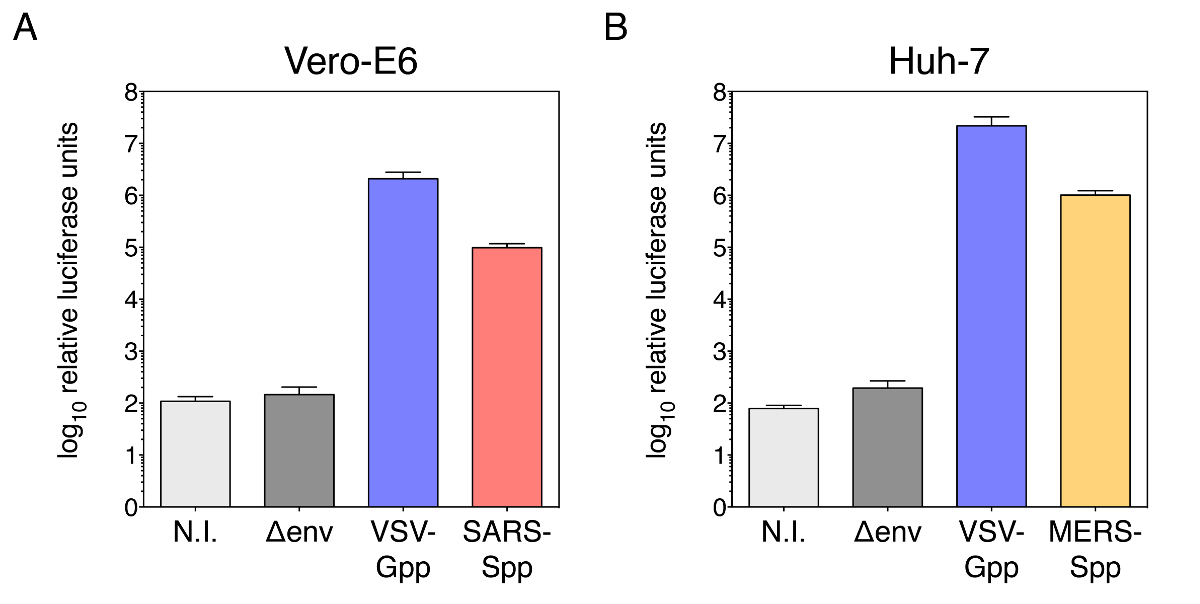
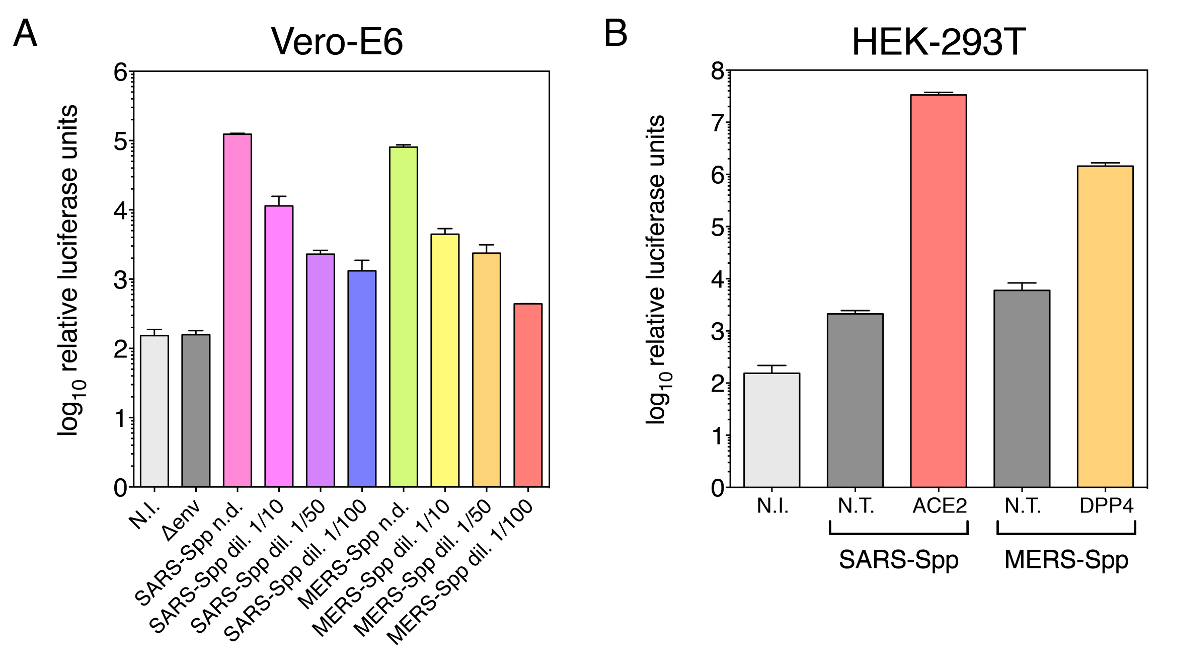
图 1显示了 sars-cov s 和中东呼吸综合征冠状粒子的传染性检测结果。如图 1a 和 1b, vsv g 伪分阳性对照粒子 (vsv-gpp) 分别在 10 6 至 107个相对荧光素酶 (rlu) 范围内给出了非常高的平均传染性。对于传染性 vero-e6 细胞的 sars-cov s 伪型颗粒 (图 1a) 感染, 在 9.8 x 10 4 rlu 左右测量出强平均传染性 。这个值几乎比未受感染的对照 (1.1 x 10 2 rlu) 或 env 粒子 (1.5 x10 2 rlu) 的测量值高出近3个数量级, 这些颗粒表面没有任何病毒包膜糖蛋白。同样, 对于 huh-7 细胞的中东呼吸综合征冠状 s 假型颗粒 (图 1b) 感染, 在 1.0 x10 6 rlu左右测量出了较高的平均传染性。这几乎比未受感染的控制 (0.8 x10 2 rlu) 或 env 粒子 (2.0 x10 2 rlu) 的测量值高出4个数量级。另外进行了一次传染性检测, 对 sars-spp 和 mers-spp 进行了连续稀释, 用于感染 ver-e6 细胞 (图 2a)。该分析证实, 所测量的荧光素酶活性取决于用于感染细胞的颗粒浓度。为了证实血管紧张素转换酶 2 (ace2) 和二肽酶 4 (dpp4) 在中备型粒子的介导和输入中的作用, 我们使用了较差的宽松 hak-293 细胞和转染它们以表示 ace2 或 dpp4 (图 2b)。然后将转染的细胞用于感染率检测。该分析表明, 在 ace2 和 dpp4 过度表达后, sars-spp 和 mers-spp 的感染分别有 ~ 4-log 和 ~ 2-log 的增加, 证实假病毒受体的使用与原生病毒相似。
此处显示的示例证明了在产生伪类型粒子时包括负 (未受感染、env 颗粒) 以及正控制 (vsv-gpp) 条件的重要性。事实上, 阳性对照 vsv-gpp 粒子使我们能够评估特定批次的伪型粒子是否成功地产生了功能性和传染性的伪病毒。大多数哺乳动物细胞系中 vsv-gpp 颗粒典型感染的预期结果在 106-107rlu 范围内.hek-29jt17 生成单元的问题 (高通道数、细胞密度问题) 或转染效率低可能会影响整体伪型粒子的生产和感染率。此外, 负控制条件也很重要, 因为它们使我们能够评估特定细胞系 (非感染条件) 中 rlu 测量的基线和不导这种情况的粒子非特定内化 (env 感染)由病毒包膜蛋白。理想情况下, 对于具有感兴趣的病毒包络蛋白的给定类型的粒子伪类型, 建议获取比负值更高的几个数量级的值, 如图 1a、b 所示。然而, 如果对于给定的细胞类型, 伪型病毒感染提供的传染性很小 (即接近非转染的 n. t. 条件下的负对照), 则不一定意味着伪类型病毒感染颗粒生产出现故障。很可能是所使用的特定细胞系对感染不允许或不允许。建议检查特定的细胞类型是否容易被正在调查的病毒感染。用表达病毒受体的质粒转染不允许的细胞, 可以让更有效的病毒进入和感染发生, 如图 2b所示, 在目标 hek-293t 中转染 ace2 和 dpp4 受体时细胞, 有一个 ~ 4-和 ~ 2-日志的传染性, 分别增加。
| 塑料/试剂 | 数量 |
| pcmv-mlvgagpol mlv 口子和 pol 编码质粒 | 300纳克 |
| ptg-luc 转移载体与荧光素酶记者 | 400纳克 |
| pcdna-sars-s, pcDNA-SARS-S 或空向量 | 300纳克 |
| 减少血清细胞培养培养基 | 至50μl |
表 1: 为伪分型颗粒生产而转染 hk-29红豆17细胞6井板的一口井所需的质粒数量和减少的血清细胞培养介质.从质粒制剂的浓度计算出所需的体积, 以达到所需的质粒 dna 量。如果用相同的质粒转染了一个以上的井, 则将体积乘以所需数量的井进行转染, 并在计算中包括一个额外的井, 以避免在以后的步骤中耗尽混合。被转染的 dna 总量为 1μg well。等质粒可应要求提供给作者。
| 试剂 | 数量 |
| 转染试剂 | 3μl |
| 减少血清细胞培养培养基 | 47μl |
表 2: 为伪型颗粒生产而转染 hek-29红豆17细胞6井板的一口井所需的转染试剂和减少的血清细胞培养介质的数量.将体积乘以所需数量的井进行转染, 并在计算中包括额外的井, 以避免在后面的步骤中耗尽混合。转染试剂: 使用质粒 dna 比为3:1。

图 1: 利用小鼠白血病病毒 (mlv) 主干和荧光素酶报告基因检测易感宿主细胞中的冠状病毒 s-伪型颗粒感染率.(a) 在 vero-e6 细胞中进行表伪型粒子感染率试验。(b) 中东呼吸综合征冠状 s 伪型粒子在 huh-7 细胞中的感染率测定。对于 (a) 和 (b), 绘制的数据对应于三个独立实验的平均相对荧光素酶单位, 误差条对应于标准偏差 (s. d.)。在 y 轴上的日志10刻度中绘制的数据。非感染对照组;env 感染缺乏病毒包膜糖蛋白和 vsv-gpp 的假型颗粒: 含有阳性对照 vsv g 包膜蛋白的假型颗粒感染。其他使用的缩写, sars-spp: 感染 sars-cov s 假型粒子, mers-spp: 感染中东呼吸综合征冠状 s 表。请点击这里查看此图的较大版本.
{kind=link}

图 2: cov s-伪病毒的浓度依赖性以及 ace2 和 dpp4 受体在 sars-spp 和 mers-spp 进入中的作用.(a) 荧光素酶活性测定 sars-spp 和 mers-spp 的浓度依赖性。假病毒被连续稀释, 用于感染 vero-e6 细胞。(b) 分别在转化为表达 ace2 的 hek-293t 靶细胞和 dpp4 受体中进行 sars-spp 和 mers-spp 的感染检测。数据在日志10中绘制的数据在 y 轴上绘制, 如图1中的重复实验中绘制的数据, 误差条与标准偏差 (s. d.) 相对应。n. t.: 非转染对照 hek-293t 细胞;ace2: 血管紧张素转换酶 2 (sars-cov 受体) 和 dpp4:dipertidil 肽酶 4 (mers-cov 受体)。使用的其他缩写与图 1中的缩写相同。请点击这里查看此图的较大版本.
{kind=link}
讨论
该协议描述了一种在 bsl-2 设置中有效生成含有风险3冠状病毒 s 蛋白 (sars-cov 和 mers-cov) 的假型粒子的方法。这些颗粒, 其中包含荧光素酶报告基因, 使我们能够很容易地量化冠状病毒 s 介导的进入事件, 通过相对简单的荧光素酶检测48,49,50,51。在使用允许细胞的感染性检测中, 我们证实所测量的荧光素酶活性取决于颗粒的浓度。此外, ace2 和 dpp4 受体转染允许更有效地进入许可差的细胞系, 如 hek-293t 细胞。该方法对其他病毒包膜糖蛋白有很强的适应性, 已被广泛使用48,49,50,51, 52,53, 55、56、57、58、59, 通常是为了补充其他检测, 如生化分析或本地病毒感染。
我们在这里描述的协议是基于逆转录酶病毒 mlv, 其中包含了一个荧光素酶记者。但是, 必须强调的是, 已经成功地开发了一系列非常广泛的其他伪类型系统来包装冠状病毒 s12、13、25、26、30,31,32和其他病毒包膜糖蛋白10,11,14,16, 17, 23,24,29,33,38,40,42,44,46. 其中一些其他系统以常用的 mlv 抗逆转录病毒核心7、8、9、10、11、12、13为基础 ,14,15,16,17, 18,19, 20,或基于广泛使用的慢病毒 hiv-1使用不同策略的伪分型系统23、24、25、26、27、28、29、30 ,31,32,33,34,35, 或以横纹布病毒泡状口炎病毒 (vsv) 为核心, 这允许纳入广泛的品种包膜糖蛋白, 并再次采用各种策略使用 37,38,39, 40,41,42, 43 44, 45,46,47.此外, 其他记者如荧光蛋白如 gfp11、13和 rfp36, 或其他酶, 如β-半乳糖苷酶16,17和分泌碱性磷酸酶 (seap)42已成功地用于测量。此外, 在本协议中提出的分析中, 采用瞬态转染来表达 mlv 和 cov s 基因和蛋白质。然而, 还有其他表达策略, 例如生成用于生产伪类型病毒的稳定细胞系7,14。由于每个系统都有其优点和缺点, 在决定哪种伪类型系统最适合调查员的需要时, 必须考虑以下重要参数: 伪 virion 核心 (mlv、hiv-1、vsv 或其他), 如何选择性的伪分型核心是在纳入一个特定的病毒包络糖蛋白, 检测读数的报告 (transfection, 荧光素酶, seap 或其他), 和转染策略 (参与联合转染的质粒的数量,转染或生成稳定的细胞系)。
该方法中有许多重要步骤需要强调。细胞密度, 特别是 hek-293t17 生产者细胞系的密度是确保成功转染的关键因素。在40-60% 的融合范围内的细胞密度被认为是最佳的。密度越高, 转染效率越低, 颗粒产量也越低。另外, 重要的是要记住, hek-293t17 细胞的粘附程度低于其他细胞系。在处理这些问题时, 应谨慎行事, 以避免不必要地将它们分离。一种选择是用多 d-赖氨酸处理细胞培养塑料表面, 以增强粘附。此外, 较高的细胞通道往往导致转染率很低。在 hek-293t17 细胞中添加转染试剂后, 还必须记住细胞的渗透率增加。这就是为什么在这一点上, 最好避免使用含有抗生素的介质, 因为它们可能会增加细胞毒性。在收集伪型粒子之前, 检查转染 hek-29nt17 细胞上清液的颜色。通常情况下, 经过48小时的转染后, 细胞培养介质的颜色采取橙色-粉红色的色彩。黄色介质通常转化为伪型颗粒产量低, 通常是细胞播种密度或高通道数问题的结果。
在该协议中, 伪类型粒子生产是以6孔板格式进行的。为了增加产生的颗粒量, 可以用相同的质粒混合转染6井板的几口井, 并将上清液汇集在一起。然后水池可以被澄清, 过滤和别名。或者, 为了扩大生产规模, 可以使用其他种类的容器 (例如25或75厘米2瓶)。在这种情况下, 应相应地扩大转染条件。在该协议中, 感染率检测使用24孔板格式和计步仪进行, 一次只能测量一个管。对于高吞吐量的筛选, 其他格式也是可能的, 如96孔板格式和板式读卡器的发光计。荧光素酶测定的卷和试剂需要相应地进行调整。在-80°c 条件下将假型颗粒储存在低温中, 可在几个月内保持其稳定性, 而不会显著降低传染性。不建议对它们进行冻融循环, 因为随着时间的推移, 这将减少它们的传染性。因此, 最好将它们存放在小的等价物中, 如 0.5–1 ml, 并在感染前解冻。
这里介绍的方法有几个限制。一个重要的事实是, 伪类型粒子只重述病毒进入事件。要分析传染性生命周期中的其他步骤, 还需要进行其他检测。此外, 当 mlv 颗粒在质膜上发芽时, 重要的是要记住, 正在研究的包膜糖蛋白还需要流量到质膜上, 以便在生产过程中融入伪病毒。因此, 重要的是要知道在细胞中, 特定的病毒包络糖蛋白在转染条件下的表达, 例如通过免疫荧光法的可视化亚细胞定位, 或者通过检查内的保留信号蛋白质。此外, 虽然该协议描述了生产和测试传染性的步骤, 但它没有详细说明如何测量病毒包膜糖蛋白与伪类型粒子的结合。一种方法是对颗粒的浓缩溶液进行西方印迹检测, 如前所述, 对中东呼吸综合征冠状病毒 s 的 50,51进行检测。在这些检测中, 将中东呼吸综合征冠状病毒的 s 包络糖蛋白与 mlv 的衣壳 (p30) 蛋白一起进行研究, 使我们能够将 s 蛋白正常化地纳入颗粒。分析病毒包膜糖蛋白纳入伪病毒的其他例子是在 hiv-1 病毒假病毒系统32中进行 sars-cov s 的整合, 埃博拉糖蛋白 (gp) 在另一种 mlv 伪型中的应用颗粒系统17, 流感血凝素 (ha) 和神经氨酸酶 (na) 在 vsv 伪病毒 38.在描述伪型粒子生产方面最近的一个发展是使用了创新的成像设备, 如纳米视觉: 它使我们能够直接对50的病毒粒子进行可视化、量化和大小。该装置提供了关于整体颗粒生产的详细信息;然而, 重要的是要记住, 它不提供有关包络糖蛋白结合的信息。这些多功能伪病毒粒子的应用前景是使用单粒子跟踪、微流体和总内部反射荧光显微镜60,61 分析单个病毒融合事件 ,62。该方法成功地应用于流感病毒和猫科动物冠状病毒颗粒以及甲型和 na 伪型的基于 vsv 的伪病毒63。目前正在开发将此类技术应用于基于 mlv 的核黄病毒 s 型假型粒子的应用。
披露声明
作者没有什么可透露的。
致谢
我们要感谢惠特克和丹尼尔实验室的所有成员提出的有益意见。国家卫生研究院提供了研究资金, 其中包括 r21 ai111085 和 r01 ai135270。t. t. 感谢康奈尔大学总统生命科学研究金和国家科学基金会研究生研究奖学金计划对第1号赠款的支持。dge-165041。法律公告感谢塞缪尔·弗莱明家庭研究生奖学金和国家科学基金会研究生研究金计划的支持。dge-165041。这项工作也得到了国家科学基金会1504846的支持 (对 s. d. 和 g. r. w.)。
材料
| Name | Company | Catalog Number | Comments |
| Human embryonic kidney (HEK) HEK-293T/17 cells | ATCC | CRL-11268 | Clone 17 cells are highly competent for transfection. |
| African green monkey kidney epithelial Vero-E6 cells | ATCC | CRL-1586 | |
| Human hepatic Huh-7 cells | Japan National Institutes of Biomedical Innovation, Health and Nutrition | JCRB0403 | |
| Inverted light microscope with 10 × objective | Nikon | TS100 | |
| Dulbecco’s modification of Eagle's medium (DMEM) with 4.5 g/L glucose, L-glutamine without sodium pyruvate | Corning Mediatech | 10-017-CV | |
| Heat-inactivated fetal bovine serum (FBS) | Thermo Fisher Scientific, Gibco | 1614071 | |
| 1 M N-2-hydroxyethylpiperazine-N'-2-ethanesulphonic acid (HEPES) | Corning Mediatech | 25-060-Cl | |
| 100 × penicillin-streptomycin (PS) solution | Corning Mediatech | 30-002-Cl | |
| Dulbecco’s phosphate buffered saline (DPBS) with Ca2+ and Mg2+ | Corning Mediatech | 21-030-CV | |
| 0.25% trypsin, 2.21 mM ethylenediaminetetraacetic acid (EDTA) 1 × solution | Corning Mediatech | 25-053-Cl | |
| Cell counting slides with grids | Kova | 87144 | |
| Opti-minimal essential medium (Opti-MEM) | Thermo Fisher Scientific, Gibco | 31985-070 | |
| Lipofectamine 2000 transfection reagent | Thermo Fisher Scientific, Invitrogen | 11668-027 | |
| 0.45 µm pore-size sterile filter | Pall | 4184 | |
| 10 mL syringes | BD | 309604 | |
| 5 × luciferase assay lysis buffer | Promega | E1531 | |
| Luciferin, substrate for luciferase assay | Promega | E1501 | |
| Sterile water | VWR | E476-1L | |
| GloMax 20/20 luminometer | Promega | 2030-100 |
参考文献
- Dimitrov, D. S. Virus entry: molecular mechanisms and biomedical applications. Nature Reviews Microbiology. 2 (2), 109-122 (2004).
- White, J. M., Delos, S. E., Brecher, M., Schornberg, K. Structures and Mechanisms of Viral Membrane Fusion Proteins: Multiple Variations on a Common Theme. Critical Reviews in Biochemistry and Molecular Biology. 43 (3), 189-219 (2008).
- Bosch, B. J., van der Zee, R., de Haan, C. A. M., Rottier, P. J. M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. Journal of Virology. 77 (16), 8801-8811 (2003).
- Belouzard, S., Millet, J. K., Licitra, B. N., Whittaker, G. R. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein. Viruses. 4 (6), 1011-1033 (2012).
- Millet, J. K., Whittaker, G. R. Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology. 517, 3-8 (2018).
- Millet, J. K., Whittaker, G. R. Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis. Virus Research. 202, 120-134 (2015).
- Steidl, S., et al. Coreceptor Switch of [MLV(SIVagm)] Pseudotype Vectors by V3-Loop Exchange. Virology. 300 (2), 205-216 (2002).
- Höhne, M., Thaler, S., Dudda, J. C., Groner, B., Schnierle, B. S. Truncation of the Human Immunodeficiency Virus-Type-2 Envelope Glycoprotein Allows Efficient Pseudotyping of Murine Leukemia Virus Retroviral Vector Particles. Virology. 261 (1), 70-78 (1999).
- Wang, W., et al. Establishment of retroviral pseudotypes with influenza hemagglutinins from H1, H3, and H5 subtypes for sensitive and specific detection of neutralizing antibodies. Journal of Virological Methods. 153 (2), 111-119 (2008).
- Wallerstrom, S., et al. Detection of antibodies against H5 and H7 strains in birds: evaluation of influenza pseudovirus particle neutralization tests. Infection Ecology, Epidemiology. 4, (2014).
- Radoshitzky, S. R., et al. Transferrin receptor 1 is a cellular receptor for New World haemorrhagic fever arenaviruses. Nature. 446 (7131), 92-96 (2007).
- Han, D. P., Kim, H. G., Kim, Y. B., Poon, L. L., Cho, M. W. Development of a safe neutralization assay for SARS-CoV and characterization of S-glycoprotein. Virology. 326 (1), 140-149 (2004).
- Moore, M. J., et al. Retroviruses pseudotyped with the severe acute respiratory syndrome coronavirus spike protein efficiently infect cells expressing angiotensin-converting enzyme 2. Journal of Virology. 78 (19), 10628-10635 (2004).
- Sharkey, C. M., North, C. L., Kuhn, R. J., Sanders, D. A. Ross River virus glycoprotein-pseudotyped retroviruses and stable cell lines for their production. Journal of Virology. 75 (6), 2653-2659 (2001).
- Bruett, L., Clements, J. E. Functional murine leukemia virus vectors pseudotyped with the visna virus envelope show expanded visna virus cell tropism. Journal of Virology. 75 (23), 11464-11473 (2001).
- Ma, M., et al. Murine leukemia virus pseudotypes of La Crosse and Hantaan Bunyaviruses: a system for analysis of cell tropism. Virus Research. 64 (1), 23-32 (1999).
- Wool-Lewis, R. J., Bates, P. Characterization of Ebola virus entry by using pseudotyped viruses: identification of receptor-deficient cell lines. Journal of Virology. 72 (4), 3155-3160 (1998).
- Op De Beeck, A., et al. Characterization of Functional Hepatitis C Virus Envelope Glycoproteins. Journal of Virology. 78 (6), 2994-3002 (2004).
- Calland, N., et al. (-)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology. 55 (3), 720-729 (2012).
- Bartosch, B., Dubuisson, J., Cosset, F. L. Infectious hepatitis C virus pseudo-particles containing functional E1-E2 envelope protein complexes. Journal of Experimental Medicine. 197 (5), 633-642 (2003).
- Giroglou, T., et al. Retroviral vectors pseudotyped with severe acute respiratory syndrome coronavirus S protein. Journal of Virology. 78 (17), 9007-9015 (2004).
- Dye, C., Temperton, N., Siddell, S. G. Type I feline coronavirus spike glycoprotein fails to recognize aminopeptidase N as a functional receptor on feline cell lines. Journal of General Virology. 88 (6), 1753-1760 (2007).
- Kobinger, G. P., Weiner, D. J., Yu, Q. -. C., Wilson, J. M. Filovirus-pseudotyped lentiviral vector can efficiently and stably transduce airway epithelia in vivo. Nature Biotechnology. 19, 225 (2001).
- Salvador, B., Zhou, Y., Michault, A., Muench, M. O., Simmons, G. Characterization of Chikungunya pseudotyped viruses: Identification of refractory cell lines and demonstration of cellular tropism differences mediated by mutations in E1 glycoprotein. Virology. 393 (1), 33-41 (2009).
- Nie, Y., et al. Highly infectious SARS-CoV pseudotyped virus reveals the cell tropism and its correlation with receptor expression. Biochemical and Biophysical Research Communications. 321 (4), 994-1000 (2004).
- Grehan, K., Ferrara, F., Temperton, N. An optimised method for the production of MERS-CoV spike expressing viral pseudotypes. MethodsX. 2, 379-384 (2015).
- Bakri, Y., et al. The Maturation of Dendritic Cells Results in Postintegration Inhibition of HIV-1 Replication. The Journal of Immunology. 166 (6), 3780-3788 (2001).
- Hsu, M., et al. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proceedings of the National Academy of Sciences. 100 (12), 7271-7276 (2003).
- Simmons, G., et al. DC-SIGN and DC-SIGNR Bind Ebola Glycoproteins and Enhance Infection of Macrophages and Endothelial Cells. Virology. 305 (1), 115-123 (2003).
- Simmons, G., et al. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proceedings of the National Academy of Sciences. 102 (33), 11876-11881 (2005).
- Gierer, S., et al. The spike-protein of the emerging betacoronavirus EMC uses a novel coronavirus receptor for entry, can be activated by TMPRSS2 and is targeted by neutralizing antibodies. Journal of Virology. 87 (10), 5502-5511 (2013).
- Bertram, S., et al. Cleavage and activation of the SARS-coronavirus spike-protein by human airway trypsin-like protease. Journal of Virology. 85 (24), 13363-13372 (2011).
- Bertram, S., et al. TMPRSS2 and TMPRSS4 facilitate trypsin-independent spread of influenza virus in Caco-2 cells. Journal of Virology. 84 (19), 10016-10025 (2010).
- Connor, R. I., Chen, B. K., Choe, S., Landau, N. R. Vpr Is Required for Efficient Replication of Human Immunodeficiency Virus Type-1 in Mononuclear Phagocytes. Virology. 206 (2), 935-944 (1995).
- Evans, M. J., et al. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature. 446 (7137), 801-805 (2007).
- Negrete, O. A., et al. EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature. 436 (7049), 401-405 (2005).
- Suda, Y., et al. Analysis of the entry mechanism of Crimean-Congo hemorrhagic fever virus, using a vesicular stomatitis virus pseudotyping system. Archives of Virology. 161 (6), 1447-1454 (2016).
- Zimmer, G., Locher, S., Berger Rentsch, M., Halbherr, S. J. Pseudotyping of vesicular stomatitis virus with the envelope glycoproteins of highly pathogenic avian influenza viruses. Journal of General Virology. 95, 1634-1639 (2014).
- Moeschler, S., Locher, S., Conzelmann, K. K., Kramer, B., Zimmer, G. Quantification of Lyssavirus-Neutralizing Antibodies Using Vesicular Stomatitis Virus Pseudotype Particles. Viruses. 8 (9), (2016).
- Tong, W., Yin, X. X., Lee, B. J., Li, Y. G. Preparation of vesicular stomatitis virus pseudotype with Chikungunya virus envelope protein. Acta Virologica. 59 (02), 189-193 (2015).
- Tani, H., et al. Involvement of ceramide in the propagation of Japanese encephalitis virus. Journal of Virology. 84 (6), 2798-2807 (2010).
- Kaku, Y., et al. Second generation of pseudotype-based serum neutralization assay for Nipah virus antibodies: sensitive and high-throughput analysis utilizing secreted alkaline phosphatase. Journal of Virological Methods. 179 (1), 226-232 (2012).
- Tani, H., et al. Analysis of Lujo virus cell entry using pseudotype vesicular stomatitis virus. Journal of Virology. 88 (13), 7317-7330 (2014).
- Ogino, M., et al. Use of Vesicular Stomatitis Virus Pseudotypes Bearing Hantaan or Seoul Virus Envelope Proteins in a Rapid and Safe Neutralization Test. Clinical and Vaccine Immunology. 10 (1), 154-160 (2003).
- Logan, N., et al. Efficient generation of vesicular stomatitis virus (VSV)-pseudotypes bearing morbilliviral glycoproteins and their use in quantifying virus neutralising antibodies. Vaccine. 34 (6), 814-822 (2016).
- Takada, A., et al. A system for functional analysis of Ebola virus glycoprotein. Proceedings of the National Academy of Sciences. 94 (26), 14764-14769 (1997).
- Whitt, M. A. Generation of VSV pseudotypes using recombinant DeltaG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. Journal of Virological Methods. 169 (2), 365-374 (2010).
- Lai, A. L., Millet, J. K., Daniel, S., Freed, J. H., Whittaker, G. R. The SARS-CoV Fusion Peptide Forms an Extended Bipartite Fusion Platform that Perturbs Membrane Order in a Calcium-Dependent Manner. Journal of Molecular Biology. 429 (24), 3875-3892 (2017).
- Millet, J. K., et al. Middle East respiratory syndrome coronavirus infection is inhibited by griffithsin. Antiviral Research. 133, 1-8 (2016).
- Millet, J. K., et al. A camel-derived MERS-CoV with a variant spike protein cleavage site and distinct fusion activation properties. Emerging Microbes, Infections. 5 (12), 126 (2016).
- Millet, J. K., Whittaker, G. R. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proceedings of the National Academy of Sciences. 111 (42), 15214-15219 (2014).
- Tse, L. V., Hamilton, A. M., Friling, T., Whittaker, G. R. A Novel Activation Mechanism of Avian Influenza Virus H9N2 by Furin. Journal of Virology. 88 (3), 1673-1683 (2014).
- Sun, X., Belouzard, S., Whittaker, G. R. Molecular architecture of the bipartite fusion loops of vesicular stomatitis virus glycoprotein G, a class III viral fusion protein. Journal of Biological Chemistry. 283 (10), 6418-6427 (2008).
- Millet, J., Whittaker, G. Murine Leukemia Virus (MLV)-based Coronavirus Spike-pseudotyped Particle Production and Infection. Bio-Protocol. 6 (23), 2035 (2016).
- Bonnin, A., Danneels, A., Dubuisson, J., Goffard, A., Belouzard, S. HCoV-229E spike protein fusion activation by trypsin-like serine proteases is mediated by proteolytic processing in the S2' region. Journal of General Virology. , (2018).
- Belouzard, S., Madu, I., Whittaker, G. R. Elastase-mediated activation of the SARS coronavirus spike protein at discrete sites within the S2 domain. Journal of Biological Chemistry. 285 (30), 22758-22763 (2010).
- Madu, I. G., Roth, S. L., Belouzard, S., Whittaker, G. R. Characterization of a Highly Conserved Domain within the Severe Acute Respiratory Syndrome Coronavirus Spike Protein S2 Domain with Characteristics of a Viral Fusion Peptide. Journal of Virology. 83 (15), 7411-7421 (2009).
- Madu, I. G., Belouzard, S., Whittaker, G. R. SARS-coronavirus spike S2 domain flanked by cysteine residues C822 and C833 is important for activation of membrane fusion. Virology. 393 (2), 265-271 (2009).
- Belouzard, S., Chu, V. C., Whittaker, G. R. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proceedings of the National Academy of Sciences. 106 (14), 5871-5876 (2009).
- Costello, D. A., Millet, J. K., Hsia, C. -. Y., Whittaker, G. R., Daniel, S. Single particle assay of coronavirus membrane fusion with proteinaceous receptor-embedded supported bilayers. Biomaterials. 34 (32), 7895-7904 (2013).
- Costello, D. A., Hsia, C. -. Y., Millet, J. K., Porri, T., Daniel, S. Membrane Fusion-Competent Virus-Like Proteoliposomes and Proteinaceous Supported Bilayers Made Directly from Cell Plasma Membranes. Langmuir. 29 (21), 6409-6419 (2013).
- Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., van Oijen, A. M. Single particle kinetics of influenza virus membrane fusion. Proceedings of the National Academy of Sciences. 105 (40), 15382-15387 (2008).
- Hsu, H. -. L., Millet, J. K., Costello, D. A., Whittaker, G. R., Daniel, S. Viral fusion efficacy of specific H3N2 influenza virus reassortant combinations at single-particle level. Scientific reports. 6, 35537 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。