
Method Article
使用新颖的数据非依赖型采集工作流程定量衰老秀 丽隐杆线虫 中的不溶性蛋白质聚集
摘要
这种新颖的工作流程以最少的起始材料从秀丽隐杆线虫中高效提取和分离 SDS 不溶性蛋白质(不溶性蛋白质),用于定量差异蛋白质组学分析。该方案使用全面的数据非依赖性采集质谱分析来量化不溶性菌群和生物信息学分析,以获得对衰老机制和病理学的生物学见解。
摘要
我们和其他人已经证明,衰老过程导致不溶性蛋白质在蛋白质组范围内积累。在 40% 以上的时间内敲除编码不溶性蛋白质的基因会导致秀丽隐杆线虫的寿命延长,这表明这些蛋白质中的许多是衰老过程的关键决定因素。这些不溶性蛋白质的分离和定量鉴定对于了解衰老过程中发生的关键生物过程至关重要。在这里,我们提出了一种经过修改和改进的方案,详细介绍了如何更有效地从秀丽隐杆线虫中提取和分离 SDS 不溶性蛋白质(不溶性蛋白质组学),以通过新颖的无标记定量蛋白质组学分析简化质谱工作流程。这种改进的方案利用高效的超声仪进行蠕虫裂解,大大提高了蛋白质提取的效率,并且与以前的方案(通常至少使用 40,000 条蠕虫)相比,我们可以使用明显更少的起始材料(约 3,000 条蠕虫)。随后使用数据依赖性采集 (DDA) 对不溶菌组进行定量蛋白质组学分析,用于蛋白质发现和鉴定,使用数据非依赖性采集 (DIA) 进行全面和更准确的蛋白质定量。定量蛋白质的生物信息学分析提供了潜在的候选者,可以很容易地用其他分子方法跟踪秀丽隐杆线虫。通过此工作流程,我们通常鉴定 1000 多种蛋白质并定量 500 多种蛋白质。这种新方案能够对秀丽隐杆线虫进行高效的化合物筛选。在这里,我们验证了这种改进的方案并将其应用于野生型秀丽隐杆线虫 N2-Bristol 菌株,并证实衰老的第 10 天 N2 蠕虫比第 2 天的幼虫表现出更大的不溶性菌群积累。
引言
蛋白质稳态随着衰老而逐渐下降,并导致蛋白质聚集增加 1,2,3。蛋白质聚集与多种神经退行性疾病有关,包括阿尔茨海默病、帕金森病、亨廷顿病和肌萎缩侧索硬化症4。衰老被认为是与蛋白质聚集相关的神经退行性疾病发病的主要危险因素。容易形成不溶性聚集体的蛋白质通常与细胞毒性和组织功能障碍有关,这可能会进一步加速其他蛋白质的聚集 5,6,7。或者,不溶性蛋白质聚集体可以激活细胞防御机制,从系统中去除蛋白质的有毒寡聚形式。在年龄相关疾病和正常衰老的情况下,敲除编码不溶性蛋白的选定基因可调节秀丽隐杆线虫 (C. elegans) 的寿命 5,8,9。因此,研究蛋白质聚集的细胞和分子机制对于了解衰老至关重要,并最终可能导致治疗神经退行性疾病的方法。
线虫秀丽隐杆线虫因其独特的特性,如寿命相对较短(约 2 周)、易于培养和遗传作,已成为研究衰老和年龄相关疾病中蛋白质聚集的最广泛使用的模式生物之一。
提取和表征不溶性蛋白质的能力在确定秀丽隐杆线虫模型中与蛋白质聚集相关的年龄相关变化方面发挥了关键作用。为了研究蛋白质聚集对正常衰老过程的贡献,我们5 和其他人2 之前提取并蛋白水解消化了年轻与老年秀丽隐杆线虫的不溶性菌,使用 iTRAQ 试剂进行化学标记(“用于相对和绝对定量的同量异位标记”),然后使用基于 MS 的方法进行定量。使用同量异位标记方法和 120 mg 湿蠕虫(约 40,000 条蠕虫),我们能够获得显著的蛋白质不溶性菌组深度和覆盖率5。定量分析表明,与幼虫的类似不溶菌组分相比,鉴定的 1200 种蛋白质中有 203 种在老年秀丽隐杆线虫的不溶菌组中显著富集5。David 等人还独立地利用 iTRAQ LC-MS/MS 工作流程来检查正常老化时蛋白质聚集体的变化2。从大约 300 毫克的蠕虫开始,他们使用两次生物重复鉴定了 ~1000 种不溶性蛋白质,并确定大约 1000 种蛋白质中的 ~700 种随着年龄的增长而积累了 1.5 倍或更多,与幼虫相比2。总体而言,这些独立结果表明,广泛的蛋白质不溶性和聚集性是正常衰老的固有部分,可能会影响寿命和神经退行性疾病的发病率 2,5。
研究不溶菌素使我们能够确定环境影响如何加速或减缓衰老过程。Klang 等人在秀丽隐杆线虫中建立了无标记蛋白质组学工作流程,以研究金属流失在长寿中的作用10。在这项研究中,至少使用了 40,000 条蠕虫来提取不溶性菌组10。数据显示,铁、铜、钙和锰的水平随着衰老而增加,给蠕虫喂食铁含量升高的饮食会显著加速与年龄相关的不溶性蛋白质积累10。使用相同的工作流程检查维生素 D 对秀丽隐杆线虫不溶性菌的影响,定量了年轻蠕虫中的 38 种蛋白质(第 2 天)和老年蠕虫中的 721 种蛋白质(第 8 天)。维生素 D 喂养显着将老年蠕虫的不溶性蛋白质从 721 种蛋白质降低到 371种蛋白质 11。进一步的研究表明,喂养维生素 D 抑制了随着年龄增长而产生的蛋白质不溶性,促进了蛋白质稳态,并延长了秀丽隐杆线虫 N2 野生型蠕虫的寿命11。因此,研究不溶菌组可以帮助确定衰老和年龄相关疾病的新型调节剂。
虽然研究不溶菌组对于促进对老化过程的理解非常宝贵,但由于需要收集大量起始样品材料而受到阻碍。Groh 等人最近推出了一种无标记蛋白质组学定量工作流程,以研究秀丽隐杆线虫随衰老的固有蛋白质聚集变化;然而,它需要大量的起始材料(350 毫克地虫)12。在本报告中,我们建立了一种改进的新提取和分离方案(图 1)。在蠕虫裂解过程中使用高效的超声仪显著提高了提取效率,随后将所需的起始材料量从 40,000 个蠕虫减少到 3,000 个蠕虫。将这种新型不溶性菌组分离方案与无标记数据非依赖型采集 (DIA) 质谱工作流程相结合,显著提高了蛋白质深度和覆盖度。这里介绍的协议具有成本效益且易于修改,以允许在其他模型系统中进行不溶菌组分析。
研究方案
注意:为了更好地理解实验程序,请参见 图 1 以获取工作流程示意图。
1. 同步衰老秀丽隐杆线虫的大规模培养
- 板制备
- 将 30 mL NGM 琼脂(含或不含 50 mM 氟脱氧尿嘧啶 (FUdR))倒入每个 90 mm 板中,让板在培养罩中干燥 2 天。
- 在接种 NGM 琼脂平板的前一天,将 50 μL OP50 细菌菌株的冷冻甘油原液等分试样接种到 2 L 烧瓶中的 1 L 高压灭菌的 LB 培养基中。
- 将细菌在 37 °C 下在振荡培养箱中以 250 rpm 培养 16 小时。
- 16 小时后,以 5,000 x g 的速度离心细菌培养物 10 分钟。
- 弃去上清液,将细菌沉淀完全重悬于 40 mL 高压灭菌的 S 基础溶液(含有 50 mM K3PO4、100 mM NaCl 的水溶液)中,以产生 25x 浓缩的 OP50 大肠杆菌培养物。
- 将 2 mL 的 25x 浓缩细菌培养物分配到每个 NGM 琼脂平板上并均匀涂抹,使细菌培养物覆盖整个琼脂表面。
- 在细菌培养罩中从板上取下盖子 2-3 小时,直到板正确干燥。
注意:确保每 15 分钟旋转一次板,使细菌均匀分布。这很重要,因为细菌往往集中在板的一侧,导致细菌密度低的区域。 - 板子干燥后,盖上盖子,将板子在室温下放置 48 小时,以确保 OP50 大肠杆菌食物源完全干燥。
注意:板可以提前 2 周准备,并在使用前储存在 4 °C。
- 秀丽隐杆线虫同步队列的维护和准备
- 为了获得分离 SDS 不溶性组分所需的蛋白质裂解物 (1.0–1.5 mg) 起始量,每个样品使用大约 3,000 个同步蠕虫。
- 将~100 N 2 野生型妊娠成虫转移到NGM 25x OP50 大肠杆菌种子板中,并将蠕虫在20°C下维持~72小时,直到板中充满妊娠成虫。
- 将 ~10 mL 的 S 基础溶液移液到每个板上,并将所有蠕虫收集到 15 mL 试管中。
注意:在此步骤和所有涉及蠕虫收集的未来步骤中,使用低吸附吸头或巴斯德移液器非常重要,以防止因附着在吸头表面而导致蠕虫损失。 - 将样品以 520 x g 旋转 30 秒。旋转后去除上清液。要去除任何 OP50 大肠杆菌,向沉淀中加入 10 mL S 基础溶液并再次旋转,然后去除上清液。重复此步骤。
- 将 10 mL 次氯酸钠漂白液(含 0.5 M KOH 和 0.48% 次氯酸钠的水溶液)添加到 15 mL 锥形管中的蠕虫沉淀中,并在室温下剧烈涡旋 2 分钟。
- 在室温下以 520 x g 离心 30 秒,以离心掉任何未完全溶解的蠕虫体和虫卵。然后去除上清液。
- 重复步骤 1.2.5 和 1.2.6 以确保完全去除所有蠕虫体并释放虫卵(在显微镜下观察管以确保)。
- 用 10 mL S 基础溶液洗涤鸡蛋沉淀至少 4 次,以确保完全去除漂白剂溶液。
注意:完全去除漂白剂很重要,因为残留的漂白剂可能会阻碍种蛋的孵化。 - 将鸡蛋重悬于 15 mL 试管中的 5 mL S 基础培养基中,并置于 20 °C 的转子中 24 小时,让鸡蛋孵化。
- 24 小时后,取 10 μL S 基础幼虫溶液计算其中的蠕虫数量。使用至少三个单独的等分试样 10 μL S 基础幼虫溶液来复制蠕虫计数。计算三种幼虫浓度的平均值。根据平均浓度,稀释蠕虫溶液,使每 200 μL 溶液得到 ~3,000 只幼虫。
注意:确保倒置轻轻摇晃试管,因为如果不打扰,幼虫往往会沉淀下来,这会影响幼虫数量。 - 充分混合并将 200 μL S 基础幼虫溶液添加到每个 90 mm NGM 细菌接种板中。
注:在整个板中加入 S 基础幼虫溶液滴剂,以使幼虫均匀分布,以避免某些部位的细菌耗竭。 - 让 S 基础溶液干燥,然后将板倒置在 20 °C 培养箱中 48 小时。
- 48 小时后,在 S 基础溶液中从板中收集蠕虫,并将其转移到含有 50 mM FUDR 的新鲜 NGM 种子板中,并在 20 °C 下孵育板。
注意:在 L4 阶段的早期转移蠕虫以确保 FUDR 效率非常重要。 - 48 小时后,通过在板上加入 10 mL S 基础溶液并将溶液转移到 15 mL 试管中来收集第 2 天的年轻成人样品。让蠕虫在重力作用下沉降,并在 S 基础溶液变清后将其去除。
- 在 5 mL S 基础溶液中洗涤蠕虫沉淀 2-3 次,以去除任何附着的细菌。去除尽可能多的 S 基础溶液,并将蠕虫沉淀(在 15 mL 试管中)冷冻在干冰/乙醇浴中。将冷冻样品储存在 -80 °C。
- 每隔一天将样品蠕虫转移到含有 FUDR 的新鲜 NGM OP50 大肠杆菌板中,直至第 10 天。按照步骤 1.2.14 和 1.2.15 中的说明收集和冻结蠕虫。
注意:对于第 10 天的样品,从更多的蠕虫数量开始,以解决由于转移和挖洞而导致的蠕虫数量损失。
2. 从 蠕虫中提取 SDS 不溶性组分
- 在 900 μL 冰冷的蠕虫裂解缓冲液混合物中解冻冷冻蠕虫沉淀,该混合物含有 20 mM Tris 碱、pH 7.4、100 mM NaCl、1 mM MgCl2 和不含 EDTA 的蛋白酶抑制剂。短暂涡旋样品,以确保蠕虫沉淀在裂解缓冲液中完全重悬。
- 将 15 mL 试管放入超声处理槽中,并以高强度设置为 10 个超声处理循环(每个循环 30 秒开,关 30 秒)。重复最多五个周期(确保蠕虫完全裂解)。
- 在冷藏室中以 3,000 x g 旋转裂解物 4 分钟。离心后,丢弃含有任何蠕虫碎片的沉淀,并小心地将上清液转移到冰上新鲜预冷的 1.5 mL 离心管中。
- 使用 BCA 测定定量蛋白质浓度。将裂解物等分试样(通常含有约 1 mg 蛋白质)转移到新鲜的预冷 1.5 mL 离心管中。
- 在冷藏室中以 20,000 x g 的速度将蛋白质裂解物离心 15 分钟。
- 在不干扰沉淀的情况下,将上清液转移到新鲜的 1.5 mL 离心管中,并将其保存为水溶性蛋白质组分。
- 在室温下用 500 μL 含有 1% SDS 的蠕虫裂解缓冲液洗涤上一步的沉淀,并在室温下以 20,000 x g 离心 15 分钟。去除上清液并保存为 SDS 可溶性组分。重复此洗涤步骤两次以去除任何 SDS 可溶性组分。第三轮洗涤后剩余的沉淀定义为 1% SDS 不溶性蛋白质组分。
- 将 SDS 不溶性蛋白沉淀重悬于 60 μL 70% 甲酸中,并剧烈涡旋以溶解蛋白质。根据需要多次重复剧烈涡旋以溶解沉淀。
- 在室温下在超声仪水浴中对沉淀物进行超声处理 30 分钟。
注:SDS- 不溶性沉淀通常难以溶解在缓冲液中。然而,最终应在此步骤中溶解整个沉淀。 - 在真空浓缩器中干燥样品 1 小时以完全去除甲酸溶液。
- 向干燥的沉淀中加入 40 μL 的 1x LDS 样品凝胶缓冲液,并将样品加热至 95 °C 10 分钟。短暂涡旋并旋转样品。将 13 μL 上样到 4-12% NUPAGE Bis-Tris 凝胶上并运行凝胶。用荧光蛋白染色剂对凝胶进行染色以进行成像。保存样品的其余部分用于质谱 (MS) 分析。
- 将剩余样品加载到 4-12% Bis-Tris 凝胶中,并运行约 20 分钟以进行 MS 凝胶内消化。
3. 使用胰蛋白酶进行凝胶内消化,以分离用于 MS 分析的蛋白质
- 制备以下溶液(新鲜):25 mM NH4HCO3 (pH 7–8)、25 mM NH4HCO3 溶于 50% 乙腈(ACN,pH 7–8)和 50% ACN 溶于 5% 甲酸中。
- 将每个凝胶切片切成小块(通常为 <1 mm2),然后将它们放入 0.65 mL 硅化管中。
- 加入约 100 μL(或足以覆盖)25 mM NH4HCO3/50% ACN 溶液,并在室温下涡旋 10 分钟。提取上清液并转移到单独的试管中(丢弃)。再重复此步骤两次。
- 在真空浓缩器中完全干燥凝胶块(~20 分钟)。
- 制备新鲜溶液,向干燥的凝胶块中加入 ~100 μL(或足以覆盖)25 mM NH4HCO3 中的 10 mM DTT(使用新鲜制备的 NH4HCO3)。涡旋并短暂旋转。让反应在 56 °C 下进行 1 小时,并在混合器上以 1400 rpm 振荡。
- 去除上清液,向凝胶块中加入 100 μL 55 mM 碘乙酰胺 (IAA) 的 25 mM NH4HCO3 溶液。涡旋并短暂旋转。让反应在 RT 下在黑暗中进行 45 分钟。
- 去除上清液并丢弃。加入 ~100 μL 25 mM NH4HCO3 并涡旋 10 分钟洗涤凝胶块。短暂旋转并去除上清液,丢弃后者。
- 向凝胶块中加入 ~100 μL(或足以覆盖)25 mM NH4HCO3 的 50% 乙腈溶液,使凝胶块脱水,并涡旋 10 分钟。然后短暂旋转以去除并丢弃上清液。如果凝胶块不是很干燥,则重复此步骤两次甚至第三次,如凝胶块的较小尺寸和浑浊的白色所示。
- 在真空浓缩器中完全干燥凝胶块(~20 分钟)。
- 最初向每个样品中加入 15 μL 胰蛋白酶溶液(250 ng 胰蛋白酶),然后加入足够的 25 mM NH4HCO3 以覆盖凝胶块 (~100 μL)。涡旋 10 分钟。然后在冷藏室中短暂旋转并在 4 °C 下孵育 30 分钟,不要混合。
- 根据需要加入 25 mM NH4HCO3 以完全覆盖凝胶块。在混合器上以 1400 rpm 的转速在 37 °C 下旋转孵育过夜 16-20 小时。
- 第二天,短暂涡旋并旋转消化液。加入约 100 μL HPLC 级水,离心,封口膜管并连续超声处理 10 分钟。超声处理后短暂离心。
- 将代表水萃取的消化液转移到干净的 0.65 mL 硅化管中。
- 向凝胶块中加入 ~100 μL 50% ACN/5% 甲酸(足以覆盖它们),在室温下涡旋 10 分钟,短暂旋转并收集溶液,然后重复一次。将包含此步骤和上一步 3.13 中提取的肽的所有溶液汇集到一个试管中。
- 涡旋提取的消化物。在真空浓缩器中完全干燥肽 (~2 h)。
- 加入 30 μL 0.2% 甲酸,将肽在冷藏室的混合器上重悬 10 分钟。
- 在室温下以 1850 x g 离心样品 5 分钟。吸出肽溶液并移入新的干净的 0.65 mL 硅化管中。然后使用 C18 脱盐探针对肽溶液进行脱盐(见下文)。
4. 使用 C18 脱盐针对消化肽进行脱盐
- 将移液器设置为 10 μL 并连接 C18 脱盐吸头。吸取 10 μL 100% ACN,然后将其丢弃,润湿脱盐吸头。重复此步骤 2 次。
- 用移液液吸取 10 μL 的 50% 乙腈、49.8% 水和 0.2% FA,然后丢弃,清洗 C18 脱盐吸头。重复此步骤 2 次。
- 通过吸取 10 μL 0.2% FA 的水中,然后将其丢弃来平衡 C18 脱盐吸头。重复此步骤 2 次。
- 将移液管设置为 10 μL,并通过在树脂中上下移液 15 次,将消化的肽从溶液中加载到树脂中。
注:重复作将确保所有肽与脱盐吸头的树脂结合。 - 用 10 μL 0.2% FA 的水中吸取 10 μL,然后丢弃,对与树脂结合的消化肽进行脱盐。重复此步骤 4 次以完成脱盐步骤。
- 在新试管中,用 10 μL 50% ACN、49.8% 水和 0.2% FA 上下移液 10 次洗脱肽。重复此步骤一次,在同一管中第二次洗脱蛋白质消化物。
- 在真空浓缩器中完全干燥脱盐肽(~20 分钟)。
- 在 15 μL 0.2% FA + 1 μL iRT(索引保留时间)肽中重悬肽。Voxtex 10 分钟,以 12,000 x g 离心 2 分钟,然后转移到自动进样器样品瓶中进行 MS 分析(见下文)。
5. 使用 DDA 和 DIA 对消化肽进行质谱分析
注:可以使用 DDA 或 DIA LC-MS/MS 方法分析样品。在本研究中,使用与高分辨率质谱仪耦合的 nano-LC 2D HPLC 系统对样品进行分析。
- 将 HPLC 系统与直接连接到四极杆飞行时间质谱仪的芯片式 HPLC 系统结合使用(也可以使用其他 LC-MS 配置和系统)。
- 使用反相 HPLC-ESI-MS/MS 分析样品。
- 构建色谱方案,将肽样品加载到 C18 柱前芯片上。用上样溶剂(0.1% 甲酸)以 2 μL/min 的流速洗涤和脱盐 10 分钟。
- 将肽转移至分析柱 C18 芯片中,使用流动相 A(2% 乙腈,0.1% 甲酸)和 B(98% 乙腈,0.1% 甲酸),以 300 nL/min 的流速以 3 小时的梯度洗脱。第一个洗脱步骤包括在 80 分钟内从 5% B 到 35% B 的线性梯度。
- 在 5 分钟内将流动相 B 升至 80%,然后将流动相 B 保持在 80% 8 min,然后变为 5% B,以重新平衡色谱柱 25 min。
- 为 DDA 设置 MS 仪器方法并定义仪器参数,如下所示。
- 对实验 1 使用以下参数:从 m/z 400–1500 扫描 MS1 母离子(累积时间为 250 ms);设置强度阈值以触发电荷态 2–5 至 200 计数的离子的 MS/MS 扫描;将母离子的动态排除设置为 60 s)。
- 对实验 2 使用以下参数:MS2 扫描范围为 m/z 100–1500 的 MS/MS 子离子扫描(每个循环 30 次子离子扫描中,每次累积时间为 100 ms);将碰撞能量扩散设置为 CES=5,并勾选“高灵敏度产物离子扫描模式”。
注意:DDA 采集方法用于构建光谱库,如步骤 6.3 中所述(见下文)。在这里,它将在每个循环的每次 MS1 扫描后采集 30 个丰度最高的母离子的 MS/MS 谱图;总循环时间约为 3.3 s。
- 获取 DIA。为 DIA 设置 MS 仪器方法并定义仪器参数,如下所示。
- 对实验 1 使用以下参数:从 m/z 400-1250 扫描 MS1 母离子(累积时间为 250 ms)。
- 对实验 2 使用以下参数:对 64 个可变 SWATH 片段进行 MS/MS 子离子扫描,MS2 扫描范围为 m/z 100-1500(每个循环 64 个子离子扫描中每个片段的累积时间为 45 毫秒);将碰撞能量扩散设置为 CES=10,勾选“高灵敏度子离子扫描模式”。
注:使用 Schilling 等人 13 描述的 64 变量窗口 DIA/SWATH 采集方法进行无标记定量,总循环时间约为 3.2 s。对于 DDA 采集,不是使用 Q1 四极杆将较窄质量范围的母离子传输到碰撞池,而是使用可变窗口宽度范围(例如 m/z 5–90)在整个 m/z 范围 (m/z 400–1250) 上递增步进,具有 64 个 SWATH 段,每个段的累积时间为 45 ms, 循环时间为 3.2 秒,其中包括一次累积时间为 250 ms 的 MS1 扫描。使用 SCIEX“可变窗口计算器”算法13 ,根据在特定 m/z 范围内观察到的典型 MS1 离子电流的复杂程度调整可变窗口宽度(较窄的窗口适用于“繁忙”的 m/z 范围,较宽的窗口适用于洗脱母离子较少的 m/z 范围)。在其他 MS 平台上,也可以实施替代的 DIA 窗口选择策略。
6. 数据分析
注意:某些数据分析设置应根据特定的实验条件进行定制。例如,选择的蛋白质数据库(fasta 文件)将取决于制备样品的物种(在本方案中我们使用秀丽隐杆线虫)。
- 使用 MS 数据库搜索引擎 分析 DDA 采集并鉴定蛋白质。生成 darabase 搜索引擎方法,如下所示:
- 对于 Sample Description Parameters (样品描述参数),选择 Identification(鉴定);在 Sample Type(样品类型)下,选择 Iodoacetic Acid(碘乙酸);在 Cysteine Alkylation 下,选择 Trypsin;在 Digestion(酶解)(假设赖氨酸和精氨酸处 C 端裂解)下,在 Instrument(仪器)下选择质谱仪的名称;,然后在 Species (物种) 下选择 Caenorhabditis elegans (秀丽隐杆线虫)。
- 对于 Specific Processing parameters(特定加工参数),选中 Biological modifications(生物修饰);在 ID Focus 下,选择 SwissProt;在 数据库 下,检查 彻底 ID;在 Search Effort(搜索工作量)下,在 Detected Protein Threshold(检测到的蛋白质阈值)下选择 0.05 (10%);,然后选中 Results Quality 下的 Run False Discovery Rate Analysis。保存搜索引擎方法,并使用生成的方法将 MS 原始文件提交到数据库搜索引擎进行处理。
注:在迭代过程中,所有 MS 和 MS/MS 扫描均由搜索引擎根据初始注释和结果自动重新校准。
- 完成搜索后,单击 “导出肽段摘要 ”,并在 Excel 中按置信阈值 99(错误发现率 (FDR) = 1%)筛选所有肽段鉴定结果。
- 构建 MS/MS 谱库,用于进一步处理 DIA 原始数据文件,并进一步进行相对数据定量。
- 打开 DIA 定量分析软件。选择“ 库 ”选项卡,然后(在页面底部)单击 “数据库搜索引擎”中的“生成光谱库”,并打开“数据库搜索引擎”FDR 报告(*FDR.xlsx 电子表格文件),该报告是在 DDA 原始数据文件数据库搜索过程中自动生成的。
- 单击 下一步,选择 Library Settings Schema 并单击 Next。选择 Uniprot_Caenorhabditis 秀丽隐杆 线虫_proteome作为数据库,然后单击 Finish,将生成光谱库。
- 使用 DIA 定量分析软件 分析 DIA 采集,以进行全面的相对蛋白质定量。DIA 数据分析管道说明如下。
- 要分析和定量肽,请打开 DIA 定量分析软件 ,使用模板架构设置分析架构。模板架构可在软件中通过单击 “设置”,然后单击 “DIA 分析”,最后单击“ BGS 出厂设置”来使用。
- 在模板架构中设置参数,如下所示:
1) 在 识别 下选择 PTM 定位 (概率截断 = 0.75)。
2) 在 定量下,选择 “主组数量” 作为肽 总和数量,将 主组 N 设置为 最大 7 和 最小 1,选择 次组数量 作为 母离子总和数量,将 次组 N 设置为 最大 10 和 最小 1,并选择 “数据过滤 ”作为 Qvalue 稀疏 ,不要选择 交叉运行归一化。
- 执行相对定量分析。
- 选择 “管道” 选项卡,单击“ 从文件设置 DIA 分析”,打开感兴趣的 MS DIA 原始文件(例如:样品和对照),单击 “分配光谱库”,选择在 6.3 中生成的库,单击 “加载”,最后单击 “下一步”。
- 选择在 6.5.1 中设置的分析架构,然后单击 Next。选择适当的数据库 fasta 文件(在此协议中:Uniprot_秀丽隐杆线虫_proteome)并单击下一步。定义条件设置,并根据样本分配不同的条件,然后单击 Next。
- 查看分析概述(实验设置摘要)并浏览到 Output Directory(输出目录 ),然后单击 Finish(完成)。最后,单击 Run Pipeline 以执行无标记定量分析。
注:“DIA 定量分析软件”中的统计模块自动执行 FDR 分析,生成比较不同条件的热图和火山图,生成已鉴定和定量的肽和蛋白质列表,并提供 Q 值以及比较不同条件的相对倍数变化。在这里,样品条件设置包括第 10 天老蠕虫和第 2 天幼蠕虫,其中两个生物重复作为对照。
结果
传统的蠕虫裂解方法有各种缺点。例如,基于探针的超声处理和磁珠打浆法允许金属尖端或磁珠直接与样品接触,从而产生过多的热量,从而导致不同的蛋白质回收率和蛋白质变性。液氮研磨,然后在裂解缓冲液中超声处理,可能非常耗时,并且需要大量的蠕虫。由于传统蠕虫裂解方法的局限性,以前的 MS 工作流程,例如历史上在秀丽隐杆线虫模型系统中使用的标记方法 iTRAQ 或无标记方法,以获取有关不溶菌的定量信息,需要大量的起始材料(至少 40,000 个蠕虫)。需要费力的蠕虫培养工作才能获得这些数量的蠕虫。此外,标记方法需要昂贵的同量异位化学标签。免标记定量方法具有成本效益,并且具有更简单、更直接的样品制备和标记方法,但需要大量蠕虫才能获得足够的 MS 分析覆盖率。
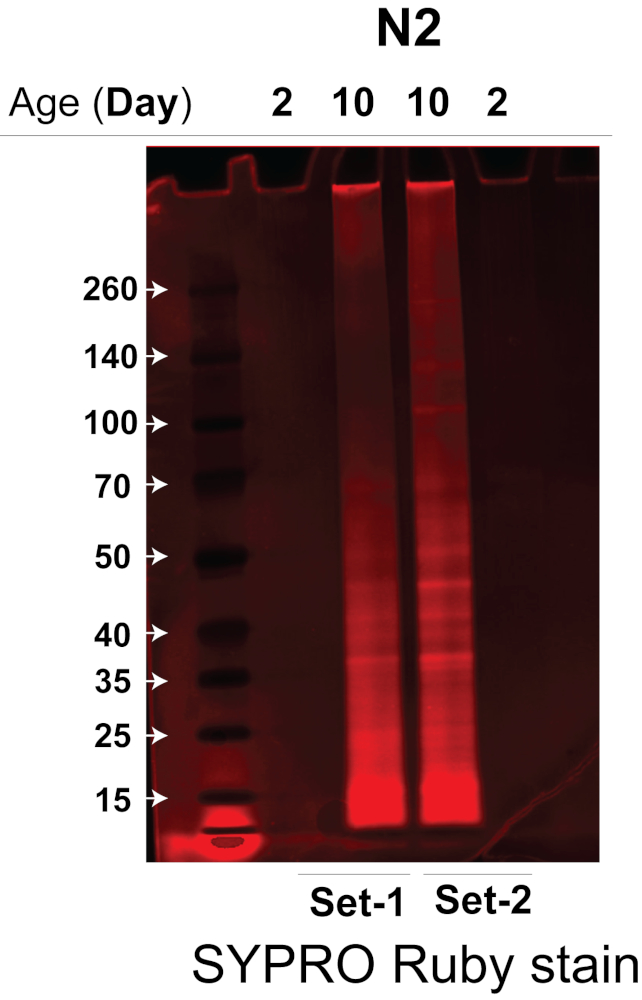
我们使用的超声仪通过在温控水浴超声仪中同时裂解多个蠕虫样品,大大提高了蠕虫裂解的效率和可重复性14,从而显著减少了所需的起始蠕虫材料量。结合高效的超声处理方法和定量 DIA 无标记 MS 方法,我们能够使用 ~3,000 条蠕虫对老年和年轻蠕虫的不溶性菌进行稳健定量。在这里,我们测试和验证了该方案的效率,并比较了来自野生型蠕虫菌株 N2-Bristol C. elegans 的老年和年轻蠕虫的不溶性。我们应用该方案从 ~3,000 个老年和年轻的 N2 秀丽隐杆线虫中提取和分离不溶菌组(每种条件两个生物学重复),然后使用四极杆飞行时间质谱仪或其他 MS 系统进行 MS 分析,使用数据依赖性采集 (DDA) 和数据非依赖性采集 (DIA/SWATH) 的组合进行蛋白质鉴定和定量。首先在 Bis-Tris 4-12% 梯度凝胶上分析不溶性蛋白质,以确定每个不溶性样品中的蛋白质量。如图 2 所示,来自 N2 老年蠕虫的不溶菌组样品(泳道 2 和 3,生物复制实验)的蛋白质明显高于来自 N2 幼蠕虫的样品(泳道 1 和 4,生物复制实验)。
凝胶内消化后,通过 HPLC-MS 分析不溶菌组的蛋白质谱。使用此工作流程,我们通常可以鉴定 1000-1500 种蛋白质,并从 SDS 不溶性组分中定量 500-1,000 种蛋白质,重现性高(未发表的数据)。在这里,我们能够通过分析 DIA 数据并去除冗余来量化来自 N2-Bristol C. elegans 不溶菌组中的 989 种蛋白质:768 种蛋白质在老年 N2 蠕虫(第 10 天)的不溶性中显着富集,27 种蛋白质显着减少与年轻相比(第 2 天),使用至少 1.5 的倍数变化和小于 0.01 的 Q 值(图 3A)。如直方图所示(图 3B),显著改变的蛋白质的倍数变化呈正态分布。老蠕虫被证明对不溶性蠕虫显著富集:观察到的最大变化表明,老蠕虫中不溶性蠕虫的相对蛋白质丰度是年轻蠕虫的 592 倍;对于 32 种蛋白质,老蠕虫中不溶性菌组中的相对蛋白质丰度比年轻蠕虫高 >250 倍,表明不溶性溶性蠕虫随年龄增长而发生巨大变化。
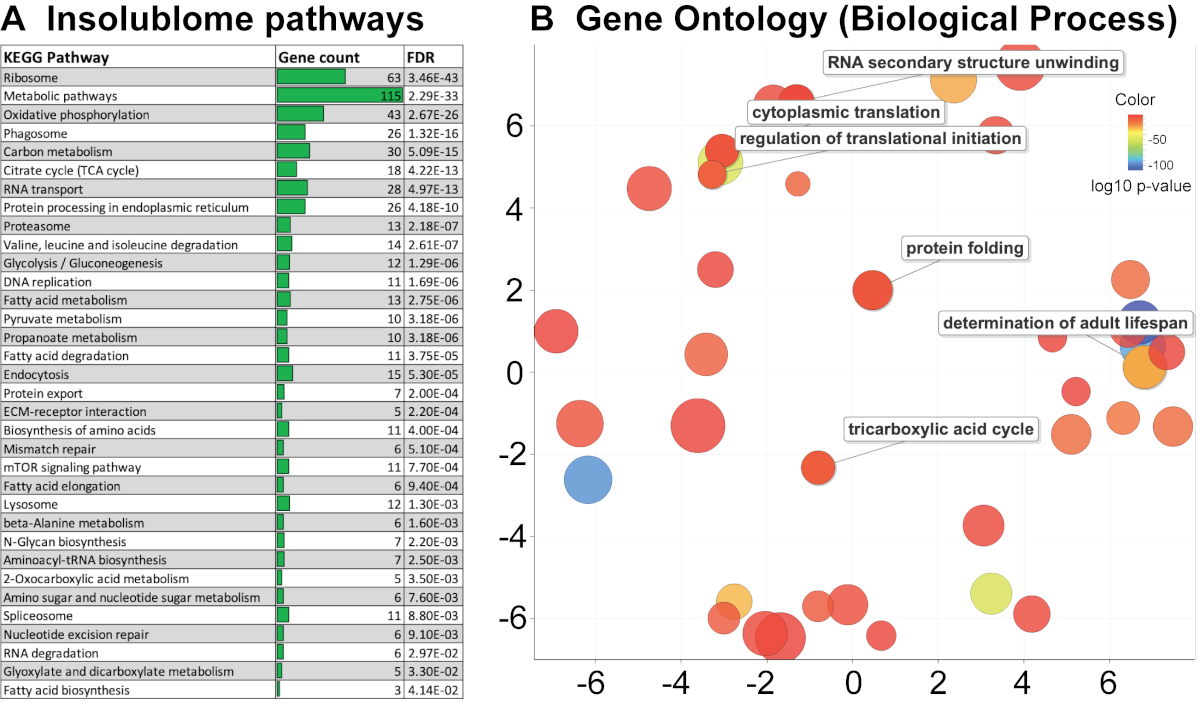
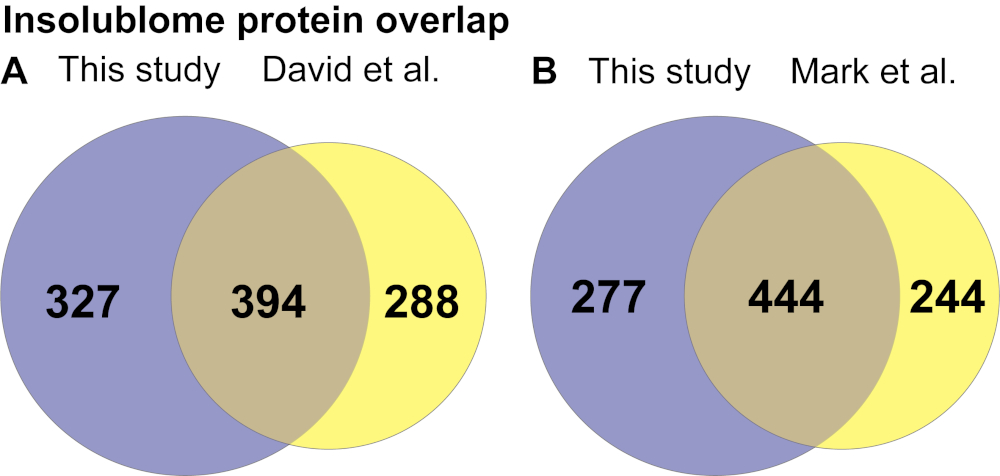
在提取了在老年蠕虫中显着增加并由蠕虫基地 (WS271) 鉴定的不溶性蛋白质列表后,进行了 KEGG 通路和基因本体论 (GO) 分析,以确定在老年不溶性蠕虫中富集的通路,以获得关于这些与衰老的关系的生物学见解。本研究中鉴定的蛋白质的 KEGG 通路分析显示,涉及核糖体、线粒体、蛋白酶体和剪接体的几种通路富集(图 4A)。基因本体分析表明,来自老年蠕虫的不溶性蛋白质包括许多特定类别的蛋白质,包括线粒体、发育、成虫寿命的决定因素和核糖体蛋白(图 4B 和 补充表 1A)。然后,我们将本研究中鉴定的蛋白质列表与 David 等人 2 和 Mark 等人 11 先前发表的工作进行了比较,如维恩图所示(图 5A,5B)。比较显示已鉴定的蛋白质 394/721 和 444/721 分别与 David 等人(图 5A)和 Mark 等人(图 5B)的研究显着重叠。本研究中不溶菌胺的 KEGG 分析揭示的生物途径在过去也已被确定,从而验证了我们的方法(补充表 1B)。这些途径和蛋白质的鉴定表明,它们可能作为进一步生物学研究它们在衰老背景下功能的候选者。
总之,使用高效的超声处理方法能够在温度控制良好且交叉污染减少的环境中同时裂解多个蠕虫样品,从而以显著减少起始蠕虫材料实现高蛋白质覆盖率。将高效的超声处理方法与 DIA 免标记蛋白质定量工作流程相结合,为不溶性蠕虫蛋白的定量提供了可靠且可重现的结果。

图 1.协议的实验工作流程。秀丽隐杆线虫在不同日期培养和收集。用超声仪裂解蠕虫后,提取 1% SDS 不溶性蛋白质组分(不溶菌蟟)并从裂解物中分离。然后通过凝胶内胰蛋白酶消化消化不溶性菌素,并通过 DIA 质谱定量,然后进行生物信息学分析。请单击此处查看此图的较大版本。
{kind=link}

图 2.不溶菌组分离的 N2-Bristol 菌株的年轻蠕虫与老蠕虫的 SDS-PAGE 凝胶。 通过 SDS-PAGE 分析 N2-Bristol 菌株的年轻蠕虫与老蠕虫的不溶物,以确定存在的蛋白质量。SDS-PAGE 凝胶用荧光蛋白染色剂染色以观察蛋白质条带。泳道 1 和 4:来自年轻 N2 蠕虫的两次生物重复实验的不溶菌组(第 2 天)。泳道 2 和 3:来自衰老 N2 蠕虫的两次生物重复实验的 Insolublome(第 10 天)。 请单击此处查看此图的较大版本。
{kind=link}

图 3.确定的候选蛋白显示老年人与年轻不溶菌组及其倍数变化分布的显着变化。 (A) 用于量化老年 N2-Bristol 蠕虫与年轻 N2-Bristol 蠕虫不溶物组的火山图。绝对折叠变化 >=1.5 且 Q 值为 <0.01 的候选人显示为红点。(B) 老蠕虫样品与幼虫样品中显著富集的 SDS 不溶性蛋白的倍数变化分布的直方图。 请单击此处查看此图的较大版本。
{kind=link}

图 4. KEGG 通路和基因本体论 (GO) 分析。 (A) 根据 p 值排列的第 10 天不溶菌组的 KEGG 通路分析,其中高度显著的通路显示在顶部。(B) 基因本体分析表明,老年蠕虫的不溶性富含许多特定类别的蛋白质,包括线粒体、发育、成虫寿命的决定因素和核糖体蛋白。散点图视图在 “语义空间” 中可视化 GO 术语,其中更相似的术语位于更近的位置。气泡的颜色反映了在 STRING 分析中获得的 p 值,而其大小反映了 GO 项在 UniProt-GOA 数据库中的通用性。 请单击此处查看此图的较大版本。
{kind=link}

图 5. 将本研究与 (A) David 等人 2 和 (B) Mark 等人 11 的研究进行比较,在第 10 天 Insolublome 蛋白重叠中发现。 请单击此处查看此图的较大版本。
{kind=link}
补充表 1 (与 图 4 和 图 5 相关)。 (A) 用 STRING 数据库分析的基因本体论(生物过程)。 (B) 本研究中鉴定的蛋白质和 KEGG 通路的详细列表,颜色代码描述了它们与已发表工作的重叠。 请点击此处下载此文件。
讨论
在该方案中,我们报告了一种改进的样品制备方法,用于从秀丽隐杆线虫中提取不溶性蛋白质。通过用高效的超声仪取代传统的蠕虫裂解(例如探针超声处理或磁珠打浆技术),我们提高了不溶性蛋白质提取的产量,并将无标记 MS 分析所需的蠕虫数量从 40,000 条蠕虫减少到 3,000 条蠕虫。使用数据库搜索引擎从 DDA 数据中鉴定蛋白质,并使用“DIA 定量分析软件”和相应的 DDA 数据库搜索结果(导入数据库搜索引擎生成的 FDR 文件报告)构建秀丽隐杆线虫光谱库。使用“DIA 定量分析软件”对老年和年轻的秀丽隐杆线虫不溶性数据进行相对量化,以处理新的 DIA 数据集和生成的光谱库。
协议中有几个步骤至关重要。与其他真核生物(如哺乳动物细胞)相比,秀丽隐杆线虫的寿命较短,使其成为研究衰老的理想系统,但在研究衰老相关现象时,分离同质的蠕虫种群至关重要。该协议中使用 FUDR 来获得同步老化蠕虫。重要的是将蠕虫在其 L4 阶段的早期转移到含有 FUDR 的 NGM 种子板上,以确保其有效性。在使用高效超声仪进行蠕虫裂解过程中,水浴温度必须设置为 4 °C,超声处理必须设置为 30 秒开启和 30 秒关闭,以防止样品过热。在第一轮超声处理 10 个循环(10 分钟)后,请务必在显微镜下检查以确保所有蠕虫都得到有效裂解。如果没有,则需要更多的超声处理循环。在凝胶内消化过程中,必须将每个凝胶切片切成适当大小 (<1 mm2) 的碎片——如果太小,它们可能会在样品制备过程中丢失,如果太大,则可能消化不充分。
对起始材料的需求大大减少了与蠕虫培养相关的费力工作,以获得用于不溶菌组分析的样品。然而,提取和分离 1% SDS 不溶性蛋白质组分涉及多个洗涤步骤,需要小心处理样品,以避免样品损失并确保结果的可重复性。用于 MS 分析的材料量足以进行 ~3 次进样,用于后续的 DDA 和 DIA 分析,但不能留作将来的实验使用。此外,尽管它可能产生混杂效应15,但我们在老化过程中使用了尽可能低浓度的 FUdR 对蠕虫进行消毒。未来的研究可能会通过使用无菌突变体或手动转移和收集蠕虫来规避 FUdR 的使用。
使用高效的超声仪进行蠕虫裂解,可以高效提取不溶菌团,从而获得良好的蛋白质覆盖率和经济高效的无标记 DIA MS 分析,以使用大大减少的蠕虫数量来定量不溶菌团。它显著减少了工作量,允许每个实验筛选更多条件。此外,免标记 MS DIA 工作流程具有成本效益,可提供与 iTRAQ、TMT 或 SILAC 等标记方法相当的蛋白质深度和覆盖度。秀丽隐杆线虫模型是一种用于衰老研究的快速筛选系统。该工作流程可以轻松修改并应用于研究该生物体和其他生物体的衰老和与年龄相关的疾病研究。例如,在正在进行的研究中,我们正在应用此工作流程来研究各种阿尔茨海默病 (AD) 秀丽隐杆线虫模型中不溶菌组和蛋白质稳态的蛋白质谱,包括 Abeta、tau 和双重 Abeta/tau 蠕虫,有或没有不同的药物干预,用于未来的高通量药物筛选。
披露声明
作者没有什么可披露的。
致谢
这项工作得到了 TripleTOF 系统(1S10 OD016281,巴克研究所)、NIH 赠款、RF1 AG057358 (GJL, JKA) 和 NIH 赠款U01AG045844 (GJL) 的 NIH 共享仪器赠款的支持。XX 得到 T32 博士后奖学金(NIH 资助 5T32AG000266,PI:Judith Campisi 和 Lisa Ellerby)的支持。MC 得到了 Larry L. Hillblom 基金会的博士后奖学金的支持。
材料
| Name | Company | Catalog Number | Comments |
| Strains used | |||
| Esherichia coli OP50 | Caenorhabditis Genetics Center (CGC) | ||
| N2 (Bristol) | Caenorhabditis Genetics Center (CGC) | ||
| Buffer/Solution | |||
| NGM (Nematode Growth Media) | Recipe: 3 g/L NaCl, 23 g/L agar; 2.5 g/L peptone; 1 mM CaCl2, 5 mg/L cholesterol, 1 mM MgSO4, 25 mM KH2PO4 | ||
| S-basal solution | Recipe: 5.85 g/L NaCl, 1g/L K2HPO4, 6 g/L KH2PO4, H2O to 1 L | ||
| Sodium hypochlorite bleach solution | Recipe: Mix 0.5 mL 5 N NaOH with 1 ml Sodium hypochlorite (5%) and make volume to 5 mL with H20. | ||
| Material/ Equipment | |||
| Agar | Difco Granulated Agar, BD Biosciences | 90000-782 | |
| Bioruptor Plus sonication device | Diagenode, USA | B01020001 | |
| Cholesterol | Sigma | c8503 | |
| 2'-deoxy-5-fluorouridine | VWR | TCD2235 | |
| Glycerol | Millipore Sigma | 356350-1000ML | |
| LB broth, Miller | Millipore Sigma | 60801-450 | |
| Sodium dodecyl sulfate (SDS? | Sigma | L4509-250G | |
| Sodium chloride | Sigma | 59888 | |
| M880 Ultrasonic bath, 117 V, holds 5.5 gallons | VWR, USA | 89375-458 | |
| Magnesium sulphate | Sigma | M506 | |
| Magnesium chloride | Sigma | 208337 | |
| NGM agar plate | VWR Disposable Petri Dishes | 25384-342 | |
| NuPAGE LDS Sample Buffer (4X) | Thermo Fisher Scientific | NP0007 | |
| NuPAGE protein gels, 4-12% | Invitrogen | NP 0335BOX | |
| Protease inhibiotr cocktail (PIC) | Roche | 11836170001 | |
| Pierce BCA Assay | Thermo Fisher Scientific | 23225 | |
| Sodium hypochlorite 5% | VWR | JT9416-1 | |
| SYPRO Ruby Protein Gel Stain | Thermo Fisher Scientific | S12000 | |
| MS Section | |||
| Acetonitrile, Burdick and Jackson LC-MS | Honeywell International Inc., Charlotte, NC, USA | 36XL66 | |
| Agilent Zorbax 300Extend C18 column | Agilent Technologies Inc., Santa Clara, CA, USA | 770995-902 | |
| Ammonium bicarbonate | Sigma Aldrich, St. Louis, MO, USA | 9830 (1 kg) | |
| Dithiothreitol (DTT) | Sigma Aldrich, St. Louis, MO, USA | D9779-5G | |
| Eppendorf Thermomixer Compact | Eppendorf AG, Hamburg, Germany | T1317-1EA | |
| Formic acid | Sigma Aldrich, St. Louis, MO, USA | F0507-500ML | |
| Indexed retention time (iRT) normalization peptide standard | Biognosys AG, Schlieren, Zurich, Switzerland | Ki-3002-2 | |
| Iodoacetamide (IAA) | Sigma Aldrich, St. Louis, MO, USA | I1149-25G | |
| Methanol, HPLC Grade | Honeywell International Inc., Charlotte, NC, USA | 34885 | |
| Nano cHiPLC Trap ChromXP C18-CL, 200 um x 6 mm, 3 um, 120A. (pre-column chip) (200 um x 6 mm ChromXP C18-CL chip, 3 um, 300 A) | Sciex LLC, Framingham, MA, USA | 804-00006 | |
| Nano cHiPLC ChromXP 75 um by 15cm, C18-CL, 3 um, 120 A (analytical column chip) | Sciex LLC, Framingham, MA, USA | 804-00001 | |
| Orthoganol quadrupole time-of-flight (QqTOF) TripleTOP 6600 mass spectrometer | Sciex LLC, Framingham, MA, USA | Per quote | |
| ProteinPilot 5.0 | Sciex LLC, Framingham, MA, USA | software download Sciex | |
| Savant SPD131DDA Speedvac Concentrator | Thermo Fisher Scientific, Waltham, MA, USA | SPD131DDA-115 | |
| Sequencing-grade lyophilized trypsin | Life Technologies | 23225 | |
| Spectronaut | Biognosys AG, Schlieren, Zurich, Switzerland | Sw-3001 | |
| SWATH 2.0 plugin into PeakView 2.2 | Sciex LLC, Framingham, MA, USA | software download Sciex | |
| Ultra Plus nano-LC 2D HPLC system | Sciex LLC, Eksigent Division, Framingham, MA, USA | Model # 845 | |
| Water, Burdick and Jackson LC-MS | Honeywell International Inc., Charlotte, NC, USA | 600-30-76 | |
| Waters 1525 binary HPLC pump system | Waters Corp., Milford, MA, USA | WAT022939 | |
| Waters 2487 Dual Wavelength UV detector | Waters Corp., Milford, MA, USA | WAT081110 | |
| Waters 717plus Autosampler | Waters Corp., Milford, MA, USA | WAT022939 | |
| Waters Fraction Collector III | Waters Corp., Milford, MA, USA | 186001878 |
参考文献
- Walther, D. M., et al. Widespread Proteome Remodeling and Aggregation in Aging C. elegans. Cell. , 919-932 (2015).
- David, D. C., et al. Widespread protein aggregation as an inherent part of aging in C. elegans. PLoS Biology. 8 (8), 47-48 (2010).
- Labbadia, J., Morimoto, R. I. The Biology of Proteostasis in Aging and Disease. Annual Review of Biochemistry. (1), 435-464 (2013).
- Ross, C. A., Poirier, M. A. Protein aggregation and neurodegenerative disease. Nature Medicine. , 10-17 (2004).
- Reis-Rodrigues, P., et al. Proteomic analysis of age-dependent changes in protein solubility identifies genes that modulate lifespan. Aging Cell. 11 (1), 120-127 (2012).
- Ayyadevara, S., et al. Proteins that accumulate with age in human skeletal-muscle aggregates contribute to declines in muscle mass and function in Caenorhabditis elegans. Aging. 8 (12), 3486-3497 (2016).
- Huang, C., et al. Intrinsically aggregation-prone proteins form amyloid-like aggregates and contribute to tissue aging in Caenorhabditis elegans. eLife. 8, (2019).
- Morimoto, R. I. Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging. Genes and Development. 22 (11), 1427-1438 (2008).
- Morimoto, R. I., Cuervo, A. M. Protein homeostasis and aging: Taking care of proteins from the cradle to the grave. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 64 (2), 167-170 (2009).
- Klang, I. M., et al. Iron promotes protein insolubility and aging in C. elegans. Aging. 6 (11), 975-991 (2014).
- Mark, K. A., et al. Vitamin D Promotes Protein Homeostasis and Longevity via the Stress Response Pathway Genes skn-1, ire-1, and xbp-1. Cell Reports. 17 (5), 1227-1237 (2016).
- Groh, N., et al. Methods to study changes in inherent protein aggregation with age in caenorhabditis elegans. Journal of Visualized Experiments. (129), 1-12 (2017).
- Schilling, B., Gibson, B. W., Hunter, C. L. Generation of High-Quality SWATH((R)) Acquisition Data for Label-free Quantitative Proteomics Studies Using TripleTOF((R)) Mass Spectrometers. Proteomics: Methods and Protocols, Methods in Molecular Biology. 1550, 223-233 (2017).
- Walther, D. M., et al. Widespread proteome remodeling and aggregation in aging C. elegans. Cell. 161 (4), 919-932 (2015).
- Angeli, S., et al. A DNA synthesis inhibitor is protective against proteotoxic stressors via modulation of fertility pathways in Caenorhabditis elegans. Aging. 5 (10), 759-769 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。