
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
大小排除色谱分析细菌外膜囊性异质性
摘要
细菌囊泡在发病机制中起着重要作用,具有广阔的生物技术应用前景。囊泡的异质性使分析和使用复杂化:因此,需要一种简单、可重复的方法来分离不同大小的囊泡。在这里,我们演示了使用大小排除色谱来分离由 聚合菌作用肌瘤产生的异质囊泡。
摘要
革兰氏阴性细菌的细胞壁由内(细胞质)和外膜(OM)组成,由薄多肽糖层分离。在整个生长过程中,外膜可以形成球形外膜囊泡(OMVs)。这些 OMV 涉及许多细胞功能,包括货物运送到宿主细胞和与细菌细胞的通信。最近,开始探讨OMV的治疗潜力,包括将其用作疫苗和药物输送工具。虽然OMV来自OM,但长期以来人们一直认为OMV的脂质和蛋白质货物与OM的脂质和蛋白质货物有显著差异。最近,已经发现了细菌可以释放多种类型的OMV的证据,并且有证据表明,体型可以影响宿主细胞吸收它们的机制。然而,这一领域的研究受到有效分离异构大小的OMV的困难的限制。密度梯度离心(DGC)传统上用于此目的:然而,这种技术是耗时和难以扩大。尺寸排除色谱 (SEC), 另一方面, 是不太繁琐, 并适合必要的未来扩大治疗使用 OMV.在这里,我们描述了一种SEC方法,它允许可重复分离异质大小的囊泡,用作测试案例,由 聚合菌作用性乙酰氨基甲基苯丙氨酸生产的OMV,其直径从小于150纳米到大于350纳米不等。我们演示了"大"(350 nm)OMV 和"小"(< 150 nm) OMV 的分离,通过动态光散射 (DLS) 验证。我们建议基于 SEC 的技术,而不是基于 DGC 的技术,以分离异构大小的囊泡,因为它易于使用、可重复性(包括用户对用户)和扩大规模的可能性。
引言
革兰阴性细菌释放囊泡从他们的外膜,所谓的外膜囊泡(OMVs),在整个生长。这些OMV通过携带许多重要的生物分子,包括DNA/RNA、蛋白质、脂质和肽类药物1、2,在细胞与宿主之间以及细菌细胞之间的细胞间交流中发挥着重要作用。特别是,由于OMV在某些毒因子和毒素3、4、5、6、7、8、9、10、11中富集,因此在细菌发病机制中的作用得到了广泛的研究。
据报道,OMV的体积从20到450纳米不等,取决于母体细菌和生长阶段,几种类型的细菌释放异质大小的OMV 8,12,13,14,这也不同于其蛋白质成分和宿主细胞进入12的机制。H. 皮洛里释放的OMV直径从20到450纳米不等,较小的OMV含有比较大的OMV更均匀的蛋白质成分。重要的是,观察到两个OMV种群通过不同的机制12被宿主细胞内化。此外,我们已经证明,聚合菌作用性肌瘤释放了小(<150nm)OMV种群以及大量(>350纳米)OMV,其中OMV含有大量分泌的蛋白质毒素,白细胞毒素(LtxA)15。虽然OMV异质性在细胞过程中的作用显然很重要,但在分离和分析不同囊泡种群的技术困难限制了这些研究。
除了在细菌发病机制中的重要性外,还建议将OMV用于一些生物技术应用,包括作为疫苗和药物输送工具16、17、18、19、20。对于这些方法的转化用途,需要清洁和单分散的囊泡制备。因此,必须采用有效和有效的分离方法。
最常见的是,密度梯度离心(DGC)用于将异质大小的囊泡体群与细胞碎片分离,包括旗菌和分泌蛋白质21:该方法也已报告为分离异构大小的OMV子聚合12,13,14的方法。但是,DGC 耗时、效率低下且用户对用户22的变量也很高,因此不适合扩展。相比之下,尺寸排除色谱 (SEC) 代表一种可扩展、高效和一致的方法来净化 OMV 21、23、24。我们发现,一个长(50厘米),重力流动,SEC列,充满了凝胶过滤介质足以有效地净化和分离OMV的亚群。具体来说,我们用这种方法将A.行动肌共体OMV分离成"大"和"小"子聚类,并去除蛋白质和DNA污染。净化工作在不到4小时的时间内完成,完成了OMV子群的完全分离和碎片的清除。
研究方案
1. 准备缓冲器
- 要准备 ELISA 洗涤缓冲器,请添加 3.94 g Tris 基、8.77 g NaCl 和 1 克牛血清白蛋白 (BSA) 到 1 L 的去离子 (DI) 水。添加 500 μL 多山梨酸盐-20。使用 Hcl 或 Naoh 将 ph 调整为 7.2。
- 要准备阻塞缓冲器,添加 3.94 g Tris 基、8.77 g NaCl 和 10 g BSA。加入 500 μL 多山梨酸酯-20 至 1 升 DI 水。使用 Hcl 或 Naoh 将 ph 调整为 7.2。
- 要准备洗礼缓冲器 (PBS),添加 8.01 g NaCl、2.7 g KCl、1.42 g Na2HPO4和 0.24 g KH2PO4 至 1 L DI 水。使用 Hcl 或 Naoh 将 ph 调整为 7.4。
注:此缓冲器的 10 倍解决方案可根据需要用 DI 水制成并稀释。
2. 准备OMV样品
- 将 A.行动肌细胞 生长到指数末期(光学密度为600纳米0.7)。在 4 °C 下以 10,000 x g 离心两次,在 10 分钟内将细胞颗粒化。通过 0.45 μm 过滤器过滤超高纳特。
- 使用 50 kDa 分子重量切断过滤器浓缩无细菌超高分子。超中心在 4 °C 下以 105,000 x g 的浓缩溶液进行 30 分钟。
- 再次在 PBS 和超中心再次重复颗粒(105,000 x g 在 4 °C 下 30 分钟)在 PBS 的 2 mL 中重新使用颗粒。
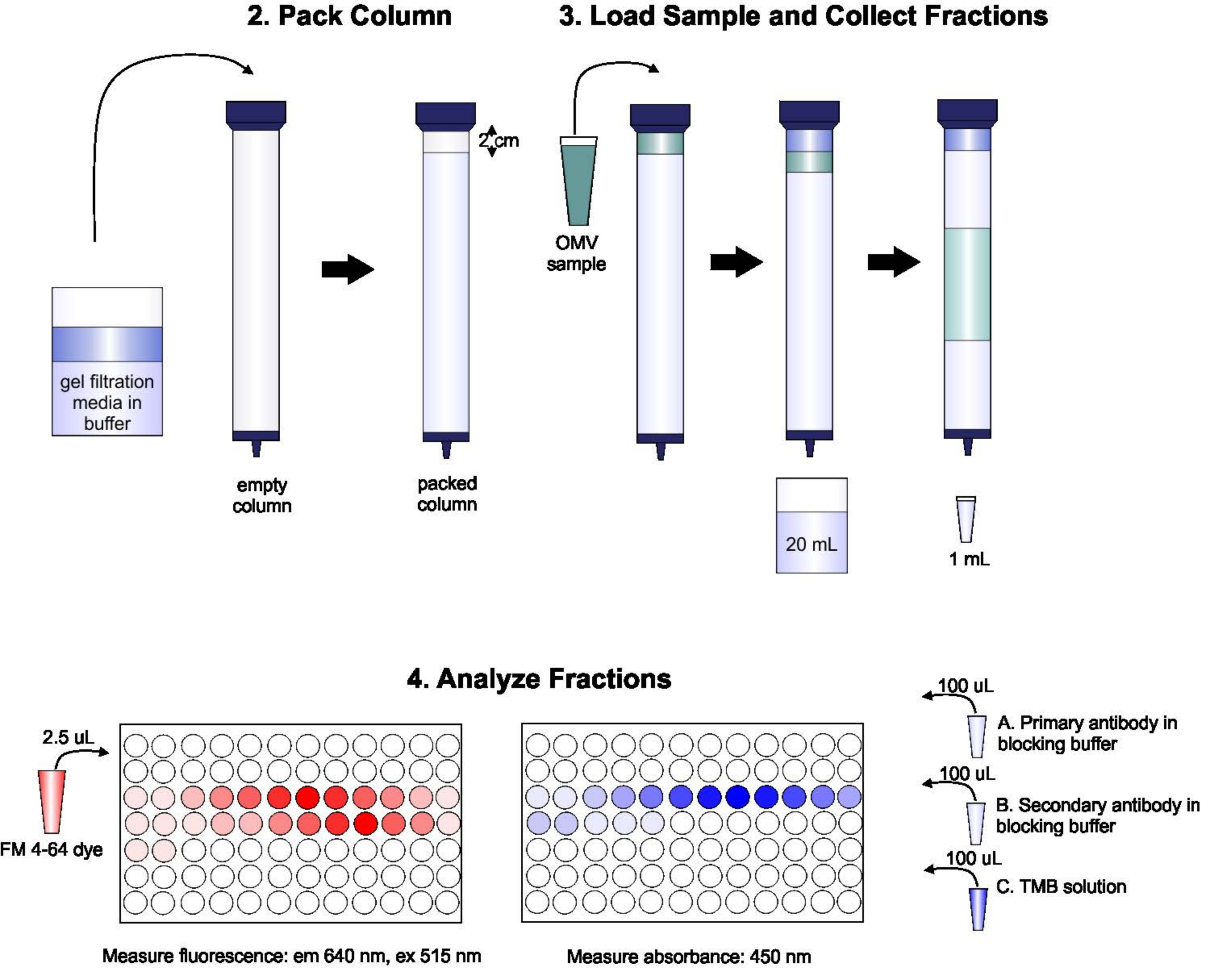
3. 包装 S-1000 列
- 将库存瓶凝胶过滤介质与玻璃搅拌棒混合,倒入玻璃瓶中,这是填充柱子所需的体积,外加约 50% 的超量(约 135 毫升)。让这些珠子坐到它们安顿下来,然后从多余的液体中排出。将珠子重新插入弹性缓冲,以便最终解决方案是大约 70%(按体积)凝胶,30% 缓冲。在真空下解加溶液。
- 使用环形支架垂直安装玻璃柱,并填充弹性缓冲器,以弄湿柱壁。排空缓冲器,直到柱子中仅剩约 1 厘米的缓冲。
- 在不产生气泡的情况下,小心地将移液器珠子放入柱子中,将柱子填充到顶部。在整个过程中继续排出多余的缓冲。在将额外的珠子添加到柱子顶部之前,请务必不要让珠子完全沉淀。柱子应包装到柱子储层底部下方约 2 厘米的高度。
4. 加载样品并收集分数
- 在真空下解脱弹性缓冲器。用两列卷(180 mL)的洗涤电位缓冲器。
- 允许剩余的缓冲区完全进入列。一旦缓冲器到达凝胶层的顶部,小心地将含有OMV(脂质浓度约为100-200 nmol/L)的2ml样品移液到珠子表面,小心不要打扰柱子顶部的任何珠子。允许样品完全进入凝胶,即当凝胶层上方没有液体时。
- 小心而缓慢地在凝胶柱顶部添加弹性缓冲。请勿打扰凝胶的顶层,因为这会导致样品稀释。
- 将单个 50 mL 管放在柱子下并打开柱子。收集前 20 mL 的 eluent。根据需要,在柱子顶部添加额外的洗礼缓冲器,以确保柱子永远不会干燥。
- 将一系列 1.5 mL 管放在柱子下。启动柱子,并在每个管中收集一系列 1 mL 样本。在收集样品时,如有必要,继续在柱子顶部添加弹性缓冲器。重复,直到收集了 96 个分数。停止列。
注:样品应储存在 -20 °C 用于长期存储,或 4 °C 用于短期存储,直到进一步分析。 - 要清洁柱,通过列运行 0.1 M NaOH 的一列体积(90 mL)。通过列运行两列卷 (180 mL) 的弹性缓冲。
5. 样品分析
- 为了测量每个分数中的脂质浓度,每个分数的移液器 50 μL 放入 96 井板的单井中。在每口井中,加入2.5微升的嗜脂染料。孵育15s。测量激发波长为 515 nm 和发射波长为 640 nm 的板读卡器上的荧光强度。要计算每个样品中所有脂质的百分比,则汇总所有排放强度,并将每个单个强度除以总强度。
- 为了测量特定蛋白质的浓度,每个分数的移液器 100 μL 进入 ELISA 免疫板的单井中。在 25 °C 下孵育 3 小时。
- 去特样品。在每口井中加入 200 μL 的 ELISA 洗涤缓冲器,然后排出。重复四次,总共洗五次。
- 在每个井中加入 200 μL 的阻隔缓冲器,并在 25 °C 下孵育 1 小时。 滗。
- 在 4 °C 下一夜之间用 100 μL 阻塞缓冲器和原发性抗体(1:10,000 表示纯化抗体;1:10 用于未净化抗体)孵育板。 滗。
- 在每口井中加入 200 μL 的 ELISA 洗涤缓冲器,然后排出。重复四次,总共洗五次。
- 在每口井中加入 100 μL 的 ELISA 洗涤缓冲器和辅助抗体(1:30,000)。在 25 °C 下孵育 1 小时。
- 在每口井中加入 200 μL 的 ELISA 洗涤缓冲器,然后排出。重复四次,总共洗五次。
- 加入 100 μL 的 3,3',5,5'-四甲基苯齐丁 (TMB) 单步解决方案,孵育 15-30 分钟或直到蓝色发展。停止 TMB 反应与 50 μL 的停止解决方案。
- 在板读卡器上,阅读每口井的吸血度,波长为 450 nm。
- 要测量蛋白质总浓度,使用紫外线光谱仪,以每个分数的波长 280 nm (A280)记录吸收量。
协议的示意图显示在 图1中。

图1:SEC程序的示意图。 柱子中小心地装着脱气凝胶过滤介质,以避免气泡和不连续性,然后用两列排的洗涤缓冲器清洗。接下来,将样品小心地输送到凝胶顶部,而不会打乱凝胶包装。打开柱子并运行,直到样品完全进入凝胶。此时,缓冲器放置在柱子顶部,并收集前 20 mL 的 eluate。接下来,收集一系列 1mL 分数。然后将这些分数放置在 96 井板或 96 井免疫板中,用于分析脂质和蛋白质含量。 请单击此处查看此图的较大版本。
{kind=link}
结果
图2显示了此方法的代表性结果。 由A.行动肌共 性菌株JP2生产的OMV首先从培养超高技术中纯化使用超浓缩15。我们以前发现,这种菌株产生两个OMV种群,一个直径约300纳米,一个直径约100纳米15。为了分离这些OMV种群,我们使用上述 SEC 协议对样本进行了净化。使用脂质染料分析每个分数的脂质含量和使用酶链接的免疫吸附剂检测 (E...
讨论
在这里,我们为细菌OMV亚聚合的简单、快速和可重复分离提供了一个协议。虽然该技术相对直截了当,但必须非常仔细地执行某些步骤,以确保在列中发生高效分离。首先,必须小心而缓慢地将凝胶加载到柱子中,以避免气泡。我们观察到,在加载柱之前,将凝胶在室温下保留数小时,使凝胶能够平衡并最大限度地减少柱内的气泡形成。当凝胶被管道插入柱子时,应沿着柱子的侧面小心地管道,?...
披露声明
作者没有利益冲突要报告。
致谢
这项工作由国家科学基金会(1554417)和国家卫生研究院(DE027769)资助。
材料
| Name | Company | Catalog Number | Comments |
| 1-Step Ultra TMB-ELISA | Thermo Scientific | 34028 | |
| Amicon 50 kDa filters | Millipore Sigma | UFC905024 | |
| Bovine Serum Albumin (BSA) | Fisher Scientific | BP9704-100 | |
| ELISA Immuno Plates | Thermo Scientific | 442404 | |
| FM 4-64 | Thermo Scientific | T13320 | 1.5 x 50 cm |
| Glass Econo-Column | BioRad | 7371552 | |
| Infinite 200 Pro Plate Reader | Tecan | ||
| Potassium Chloride (KCl) | Amresco (VWR) | 0395-500G | |
| Potassium Phosphate Monobasic Anhydrous (KH2PO4) | Amresco (VWR) | 0781-500G | |
| Sephacryl S-1000 Superfine | GE Healthcare | 17-0476-01 | |
| Sodium Chloride (NaCl) | Fisher Chemical | S271-3 | |
| Sodium Phosphate Dibasic Anhydrous (Na2HPO4) | Amresco (VWR) | 0404-500G | |
| Tris Base | VWR | 0497-1KG | |
| Tween(R) 20 | Acros Organics | 23336-2500 |
参考文献
- Kuehn, M. J., Kesty, N. C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes and Development. 19, 2645-2655 (2005).
- Kulp, A., Kuehn, M. J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annual Reviews Microbiology. 64, 163-184 (2010).
- Kato, S., Kowashi, Y., Demuth, D. R. Outer membrane-like vesicles secreted by Actinobacillus actinomycetemcomitans are enriched in leukotoxin. Microbial Pathogenesis. 32 (1), 1-13 (2002).
- Nice, J. B., et al. Aggregatibacter actinomycetemcomitans leukotoxin is delivered to host cells in an LFA-1-independent manner when associated with outer membrane vesicles. Toxins. 10 (10), 414 (2018).
- Haurat, M. F., et al. Selective sorting of cargo proteins into bacterial membrane vesicles. Journal of Biological Chemistry. 286 (2), 1269-1276 (2011).
- Horstman, A. L., Kuehn, M. J. Enterotoxigenic Escherichia coli secretes active heat-labile enterotoxin via outer membrane vesicles. The Journal of Biological Chemistry. 275 (17), 12489-12496 (2000).
- Wai, S. N., et al. Vesicle-mediated export and assembly of pore-forming oligomers of the Enterobacterial ClyA cytotoxin. Cell. 115, 25-35 (2003).
- Balsalobre, C., et al. Release of the Type I secreted α-haemolysin via outer membrane vesicles from Escherichia coli. Molecular Microbiology. 59 (1), 99-112 (2006).
- Donato, G. M., et al. Delivery of Bordetella pertussis adenylate cyclase toxin to target cells via outer membrane vesicles. FEBS Letters. 586, 459-465 (2012).
- Kim, Y. R., et al. Outer membrane vesicles of Vibrio vulnificus deliver cytolysin-hemolysin VvhA into epithelial cells to induce cytotoxicity. Biochemical and Biophysical Research Communications. 399, 607-612 (2010).
- Maldonado, R., Wei, R., Kachlany, S. C., Kazi, M., Balashova, N. V. Cytotoxic effects of Kingella kingae outer membrane vesicles on human cells. Microbial Pathogenesis. 51 (1-2), 22-30 (2011).
- Turner, L., et al. Helicobacter pylori outer membrane vesicle size determines their mechanisms of host cell entry and protein content. Frontiers in Immunology. 9, 1466 (2018).
- Zavan, L., Bitto, N. J., Johnston, E. L., Greening, D. W., Kaparakis-Liaskos, M. Helicobacter pylori growth stage determines the size, protein composition, and preferential cargo packaging of outer membrane vesicles. Proteomics. 19 (1-2), 1800209 (2019).
- Rompikuntal, P. K., et al. Perinuclear localization of internalized outer membrane vesicles carrying active cytolethal distending toxin from Aggregatibacter actinomycetemcomitans. Infections and Immunity. 80 (1), 31-42 (2012).
- Nice, J. B., et al. Aggregatibacter actinomycetemcomitans leukotoxin is delivered to host cells in an LFA-1-independent manner when associated with outer membrane vesicles. Toxins. 10 (10), 414 (2018).
- Stevenson, T. C., et al. Immunization with outer membrane vesicles displaying conserved surface polysaccharide antigen elicits broadly antimicrobial antibodies. Proceedings of the National Academy of Sciences. 115 (14), 3106-3115 (2018).
- Gujrati, V., et al. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano. 8 (2), 1525-1537 (2014).
- Huang, W., et al. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. Journal of Controlled Release. 317, 1-22 (2020).
- Chen, D. J., et al. Delivery of foreign antigens by engineered outer membrane vesicle vaccines. Proceedings of the National Academy of Sciences. 107 (7), 3099-3104 (2010).
- Chen, L., et al. Outer membrane vesicles displaying engineered glycotopes elicit protective antibodies. Proceedings of the National Academy of Sciences. 113 (26), 3609-3618 (2016).
- Singorenko, P. D., et al. Isolation of membrane vesicles from prokaryotes: A technical and biological comparison reveals heterogeneity. Journal of Extracellular Vesicles. 6 (1), 1324731 (2017).
- Zeringer, E., Barta, T., Li, M., Vlassov, A. V. Strategies for isolation of exosomes. Cold Spring Harbor Protocols. 2015 (4), 319-323 (2015).
- Benedikter, B. J., et al. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Science Reports. 7 (1), 15297 (2017).
- Mol, E. A., Goumans, M. J., Doevendans, P. A., Sluijter, J. P. G., Vader, P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomedicine. 13 (6), 2061-2065 (2017).
- Lally, E. T., Golub, E. E., Kieba, I. R. Identification and immunological characterization of the domain of Actinobacillus actinomycetemcomitans leukotoxin that determines its specificity for human target cells. Journal of Biological Chemistry. 269 (49), 31289-31295 (1994).
- Chang, E. H., Giaquinto, P., Huang, J., Balashova, N. V., Brown, A. C. Epigallocatechin gallate inhibits leukotoxin release by Aggregatibacter actinomycetemcomitans by promoting association with the bacterial membrane. Molecular Oral Microbiology. 35 (1), 29-39 (2020).
- Klimentová, J., Stulík, J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiological Research. 170, 1-9 (2015).
- Dauros Singorenko, P., et al. Isolation of membrane vesicles from prokaryotes: a technical and biological comparison reveals heterogeneity. Journal of Extracellular Vesicles. 6 (1), 1324731 (2017).
- Monguió-Tortajada, M., Gálvez-Montón, C., Bayes-Genis, A., Roura, S., Borràs, F. E. Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cellular and Molecular Life Sciences. 76 (12), 2369-2382 (2019).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。