
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
在斑马鱼胃中使用双光子显微镜进行深度和空间控制的体积消融
摘要
胚胎发育需要细胞运动的大规模协调。双光子激发介导的激光烧蚀允许对大群深层细胞进行空间控制的三维烧蚀。此外,该技术可以探测 体内 集体迁移细胞对其机械环境中的扰动的反应。
摘要
形态发生涉及许多细胞运动,以将细胞组织成组织和器官。为了适当的发展,所有这些运动都需要紧密协调,积累的证据表明,这至少部分是通过机械相互作用实现的。在胚胎中测试这一点需要直接的身体扰动。激光烧蚀是一种日益常用的选择,可以缓解机械约束或物理隔离两个细胞群。然而,许多烧蚀都是用紫外线(UV)激光进行的,其提供的轴向分辨率和组织穿透有限。这里描述了一种使用双光子显微镜消融深度,显着和空间上明确定义的体积的方法。消融在表达轴向中端肠胚层中绿色荧光蛋白的转基因斑马鱼品系中表现出来,并用于切断轴向中胚层,而不影响上覆的外胚层或下面的卵黄细胞。在消融之前和之后通过实时成像监测细胞行为。消融方案可用于不同的发育阶段,任何细胞类型或组织,规模从几微米到超过一百微米不等。
引言
细胞 - 细胞相互作用在发育中起着至关重要的作用。细胞提供信号,它们的直接邻居或更远的细胞可以感知,从而影响它们的命运和/或行为。其中许多信号本质上是化学性质的。例如,在表征良好的诱导事件中,一个细胞群产生可扩散的分子,影响另一个细胞群的命运1。然而,其他信号是机械的。细胞对邻居施加力和约束,邻居感知并做出反应2。
研究这些细胞 - 细胞 相互作用在体内 的重要性的一种方法是消除一些细胞并观察随后的发展。不幸的是,去除或破坏细胞的可用技术是有限的。细胞可以通过手术切除3,4,使用针头或小线,但这种治疗是侵入性的,不是很精确,通常在立体显微镜下进行,阻止在显微镜下立即成像。此外,靶向深层细胞意味着在覆盖的组织中刺穿一个洞,从而产生不必要的扰动。基因编码的光敏剂,如KillerRed,已被用于通过光照诱导细胞死亡5。光敏剂是在光照射下产生活性氧的发色团。它们的主要局限性是它们需要长光照明(约15分钟),如果细胞移动,这可能很难实现,并且它们通过细胞凋亡诱导细胞死亡,这不是立即的。
最后,激光烧蚀在过去15年中得到了发展和广泛的应用6,7,8,9,10,11,12。激光束聚焦在目标细胞/组织上。它通过加热,光消融或血浆诱导的消融诱导其消融;所涉及的过程取决于功率密度和曝光时间13。大多数烧蚀方案使用紫外激光器来实现其高能量。然而,紫外线既被生物组织吸收又被散射。因此,靶向深层细胞需要高激光功率,然后在更浅表的,平面外的组织中诱导损伤。这限制了紫外激光器在浅表结构上的使用,并解释了它们相对较低的轴向分辨率。非线性光学(所谓的双光子显微镜)使用光的非线性特性来激发荧光团,荧光团在红外域中具有大约一半能量的两个光子。当应用于消融术时,这有三个主要优点。首先,与紫外光相比,红外光的散射更少,被生物组织吸收更少14,从而可以在不增加所需激光功率的情况下到达更深的结构。其次,使用飞秒脉冲激光器可提供非常高的功率密度,通过等离子体感应产生烧蚀,这与加热相反,不会在空间上扩散15。第三,诱导等离子体形成的功率密度仅在焦点处达到。由于这些特性,双光子激光烧蚀可用于精确靶向深层细胞,而不会影响周围的组织环境。
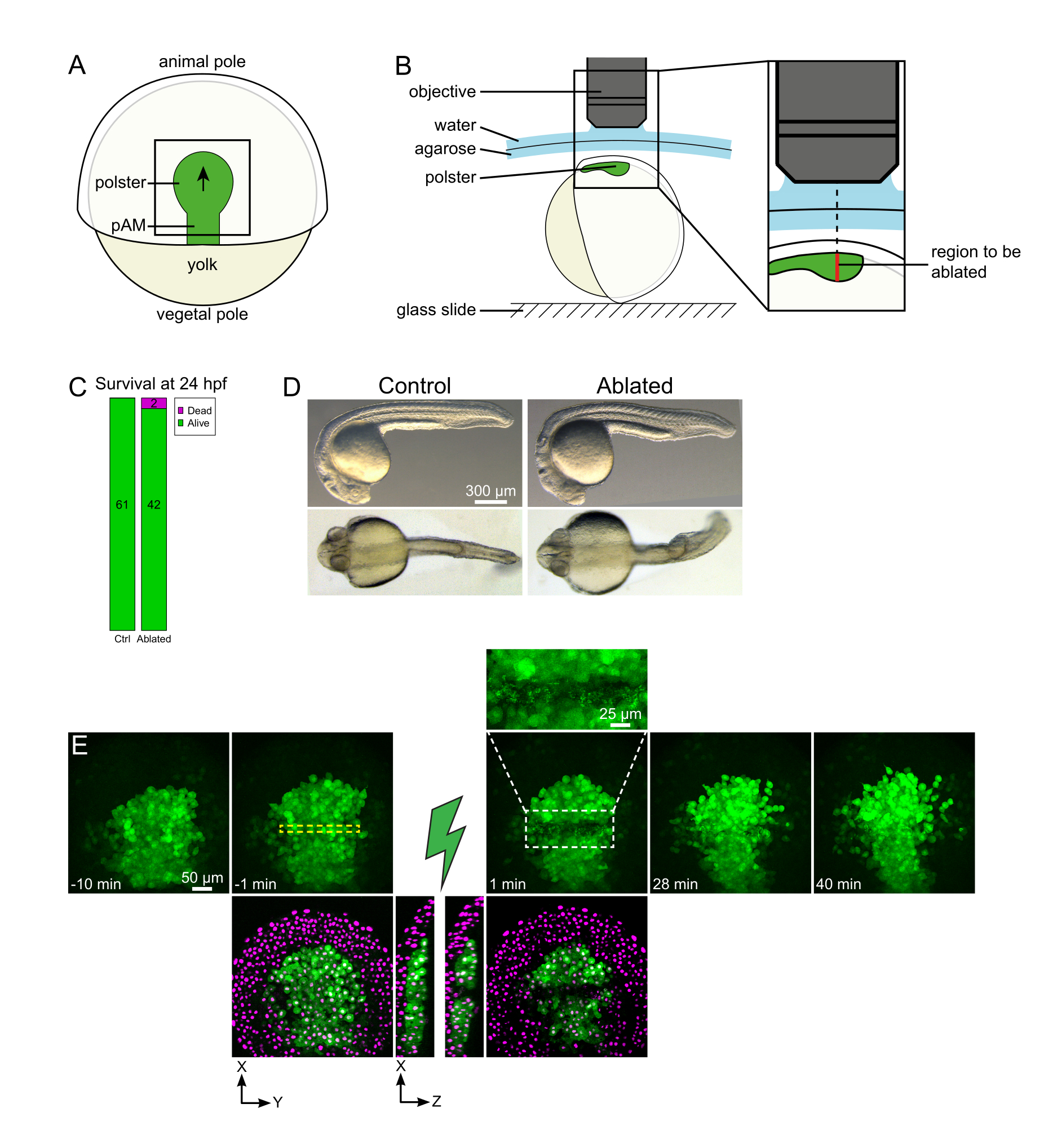
集体迁移是发育过程的一个很好的例子,其中细胞 - 细胞相互作用是根本的。集体迁移被定义为相邻细胞影响一个细胞行为的细胞迁移16。这些相互作用的性质(化学或机械)以及它们如何影响细胞迁移可能会有很大差异,并且通常不完全理解。移除细胞并观察其如何影响其他细胞的能力对于进一步解开这些集体过程至关重要。几年前,我们使用手术方法确定,斑马鱼原肠胚形成过程中的洄游是集体迁徙17。Polster是一组细胞,构成胚胎背侧的第一个内化细胞18。这些细胞在 Tg(gsc:GFP) 转基因系中以绿色标记,位于胚胎深处,在几层外胚层细胞的下方。在原肠胚形成过程中,该组导致轴向中胚层的延伸,从胚胎组织者迁移到动物极19,20,21,22,23 (图1A)。我们确定细胞需要与邻居接触,以将其迁徙定位在动物极点的方向上。然而,更好地了解这种集体迁移的细胞和分子基础涉及去除一些细胞,看看这如何影响其余的细胞。因此,我们使用双光子显微镜设置开发了大而深体积的烧蚀。在这里,我们演示了使用该协议来切断中间的polster,并通过跟踪用Histone2B-mCherry标记的细胞核来观察对细胞迁移的影响。
研究方案
所有动物工作均由伦理委员会N 59和国家教育部批准,文件编号为APAFIS#15859-2018051710341011v3。下面描述的一些步骤特定于我们的设备和软件,但可以很容易地适应不同的设备。
1. 注射准备
- 在胚胎培养基(EM)中制备75毫升1%琼脂糖溶液。
- 将注射模具放入90毫米培养皿中,倒入约50毫升琼脂糖,足以让模具漂浮。让琼脂糖凝固并除去注射模具。
- 通过将1 mL琼脂糖倒入30毫米培养皿中来制备琼脂糖包衣的培养皿。
- 将储备溶液稀释在不含RNase的水中,制备4μL 30ng / μL组蛋白2B-mCherry mRNA溶液并保持在冰上。
注意:在操作mRNA时要小心戴上手套,以避免RNase介导的降解。 - 使用微量移液管拉拔器从毛细管中拉出注射针。
2. 胚胎制备
- 一旦鱼产卵,在EM的90毫米培养皿中收集,冲洗和收获。将胚胎置于28.5°C培养箱中。
- 等待 20 分钟,让第一个单元格变为可见。
- 将30个胚胎转移到充满EM的注射板中。使用略钝的镊子将胚胎挤压在凹槽中,并用动物杆向上定向。
- 使用微量加载器尖端,用2μLmRNA溶液填充注射针。将针头插入毛细管支架中,置于与聚四氟乙烯(PTFE)管连接到空气喷射器的微型机械手中。
- 在立体显微镜下,小心地打破针尖。
- 通过将针头插入细胞中,将mRNA溶液注射到1细胞期胚胎中。
注意:注射的体积约为细胞体积的三分之一。 - 将注射的胚胎放回28.5°C培养箱中。
3. 双光子显微镜的制备
注:本协议使用两个激光器。一个用于成像GFP(在920nm处)并进行烧蚀(在820nm处)。它将被称为绿光/烧蚀激光。另一个用于在1160nm处对mCherry进行成像。它将被称为红色激光。
- 将绿色/烧蚀激光设置为 820 nm(烧蚀波长),将红色激光设置为 1160 nm(mCherry 激发)。
- 在光路上使用可移动的反射镜,在扫描头的入口和出口处对齐绿色/烧蚀和红色激光束。
注:这增加了激光束焦点,并最大限度地减少了激发和烧蚀的焦散量。 - 测量物镜下820nm处绿光/烧蚀激光器的最大功率。为此,请将功率计放在物镜下方,关闭黑色腔室,将绿色/烧蚀激光功率设置为100%,然后打开百叶窗。计算达到 300 mW 所需的激光功率百分比。
- 将绿光/烧蚀激光器回设为920 nm(GFP激发),并将激光功率设置为7%。将红色激光功率设置为 15%。
- 激活外延光电倍增管(PMT)探测器,用于绿线和红线;将绿线和红线 PMT 灵敏度设置为 65。
- 将视场设置为 400 x 400 μm,将图像分辨率设置为 512 x 512 像素,将扫描频率设置为 800 Hz。
- 选择 "3D 延时成像" 模式。然后,创建一个文件夹,并在每次采集后激活 自动保存 数据。
- 组装加热室并将其设置为28°C。 等待至少10分钟,使腔室和物镜变暖。
4. 安装胚胎
- 在荧光立体显微镜下,以70%的外延率鉴定表达GFP的胚胎。
注意:选择轴向中胚层中具有明亮信号且无背景荧光的胚胎,以获得更好的成像质量。 - 使用塑料巴斯德移液管在琼脂糖涂层皿(步骤1.3)中转移三到四个选定的胚胎,并使用细镊子小心地将其去脉。
注意:去鞘胚胎非常脆弱,与空气或塑料接触时会破裂。 - 将 1 mL 0.2% 琼脂糖倒入 1x 青霉素-链霉素 EM 中,放入一个小玻璃瓶中。将小瓶置于预热的42°C干块加热器中。
注意:必须快速执行以下步骤,以便在琼脂糖凝固之前允许胚胎定向。 - 使用火抛光玻璃移液器将去壳的胚胎转移到0.2%琼脂糖玻璃小瓶中。注意不要在琼脂糖中添加过多的EM,以避免稀释它。从移液器中丢弃剩余的EM,并在胚胎从移液器中脱落之前,将胚胎与足够的琼脂糖一起吸回以覆盖玻璃底培养皿的载玻片。
- 将琼脂糖和胚胎吹在盘子的载玻片上。注意不要让胚胎接触空气或盘子的塑料面。接下来,用琼脂糖填充载玻片周围的腔室。
- 使用睫毛定向胚胎,使目标区域位于顶部(图1B)。
注意:在定向胚胎时,请注意仅触摸胚泡,而不是非常脆弱的蛋黄。琼脂糖将在1分钟左右凝固,具体取决于室温。 - 等待约5分钟,琼脂糖完全凝固,然后加入几滴青霉素 - 链霉素EM。
5. 定位胚胎和消融前成像
- 将玻璃底盘放在加热室中物镜下方。将物镜浸入青霉素 - 链霉素EM中并关闭加热室。
- 移动滑块以设置目镜的光路。然后,使用目镜,荧光灯和载物台控制,找到一个胚胎并将焦点设置为胚胎表面。
- 关闭荧光灯,将光路设置为PMT,然后关闭黑色腔室。
注:小心关闭黑色腔室中的所有光源,因为它可能会损坏PMT。 - 开始实时成像并定位轴向中胚层。调整绿色/烧蚀和红色激光功率以获得良好的信号(即,对于GFP表示区域,每像素1,000至20,000光子)。使用红色通道将阶段移动到胚胎的最顶部,并将此位置设置为Z = 0。
- 选择 1 分钟的时间步长和 2 μm 的 Z 步长。110 μm的Z形程足以覆盖整个埾星,并且在这些设置下不到1分钟即可获得。将第一片片设置在轴向中胚层上方15μm处(在更浅的外胚层中)。
注:堇子沿曲线移动,因此Z层的底部应设置为比堇子最深位置深30μm,以适应其在延时成像期间的运动(图1E)。 - 录制10-15分钟的消融前电影。

图1:激光消融术的成功结果(A)背侧70%外延的胃泌尿胚胎方案;pAM:后轴中胚层;黑色箭头标记了樁匐迁移的方向;黑色方块表示极点中烧蚀的典型视场。(B)用于切断的胚胎安装方案。侧视图。胚胎的安装使得燎星的平面垂直于光轴。(C)受精后24小时对照和烧蚀胚胎的存活和形态。比例尺为300μm.(E)在表达Histone2B-mCherry的Tg(gsc:GFP)胚胎的polster中激光烧蚀的时间序列。仅带有绿色通道的视图是最大投影。特写镜头显示了含有细胞碎片的烧蚀区域。具有绿色和红色(显示为洋红色)通道的视图是烧蚀前后的 XY 和 XZ 切片(绿色闪电表示烧蚀)。XZ切片显示,上覆组织(没有GFP表达的洋红色细胞核)没有受到底层结构消融的影响。黄色虚线框对应于为激光烧蚀处理选择的ROI。比例尺在大视图中为50 μm,在特写镜头中为25 μm。请点击此处查看此图的放大版本。
{kind=link}
6. 目标定位和激光烧蚀
- 在实时成像上定位光栅轮廓,并使用电光调制器感兴趣区域 (EOM ROI) 工具绘制一个跨越光栅宽度的 20 像素 (15 μm) 大矩形。将此矩形放在 polster 的中间(图 1E)。
- 请注意包含 polster 像元的最高平面和最低平面的轴向位置。在这两个平面之间每10μm将进行烧蚀。请注意,ROI 不会与任何这些平面上的蛋黄单元格重叠。
- 将载物台放置在间隔的最低 Z 位置。烧蚀必须自下而上地进行,因为碎屑会吸收光线。
- 将绿色/烧蚀激光波长设置为820 nm,并设置 功率百分比 以获得300 mW的退出功率(步骤3.3)。
- 将 成像频率 设置为 200 Hz。
- 将绿色/烧蚀激光成像 EOM 设置为 0,然后选择 ROI 处理 模式。
- 打开 EOM 并将处理设置为立即开始(在 0 帧之后)。
- 将 "成像模式" 设置为"延时摄影",然后停用 "自动保存"。
- 将 "时间步长" 设置为"快速"模式。
- 将 处理帧数 和 帧数 设置为与目标深度对应的值(表 1)。
| 深度(微米) | 治疗框架 |
| -30 | 1 |
| -35 | 1-2 |
| -40 | 1-2 |
| -45 | 2 |
| -50 | 2-3 |
| -55 | 3 |
| -60 | 3-4 |
| -65 | 4 |
| -70 | 4 |
| -75 | 4-5 |
| -80 | 4-5 |
| -85 | 5 |
| -90 | 5 |
| -95 | 5-6 |
| -100 | 6 |
| -105 | 6 |
表1:建议的激光治疗框架数量作为胚胎中靶细胞深度的函数(0是胚胎表面)。
- 开始成像。采集是黑色的,因为在EOM处理期间PMT的快门关闭。
- 将舞台上移到列表的下一个 Z 位置(步骤 6.2)。
- 重复步骤 6.10 到 6.12,直到到达 polster 的顶部。
7. 烧蚀后验证和成像
- 将绿光/烧蚀激光器设置为920 nm和5%功率。将绿色/烧蚀激光成像EOM设置为100,然后选择 全场 模式。
- 将 "成像频率" 设置为 800 Hz。关闭 EOM。
- 在实时模式下浏览整个堆栈,以检查每个平面是否已被烧蚀。如果不是这种情况,请返回步骤 6.2。
注意:消融有时会引起相邻组织的垂直移动,因此可能必须重新定义Z层。 - 将 "成像模式" 设置为"3D 延时摄影",然后重新激活 "自动保存"。记录40-60分钟的消融后电影。
- 在消融后的电影中,检查靶细胞是否被有效消融。荧光回收或靶细胞占据空间并阻止跟随细胞通过,表明靶细胞仅被光漂白而没有消融(图1E和图2A)。

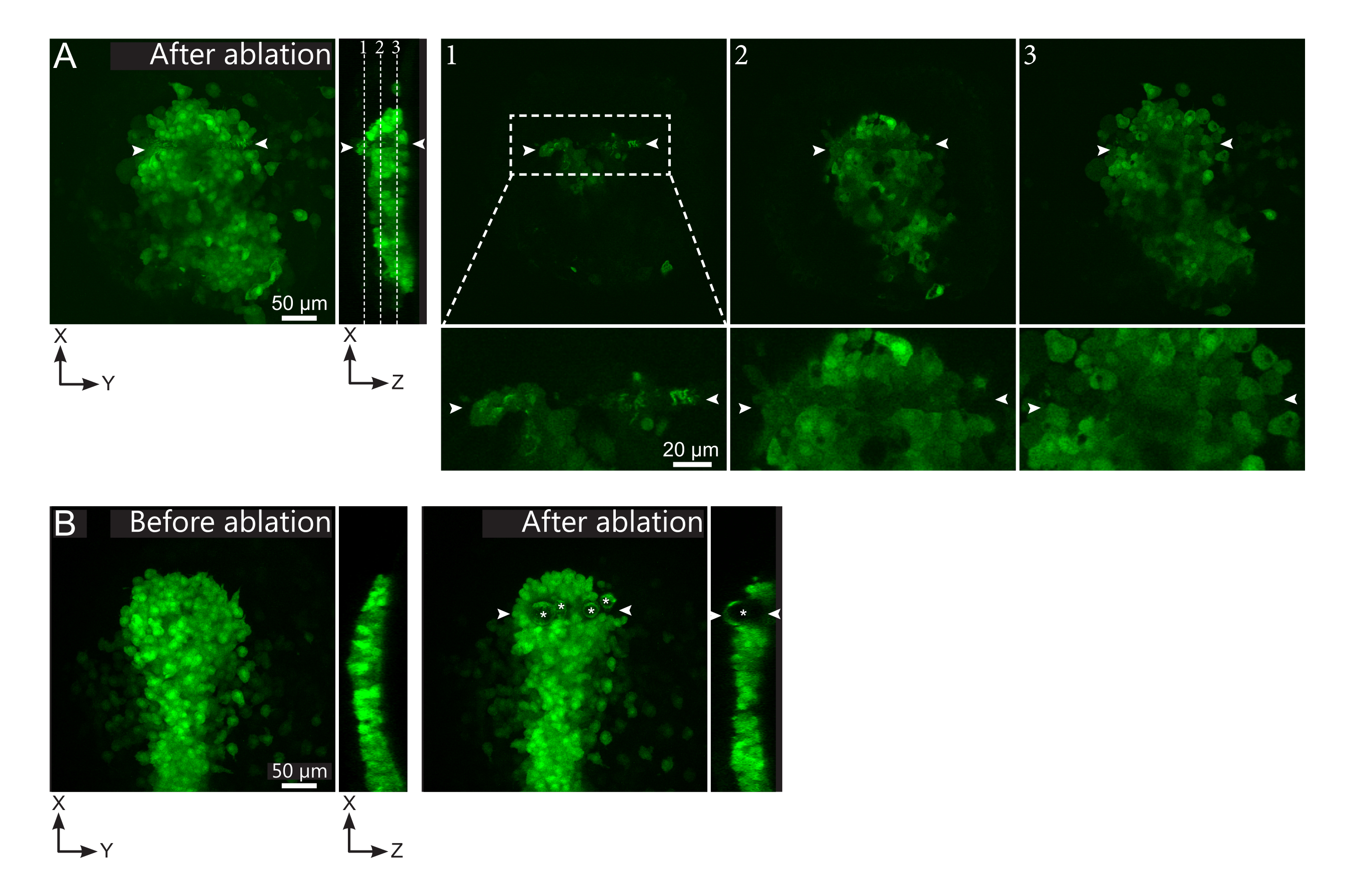
图2:激光烧蚀的阴性结果。 (A)激光烧蚀中潜在故障的典型示例。大型 XY 视图是最大投影,XZ 视图是重建剖面。激光处理区域位于两个白色箭头之间。三个焦平面在重建部分中突出显示,并显示在右侧。它们对应于三种不同类型的故障。平面1显示,栉体上方的细胞已被烧蚀。这可以通过在栉星上方的焦平面上存在自发荧光碎片(见特写)来识别(参见重建部分上平面1的位置)。这可能是由于对要消融的区域的定义不正确造成的。平面2显示了已经漂白但未消融的细胞。它们可以被识别为低荧光信号仍然显示完整的细胞轮廓(见特写)。平面3显示了完整的细胞,这些细胞几乎没有通过激光处理漂白。这可能是由于对要消融的区域的定义不正确或处理不当造成的。在平面2和3中描述的情况下,可以将消融处理重新应用于未消融的靶细胞。比例尺在大视图中为50 μm,在特写镜头中为20 μm。(B)由于激光处理过强而由空化形成的气泡(用白色星号标记)的典型例子。这种气泡不仅限于Z平面,有时甚至跨越了污染物的整个高度,使相邻组织变形。比例尺为50μm, 请点击此处查看此图的放大版本。
{kind=link}
8. 数据分析
- 使用图像分析软件打开延时系列,并设置正确的像素大小。
- 在 Spot 函数中,将 "物体大小" 设置为 10 μm,因为这是原肠胚形成过程中的平均原子核大小。然后,运行 Spot 函数以检测和跟踪原子核。
注意:通过考虑较低的轴向分辨率,沿Z轴拟合12μm长的椭圆体形状,可以略微改善检测。 - 使用筛选器删除误报。在 Tg(Gsc:GFP) 系中,来自胚胎轴的细胞和一些内胚层细胞被标记为绿色。因此,对绿色强度进行滤波可以快速选择这些电池(图3A)。
- 将连续点之间的最大距离设置为与像元速度兼容的值。
注意:请仔细考虑两帧之间的时间间隔。Polster细胞以2.8±0.8μm/min迁移。因此,允许4μm的最大位移持续1分钟的时间步长可以去除大多数伪影轨迹。 - 允许在一个或两个时间点上出现间隙可提供更长的连续跟踪,但可能会引入跟踪错误。如果在一次性点未正确检测到原子核,请考虑使用不同的参数/过滤器重新运行斑点检测。
- 目视检查曲目,并在必要时进行更正。
- 将结果导出为.xlsx文件。使用已发布的电子表格例程24 (图 3B)和数据分析软件上的自定义例程(可根据要求提供)处理文件。

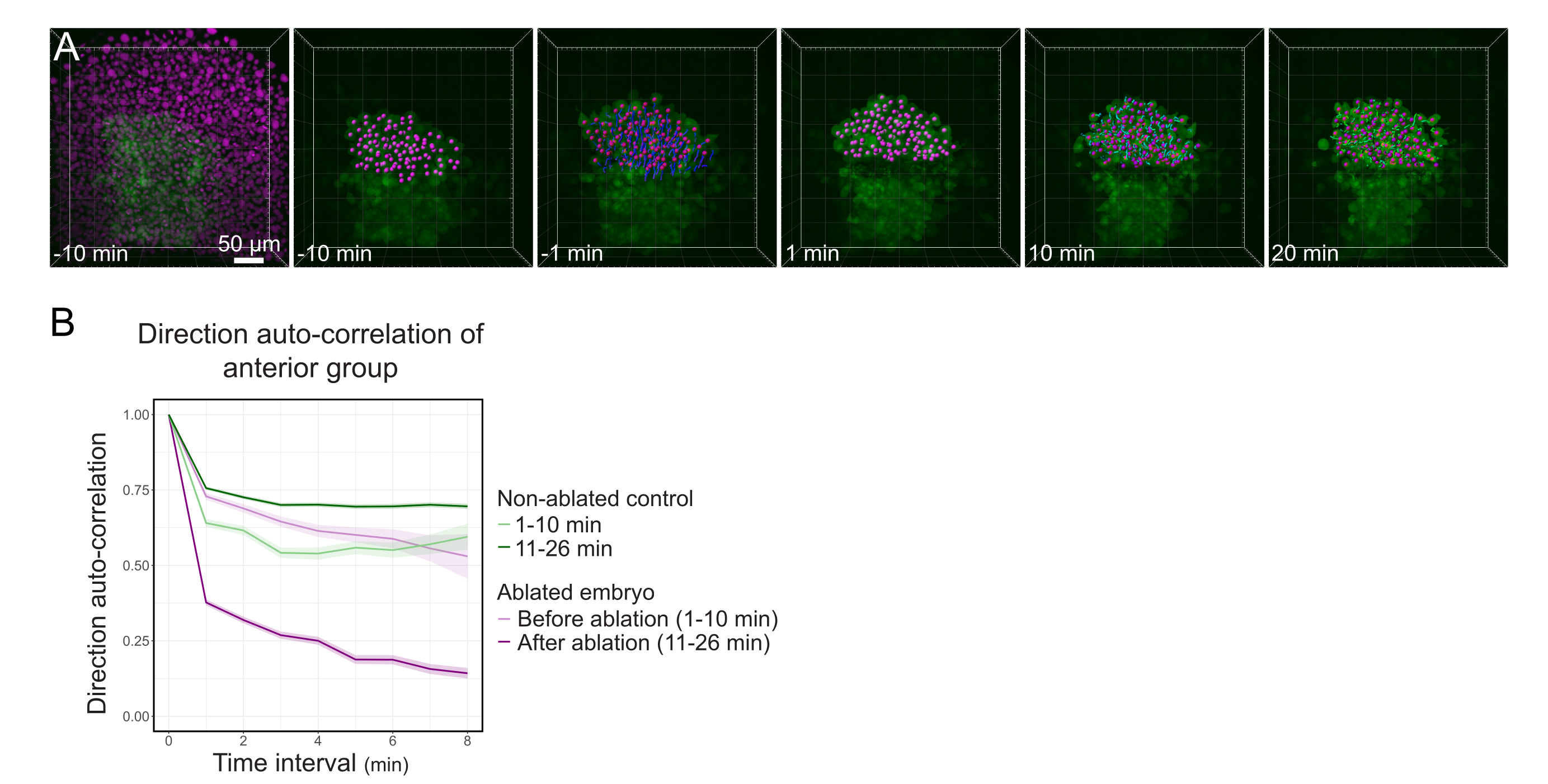
图3:分离出的桔梗前半部分会影响细胞的方向性。 (A)在激光消融切断中间的栉水之前和之后,3D重建表达Histone2B-mCherry的 Tg(gsc:GFP) 胚胎(以洋红色显示)。属于燎星前半部分的细胞核用洋红色点标记,并在消融前后随时间跟踪(见 电影S1)。(B)作为迁移持久性的量度,在消融之前和之后属于Polster前部的细胞的方向自相关。细胞在消融前显示连续运动,消融后急剧减少,表明集体导向迁移的损失。作为对照,还在形成非消融胚胎前半部分的细胞上测量方向自相关。图形包络线指示标准误差。 请点击此处查看此图的放大版本。
{kind=link}
结果
为了切断中间的污染物,注射了Histone2B-mCherry mRNA的 Tg(gsc:GFP) 胚胎在70%的表观阶段安装,如步骤4所述。通过GFP表达鉴定出极点,并且将胚胎安装,以使极点的平面垂直于光轴(图1B)。将胚胎倾斜远离这个位置将使程序复杂化。光必须穿过更多的组织才能到达消融平面,并且消融平面将相对于胚胎轴倾斜。在验证了所有细胞核都正确标记后,记录了10分钟的消融前?...
讨论
在这里,我们描述了一种协议,该协议使用非线性光学来执行深度和空间上定义良好的体积烧蚀。该协议最关键的一步是找到提供足够能量以允许消融的处理条件,但不能过多的能量,以避免过多的碎片或空化。目标部位传递的能量主要取决于:(1)激光退出功率,(2)激光对准的质量,(3)光通过到达烧蚀平面的组织的性质,(4)烧蚀平面的深度。因此,在每次实验之前,测量激光退出功率?...
披露声明
作者声明没有竞争利益。
致谢
我们感谢Emilie Menant的鱼类护理,Polytechnique Bioimaging Facility,特别是Pierre Mahou,感谢在法兰西岛地区(interDIM)和国家研究机构(ANR-11-EQPX-0029 Morphoscope2,ANR-10-INBS-04 France BioImaging)部分支持的设备上提供实时成像。这项工作得到了ANR拨款15-CE13-0016-1,18-CE13-0024,20-CE13-0016以及欧盟地平线2020研究和创新计划的支持,该计划根据Marie Skłodowska-Curie赠款协议No 840201,高级和研究部和国家科学研究中心。
材料
| Name | Company | Catalog Number | Comments |
| 25x water immersion objective | Olympus | XLPLN25XWMP2 | |
| Agarose | PanReac AppliChem | A8963,0500 | |
| Data analysis software : Matlab | Math Works | ||
| Electro-optic modulator (EOM) | ConOptics | 350-80LA | |
| Embryo Medium (EM) solution | Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th Edition. University of Oregon Press, Eugene (Book). (2000). | ||
| Environmental chamber chamber | Okolab | H201-T-UNIT-BL | |
| EOM driver | ConOptics | 302RM | |
| Fluorescence source | Lumencor | SOLA | |
| Glass bottom dishes | MatTek | P35G-0-10-C | |
| Glass capillaries | Harvard Apparatus | 300085 | Outside diameter 1.0 mm, inside diameter 0.58 mm |
| Glass pipettes | Volac | D810 | Tip should be fire polished |
| Green/ablation laser | Spectra Physics | Mai Tai HP DeepSee | |
| Histone2B-mCherry mRNA | Synthesized from pCS2-H2B-mCherry plasmid (Dumortier& al. 2012) | ||
| Image analysis software: IMARIS | Bitplane | ||
| ImSpector software | Abberior Instruments Development Team | ||
| Injection mold | Adapative Science Tools | I-34 | |
| Microloader tips | Eppendorf | 5242956003 | |
| Micromanipulator | Narishige | MN-151 | |
| Micropipette puller | Sutter | P-1000 | |
| mMESSAGE mMACHINE SP6 Transcription Kit | Invitrogen | AM1340 | |
| Penicillin-Streptomycin | Thermofisher | 15140-122 | 10 000 units penicillin and 10 mgstreptomycin per ml |
| Photomultiplier tube (PMT) | Hammamatsu | H7422-40 | |
| PicoPump (Air injector) | World Precision Instrument | PV820 | |
| Red laser | Spectra Physics | OPO/Insight DeepSee | |
| RNAse free water for injection | Sigma | W3500 | |
| Spreadsheet software: Excel | Microsoft | ||
| Stereomicroscope | Nikon | SMZ18 | |
| Tg(gsc:GFP) zebrafish line | Doitsidou, M. et al. Guidance of primordial germ cell migration by the chemokine SDF-1. Cell. 111 (5), 647–59, doi: doi.org/10.1016/S0092-8674(02)01135-2 (2002). | ||
| TriM Scope II microscope | La Vision Biotech |
参考文献
- Slack, J. M. W. Embryonic induction. Mechanisms of Development. 41 (2-3), 91-107 (1993).
- Fernandez-Sanchez, M. -. E., Brunet, T., Röper, J. -. C., Farge, E. Mechanotransduction's impact on animal development, evolution, and tumorigenesis. Annual Review of Cell and Developmental Biology. 31, 373-397 (2015).
- Shih, J., Fraser, S. E. Characterizing the zebrafish organizer: microsurgical analysis at the early-shield stage. Development. 122 (4), 1313-1322 (1996).
- Selleck, M. A. J. Culture and microsurgical manipulation of the early avian embryo. Methods in Cell Biology. 51 (51), 1-21 (1996).
- Bulina, M. E., et al. A genetically encoded photosensitizer. Nature Biotechnology. 24 (1), 95-99 (2006).
- Fang-Yen, C., Gabel, C. V., Samuel, A. D. T., Bargmann, C. I., Avery, L. Laser microsurgery in Caenorhabditis elegans. Methods in Cell Biology. 107, 177-206 (2012).
- Colombelli, J., Grill, S. W., Stelzer, E. H. K. Ultraviolet diffraction limited nanosurgery of live biological tissues. Review of Scientific Instruments. 75 (2), 472-478 (2004).
- Smutny, M., Behrndt, M., Campinho, P., Ruprecht, V., Heisenberg, C. -. P. UV laser ablation to measure cell and tissue-generated forces in the zebrafish embryo in vivo and ex vivo. Methods in Molecular Biology. 1189, 219-235 (2015).
- Behrndt, M., et al. Forces driving epithelial spreading in zebrafish gastrulation. Science. 338 (6104), 257-260 (2012).
- Volpe, B. A., Fotino, T. H., Steiner, A. B. Confocal microscope-based laser ablation and regeneration assay in zebrafish interneuromast cells. Journal of Visualized Experiments: JoVE. (159), (2020).
- Bonnet, I., et al. Mechanical state, material properties and continuous description of an epithelial tissue. Journal of the Royal Society, Interface. 9 (75), 2614-2623 (2012).
- Rauzi, M., Lenne, P. F., Lecuit, T. Planar polarized actomyosin contractile flows control epithelial junction remodelling. Nature. 468 (7327), 1110-1115 (2010).
- Niemz, M. H. . Laser-Tissue Interactions. Encyclopedia of Biomaterials and Biomedical Engineering, Second Edition - Four Volume Set. , (2019).
- Smith, A. M., Mancini, M. C., Nie, S. Bioimaging: second window for in vivo imaging. Nature Nanotechnology. 4 (11), 710-711 (2009).
- Rauzi, M., Lenne, P. -. F. Cortical forces in cell shape changes and tissue morphogenesis. Current Topics in Developmental Biology. 95, 93-144 (2011).
- Theveneau, E., David, N. B. Migrations cellulaires collectives. Medecine/Sciences. 30 (8-9), 751-757 (2014).
- Dumortier, J. G., Martin, S., Meyer, D., Rosa, F. M., David, N. B. Collective mesendoderm migration relies on an intrinsic directionality signal transmitted through cell contacts. Proceedings of the National Academy of Sciences of the United States of America. 109 (42), 16945-16950 (2012).
- Solnica-Krezel, L., Stemple, D. L., Driever, W. Transparent things: cell fates and cell movements during early embryogenesis of zebrafish. BioEssays. 17 (11), 931-939 (1995).
- Montero, J. -. A., Kilian, B., Chan, J., Bayliss, P. E., Heisenberg, C. -. P. Phosphoinositide 3-kinase is required for process outgrowth and cell polarization of gastrulating mesendodermal cells. Current Biology. 13 (15), 1279-1289 (2003).
- Ulrich, F., et al. Slb/Wnt11 controls hypoblast cell migration and morphogenesis at the onset of zebrafish gastrulation. Development. 130 (22), 5375-5384 (2003).
- Kai, M., Heisenberg, C. -. P., Tada, M. Sphingosine-1-phosphate receptors regulate individual cell behaviours underlying the directed migration of prechordal plate progenitor cells during zebrafish gastrulation. Development. 135 (18), 3043-3051 (2008).
- Smutny, M., et al. Friction forces position the neural anlage. Nature Cell Biology. 19 (4), 306-317 (2017).
- Johansson, M., Giger, F. A., Fielding, T., Houart, C. Dkk1 controls cell-cell interaction through regulation of non-nuclear β-Catenin pools. Developmental Cell. 51 (6), 775-786 (2019).
- Gorelik, R., Gautreau, A. Quantitative and unbiased analysis of directional persistence in cell migration. Nature Protocols. 9 (8), 1931-1943 (2014).
- Grill, S. W., Howard, J., Schäffer, E., Stelzer, E. H. K., Hyman, A. A. The distribution of active force generators controls mitotic spindle position. Science. 301 (5632), 518-521 (2003).
- Desprat, N., Supatto, W., Pouille, P. -. A. A., Beaurepaire, E., Farge, E. Tissue deformation modulates twist expression to determine anterior midgut differentiation in Drosophila embryos. Developmental Cell. 15 (3), 470-477 (2008).
- Farhadifar, R., Röper, J. -. C., Aigouy, B., Eaton, S., Jülicher, F. The influence of cell mechanics, cell-cell interactions, and proliferation on epithelial packing. Current Biology. 17 (24), 2095-2104 (2007).
- Willier, B. H., Oppenheimer, J. M. . Foundations of Experimental Embryology. , (1964).
- Ashby, W. J., Zijlstra, A. Established and novel methods of interrogating two-dimensional cell migration. Integrative Biology: Quantitative Biosciences from Nano to Macro. 4 (11), 1338-1350 (2012).
- Bosze, B., et al. Pcdh18a regulates endocytosis of E-cadherin during axial mesoderm development in zebrafish. Histochemistry and Cell Biology. 154 (5), 463-480 (2020).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。