
Method Article
Vibratom Schnitte Maus-Retina zu Photorezeptor Kulturen vorbereiten
In diesem Artikel
Zusammenfassung
Neuronale Netzhaut einer Maus im Alter von 8 Tagen auf einem 4% igen Gelatineblock. Nach Isolierung der Photorezeptorschicht (200 um) durch Vibratom werden die Photorezeptoren nach der mechanischen und enzymatischen Spaltung der Kultur beimpft. Die Photorezeptorschicht kann für die molekulare, biochemische Analysen oder Transplantation verwendet werden.
Zusammenfassung
Die Retina ist ein Teil des zentralen Nervensystems, die Architektur in Schichten von den Photorezeptoren der Netzhaut-Pigmentepithel in der am weitesten entfernten Teil auf der Netzhaut unter Berücksichtigung der Richtung der Licht organisiert ist, mit Neuronen, die beiden Stäbchen und Zapfen in Kontakt, und die Ganglienzellen in der proximalsten Abstand. Diese Architektur ermöglicht die Isolierung der Photorezeptorschicht durch Vibratom Schnitte. Die seziert neuronalen Netzhaut einer Maus im Alter von 8 Tagen Flach eingebettet in 4% Gelatine auf einer Scheibe von 20% Gelatine Photorezeptorschicht nach unten. Mit einem Vibratom und ein Doppel Rasierklinge wird der 100 um dicken inneren Netzhaut geschnitten. Dieser Abschnitt enthält die Ganglienzellen und die innere Schicht mit insbesondere den bipolaren Zellen. Ein Zwischenabschnitt 15 um verworfen wird, bevor 200 & mgr; m der äußeren Netzhaut, die die Photorezeptoren wird gewonnen. Die Gelatine wird durch Erhitzen bei 37 ° C entfernt. Stücke von äußeren Schicht sind Inkubatorted in 500 ul Ringer-Lösung mit 2 Einheiten aktiviert Papain für 20 min bei 37 ° C. Die Reaktion wird durch Zugabe von 500 ul 10% fötalem Kälberserum (FCS) in Dulbeccos modifiziertem Eagle Medium (DMEM) angehalten wird, dann 25 Einheiten DNAse I wird vor der Zentrifugation bei RT mehrere Male zugegeben gewaschen, um Serum zu entfernen, und die Zellen werden erneut suspendiert in 500 ul DMEM und bei 1 x 10 5 Zellen / cm 2 ausgesät. Die Zellen werden 5 Tage in vitro gezüchtet und ihre Lebensfähigkeit mit Live / Dead-Assay erzielt. Die Reinheit der Kultur wird zunächst durch mikroskopische Beobachtung während des Experiments bestimmt. Die Reinheit wird dann durch Impfen und Fixierung von Zellen auf einem histologischen Objektträgers und Analyse mit einem polyklonalen Kaninchen-anti-SAG, einen Photorezeptor Marker und monoklonalen Maus-Anti-RHO, eine Stange Photorezeptor spezifischer Marker validiert. Alternativ kann die Photorezeptorschicht (97% Stangen) zum Gen- oder Proteinexpressionsanalyse und für die Transplantation verwendet werden.
Einleitung
Die Retina ist ein integraler Teil des zentralen Nervensystems, die eine konservierte Architektur innerhalb der Wirbeltiere hat. Die Neuronen des neuronalen Netzhaut sind in Schichten organisiert, mit dem entferntesten für das einfallende Licht, die Photorezeptorschicht in engem Kontakt mit der retinalen Pigmentepithel (RPE) auf der Rückseite des Auges. Stäbchen und Zapfen photorecptors sind lichtempfindlichen Zellen, die auf Opsin empfindlicher Moleküle für Photonenerfassung verlassen. Diese Moleküle werden auf Platte Membranen einer zellulären Struktur, auf der äußeren Segment des Photorezeptors, die in Richtung des RPE 1 weist befindet umschlossen. Diese Struktur, die häufig früh bei der Photorezeptordegeneration betroffen ist, wird mit einer Rate von 10% pro Tag erneuert. Die sogenannte innere Schicht enthält die meisten der anderen Neuronen, die von den Photorezeptoren, die bipolare, amakrinen und horizontalen Zellen sowie der Ganglienzellen empfangenen Signals zu berechnen. Diese letzteren mit ihren Axonenbilden einen Strahl, der Sehnerv. Diese Schichtung ist so konserviert, dass Biologen den Begriff verdrängt Amakrinzellen verwendet, wenn die Zellen außerhalb der inneren plexiformen Schicht 2 gefunden. Neuronenschichten innerhalb eines Anker radialer Müller Gliazellen verteilt. Bipolare Zellen verknüpfen Photorezeptoren zu Ganglienzellen. Sie werden zwischen dem externen Plexiformschicht und der inneren Plexiformschicht entfernt. Die Ganglienzellen bilden die inneren Plexiformschicht in Verbindung mit den bipolaren Zellen. Die amacrin Zellen werden als Assoziation Zellen im Innen Plexiformschicht zwischen den bipolaren Zellen und Ganglionzellen befindet gestattet. Die äußere plexiformen Schicht enthält Horizontalzellen. Diese einzigartige Anordnung von neuronalen Schichten des zentralen Nervensystems erlaubt die Isolierung der Photorezeptorschicht von der inneren Zellschicht, die durch das Schneiden der flachen montierten Netzhaut mit einem Vibratom.
Ursprünglich wurde diese Technik verwendet werden, um Photorezeptoren für tran isolierensplantation in das Auge des rd1 Maus, ein Modell der menschlichen Retinitis pigmentosa (RP) 3. Die rd1 Maus trägt eine rezessive Mutation im Gen, das für PDE6B stab spezifischen Phosphodiesterase beta-Untereinheit kodiert. Rezessive Mutationen dieses Gens führen zu RP bei Menschen 4. Nach Stäbchen-Photorezeptoren sind degeneriert, verliert der Patient Nachtsicht, und überraschend Zapfen-Photorezeptoren, die nicht das mutierte Gen exprimieren, entartet in einem zweiten Schritt. Da die Zapfen sind für das Farbsehen und die Sehschärfe erforderlich, die Patienten werden zunehmend blind und eine wirksame Behandlung für die Krankheit ist noch nicht entwickelt. Durch Pfropfen von Photorezeptorschicht von einer Wildtyp-Maus die Zapfen-Degeneration des Host-Maus verzögert 3,5. Die in der Kolbenstangenkonus degenerative Modells verloren Stangen nicht durch ein Transplantat ersetzt werden, da der synaptischen Verbindung zwischen den Stäben und bipolare Zellen nur in einem bestimmten Stadium der r erhalten werden,etinal Entwicklung, durch das Einsetzen Nrl Ausdruck 6 gekennzeichnet. Die Schicht von Photorezeptoren durch eine Operation in dem subretinalen Raum des stangenlosen rd1 Netzhaut eingeführt wurde, zwischen der RPE und der äußeren Netzhaut entspricht, nur 3% der verbleibenden Photorezeptoren, die Kegel. Zwei Wochen nach der Operation, überlebten 40% der Kegel aus dem Tier mit einem normalen Photorezeptorschicht transplantiert, verglichen mit dem Tier, das mit einer normalen Schicht inneren Netzhautzellen oder zur scheinoperierten Tier transplantiert. Die Topographie des Konus Überleben ausgebreiteten über die gesamte Oberfläche des mutierten Retina an der Position des transplantierten Gewebes befindet, angibt, dass die Schutzwirkung aufgrund eines diffundierbaren Molekül 7.
Als nächstes untersuchten wir eine Co-Kultur-System als auch Kulturmedien, die Tatsache, dass die Schutzwirkung beruht auf der Sekretion eines Proteins durch Stangen 8,9 zu validieren. Wir vermuten, dass dieses Protein wird expr werdenkontinuierlich und zwar grundsätzlich über Stangen und ihrem Tod in der ersten Phase der Krankheit wird sekundären Zapfen-Degeneration durch den Verlust eines Schutzsignals von Stangen in einer nicht-zellautonom 10 auszulösen. Wegen der Bedeutung von Kegel vermittelten zentralen Sehens in Primaten diese putative Protein vor erheblich therapeutisches Werkzeug für RP. Die Erhaltung der Zapfen in RP theoretisch verhindern, insgesamt 1,5 Millionen Patienten weltweit blinde 11 zu werden. Wir haben einen hohen Gehalt Screening und eine Kegel angereicherten Kulturmodell verwendet, um eine cDNA-Rod-abgeleitete Cone Lebensfähigkeit Factor (RdCVF) Codierung von einem Netzhaut-cDNA-Bank 12 zu identifizieren. RdCVF ist die gespleißte Produkt der NXNL1 Gen, interessanter homolog zu dem Gen für die Proteine im Thioredoxin Redoxhomöostase 13 beteiligt. Die zweite gespleißte Produkt des Gens ist RdCVFL ein Enzym, das seine Ziel, des Tau-Proteins gegen oxidativen d schütztAmage 14. Die Verabreichung von RdCVF verhindert, dass der sekundäre Degeneration der Zapfen und den Verlust ihrer Sehfunktion in einem rezessiven und dominanten Modell der RP 12,15. Dies zeigt zwei wichtige Aspekte dieses innovativen Therapieansatz 16. Erstens kann es in den meisten Fällen in einem RP-Gen-unabhängige Art und Weise aufgebracht werden. Zweitens wird im Gegensatz zu den konkurrierenden Faktor CNTF, RdCVF Überleben mit der Erhaltung der Sehfunktion 17 verbunden. Das Fehlen von funktionellen Effekt kann den Grund der Abwesenheit von klinischen Nutzen der Verabreichung von CNTF auf RP-Patienten 18 zu erklären. RdCVF ist wahrscheinlich einer der wichtigsten Überlebenssignal zwischen Stäbchen und Zapfen, da Kegel Rettungs in vitro durch RdCVF Immundepletion 12 gehemmt. Darüber hinaus ist die Störung des stab abgeleitet Konus Lebensfähigkeit Gens führt zu Funktionsstörungen und die Anfälligkeit für oxidativen Stress 19 Photorezeptor.
Die Verwendung vonPhotorezeptorschicht am Ursprung Identifizierung RdCVF und eines neuartigen Redoxsignals bei neurodegenerativen Erkrankungen 20 beteiligt. Diese Handschrift beschreibt die Methode zur Isolierung und Kultivierung von Zellen aus der Photorezeptorschicht, die Aktivität RdCVF charakterisieren Protokoll. Die Fotorezeptoren können in Kultur für 5 bis 7 Tagen 21 aufrechterhalten werden. Diese Technik kann auch verwendet werden, um die Expression von spezifischen Genen Photorezeptor zu untersuchen.
Protokoll
HINWEIS: Das Verfahren wurde von der Ethikkommission Darwin von der Universität Pierre und Marie Curie (Ce5 / 2009/048) genehmigt
1. Herstellung von Gelatine-Lösung, Instrumente, Vibratom, Kultur, Medien und Kulturplatte
- Vorbereiten einer 20% igen Gelatinelösung, mindestens einen Tag vor dem Experiment.
- Unter der Haube, fügen Sie 500 ul Gentamycin [10 mg / ml], um einen 500-ml-Flasche mit CO 2 unabhängige Medium (CO 2 -i).
- In einem separaten Becherglas mit 20 ml CO2-I (aus Schritt 1.1.1) und Wärme bei 95 ° C auf einem Heizblock unter ständigem Rühren für 15 min. Beachten Sie einen Farbwechsel nach Gelb zeigt an, dass die Lösung für die weitere Verwendung.
- Man wiegt 4 g Gelatine und allmählich arm zu der oben hergestellten Lösung (Schritt 1.1.2) mit erhöhtem Rühren und Erwärmen auf 95 ° C für 45 min, um eine gelb gefärbte Lösung erhalten. Lassen Sie die Gelatine-Lösung unter Kühlungdie Haube für ca. 5 min.
- Pour 4 ml der Gelatine-Lösung in eine 35 mm-Durchmesser-Kulturschalen ohne Einführung von Luftblasen. Halten Sie die Schale bei 21 ° C für 30 min. Decken Sie das Geschirr mit Kunststofffolie, und drehen Sie die Speisen auf den Kopf. Wahlweise speichern die Gerichte bei 4 ° C für bis zu 2 Monate vor dem Gebrauch.
- Bereiten Sie eine 4% ige Gelatinelösung.
- Unter der Haube 25 ml CO 2 -i Medium in einem sterilen Kunststoffbehälter und Wärme bei 42 ° C auf einem Thermoblock. Entfernen des Mediums aus dem Heizblock und nach und nach 1 g Gelatine zu der heißen Medium und Rühren. Schnell wieder den Behälter auf den Heizblock (42 ° C) und warm halten während des Prozesses.
- Aufstellen des Vibratom Gerät:
- Entfernen der Schalen, enthaltend 20% bei 4 ° C gelagert Gelatinelösung. Schneiden Sie die Gelatine mit einem Skalpell. Drehen Sie die Gelatine in Scheiben schneiden und kleben Sie es auf dem Schwarzstützscheibe des Vibratom mit einem Tropfen super Leim.
- Brechen Sie eine Rasierklinge in zwei Hälften, und legen Sie eine Hälfte in die Vibratom Halteaufnahme. Setzen Sie die schwarze Trägerscheibe auf die Vibratom Gerät. Schalten Sie den schwarzen Knopf auf der rechten Seite. Befestigen Sie die Halteaufnahme auf den Kopf des Vibratom. Es wird ausreichend viel CO 2 -i Mediums im Vibratom Tank, um die Gelatine Block abzudecken.
- Schalten Sie die Vibratom geschnitten und drei 100 & mgr; m Scheiben Gelatineblock. Halten Sie die Gelatine-Block für die weitere Verwendung.
- Bereiten Sie 40 ml Kulturmedium:
- Bereiten Dulbeccos modifiziertem Eagle-Medium (DMEM) mit 10% fötalem Kälberserum (FCS) unter der Haube. Herstellung von 1 mg / ml Poly-D-Lysin unter Verwendung von Phosphat-gepufferter Saline (PBS), pH 7,4.
- In einer 96-Well-Platte, add 2 ug / cm 2 von Poly-D-Lysin zur Beschichtung der Boden der Vertiefungen. Inkubiere die Platte für 45 min bei 37 ° C in 5% CO 2 -Inkubator. Nach der Inkubation entfernen PBS und ersetzen mit 200 & mgr;/ Well DMEM und Ort wieder bei 37 ° C in 5% CO 2 -Inkubator vor dem Gebrauch.
2. Präparation der Entire Retina
- Entkernen Sie die Maus:
- Opfern Sie die Maus nach der europäischen Richtlinie 2010/63: durch Genickbruch. Schneiden Sie den Kopf mit geschwungenen-Schere und legen Sie sie in einem Durchmesser von 100 mm Petrischale, nachdem sie mit Desinfektionsmittel desinfiziert das Auge.
- Entfernen Sie die Augen, nachdem er den Sehnerv abgelöst durch Schneiden sie sehr vorsichtig mit einem gebogenen Griff. Dann legen beide Augen in einer 35 mm Petrischale mit steriler CO 2 -i Medium bei 21 ° C.
- Entfernen der Netzhaut:
- Machen Sie ein Loch in der Höhe des Auges-Glied mit Hilfe einer 18 G-Nadel, um die Schere ins Auge Welt einzuführen. Einzuführen geraden Schere in das Loch und sorgfältig geschnitten Sklera unterhalb der Iris auf dem ganzen Umfang des Augapfels zu Ziliarkörper entfernen.
- Entfernen Sie die Hornhaut und die Linse und halten Sie die Hinterkammer des Auges mit der Netzhaut zusammenhängend an der Lederhaut. Mit zwei feinen Griff, entfernen Sie die Glaskörper in der Hinterkammer befindet, ohne die Netzhaut. Führen Sie vier radiale Einschnitte (Netzhaut / Lederhaut) zu Abflachung der Netzhaut zu ermöglichen.
- Aufschwung des Augapfels und ziehen ihn als "orange". Trennen Sie die Netzhaut von der Lederhaut und Netzhaut-Pigmentepithel (RPE). Trennen Sie die Netzhaut noch auf dem Stand des Sehnervs sehr vorsichtig mit feinen gekrümmten Schere befestigt.
- Nehmen Sie die verbleibenden Glaskörper von der Peripherie in Richtung der Mitte der Netzhaut mit zwei feinen Griffen. Stellen Sie sicher, dass die gesamte Glaskörper vollständig entfernt wird, um die Abflachung der Netzhaut zu ermöglichen.
- Schneiden Sie das Ende einer Plastikpipette und übertragen die Netzhaut zu einem Durchmesser von 35 mm Petrischale mit CO 2 -i Medium mit der Plastikpipette.
3. Abdichtung von Flach mounted Retina auf Gelatine-Block
- Entfernen 20 - 30 ml CO 2 -i Medium aus dem Vibratom Tank (die Gelatine Block frei ist), um die Abdichtung des Flach montiert Retina ermöglichen.
- Übertragen Sie die Netzhaut auf einen Glasobjektträger mit einer Plastikpipette und einen Tropfen CO 2 -i Medium vor der Übertragung in die Gelatine-Schnitte mit dem Photorezeptor nach unten auf die Gelatine in Scheiben schneiden.
- Befestigen der Netzhaut zu der Gelatineblock durch leichtes Einspritzen erwärmt 4% Gelatine auf einer Seite des Blocks zwischen der Netzhaut und der Gelatine-Block und gleichzeitig Ausstoßen warmen Gelatine auf der anderen Seite.
- Aspirat 4% Gelatine mit einer Pasteur-Pipette. Warten Sie 10 Minuten, um insgesamt Abdichtung ermöglichen. In CO 2 -i Medium, um den Block und die Klinge vollständig einzutauchen.
4. Schnitte der Retina
HINWEIS: Dieser Schritt ist wichtig, um eine Schicht von Photorezeptorzellen, ohne die anderen Netzhautzellen zu erhalten (Abbildung 1).
- Ausgehend von der Spitze auf die Gelatine-Block, schneiden Sie 100 um Serienschnitte, um die Netzhaut zu erreichen. Dann schneiden Sie 100 bis 120 um Abschnitte der inneren Schicht. Je nach Anwendungsfall kann diese Schicht verworfen oder in flüssigem Stickstoff gelagert werden. Sicherzustellen, dass die Geschwindigkeit des Vibratom ist sehr langsam (in 1 oder 2) während des Schneid der inneren Schicht oder Schichten Photorezeptoren.
- Führen Zwischen 15 um Abschnitte der Netzhaut, die der Grenzfläche zwischen der inneren und der äußeren Netzhaut. Beachten Sie diesen Abschnitt unter dem Mikroskop. Das Fehlen der Blutgefäße gibt an, dass die äußere Netzhaut erreicht.
- Schneiden 200 um der äußeren Netzhaut (der Photorezeptor-Schicht) mit Gelatine und die Übertragung auf einen Durchmesser von 35 mm Schale mit 3 ml CO 2 -i Medium gefüllt. Halten Sie es auf Eis, bis alle Photorezeptorschicht Schnitte aller Netzhaut erhalten werden.
5. distanziert Sehzellen
- Nehmen Sie die Schale (n), welche die Photorezeptorschicht (en) und so (sie) in einem Brutschrank bei 37 ° C für 10 min.
- Mit einer Pinzette vorsichtig trennen die Photorezeptorschicht von der Gelatine und übertragen sie in einem neuen Gericht mit 3 ml Ringer-Lösung gefüllt. Wiederholen Sie diesen Schritt (5.2) für jeden Photorezeptorschicht Vorbereitung.
- (; 5,5 mM L-Cystein, 1,1 mM EDTA 60 & mgr; M β-Mercaptoethanol) 2 Einheiten Papain mit 25 ul Aktivatorlösung Inkubieren in einem 5-ml-Polypropylen steriles Röhrchen und Inkubation er 30 min bei 37 ° C in 5% CO 2 Inkubator . Die Aktivierung von Papain bei 37 ° C erreicht. Während dieser Inkubation schneiden die Photorezeptorschicht in 2 mm 2 Stück (nicht zu klein) und übertragen Sie diese Stücke in eine 5-ml-Röhrchen.
- Zweimal Spülen der Netzhaut mit 1,5 ml einer Lösung, Ringer-Lösung, gefolgt von Schwerkraftsedimentation. Löschen aller restlichen Ringerlösung aus dem Röhrchen mit der Probe.
- Hinzufügen 475 ul Ringer-Lösung, um das Röhrchen mit Aktiv Papain und Mischung. Diese Lösung wird auf das Rohr, das die Netzhaut.
- Das Röhrchen mit der Netzhaut für 20 min bei 37 ° C in 5% CO 2 -Inkubator. Die Reaktion durch Zugabe von 1 ml 10% FCS in DMEM. Hinzufügen 25 U Desoxyribonuclease I (DNAse I) bis verdaute DNA von Todeszellen. Sorgfältig zu homogenisieren die Zellsuspension mit einem 1 ml-Pipette. Spin bei 50 × g für 6 Minuten bei RT.
- Überstand verwerfen, um Spuren von Serum zu entfernen und DMEM-Medium mit Ergänzungen (siehe Tabelle der spezifischen reagents_equipment) an die Zellpellet resuspendieren und sorgfältig die Zellsuspension mit einer 1 ml Pipette. Spin bei 50 × g für 6 Minuten bei RT. Wiederholen Sie diesen Schritt (5.7) noch einmal.
6 Kultivierung Sehzellen
- Die Photorezeptorschicht von der Maus am PN8 enthält etwa 1 bis 1,5 Millionen Zellen mit dieser Technik. Seed die Sehzellen in 1x 10 5 Zellen / cm 2 in einer Kulturplatte mit Kulturmedium und die Kultur werden die Zellen für 5 Tage bei 37 ° C in 5% CO 2 -Inkubator. Am Tag 5 nach der Immunchemie und Western-Blot-Studie folgenden Methoden, die von den Antikörperlieferanten nicht vorzunehmen.
HINWEIS: Die Schritte, um die Kultur zu stoppen und Vorversuche sind in Bezug 21 beschrieben.
Ergebnisse
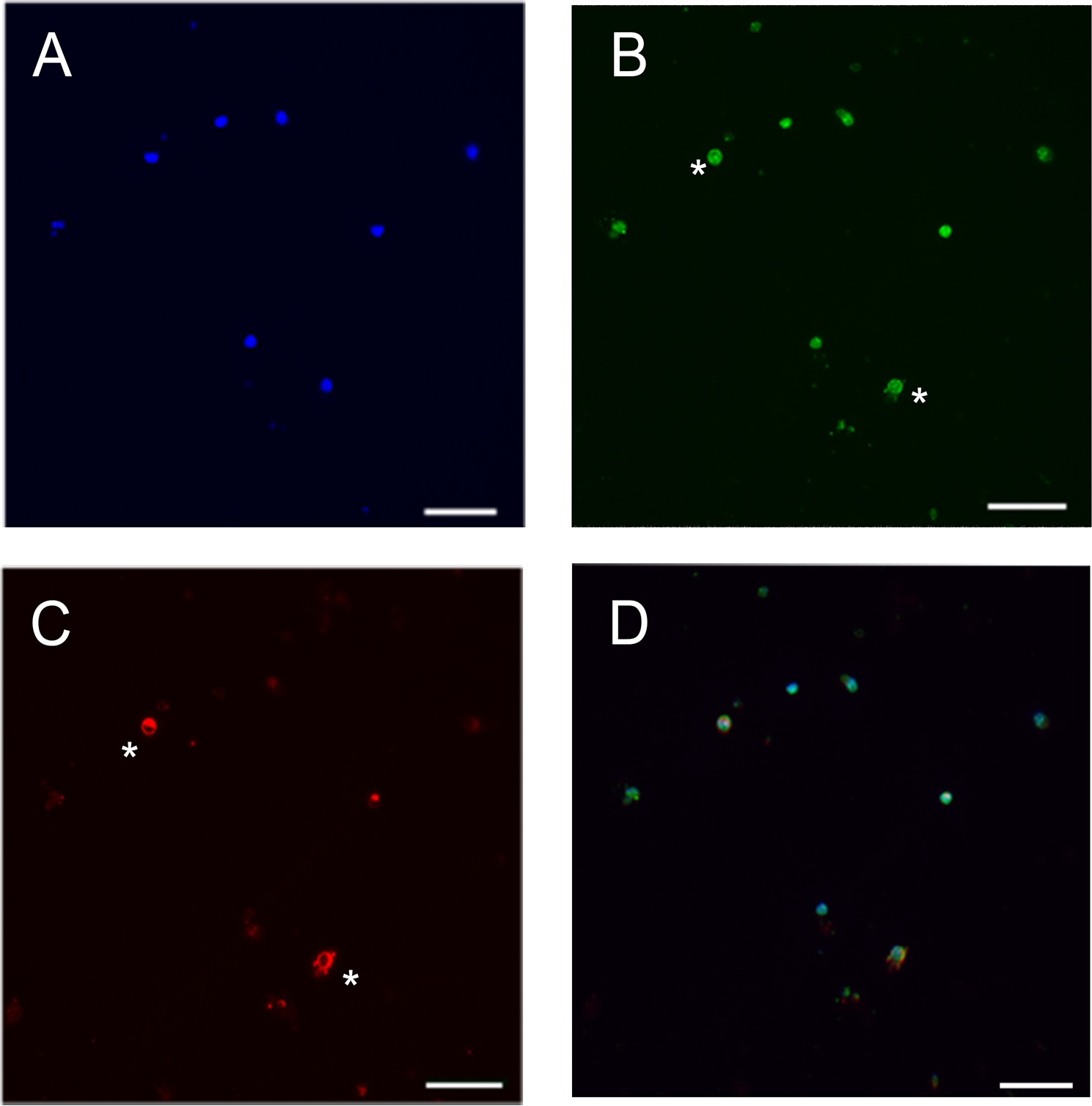
Abgesehen von der Transplantation wurden die Fotorezeptorschichten auch verwendet, um Zellsignalisierung durch Impfen Zellen zur Herstellung von Photorezeptorkulturen 12,22 studieren. Darüber hinaus werden sie zur Genexpression und zirkadianen Rhythmus 23,24 studieren. Wir haben die Zellen Photorezeptorschicht aus einer Wildtyp-Maus am postnatalen Tag 8 für 5 Tage bei 37 ° C in einem Inkubator mit 5% CO 2 (1E zur Photorezeptor-Kulturen in Abwesenheit von FCS vorbereitet und verwaltet, die Zellen ). Die Zellen wurden dann mit 4% paraformadehyde fixiert und dann für immunocytochemische Analyse unter Verwendung von Methoden, die von den Antikörper-Anbieter entweder mit Maus-monoklonalem anti-RHO vorgesehen fort (1: 250, Millipore Mab5316) oder polyklonalem Kaninchen-anti-SAG (1: 200, eine großzügige Geschenk des Igal Gery und David Hicks). Wir verwendeten anti-RHO und Anti-SAG (obwohl Anti-SAG-Label Zapfen und Stäbchen) aufgrund der Tatsache, dass Anti-SAG ist ein früher Marker, als anti-RHO. Der Anteil der cells positiv für die anti-RHO Antikörper ist niedriger als positiv für die Anti SAG Antikörper (Abbildung 2). Dies kann durch die Tatsache, dass die SAG, der Stab Arrestin wird auch von Konen, die 3% der Photorezeptoren zu machen in der isolierten Schicht, oder auch durch die Tatsache, dass während der postnatalen Reifung der Netzhaut, der Ausdruck der SAG voraus, dass ausgedrückt erklärt werden RHO 25,26.
Die Photorezeptorschicht wurde auch verwendet, um die spezifische Expression von Genen durch Photorezeptoren der Wildtyp Netzhaut und Gehirn von Tieren im Alter von 35 Tage RNA wurden mit CsCl Ultrazentrifugation 27 erstellt und Maus DNA-Chip-Array hybridisiert überwachen. Die Boten für Rhodopsin (Rho), S-Arrestin (Sag) und kegel Transducin (GNAT2) sind speziell in der Netzhaut im Vergleich zum Gehirn (3A) ausgedrückt. Der Ausdruck ist prominent in der Photorezeptorschicht (PR) als in der gesamten Netzhaut, die umfasstdie innere Retinaschicht zeigt, dass diese Gene in der Tat von Photorezeptoren exprimiert. Für GNAT2 die relative Expression verglichen mit Rho und Sag zeigt, daß es durch Zapfen in dem isolierten Photorezeptorschicht enthalten ausgedrückt. Die Expression Recoverin (Rcvrn) in der Photorezeptorschicht im Vergleich zu der gesamten Retina erhöht wird (3B).
Die Photorezeptorschicht wurde auch verwendet, um die Expression von RHO und GNAT2 mittels Western Blotting folgenden Verfahren durch den Antikörper Lieferanten bereitgestellt zu überwachen und an die innere Retinaschicht (4) zu vergleichen. Beachten Sie das Fehlen von RHO und GNAT2 28, die Marker von Stäbchen und Zapfen, die jeweils in den gesamten Netzhaut und in der Netzhaut des rd1 Maus am postnatalen Tag 35.

FiAbbildung 1. Schematische Darstellung der Vibratom Schnitte von Maus Netzhaut. (A) Installation des Flach montierte Netzhaut mit den Photorezeptor nach unten auf die Gelatine in Scheiben schneiden. (B) Abschnitt der inneren Netzhaut mit dem Vibratom Klinge. (C) Abschnitt der äußeren Netzhaut aufgenommen Gelatine. (D) Kontrolle der die Anwesenheit des Photorezeptors an der Kante des Fragments der Netzhaut nach der Isolierung des Photorezeptorschicht. Maßstab für grüne Fluoreszenz verwendet wird von einer weißen Lichtmikroskop verwendet. Die Photorezeptoren sind in der Bildrand befindet. (E) Kultivierte Sehzellen (post-fünf Tage). (F) Höhere Vergrößerung von Panel E.

Abbildung 2: Differentielle Expression von RHO und SAG in Maus Photorezeptor Kultur. (A) Färbung der Kerne (blau), (B) SAG-Färbung (grün), (C) RHO-Färbung (rot). (D) zusammengeführten Bild. Da Färbung der SAG früher als RHO Färbung ausgedrückt, sind nur zwei Zellen doppelt markierten SAG und RHO (siehe *). Maßstab:. 23 um Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.
{kind=link}

Abbildung 3. Vergleich der Expression der drei Gene im Gehirn und ganze Netzhaut und Photorezeptoren der Wildtyp-Tiere im Alter von 35 Tage. (A) Rhodopsin (Rho), S-Arrestin (Sag) und kegel Transducin (GNAT2). (B) Calbindin (Calb1) und Recoverin (Rcvrn). Die Daten werden als relative Expression angezeigt. Die relative Ausdruck ist ein Ausdruck Wertnach normaliszation mit der Software Robust Multiarray Durchschnitt (RMA) in der Wissensdatenbank kbass (http://kbass.institut-vision.org/KBaSS/) von Microarray-Daten erhalten.

Abbildung 4. Expression von RHO und GNAT2 im äußeren Netzhaut von einer Wildtyp-Netzhaut im postnatalen Tag 35 Abwesenheit ihrer Expression in der inneren Netzhaut und in der Netzhaut des rd1 Maus am postnatalen Tag 35 ACTB, beta -Actin. Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.
{kind=link}
Diskussion
Die Netzhaut ist ein Modell Orgel in der Biologie. Untersuchung der Netzhaut führte zu 6 großen Entdeckungen in der Biologie. Es ist am Anfang des ersten Tumorsuppressorgens RB1. Es zeigt die molekulare Verbindung zwischen den Rezeptor-Tyrosinkinasen und die MAP-Kinasen durch die Interaktion mit Sohn sevenless. Es war in der Entdeckung PAX6, der ersten übergeordneten Steuerung Gen für Orgel Morphogenese beteiligt. Es ist in der Mitte der genetische Assoziation von Complement-Faktor H (CFH) mit altersbedingter Makuladegeneration (AMD), der erste der Anfälligkeit für Krankheiten Gen durch genomweiten Assoziations Bildschirm (GWAS) identifiziert. Schließlich führte sie zu dem ersten erfolgreichen Gentherapie zur Leber kongenitale Amaurose, die erste korrigierende Gentherapie-Studie bei menschlichen irgend Erbkrankheit. Die Struktur dieses Organs in den meisten Wirbeltierarten konserviert. Ihre Zugänglichkeit für die Manipulation in vivo hat frühzeitig funktionellen Genomik Untersuchungen auf dieser integraler Bestandteil des Cent gefördertral Nervensystem.
Wir zeigen hier, wie die Photorezeptorschicht von der Innenschicht der Retina durch Vibratom Schnitte trennen. Dieser Schritt ist wichtig, um reine Kulturen Photorezeptor zu erhalten. Unsere Dissektion Protokoll macht die Kontamination durch RPE-Zellen sehr unwahrscheinlich.
Eine der wichtigsten Herausforderungen ist das Abflachen der Netzhaut, die in Abschnitt ordnungsgemäß der inneren und der äußeren Netzhaut notwendig ist. Die Aufteilung der Netzhaut am besten auf der Netzhaut mit kleinem Durchmesser wie die aus Nagern und dieser Durchmesser ist eine Einschränkung der Technik mit derzeitig erhältlichen Material.
Es ist ratsam, vor dem Starten eines biologisch sinn Experiment an einer Probe von der Netzhaut zu praktizieren. Wir zeigen das Verfahren durch aus kultivierten Sehzellen erhalten repräsentative Ergebnisse, unter Verwendung des Materials, um die Expression Studie beider mRNAs und Proteine durchzuführen. Expressionsstudien können auch alternativ auf Sectio durchgeführt werdenns durch die Laser Mikrodissektion erhalten, aber die Kulturen am besten mit Vibratom Schnitte. Wir könnten die Technik der Mikrodissektion Laser mit einer ganz anderen Strategie verwendet haben. Aber, um die Photorezeptorschicht mit der Mikrodissektion Vorrichtung zur Kultur zu sammeln, würde es notwendig sein, Fixiermittel zu vermeiden und dadurch die derzeitige Methodik erheblich erschweren.
Wir entwickeln ein Protokoll auf die Untersuchung der Kinetik der Gen und Proteinexpression in Vibratom Abschnitte während der Degeneration von Photorezeptoren in Modellen der Retinitis pigmentosa gerichtet. Wir glauben, dass die detaillierte Beschreibung des Protokolls wird nützlich für Forscher auf dem Gebiet der Netzhaut-Biologie und ganz besonders für die Proteomik und metabolomic Studien.
Offenlegungen
The authors have nothing to disclose.
Danksagungen
Igal Gery and David Hicks for anti-SAG antibodies. Ram Fridlich for reading the manuscript.
Materialien
| Name | Company | Catalog Number | Comments |
| Cytidine 5′-diphosphocholin* | Sigma-Aldrich | C0256 | 4.7 µM Cytidine 5′-diphosphocholin |
| Cytidine 5′-diphosphoethanolamine* | Sigma-Aldrich | C0456 | 2.7 µM Cytidine 5′-diphosphoethanolamine |
| Linoleïc acid/ bovine serum albumin (BSA)* | Sigma-Aldrich | L8384 | 100 µg/ml linoleïc acid/ bovine serum albumin (BSA) |
| Triiodo-L-thyronine* | Sigma-Aldrich | T6397 | 0.03 µM Triiodo-L-thyronine |
| 96-well plates | Greiner bio-one | 655-095 | |
| Binocular microscope | Leica | MZ-75 | |
| CO2 independent (CO2-i) | Life Technologies | 18045054 | |
| DMEM | Life Technologies | 41966029 | |
| Forceps n°5 Dumont | Bionic | 11254-20 | |
| Gelatin from porcin skin type A | Sigma-Aldrich | G2500 | |
| GeneChip | Affymetrix | U74v2 | |
| Gentamicin solution | Life Technologies | 15710049 | |
| Hydrocortison* | Sigma-Aldrich | H0888 | 0.55 µM hydrocortison |
| Insulin* (I) | Sigma-Aldrich | I1884 (ITS) | 0.86 µM insulin (I) |
| Papain | Worthington-biochem | WOLS03124 | |
| Poly-D-lysine | Sigma-Aldrich | P-6407 | |
| Progesterone* | Sigma-Aldrich | P7556 | 2.0 µM progesterone |
| Prostaglandin* | Sigma-Aldrich | P5172 | 0.28 µM prostaglandin |
| Putrescine* | Sigma-Aldrich | P5780 | 182 µM putrescine |
| Scalpel Albion | EMS | 72000 | |
| Scissor | Moria | 15396-00 | |
| Sodium pyruvate* | Sigma-Aldrich | S8636 | 1 mM sodium pyruvate |
| Sodium selenite* (S) | Sigma-Aldrich | I1884 (ITS) | 0.29 µM Na2SeO3 (S) |
| Taurine* | Sigma-Aldrich | T8691 | 3 mM taurine |
| Transferrin* (T) | Sigma-Aldrich | I1884 (ITS) | 0.07 µM transferrin (T) |
| Vibratome apparatus | Leica | VT1000-S | |
| * Supplements | |||
Referenzen
- The Organization of the Retina and Visual System. Webvision. Kolb, H., Nelson, R., Fernandez, E., Jones, B. , (2014).
- Perez De Sevilla Muller, L., Shelley, J., Weiler, R. Displaced amacrine cells of the mouse retina. J Comp Neurol. 505 (2), 177-189 (2007).
- Mohand-Said, S., et al. Photoreceptor transplants increase host cone survival in the retinal degeneration (rd) mouse. Ophthalmic Res. 29 (5), 290-297 (1997).
- Daiger, S., Sullivan, L., Bowne, S. RetNet, the Retinal Information Network. , The University of Texas Health Science Center. Houston, TX. Available from: https://sph.uth.edu/retnet (2014).
- Mohand-Said, S., Hicks, D., Dreyfus, H., Sahel, J. A. Selective transplantation of rods delays cone loss in a retinitis pigmentosa model. Arch Ophthalmol. 118 (6), 807-811 (2000).
- MacLaren, R. E., et al. Retinal repair by transplantation of photoreceptor precursors. Nature. 444 (7116), 203-207 (2006).
- Yang, Y., et al. Transplantation of photoreceptor and total neural retina preserves cone function in P23H rhodopsin transgenic rat. PLoS One. 5 (10), (2010).
- Mohand-Said, S., et al. Normal retina releases a diffusible factor stimulating cone survival in the retinal degeneration mouse. Proc Natl Acad Sci USA. 95 (14), 8357-8362 (1998).
- Fintz, A. C., et al. Partial characterization of retina-derived cone neuroprotection in two culture models of photoreceptor degeneration. Invest Ophthalmol Vis Sci. 44 (2), 818-825 (2003).
- Cronin, T., Leveillard, T., Sahel, J. A. Retinal degenerations: from cell signaling to cell therapy; pre-clinical and clinical issues. Curr Gene Ther. 7 (2), 121-129 (2007).
- Wright, A. F. A searchlight through the fog. Nat Genet. 17 (2), 132-134 (1997).
- Leveillard, T., et al. Identification and characterization of rod-derived cone viability factor. Nat Genet. 36 (7), 755-759 (2004).
- Lillig, C. H., Holmgren, A. Thioredoxin and related molecules--from biology to health and disease. Antioxid Redox Signal. 9 (1), 25-47 (2007).
- Fridlich, R., et al. The thioredoxin-like protein rod-derived cone viability factor (RdCVFL) interacts with TAU and inhibits its phosphorylation in the retina. Mol Cell Proteomics. 8 (6), 1206-1218 (2009).
- Yang, Y., et al. Functional cone rescue by RdCVF protein in a dominant model of retinitis pigmentosa. Mol Ther. 17 (5), 787-795 (2009).
- Bennett, J. Strategies for delivery of rod-derived cone viability factor. Retina. 25 (8 Suppl), S47(2005).
- Komaromy, A. M., et al. Transient photoreceptor deconstruction by CNTF enhances rAAV-mediated cone functional rescue in late stage CNGB3-achromatopsia. Mol Ther. 21 (6), 1131-1141 (2013).
- Birch, D. G., Weleber, R. G., Duncan, J. L., Jaffe, G. J., Tao, W. Randomized trial of ciliary neurotrophic factor delivered by encapsulated cell intraocular implants for retinitis pigmentosa. Am J Ophthalmol. 156 (2), 283-292 (2013).
- Cronin, T., et al. The disruption of the rod-derived cone viability gene leads to photoreceptor dysfunction and susceptibility to oxidative stress. Cell Death Differ. 17 (7), 1199-1210 (2010).
- Leveillard, T., Sahel, J. A. Rod-derived cone viability factor for treating blinding diseases: from clinic to redox signaling. Sci Transl Med. 2 (26), 26ps16(2010).
- Fontaine, V., Hicks, D., Dreyfus, H. Changes in ganglioside composition of photoreceptors during postnatal maturation of the rat retina. Glycobiology. 8 (2), 183-190 (1998).
- Fontaine, V., Kinkl, N., Sahel, J., Dreyfus, H., Hicks, D. Survival of purified rat photoreceptors in vitro is stimulated directly by fibroblast growth factor-2. J Neurosci. 18 (23), 9662-9672 (1998).
- Reichman, S., et al. The homeobox gene CHX10/VSX2 regulates RdCVF promoter activity in the inner retina. Hum Mol Genet. 19 (2), 250-261 (2010).
- Sandu, C., Hicks, D., Felder-Schmittbuhl, M. P. Rat photoreceptor circadian oscillator strongly relies on lighting conditions. Eur J Neurosci. 34 (3), 507-516 (2011).
- Dorrell, M. I., Aguilar, E., Weber, C., Friedlander, M. Global gene expression analysis of the developing postnatal mouse retina. Invest Ophthalmol Vis Sci. 45 (3), 1009-1019 (2004).
- Brooks, M. J., Rajasimha, H. K., Roger, J. E., Swaroop, A. Next-generation sequencing facilitates quantitative analysis of wild-type and Nrl(-/-) retinal transcriptomes. Mol Vis. 17, 3034-3054 (2011).
- Delyfer, M. N., et al. Transcriptomic analysis of human retinal surgical specimens using jouRNAI. J Vis Exp. (78), (2013).
- Ying, S., et al. A CAT reporter construct containing 277bp GNAT2 promoter and 214bp IRBP enhancer is specifically expressed by cone photoreceptor cells in transgenic mice. Curr Eye Res. 17 (8), 777-782 (1998).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten