
Method Article
Mukociliäre Epithelorganoide aus Xenopus Embryonal cells: Generation, Culture und High-Resolution Live Imaging
In diesem Artikel
Zusammenfassung
Wir beschreiben ein einfaches Protokoll zur Entwicklung von mukociliären Epithelorganoiden aus tiefen Ektodermzellen, die aus Xenopus laevis Embryonen isoliert sind. Die multipotenten Vorläufer regenerieren epitheliale Kelchenzellvorläufer und ermöglichen eine Live-Tracking der Initiation und Progression der Zellübergänge auf der Oberfläche von Organoiden.
Zusammenfassung
Mukociliale Epithel bietet die erste Verteidigungslinie durch entfernen fremde Partikel durch die Wirkung der Schleimproduktion und Zilien-vermittelte Clearance. Viele klinisch relevante Defekte im mukociliären Epithel werden abgeleitet, da sie tief im Körper auftreten. Hier führen wir ein traktierbares 3D-Modell für mukociliäre epithelium ein, die aus multipotenten Vorläufern erzeugt werden, die mikrochirurgisch aus Xenopus laevis Embryonen isoliert wurden. Die mukociliären Epithel-Organoide werden mit neu generiertem Epithel aus tiefen Ektodermzellen bedeckt und später mit ausgeprägten gemusterten multiciliatierten Zellen, sekretorischen Zellen und schleimproduzierenden Kelchzellen verziert, die innerhalb von 24 h nicht von der nativen Epidermis zu unterscheiden sind. Die vollständigen Sequenzen dynamischer Zellübergänge von mesenchymal zu epitheliaal, die auf der apikalen Oberfläche von Organoiden entstehen, können durch hochauflösende Live-Bildgebung verfolgt werden. Diese in vitro kultivierten, selbstorganisierenden mukociliären Epithel-Organoide bieten deutliche Vorteile bei der Untersuchung der Biologie des mukociliären Epithels mit hoher Effizienz in der Erzeugung, definierten Kulturbedingungen, Kontrolle über Anzahl und Größe und direktem Zugang zur Live-Bildgebung während der Regeneration des differenzierten Epithels.
Einleitung
Verletzungen, Infektionen und Erkrankungen des Mukociliaepithels sind mit einer beeinträchtigungen Produktion und Clearance von Schleim verbunden, die häufig bei Lungenerkrankungen wie chronisch obstruktiver Lungenerkrankung, Asthma, Mukoviszidose, Bronchiektase und primärer Ziliardyskinesie1,2,3,4gefunden wird. Als vielversprechendes Modell mit therapeutischem Potenzial1,5,6entsteht ein neuer Fortschritt in der Organoidtechnologie, das von der Basalzelle abgeleitet ist, die Tracheosphäre genannt wird, die die Regeneration des mukociliären Epithels rekapituliert. Allerdings ist seine Verwendung derzeit begrenzt, zum Teil aufgrund des Mangels an definierten Kulturbedingungen und der geringen Effizienz in organoiden Produktionen. Mukociliale Epithel in der menschlichen Atemwege und Frosch Epidermis sind bemerkenswert ähnlich in Gewebemorphologie, zelluläre Zusammensetzung, und seine Funktion7,8,9,10,11,12. In beiden Organismen bietet das mukoziliarische Epithel eine Erstlinienverteidigung durch Sekretierung von Schleim und antimikrobiellen Substanzen und klärt schädliche Partikel und Krankheitserreger durch die synchronisierte Wirkung von Zilien.
Hier beschreiben wir ein einfaches Protokoll zur Erzeugung von mukociliären Epithelorganoiden mit den multipotenten Vorläufern von Xenopus laevis Embryonen13,14. Zuvor berichteten wir14, dass sich in Ermangelung exogener Wachstumsfaktoren und der extrazellulären Matrix die mikrochirurgisch isolierten tiefen Zellen aus dem frühen Gastorulastadium ektoderm spontan zu Aggregaten zusammenfügen, Epithel auf seiner Oberfläche regenerieren und durch Interkalieren von multiciliated und anderen Zubehörzellen innerhalb von 24 h zu mukociliärem Epithel reifen. Neben der rasanten Entwicklung bietet dieses Protokoll eine deutliche Möglichkeit für den direkten Zugriff auf die Übergänge von multipotenten tiefen Ektodermzellen in epitheliale Kelchenzellvorläufer, die die Regenerationsschritte eines gestörten Epithels14 rekapitulieren, die nicht von intakten Embryonen und Ektoderm (auch bekannt als Tierkappe)15,16,17. Die Anzahl und Größe der produzierten Organoide sind mit hoher Effizienz skalierbar, indem die Ausgangsmaterialien von Xenopus-Embryonen gesteuert werden. Organoide in der schwimmenden Kultur können einfach sortiert und im gewünschten Stadium für weitere Analysen übertragen werden, einschließlich hochauflösender Bildgebung, mechanischer Tests, medikamentöser Behandlung und genetischer Charakterisierung14. Diese spontane, gewebemechanikgetriebene Regeneration des Epithels auf der Oberfläche embryonaler Zellaggregate führt zu mukociliären Epithelorganoiden und liefert ein neuartiges dreidimensionales (3D) Modell, um die Biologie des mukociliären Epithels zu untersuchen.
Protokoll
Tierverwendungs- und Versuchsprotokolle wurden vom institutionellen Ausschuss für Tierpflege und -nutzung (IACUC) des Institute for Basic Science (IBS 18-01) und des Korea Advanced Institute of Science and Technology (KA2017-22) genehmigt.
1. Embryonen
- Erhalten Sie X. laevis Embryonen nach einem Standardverfahren: sammeln Sie manuell Eier von stimulierten weiblichen Fröschen und führen Sie in vitro Fertilisation18,19.
- Entgelee die befruchteten Embryonen mit sanfter Erregung für ca. 5 min in 2% Cystein in 1/3x modifizierter Barth-Kochsaline (MBS; siehe Rezept für 1X MBS unten) bei pH 819.
- Optionaler Schritt für live-Imaging: um bestimmte Proteine fluoreszierend zu kennzeichnen und deren Dynamik im Organoid zu beobachten, fahren Sie mit Abschnitt 5 fort.
- (Optional) Um die Kontamination oberflächlicher Ektodermzellen innerhalb von Organoiden zu überwachen, beschriften Sie die apikale Oberfläche der Embryonen mit NHS-Rhodamine im Stadium 914. Inkubieren Sie Embryonen in 1 mg/ml NHS-Rhodamine in 1/3x MBS (pH 9.0) für 30 min mit sanfter Nutation. Waschen Sie Embryonen dreimal, indem Sie 15 min auf eine Petrischale übertragen, die mit 1/3x MBS gefüllt ist.
- Den Embryo in 1/3x MBS bei der bevorzugten Temperatur (14–26 °C) kulturieren, bis die ersten Anzeichen von Stadium 10 nachgewiesen werden (d. h. das Auftreten von dunkelpigmentierten Zellen um das Blastopor in der Gemüseansicht).
2. Herstellung von mikrochirurgischen Werkzeugen, Lösungen und Kulturgefäßen
- Bereiten Sie die Werkzeuge benötigt, einschließlich eines Paares von chirurgischen Grade Zangen und Haarwerkzeuge (Haarschlaufe und Haarmesser) für die Mikrochirurgie20.
- Bereiten Sie die folgenden Kulturmedien für Embryonen vor: 1/3X MBS, wobei 1X MBS mit NaCl (88 mM), KCl (1 mM), NaHCO3 (2,4 mM), MgSO4 (0,82 mM), Ca(NO3)2 (0,33 mM), CaCl2 (0,41 mM) und HEPES (10 mM) hergestellt wird. PH auf 7,4 mit 10 M NaOH einstellen.
HINWEIS: Optional: Tropfen Phenolrot hinzufügen, um den pH-Wert anzuzeigen. - Bereiten Sie die folgenden Kulturmedien für embryonale Gewebe und Organoide vor: Danilchik es for Amy (DFA)21 ergänzt mit frischen 1% Antibiotikum und Antimykotiklösung. Bereiten Sie DFA mit NaCl (53 mM), Na2CO3 (5 mM), Kaliumgluconat (4,5 mM), Natriumgluconat (32 mM), CaCl2 (1 mM) und MgSO4 (1 mM) vor. Stellen Sie den pH-Wert mit körnigem Bicine auf 8,3 ein. Filtern Sie DFA (0,2 m Flaschen-Top-Filter), aliquot und lagern Sie es bei -20 °C.
- Bereiten Sie kalzium- und magnesiumfreie DFA zur Trennung von tiefen Zellen vom Ektoderm vor, indem Sie das obige Rezept verwenden und CaCl2 und MgSO4auslassen. Aliquot und lagern bei-20 °C.
- Bereiten Sie nicht klebende PCR-Röhren für die embryonale Zellaggregation vor.
- Um eine spontane Aggregation der isolierten embryonalen Zellen zu induzieren, bereiten Sie nicht klebende PCR-Röhren vor, indem Sie runde PCR-Röhren mit 200 l von 1% BSA (1 g BSA in 100 ml destilliertem Wasser) über Nacht bei 4 °C oder für 2 h bei Raumtemperatur beschichten. Jedes Rohr wird verwendet, um ein Organoid zu montieren.
- Spülen Sie BSA-beschichtete PCR-Rohre mit DFA dreimal, um restliche BSA zu entfernen.
- Füllen Sie PCR-Röhren mit 200 l DFA.
3. Isolierung von tiefen Ektodermzellen
- Wählen und sammeln Sie Embryonen, wie sie früh Stufe 10 mit Haarwerkzeugen unter einem Stereoskop erreichen.
- Mit einer Einweg-Transferpipette die ausgewählten Embryonen in eine MIT DFA gefüllte Petrischale übertragen.
- Entfernen Sie die Vitelline-Membran der Embryonen mit scharfen Zangen von der gemüseseitigen Seite, ohne die tierische Seite des Embryos zu stören.
HINWEIS: Achten Sie darauf, Embryonen nicht der Luft auszusetzen. Das Einbringen von Luftblasen in die Lösung oder das Einbringen von Embryonen an die Oberfläche führt dazu, dass der Embryo platzt. - Um die Tierkappe zu isolieren, positionieren Sie die tierische Seite des Embryos nach oben.
- Schätzen Sie visuell das Ausmaß der zu verbrauchenden Tierkappe und machen Sie den ersten Schnitt entlang der Kante der Tierkappe mit einem Haarmesser. Ziehen Sie das Haarmesser nach außen, um einen Schnitt zu machen.
- Wiederholen Sie Schritt 3.5, um eine Kette von kleinen Schnitten zu erstellen, um die Tierkappe zu verbrauchen.
- Trimmen Sie die dickschichtige Kante der Tierkappe mit einem Haarmesser, um die Aufnahme von Mesoderm-Vorläufern zu verhindern.
HINWEIS: Um die Heilung und Aggregation isolierter Tierkappen zu verhindern, fahren Sie innerhalb von 10 min mit dem nächsten Schritt fort. Typischerweise isolieren wir 5–10 Tierkappen gleichzeitig, um mehrere Organoide zu montieren. - Um tiefe Ektodermzellen von der Tierkappe zu trennen, übertragen Sie die ausgeschnittenen Tierkappen auf eine Mitschnittschale, die mit kalzium- und magnesiumfreiem DFA gefüllt ist, mit einer Einweg-Transferpipette. Achten Sie darauf, während des Transfers keine Luftblasen einzuführen.
- Um genügend Platz für die nächsten Schritte zu halten, positionieren Sie mit den Haarwerkzeugen die Tierkappen, um der Tierseite zu begegnen und einen großzügigen Abstand zu anderen Explants zu wahren.
- Warten Sie 5–10 min und überwachen Sie dann die Explanten unter einem Stereoskop. Sobald gelockerte tiefe Zellen vom Rand der dunkelpigmentierten oberflächlichen Schicht gefunden wurden, beginnen Sie, die oberflächliche Schicht mit einem Haarmesser unter dem Stereoskop von den hellen tiefen Ektodermzellen wegzuheben.
- Die oberflächliche Schicht vorsichtig mit einem Haarmesser ablösen, beginnend am Rand.
- Sammeln Sie die tiefen Ektodermzellen mit minimalem Aspirationsgrad (10-u201215-L), um die Menge an kalzium- und magnesiumfreiem DFA zu begrenzen, die im nächsten Schritt an die Aggregationsmedien übertragen wird.
HINWEIS: Abgetrennte oberflächliche Zellen können aus dem Medium entfernt werden, um eine Kontamination der verbleibenden tiefen Ektodermzellen zu verhindern.
4. Erzeugung von mukociliären Epithelorganoiden
- Übertragen Sie gesammelte tiefe Ektodermzellen auf ein nicht klebendes PCR-Rohr, das 200 l DFA enthält. Pipette das Medium (2-u20123 mal) vorsichtig, um übertragene Zellen in der PCR-Röhre zu dispergieren.
HINWEIS: Das Timing, das als Stunden nach der Aggregation (hpa) bezeichnet wird, beginnt in diesem Schritt. Die Größe der resultierenden Organoide wird durch die Anzahl der tiefen Ektodermzellen gesteuert, die einer PCR-Röhre hinzugefügt werden. Tiefe Ektodermzellen aus einer oder mehreren Tierkappen können je nach gewünschter Größe des Organoids verwendet werden. - Schließen Sie die PCR-Röhre. Halten Sie PCR-Röhren aufrecht, um spontane Aggregation an der Unterseite zu induzieren.
- Überwachen Sie den Aggregationsprozess unter einem Stereoskop. Zellen sammeln sich in der Regel innerhalb einer Stunde am unteren Rand des PCR-Rohrs und fügen sich je nach Größe innerhalb von 2–3 h zu kugelförmigen Aggregaten zusammen.
- Um Live-Bildgebung oder Arzneimitteltests während der Entwicklung der mukociliären Epithel-Organoide durchzuführen, sammeln Sie Aggregate bei 2 PSa mit einer 200-L-Pipette, die mit vergrößerten Spitzen (mit sterilisierter Schere geschnitten) ausgestattet ist, um Schäden an den Aggregaten während der Entnahme zu vermeiden.
- Damit sich Aggregate in der Kultur zum mukociliären Epithelorganoid entwickeln können, sammeln Sie kugelförmige Aggregate aus dem PCR-Rohr bei 5 PSa und übertragen sie in eine DFA-gefüllte Petrischale.
- Positionieren Sie die Aggregate weit von anderen entfernt, um zu verhindern, dass sie verschmelzen. Innerhalb von 24 h Kultur bei Raumtemperatur ohne zusätzliche Faktoren können reife mukociliäre Epithelorganoide beobachtet werden, die sich mit der Wirkung von schlagenden Zilien drehen, die die Oberfläche des differenzierten Epithels abdecken.
5. (Optional) Hochauflösende Live-Bildgebung der Entwicklung von Organoiden
- Bereiten Sie mRNA für die Mikroinjektion vor.
- Um die Epithelisierung zu visualisieren, die im Anfangsstadium der Organoidbildung auftritt, bereiten Sie mRNA für die epithelialenspezifischen Zonula-Okkludensprotein-1 (ZO-1) und für die Darstellung der Zellmembranen durch Amplizierung von pCS2-ZO1-RFP und pCS2-mem-GFP Plasmiden (ein Geschenk von Davidson Davidson) vor.
- Extrahieren und linearisieren Sie die Plasmid-DNA, und transkribieren Sie dann die gekappte mRNA mit einem SP6/T7 In-vitro-Transkriptionskit.
- Aliquot die transkribierte mRNA und lagern Sie sie bei -80 °C.
- Mikroinjizieren Sie die mRNA in einen befruchteten Embryo
- Befruchtete Embryonen in 3% Ficoll in 1x MBS geben.
- Laden Sie 3–4 l mRNA mit einer Mikroladerspitze in eine gezogene Glasnadel (eine lange und feine verjüngte Nadelspitze mit einem Innendurchmesser von 10 bis 201230 m).
- Befestigen Sie die Nadel an einem Mikroinjektor und passen Sie die Zeit und den Druck an, um ein konstantes Volumen an mRNA für die Mikroinjektion zu liefern.
- Injizieren Sie die mRNA direkt unter der apikalen Oberfläche des Tierpols. Ein deutlich blassgefärbter kreisförmiger Fleck, der durch die Ausdehnung des Kortex verursacht wird, ist zum Zeitpunkt der Mikroinjektion sichtbar.
- Übertragen Sie die injizierten Embryonen auf 1/3X MBS und kultivieren Sie sie auf Stufe 9.5.
- Sammeln Sie die fluoreszierend markierten Embryonen unter einem Fluoreszenz-Steroskop (Erregungs-/Emissionseinstellungen für GFP (488/510) und RFP (532/588)).
- Fahren Sie mit Schritt 1.3 fort.
- Montieren und kulturieren Sie das Organoid (Abschnitte 3 und 4) bis zum gewünschten Entwicklungsstadium.
- Führen Sie Live-Bildgebung durch.
- Bereiten Sie eine Glasboden-Bildgebungskammer vor, indem Sie ein Deckglas mit Siliziumfett in eine speziell gefräste Acrylkammer kleben.
HINWEIS: Versiegeln Sie die Kammer fest, um ein Auslaufen der Kulturmedien zu verhindern. - Füllen Sie die Bildgebungskammer mit DFA.
- Wählen Sie ein sechseckiges Transmissionselektronenmikroskopie (TEM) Raster (75 Mesh) aus einem Behälter mit Zangen und wenden Sie eine winzige Menge Fett an den Rand des Gitters.
HINWEIS: Die Größe des Netzes sollte kleiner als der Durchmesser des Aggregats sein, damit das Aggregat auf dem Raster sitzt. - Drücken Sie leicht nach unten, um das TEM-Gitter an der Unterseite der Bildkammer zu befestigen.
- Übertragen Sie die Aggregate in die Bildkammer und positionieren Sie sie innerhalb des Rasters.
HINWEIS: Vermeiden Sie die Positionierung der Aggregate neben dem Fett. Während des gesamten Experiments sollten die Aggregate auf dem TEM-Gitter sitzen, ohne die Unterseite der Kammer zu kontaktieren, um eine physische Kompression zu verhindern. - Füllen Sie die Bildgebungskammer mit DFA und versiegeln Sie sie mit einem Deckglas und Fett.
HINWEIS: Die Kammer sollte luftdicht und ohne Luftblasen sein, um Turbulenzen oder Bewegungen während der Bildgebung zu verhindern. - Um das Fortschreiten der mukociliären epithelialen Organoidbildung zu verfolgen, sammeln Sie Zeitraffer-Z-Stack-Bilder von Aggregaten (von 2 PSa) mit einem konfokalen Mikroskop.
HINWEIS: Wir sammeln in der Regel alle 15 min 120-m-dicke Z-Stacks mit einem 20-fachen Objektiv, um das dynamische Zellverhalten zu verfolgen, aber diese Spezifikationen sollten für das Ziel von Experimenten optimiert werden.
- Bereiten Sie eine Glasboden-Bildgebungskammer vor, indem Sie ein Deckglas mit Siliziumfett in eine speziell gefräste Acrylkammer kleben.
6. (Optional) Bildgebung entwicklung Organoide durch Fixierung und Immunfärbung
- Fixieren Sie Organoide im gewünschten Entwicklungsstadium, indem Sie sie in eine Glasflasche übertragen, die mit einer fixativen Lösung gefüllt ist.
HINWEIS: Fügen Sie ein Volumen fixativer Lösungen hinzu, das >20-mal so hoch ist wie das der Samples, um eine vollständige Fixierung sicherzustellen. Führen Sie die folgenden Prozesse auf einem Nutator durch, sofern nicht anders angegeben. Im Allgemeinen werden Organoide mit 4% Paraformaldehyd (PFA) in PBS fixiert. Es können jedoch verschiedene Fixative erforderlich sein, um bestimmte Proteine zu erkennen. Zum Beispiel verwendeten wir 4% PFA mit 0,25% Glutaraldehyd in PBS, um F-Actin und acetyliertes Tubulin zu erkennen. Um Intelectin (ITLN) und ZO-1 zu erkennen, werden Organoide mit eiskalter Dent-Lösung (4:1 Methanol:Dimethylsulfoxid) für die Nacht bei -20 °C fixiert. Die festen Organoide von Dent sollten vor dem Waschen seriell dehydriert werden (Schritt 6.3). Die Dauer der Antikörperinkubation und des Waschens kann für spezifische Bedürfnisse optimiert werden. - Organoide für 15 min bei Raumtemperatur (RT) oder über Nacht bei 4 °C fixieren.
- Waschen Sie 3 mal mit PBST (PBS mit 0,1% Triton X-100) für 15 min bei RT.
- Blockieren Sie unspezifische Bindung mit 10% Ziegenserum in PBST (PBSGT) für 1 h bei RT.
- Inkubieren Sie mit primärem Antikörper (1:200) in PBSGT über Nacht bei 4 °C.
- Waschen Sie 3 mal mit PBST für 15 min bei RT.
- Inkubieren Sie mit Sekundärantikörper (1:200) in PBSGT über Nacht bei 4 °C.
- Waschen Sie 3 mal mit PBST für 15 min bei RT.
- Übertragen Sie die fixierten und immunbefleckten Organoide in eine Bildkammer und führen Sie eine konfokale Bildgebung durch.
Ergebnisse
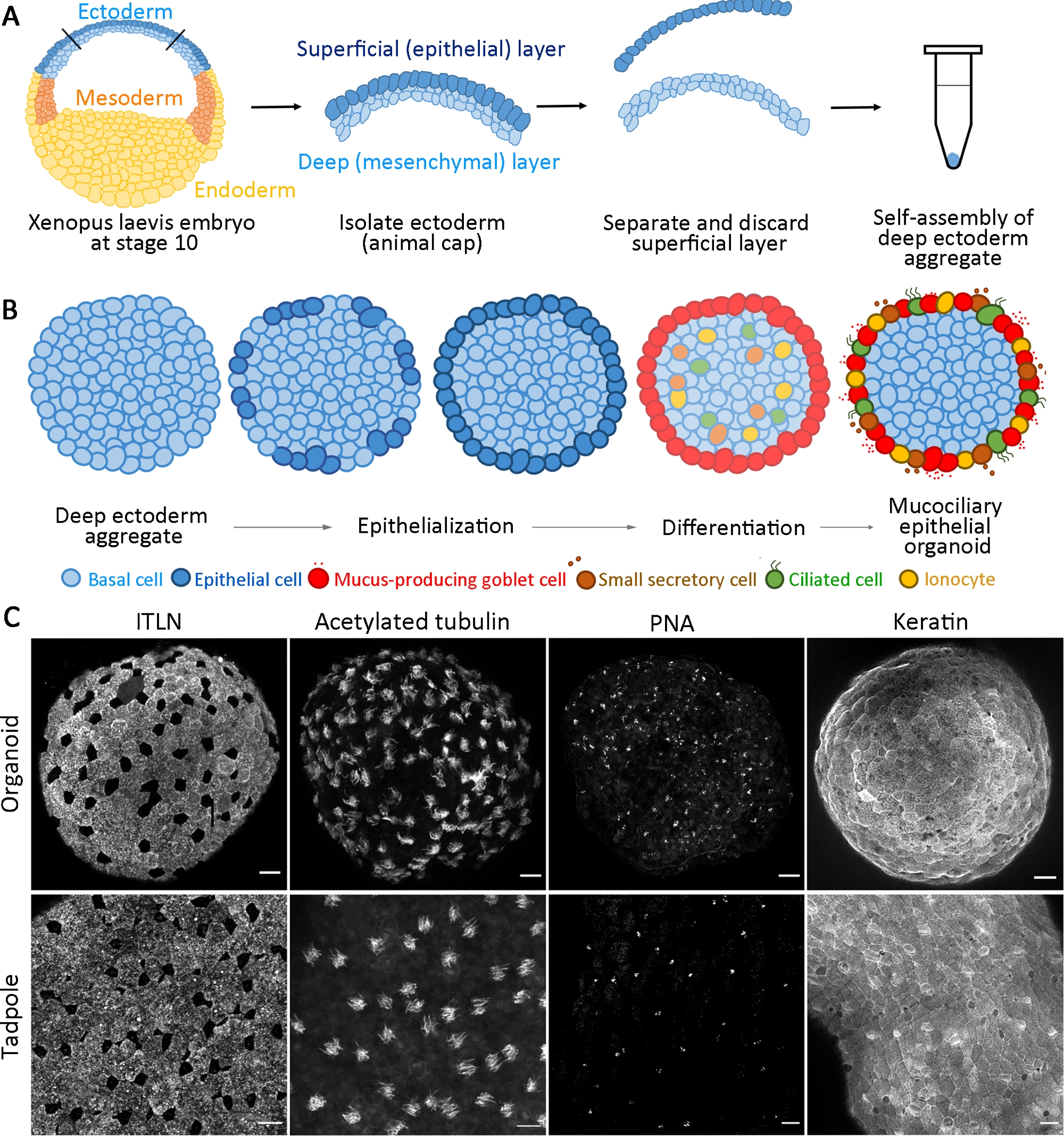
Dieses standardisierte Protokoll erzeugt ein mukociliäres Epithelorganoid aus multipotenten Vorläufern, die ab dem frühen Gastorulastadium X isoliert sind. laevis-Embryonen innerhalb von 24 h der Kultivierung14. Gesammelte tiefe Ektodermzellen bauen sich selbst zu einem Aggregat in einem nicht klebenden PCR-Rohr zusammen und durchlaufen Oberflächenepithelisierung und Kelchzelldifferenzierung. Die neu epithelisierte Oberfläche von Aggregaten bietet ein Substrat ähnlich dem nativen Epithel, das in vivo für interkalierende innere Zellen (z.B. multiciliated und andere Zubehörzellen) gefunden wurde und entwickelt sich zu mukociliären Epithelorganoiden(Abbildung 1A,B). Innerhalb von 24 h nach der Aggregation regenerieren selbstorganisierte mukociliäre Epithel-Organoide eine reife Epidermis, die von der Epidermis einer Kaulquappe nicht zu unterscheiden ist. Die Organoide bestehen aus vollständig differenziertem Epithel (Keratin), schleimsekretsierenden Kelchenzellen (ITLN), multiciliatierten Zellen (acetyliertes Tubulin) und kleinen sekretoreichen Zellen (Erdnussagglutinin, PNA) (Abbildung 1C).
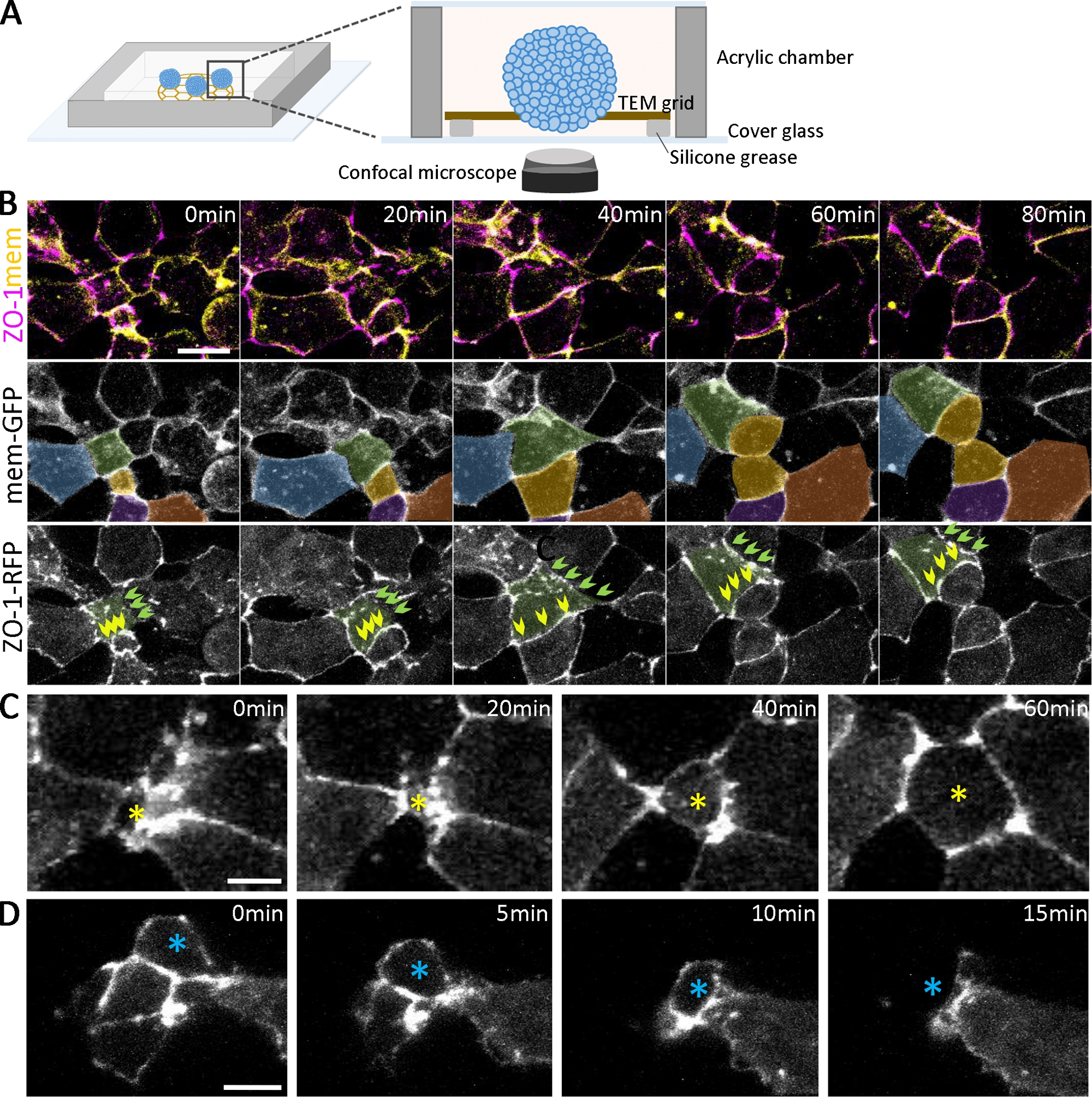
Neben der Bestätigung der Entwicklung verschiedener Zelltypen mit Immunostainierung kann der Dynamik der Organoidentwicklung auch eine Live-Bildgebung folgen (Abbildung 2A). Um die Epithelisierung zu untersuchen, die im frühen Stadium der Organoidbildung entsteht (Abbildung 1B), haben wir Embryonen markiert, indem wir fluoreszierend getaggte enge Knotenproteine (ZO-1-RFP) und Membranlokalisierungsproteine (mem-GFP) exizierten. Bei der Dual-Labeling können die sequenziellen Schritte der ZO-1-positiven engen Knotenbildung während der Epithelisierung markiert und quantitativ analysiert werden (Abbildung 2). Für Zellen(Abbildung 2B, grün gefärbt) in verschiedenen Stadien der Epithelisierung (bei 0 min) haben einige Regionen der Zellzelladhäsion verstreute Punktia von ZO-1(Abbildung 2B, grüne Pfeile). Im Gegensatz dazu haben andere Bereiche einen vollständig montierten, zusammenhängenden ZO-1-Ausdruck(Abbildung 2B, gelbe Pfeile). Im Laufe der Zeit verschmelzen die Puncta und verbinden sich zu zusammenhängenden engen Knoten(Abbildung 2B,grüne Pfeile), und zusammenhängende enge Knoten behalten ihre Morphologie auch während der Zellteilung bei(Abbildung 2B,gelbe Pfeile). Wenn die engen Knoten reifen, bewegen sich Zellen dynamisch in und aus der Oberfläche entlang der apikalen Ebenen der Organoide (Abbildung 2C,D). Darüber hinaus ist durch die Verfolgung von Zellen, die auf der Oberfläche differenzierender Organoide(Abbildung 2B, farbkodierte Zellen) räumlich begrenzt sind, eine Multiskalenanalyse möglich, die von einzelnen Punktzeichen bis hin zu zusammenhängenden engen Knoten, Zell-Zell-Grenzen und Teilmengen von Zellpopulationen innerhalb von Organoiden reicht.

Abbildung 1: Erzeugung von mukociliären Epithelorganoiden.
(A) Ein Schaltplan, der das Protokoll zur Zusammenstellung tiefer Ektodermaggregate aus X. laevis-Embryonen zeigt. (B) Ein Schema für ein Modell der mukociliären epitheliale Organoidbildung, die aus multipotenten tiefen Ektodermzellen (Querschnittsansicht) stammt. Oberflächenpositionierte Zellen werden in Epithelzellen übertragen und in Kelchenzellen differenziert. Die Differenzierung von Zilienzellen, Sekretorienzellen und Ionozyten intercalate radial in die Oberfläche und regenerieren eine reife Epidermis. (C) Maximale z-Projektion von mukociliären Epithel, immunostainiert für ITLN (Schleim produzierende Kelchzellen), acetyliertes Tubulin (Zilienzellen), PNA (kleine sekretorische Zellen) und Keratin (Epithelzellen) bei Organoiden bei 24 PSa (obere Platte) und Kaulquadermik (unteres Panel). Skalenbalken = 30 m. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Live-Bildgebung der Entwicklung von Organoiden.
(A) Ein Schaltplan der Bildkammer für lebende Organoide (nicht zu skalieren). (B) Zeitraffersequenzen von konfokalen Stapeln, die aus tiefen Ektodermzellaggregaten gesammelt werden, die ZO-1-RFP und Mem-GFP aus 2,5 psa exdrücken. Skalenbalken = 20 m. Zellen sind pseudofarben für die Verfolgung im Laufe der Zeit. Grün gefärbte Zellen haben unterschiedliche Zellhaftungsstatus, einschließlich einer sich zunehmend entwickelnden ZO-1-positiven Haftung (grüne Pfeile) und einer, die zusammenhängende ZO-1-positive Haftung (gelbe Pfeile) im Laufe der Zeit aufrechterhält. (C, D) Zeitraffer-Konfokalbilder von ZO-1-RFP-exezierenden tiefen Ektodermzellenaggregaten zeigen die radial interkalierenden Zellen, die sich an die Oberfläche bewegen (C, gelber Stern) und sich innerhalb der Aggregate (D, blauer Stern) bewegen. Skalenbalken = 10 m. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Mukociliäre Epithelorganoide, die aus tiefen Ekodermzellen des X. laevis Embryos erzeugt werden, sind ein leistungsfähiges Werkzeug, um die Epithelisierung und Differenzierung multipotenter Vorläufer in vitro zu untersuchen. Im Gegensatz zum weit verbreiteten Tierkappentest16, der für die In-vitro-Organogenese13 verwendet wird, und der Entwicklung von mukociliären Epithelien15,17,22, die das intakte Ektoderm nutzen, bieten die in diesem Protokoll vorgestellten tiefen ekodermabgeleiteten Organoide eine deutliche Gelegenheit, die gewebemechanikgetriebenen Regenerationsphasen des Oberflächenepithels14zu überwachen. Bei etwa 2 PSa beginnen die neu generierten ZO-1-positiven Epithelzellen (Abbildung 2) auf der apikalen Oberfläche von Organoiden zu erscheinen und ihre Population zu erhöhen, um das gesamte Organoid zu bedecken, da das Gewebe die Compliance verfestigt oder reduziert14. Die Regeneration des Epithels und die nachfolgenden Abstammungsspezifikationen für Schleim produzierende Kelchenzellen verlaufen spontan innerhalb eines Tages in einem chemisch definierten Kulturmedium. Diese sich schnell entwickelnden mukociliären Epithelorganoide bieten eine Plattform, um dynamischezellverhaltene Zellen in Echtzeit, in hoher Auflösung, während progressiver Schritte der epitheliaalen Regeneration zu untersuchen. Sie ermöglichen auch die Untersuchung grundlegender Fragen, die während der Entwicklung des mukociliären Epithels, der Homöostase und der damit verbundenen Krankheiten2,9,23auftreten. Insbesondere die mechanische Empfindlichkeit der tiefen Vorläuferzellen während des Übergangs zu epithelialer Kelchenzellvorläufern, die in den Organoiden14 identifiziert wurden, kann dazu dienen, Atemwegserkrankungen im Zusammenhang mit einer abnormalen Basaldifferenzierung zu verknüpfen, bei der Schleimsekretion von Kelchenzellen über- oder unterproduziert sind23.
Während dieses Protokoll einen einfachen Ansatz bietet, um diese Organoide zu erzeugen, gibt es mehrere kritische Schritte für den Erfolg in Experimenten. Um eine Kontamination oberflächlicher Epithelzellen während der Isolierung von tiefen Ektodermzellen von der Tierkappe zu verhindern, sollte man die in Kalzium- und Magnesiumfreie DFA unter einem Stereoskop platzierte Tierkappe überwachen und den richtigen Zeitpunkt erkennen, um die Trennung der dunkelpigmentierten oberflächlichen Schicht der Tierkappe einzuleiten. Wenn das Gewebe zu lange in kalzium- und magnesiumfreiem DFA gehalten wird, wird sich das gesamte Gewebe dissoziieren und eine Unterscheidung zwischen tiefen und oberflächlichen Zellen wäre für tiefe Ektodermaggregate unmöglich. Um das Fehlen oberflächlicher Zellen in tiefen Ektodermaggregaten zu bestätigen, empfehlen wir, die apikale Oberfläche des Embryos vor der Mikrochirurgie fluoreszierend mit NHS-Rhodamine (Schritt 1.414) zu kennzeichnen; dies würde eine einfache Identifizierung von Oberflächenzellen ermöglichen, wenn sie in den resultierenden Organoiden vorhanden sind. Da die epitheliale Regeneration durch die Gewebemechanik14reguliert wird, ist es wichtig, unbeabsichtigte Krafterzeugung für selbstorganisierende Organoide zu vermeiden. Insbesondere empfehlen wir, den Kontakt mit dem Glasboden der Bildgebungskammer während der Live-Bildgebung zu vermeiden, indem Aggregate an den Rändern der TEM-Gitter platziert werden, da dies einen freien Kontakt mit dem Bildfenster von Live-Aggregaten ermöglicht (Schritt 5.1.2.). Dieses in vitrokultivierte,selbstorganisierte 3D-Modell für mukociliäres Epithel wird als traktierbares Werkzeug dienen, um die grundlegenden Fragen zu beantworten, die sich bei der Regeneration von Epithel und der Abstammungsspezifikation von Kelchzellen stellen.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Wir danken den Mitgliedern von Kim lab und Lance Davidson für ihre Kommentare und Unterstützung. Diese Arbeit wurde vom Young Scientist Fellowship to HYK vom Institute for Basic Science (IBS-R0250Y1) unterstützt.
Materialien
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Dual-stage Glass Micropipette Puller | Narishige | PC-100 | |
| Picoliter microinjector | Warner Instruments | PLI-100A | |
| Confocal Laser Microscope | |||
| Stereoscope | |||
| Tools | |||
| Forcep | Dumont | Dumont #5 | |
| Hair knife | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| Hair loop | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| hCG injection | |||
| human chorionic gonadotropin | Sigma | cg10-10vl | |
| MBS solution | |||
| 10M Sodium hydroxide | Sigma | 72068 | |
| Calcium chloride | Sigma | C3881 | |
| Calcium nitrate | Sigma | C1396 | |
| HEPES | Sigma | H4034 | |
| Magnesium sulfate | Sigma | 230391 | |
| Phenol-red | Sigma | P0290 | |
| Potassium chloride | Sigma | 7447-40-7 | |
| Sodium bicarbonate | Sigma | S6014 | |
| Sodium chloride | Sigma | S9625 | |
| Sodium hydroxide reagent grade, 97%, powder-25g | Sigma | 655104 | |
| dejellying solution | |||
| L-Cysteine hydrochloride monohydrate | Sigma | C7880 | |
| Sodium hydroxide 10M | Sigma | 72068 | |
| Ficoll solution | |||
| Ficoll | Sigma | F4375 | |
| DFA solution | |||
| Sodium chloride | Sigma | S9625 | |
| 0.22mm Filter | Millipore | S2GPT05RE | |
| Antibiotic Antimycotic Solution | Sigma | A5955 | |
| Bicine | Sigma | B3876 | |
| Calcium chloride | Sigma | C3881 | |
| Magnesium sulfate | Sigma | 230391 | |
| Potassium gluconate | Sigma | G4500 | |
| Sodium carbonate | Sigma | 222321 | |
| Sodium gluconate | Sigma | G9005 | |
| mRNA in vitro transcription | |||
| SP6/T7 in vitro transcription kit | Invitrogen | AM1340 | |
| mRNA microinjection | |||
| Borosilicate glass capillary tubes | Harvard Apparatus | GC100-10 | |
| Eppendorf microloader pipette tips | ThermoFisher | A25547 | |
| Mineral oil | Sigma | M5904 | |
| PCR tube coating | |||
| BSA | Thermofisher | 26140079 | |
| PCR tubes | SSI | SSI-3245-00 | |
| Imaging | |||
| Custom-milled acrylic chamber | |||
| Coverglass 24mmX50mm | Duran | B01_001650 | |
| SPI Hexagonal TEM Grids, Gilded Nickel (50mesh) | SPI | 275HGN-XA | |
| SPI Hexagonal TEM Grids, Gilded Nickel (75mesh) | SPI | 2775GN-XA | |
| Silicone grease | Shinetsu | HIVAC-G | |
| Fixation | |||
| 20ml screw top-cap vial | Wheaton | WH.986580 | |
| 2ml screw top-cap vial | |||
| Benzyl alcohol | Sigma | 305197 | |
| Benzyl benzoate | Sigma | B6630 | |
| Dimethyl sulfoxide (DMSO) | Sgima | D4540 | |
| Glutaraldehyde 10% EM GRADE | Electron Microscopy Sciences | 16120 | |
| Goat serum | Jackson | 005-000-121 | |
| Methanol | Sigma | 322415 | |
| Paraforlamdehyde | Sigma | P6148 | |
| Phosphate-buffered saline (PBS) | LPS Solution | CBP007B | |
| Triton X-100 | Sigma | T8787 | |
| Primary antibody (1:200) | |||
| acetylated tubulin | Sigma | clone 6-11B-1 | |
| Itln1 | Proteintech | 11770-1-AP | |
| Keratin | Developmental Studies Hybridoma Bank | 1h5 | |
| ZO1 | Invitrogen | 402200 | |
| Vectors | |||
| pCS2-mem-GFP | Gift from Dr. Lance Davidson | ||
| pCS2-ZO1-RFP | Gift from Dr. Lance Davidson |
Referenzen
- Barkauskas, C. E., et al. Lung organoids: current uses and future promise. Development. 144 (6), 986-997 (2017).
- Puchelle, E., Zahm, J. M., Tournier, J. M., Coraux, C. Airway Epithelial Repair, Regeneration, and Remodeling after Injury in Chronic Obstructive Pulmonary Disease. Proceedings of the American Thoracic Society. 3 (8), 726-733 (2006).
- Tilley, A. E., Walters, M. S., Shaykhiev, R., Crystal, R. G. Cilia dysfunction in lung disease. Annual Review of Physiology. 77, 379-406 (2015).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clinical Microbiology Reviews. 24 (1), 210-229 (2011).
- Rock, J. R., et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12771-12775 (2009).
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300(2019).
- Dubaissi, E., et al. A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development. 141 (7), 1514-1525 (2014).
- Hayes, J. M., et al. Identification of novel ciliogenesis factors using a new in vivo model for mucociliary epithelial development. Developmental Biology. 312 (1), 115-130 (2007).
- Walentek, P., Quigley, I. K. What we can learn from a tadpole about ciliopathies and airway diseases: Using systems biology in Xenopus to study cilia and mucociliary epithelia. Genesis. 55 (1-2), (2017).
- Werner, M. E., Mitchell, B. J. Understanding ciliated epithelia: The power of Xenopus. Genesis. 50 (3), 176-185 (2012).
- Dubaissi, E., Papalopulu, N. Embryonic frog epidermis: a model for the study of cell-cell interactions in the development of mucociliary disease. Disease Models & Mechanisms. 4 (2), 179-192 (2011).
- Walentek, P., et al. A novel serotonin-secreting cell type regulates ciliary motility in the mucociliary epidermis of Xenopus tadpoles. Development. 141 (7), 1526-1533 (2014).
- Asashima, M., et al. In vitro organogenesis from undifferentiated cells in Xenopus. Developmental Dynamics. 238 (6), 1309-1320 (2009).
- Kim, H. Y., Jackson, T. R., Stuckenholz, C., Davidson, L. A. Tissue mechanics drives regeneration of a mucociliated epidermis on the surface of Xenopus embryonic aggregates. Nature Communications. 11 (1), 665(2020).
- Haas, M., et al. DeltaN-Tp63 Mediates Wnt/beta-Catenin-Induced Inhibition of Differentiation in Basal Stem Cells of Mucociliary Epithelia. Cell Reports. 28 (13), 3338-3352 (2019).
- Green, J. Molecular Methods in Developmental Biology: Xenopus and Zebrafish. Guille, M. , Humana Press. 1-13 (1999).
- Stubbs, J. L., Davidson, L., Keller, R., Kintner, C. Radial intercalation of ciliated cells during Xenopus skin development. Development. 133 (13), 2507-2515 (2006).
- Nieuwkoop, P. D., Faber, J. Normal table of Xenopus laevis (Daudin) : a systematical and chronological survey of the development from the fertilized egg till the end of metamorphosis. , Garland Pub. (1994).
- Sive, H. L., Grainger, R. M., Harland, R. M. Early development of Xenopus laevis : a laboratory manual. , Cold Spring Harbor Laboratory Press. (2000).
- Joshi, S. D., Kim, H. Y., Davidson, L. A. Microscopy tools for quantifying developmental dynamics in Xenopus embryos. Methods in Molecular Biology. 917, 477-493 (2012).
- Sater, A. K., Steinhardt, R. A., Keller, R. Induction of neuronal differentiation by planar signals in Xenopus embryos. Developmental Dynamics. 197 (4), 268-280 (1993).
- Sedzinski, J., Hannezo, E., Tu, F., Biro, M., Wallingford, J. B. Emergence of an Apical Epithelial Cell Surface In Vivo. Developmental Cell. 36 (1), 24-35 (2016).
- Rock, J. R., Randell, S. H., Hogan, B. L. M. Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Disease Models & Mechanisms. 3 (9-10), 545-556 (2010).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten