
Method Article
Tiefenfluoreszenzbeobachtung in Reistrieben mittels Clearing-Technologie
In diesem Artikel
Zusammenfassung
Das vorliegende Protokoll beschreibt eine Clearing-Technik für Reissprossen, die aufgrund der harten, dicken oder geschichteten Natur der Gewebe schwer auf interne Strukturbeobachtungen vorzubereiten sind. Diese Methode ermöglicht kontinuierliche und tiefe Fluoreszenzbeobachtungen, auch in erwachsenen Reispflanzen.
Zusammenfassung
Die kürzlich entwickelte Clearing-Technologie, die Brechungsindex-Fehlanpassungen eliminiert und autofluoreszierendes Material verringert, hat es ermöglicht, Pflanzengewebe in drei Dimensionen (3D) zu beobachten und gleichzeitig ihre inneren Strukturen zu erhalten. Bei Reis (Oryza sativa L.), einer einfarbigen Modellpflanze und einer weltweit wichtigen Kulturpflanze, wurde über Reinigungstechnologie in relativ leicht zu beobachtenden Organen wie Wurzeln und Blättern berichtet. Anwendungen der Clearing-Technologie in Sprossapikalmeristem (SAM) und Stängeln wurden ebenfalls berichtet, jedoch nur in begrenztem Maße wegen der schlechten Penetration der Clearing-Lösung (CS) in diesen Geweben. Die begrenzte Effizienz der Clearing-Lösungen in diesen Geweben wurde der Autofluoreszenz, Verdickung und Verhärtung der Gewebe im Stamm zugeschrieben, wenn sich die Gefäßbündel und die Epidermis entwickeln, und der Schichtung des SAM mit wasserabweisenden Blättern. Das vorliegende Protokoll berichtet über die Optimierung eines Clearing-Ansatzes zur kontinuierlichen und 3D-Beobachtung der Genexpression von der SAM/jungen Rispe bis zur Basis der Triebe während der Entwicklung. Fixierte Gewebeproben, die einen fluoreszierenden Proteinreporter exprimierten, wurden mit einem vibrierenden Mikroschneider in Schnitte geschnitten. Wenn eine geeignete Dicke erreicht war, wurde der CS aufgetragen. Durch die gezielte Ausrichtung auf das zentrale Gewebe erhöhte sich die Penetrationsrate und Gleichmäßigkeit des CS und die Zeit, die benötigt wurde, um das Gewebe transparent zu machen, nahm ab. Darüber hinaus ermöglichte die Räumung der beschnittenen Abschnitte die Beobachtung der inneren Struktur des gesamten Triebs aus einer Makroperspektive. Diese Methode hat potenzielle Anwendungen in der tiefen Bildgebung von Geweben anderer Pflanzenarten, die schwer zu klären sind.
Einleitung
Die kürzlich entwickelte Clearing-Technologie hat es ermöglicht, das tiefe Gewebe von Pflanzen zu beobachten und gleichzeitig ihre innere Struktur zu erhalten 1,2,3. In der Dikotosen-Modellpflanze Arabidopsis wurden viele Studien zur fluoreszierenden Proteinbildgebung mit Clearing-Technologie durchgeführt, um Brechungsindex-Fehlanpassungen zu eliminieren und autofluoreszierende Materialien zu entfernen 4,5,6. Obwohl der Einsatz der Clearing-Technologie7,8 und der 3D-Bildgebung bei zellulärer Auflösung9 in Reis (Oryza sativa L.), einer monokotylen Modellpflanze und einer weltweit wichtigen Kulturpflanze, berichtet wurde, sind diese auf relativ dünne und weiche Organe wie Wurzeln, Blätter und Sprossapikalmeristem (SAM) beschränkt, die leicht zu beobachten sind.
Der Spross ist das Hauptorgan, das die oberirdischen Teile von Gefäßpflanzen bildet. In Reis bestehen Triebe aus einer Reihe vertikal gestapelter "Phytomere", bestehend aus axillären Knospen, Blättern und dem Stamm10. An der Spitze des Sprosses besteht das SAM aus undifferenzierten Stammzellen in der Mitte. Phytomere werden durch die Differenzierung von Zellen gebildet, die aus dem SAM stammen. Nachdem die Pflanzen von der vegetativen in die Fortpflanzungsphase übergegangen sind, verlängern sich die Reisstiele und der SAM differenziert sich in jungeRispen 10. Diese Entwicklungsveränderung geht einher mit Schwankungen in der Expression verschiedener Gene in den Stängeln und SAM/jungen Rispen. Um die Mechanismen zu verstehen, die der Zelldifferenzierung in verschiedene Gewebe zugrunde liegen, ist es wichtig, die Zellmorphologie und Genexpression in internen Sprossgeweben strukturell zu beobachten. Die tiefe Bildgebung von Stängeln (Knoten und Internodien) im Spross stellt jedoch eine Herausforderung dar, da die Clearing-Lösungen ineffizient sind, um in das Gewebe einzudringen. Stängel erfahren nach der Differenzierung vom SAM sofort eine schnelle Volumenzunahme durch seitliches Wachstum. Die Verhärtung der Reisknotengewebe aufgrund der Verdickung der Gefäßbündel und die horizontale komplexe Verbindung der nodalen vaskulären Anastomose sowie die hohe Abstoßung der Reissprossen tragen dazu bei, das Eindringen des CS in Stängel10 zu begrenzen.
Diese Studie zielte darauf ab, Veränderungen in der Genexpression in Reistriebgeweben mit einer strukturellen Tiefenfluoreszenztechnik zu beobachten. Diese Arbeit optimiert ein Clearing-Protokoll für Reis, um die Genexpression von der SAM / jungen Rispe bis zur Basis kontinuierlich in einer 3D-Struktur und nicht auf einer flachen Oberfläche mit einem konfokalen Lasermikroskop zu beobachten.
Protokoll
1. Herstellung der Fixierlösung
- 70 ml steriles Wasser in eine Glasflasche geben und 10 g Paraformaldehyd (PFA) hinzufügen.
ACHTUNG: PFA ist giftig; Daher wird das Tragen von Handschuhen empfohlen. - Fügen Sie 2 ml 1 N NaOH hinzu, um die PFA-Lösung aufzulösen.
- Die PFA-Lösung wird unter Dauerrühren (300-500 U/min) bei 60 °C ca. 1 h lang gelöst, bis die PFA-Lösung transparent ist.
- Fügen Sie sterilisiertes Wasser hinzu, um das Volumen der PFA-Lösung auf 100 ml zu erhöhen.
HINWEIS: Die PFA-Lösung wurde vor Gebrauch frisch zubereitet. Darüber hinaus kann eine bei 4 °C gelagerte PFA-Lösung für ca. 2 Monate verwendet werden. - Unmittelbar vor der Probenahme 25 ml 60 mM HEPES (pH 7,4), 3 ml 1 M Saccharose und 2 ml steriles Wasser in 20 ml 10 % PFA geben, um eine 50 ml Fixierlösung herzustellen.
2. Vorbereitung der Clearinglösung (CS)
- Wiegen und mischen Sie 7,5 g Natriumdesoxycholat, 5 g Xylit und 12,5 g Harnstoff (siehe Materialtabelle) in 50 ml sterilem Wasser.
HINWEIS: Natriumdesoxycholatpulver wurde in einer allgemeinen Laborzugkammer gewogen, um eine Dispersion in der Luft zu verhindern. - Lösen Sie die Zutaten (Schritt 2.1.) mit einem Magnetrührer auf, um das CS vorzubereiten.
HINWEIS: In der vorliegenden Studie wurde eine intern hergestellte Clearinglösung (CS) verwendet, aber auch eine kommerziell erhältliche Clearinglösung, ClearSee (siehe Materialtabelle), kann verwendet werden. - Das CS in ein 50 mL konisches Röhrchen geben und bei Raumtemperatur (15-30 °C) im Dunkeln lagern.
HINWEIS: Die CS kann länger als 1 Jahr gelagert werden.
3. Probenahme
- Schneiden Sie die Wurzeln vorsichtig, ohne die Pflanzen zu beschädigen. Waschen Sie die Pflanzen mit Wasser, um den Schmutz zu entfernen.
HINWEIS: Diese Methode untersuchte die Beobachtung von Geweben aus dem dreiblättrigen Stadium (LS) zur Kennzeichnung von LS. Die vorliegende Studie verwendete die Reissorten Nipponbare (Nip) und Taichung 65 (T65) bei 8-10 LS. - Die alten äußeren Blätter mit Händen und Pinzette abziehen. Es waren noch etwa 2-3 Blätter übrig.
- Schneiden Sie das Gewebe von der SAM/jungen Rispe mit einem einschneidigen Rasiermesser mit einer Gleitbewegung zur Basis, um eine Zerquetschung der Zellen zu vermeiden (Abbildung 1A-C). Wenn die Position der SAM/jungen Rispe nicht sichtbar ist, schneiden Sie sie länger.
- Da der Querschnitt des Reistriebs eine ovale Form hat (Abbildung 1D), rasieren Sie eine Seite des Ovals mit einem zweischneidigen Rasiermesser dünn in Richtung seiner Längsachse ab (Abbildung 1E-F).
HINWEIS: Dieser Schritt erleichterte es der Fixierlösung, in die Probe einzudringen. Eine horizontale Schnittfläche erleichtert das Anhaften der Probe an einer vibrierenden Mikroschneidemaschine (siehe Materialtabelle). - Bestätigen Sie die Position der SAM/jungen Rispe durch das Auftreten einer weißlichen Farbe in der Probe (Abbildung 1G). Schneiden Sie überschüssiges Gewebe ab.
HINWEIS: Die Länge der Probe sollte auf die maximale Länge eingestellt werden, die mit einem vibrierenden Mikroslicer zugeschnitten werden kann. Wenn die Probe zu lang ist, teilen Sie sie in mehrere Stücke.
4. Fixierung der Proben
- 1 ml der Fixierlösung (hergestellt in Schritt 1) in ein 1,5 ml Mikrozentrifugenröhrchen pipettieren und sofort in die Proben geben.
HINWEIS: Stellen Sie sicher, dass die Probenmenge 20% des Volumens der Fixierlösung nicht überschreitet. - Schneiden Sie ein Stück Paraffinfolie in 2,5 cm2. Formen Sie die Paraffinfolie zu Kugeln und legen Sie sie auf die Proben, um zu verhindern, dass sie aus der Fixierlösung herausschwimmen.
- Legen Sie das Röhrchen mit der Probe in einen Exsikkator. Schließen Sie dann den Deckel des Exsikkators und starten Sie die Vakuumpumpe, um das Innere des Exsikkators langsam bis -0,095 MPa zu entlasten.
- Nach dem Schließen des Exsikkators bei −0,095 MPa schalten Sie die Vakuumpumpe aus und lassen Sie sie 30 min bei Raumtemperatur stehen.
HINWEIS: Das Vakuumniveau war so, dass allmählich Blasen in den Proben auftraten. - Öffnen Sie den Exsikkator leicht und bringen Sie ihn langsam wieder auf Atmosphärendruck. Reduzieren Sie den Druck des Exsikkators auf −0,095 MPa, schalten Sie die Vakuumpumpe aus und lassen Sie die Mischung 30 min bei Raumtemperatur stehen.
- Öffnen Sie den Exsikkator leicht und bringen Sie ihn langsam wieder auf Atmosphärendruck. Wenn die Probe richtig fixiert ist, sinkt sie in die Fixierlösung. Wenn es nicht gesunken ist, reduzieren Sie den Druck wieder.
- Entfernen Sie die Paraffinfolie, schließen Sie den Deckel und bewahren Sie die Tuben über Nacht bei 4 °C auf.
HINWEIS: Die Proben wurden für mindestens 2 h in fixierender Lösung fixiert. Es kann für ~ 1 Woche gelagert werden.
5. Trimmen der fixierten Probe
- Entfernen Sie die überschüssige Fixierlösung mit fusselfreien Tüchern aus der Probe.
HINWEIS: Überschüssige Fixierlösung verhindert, dass die Probe ordnungsgemäß auf dem Tablett haftet. - Fixieren Sie die Probe mit Sofortkleber auf einer vibrierenden Mikroschneideschale. Legen Sie es mit der flachen Seite nach unten auf ein Tablett, das während der Probenahme abrasiert wurde.
HINWEIS: Aufgrund der Struktur des Reistriebs muss er von der Basis in Richtung SAM / junge Rispe geschnitten werden. - Stellen Sie die Beschneidungsbedingungen entsprechend der festgelegten Probe ein.
HINWEIS: In dieser Studie wurde das Trimmen auf DICK, 130 μm eingestellt; BREITE, 15 mm; HÄUFIGKEIT, 15%; und SCHNITTGESCHWINDIGKEIT, 32 mm. - Stellen Sie die Probenposition mit der RVS- oder FWD-Taste und der UP- oder DOWN-Taste ein. Füllen Sie die Schale mit entionisiertem Wasser, bis alle Proben eingetaucht sind. Drücken Sie nach dem Befüllen des Fachs die Taste START. Legen Sie die zugeschnittenen Proben mit einem Tropfen 1x PBS auf einen Glasobjektträger.
HINWEIS: Verschieben Sie die Position der Probe nach jedem Trimmen auf die andere Seite, da die harte Probe die Rasierklinge verschleißt. - Nach Überprüfung der Probe, die die Zielstelle enthält, unter einem Stereomikroskop wird die zugeschnittene Probe auf eine Multiwell-Platte mit 1 ml 1x PBS überführt.
HINWEIS: Wählen Sie die Anzahl der Vertiefungen entsprechend der Größe der Stichprobe. In dieser Studie wurde eine 12-Multiwell-Platte verwendet.
6. Behandlung mit der Clearinglösung
- Entfernen Sie das 1x PBS von der Multiwell-Platte und fügen Sie frisches 1x PBS hinzu. 1 min stehen lassen.
- Entfernen Sie 1x PBS und fügen Sie das CS hinzu.
HINWEIS: Die Menge an CS muss das Fünffache des Probenvolumens betragen. - Legen Sie die Multiwell-Platte mit der Probe in einen Exsikkator. Schließen Sie dann den Deckel des Exsikkators und starten Sie die Vakuumpumpe, um das Innere des Exsikkators langsam bis -0,09 MPa zu entlasten.
- Nach dem Schließen des Exsikkators bei −0,09 MPa die Vakuumpumpe ausschalten und 1 h bei Raumtemperatur stehen lassen.
- Öffnen Sie den Exsikkator leicht und bringen Sie ihn langsam wieder auf Atmosphärendruck.
- Gießen Sie entionisiertes Wasser in den Spalt zwischen den Brunnen, um die Verdunstung des CS zu verhindern. Lagern Sie die Multiwell-Platten bei Raumtemperatur im Dunkeln zur Reinigung.
HINWEIS: Schütteln Sie die Multiwell-Platte alle 1-2 Tage vorsichtig, um das Chlorophyll und andere Komponenten zu diffundieren. - Wenn der CS grün wird, ersetzen Sie ihn durch eine neue Lösung.
7. Zellwandfärbung mit chemischem Farbstoff
- Die weiße Calcofluorlösung (1 ml, Endkonzentration: 1 mg/ml in der Reinigungslösung, siehe Materialtabelle) wird in eine Multiwell-Platte pipet. Die Proben in die Farbstofflösung geben und 1 h inkubieren.
HINWEIS: Das Volumen des Farbstoffs darf 10% des Volumens des CS nicht überschreiten. - Waschen Sie die gefärbten Proben in einer anderen Multiwell-Platte mit 1 ml CS für mindestens 1 h.
8. Beobachtung mit einem konfokalen Lasermikroskop
- Geben Sie einen Tropfen CS auf einen Objektträger und übertragen Sie die behandelten (mit CS) Proben auf den Objektträger.
HINWEIS: Da die behandelten Proben weich sind, müssen sie vorsichtig mit einer Pinzette behandelt werden. - Decken Sie die Probe mit einem Glasdeckglas ab, um das Verdampfen des CS zu verhindern.
HINWEIS: Der CS fällt leicht aus. - Beobachten Sie die behandelten Proben unter einem konfokalen Lasermikroskop.
HINWEIS: Nach der Beobachtung können die behandelten Proben für spätere weitere Beobachtungen an die CS zurückgegeben werden.
Ergebnisse
Zunächst wurde validiert, inwieweit das harte und dicke Gewebe erwachsener Reispflanzen durch die Clearing-Behandlung transparent gemacht werden konnte. Nip am 9-10 LS, dem Stadium der Rispenbildung, und 12-16 mm lange Gewebe, die von der jungen Rispe bis zur Basis des Triebs herausgeschnitten wurden, wurden verwendet. Diese Proben, einschließlich der jungen Rispe, wurden in der fixierenden Lösung fixiert und vor und nach 1-4 Wochen der Clearing-Behandlung beobachtet (Abbildung 2). Die unbeschnittenen Proben blieben grün und wurden nach 4 Wochen oder gar nach 3 Monaten Behandlung mit der Clearing-Lösung nicht transparent (Abbildung 2A). In Proben, die von allen Blättern abgezogen wurden, wurden die Internodien nach 1 Woche Behandlung mit dem CS weiß, obwohl die Knoten grün blieben. Diese Probe wurde nach 4 Wochen nicht transparent (Abbildung 2B). Nach 3 Monaten der Behandlung mit dem CS wurde nur die junge Rispe transparent. Die handgetrimmten Proben wurden nach 1 Woche Behandlung mit dem CS mit Ausnahme der äußeren Blätter und Gefäßbündel weiß. Einige Teile der Blätter, junge Rispe und Internodien wurden durchscheinend, aber nach 4 Wochen nicht mehr transparent (Abbildung 2C). Nach 3 Monaten wurden nur die junge Rispe und die inneren Blätter transparent. Die Proben, die mit dem vibrierenden Mikroschneider auf 130 μm Dicke zugeschnitten wurden, wurden nach 1 Woche Behandlung mit dem CS durchscheinend, wobei die Blätter transparent wurden. Nach 2 Wochen war die Probe nahezu transparent und blieb nach 4 Wochen unverändert (Abbildung 2D). Nach 3 Monaten wurden die Proben vollständig transparent.
Als nächstes wurde die Tiefe validiert, in der die Clearing-Behandlung Genexpressionsbeobachtungen im Dreh von T65 bei der 8 LS ermöglichte. Die UBQpro:: NLS-sGFP-nClover3-mNeonGreen (UBQpro ::NGCN, siehe Materialtabelle) exprimierten Triebe wurden verwendet. Die Gewebeproben wurden in der Fixierlösung fixiert, bevor sie mit einem vibrierenden Mikroslicer auf 130 μm Dicke zugeschnitten wurden. Die Proben wurden dann unter einem konfokalen Lasermikroskop (Laser: 488 nm/13%, Objektiv: 20x, Pinhole: 0,93 AE, Mittelwert: 16, Detektorgewinn: 800) in Abständen von 10 μm in Richtung der z-Achse jede Woche von 0-4 Wochen Behandlung mit dem CS beobachtet. Die repräsentativen Ergebnisse sind in Abbildung 3 dargestellt. Alle Zahlen wurden mit der gleichen Verarbeitung angepasst, um die Fluoreszenzintensität zu vergleichen. Im Knoten wurden ohne die Behandlung schwache fluoreszierende Signale in einer Tiefe von −40 μm beobachtet (Abbildung 3A). Die beobachtbare Tiefe betrug −60 μm nach 1 Woche der CS-Behandlung und −90 μm nach 2 Wochen, aber die Autofluoreszenz des Zytoplasmas war in beiden Wochen in einer Tiefe von −20 μm bemerkbar. Die Autofluoreszenz wurde nach 3 Wochen etwas schwächer und war nach 4 Wochen nicht mehr sichtbar. Fluoreszierende Signale wurden in einer Tiefe von −80 μm nach 3 Wochen und in einer Tiefe von −100 μm nach 4 Wochen beobachtet. Im Internodium wurden fluoreszierende Signale in einer Tiefe von −60 μm ohne CS-Behandlung beobachtet (Abbildung 3B). Die beobachtbare Tiefe betrug −90 μm nach 1 Woche bis 3 Wochen der CS-Behandlung und −100 μm nach 4 Wochen, aber die Autofluoreszenz im Zytoplasma war in einer Tiefe von −20 μm nach 1 Woche bemerkbar. Die Autofluoreszenz war nach 2 Wochen nicht mehr sichtbar. In den Blättern wurden fluoreszierende Signale in einer Tiefe von −90 μm ohne CS-Behandlung beobachtet (Abbildung 3C). Die fluoreszierenden Signale wurden tiefer in den Blättern beobachtet als in den Knoten und Internodien. Die fluoreszierenden Signale waren jedoch schwächer. Die fluoreszierenden Signale wurden nach 1 Woche CS-Behandlung klar und wurden in einer Tiefe von −120 μm beobachtet. Nach 2 Wochen und 3 Wochen wurden fluoreszierende Signale in einer Tiefe von −110 μm und nach 4 Wochen in einer Tiefe von −120 μm beobachtet.
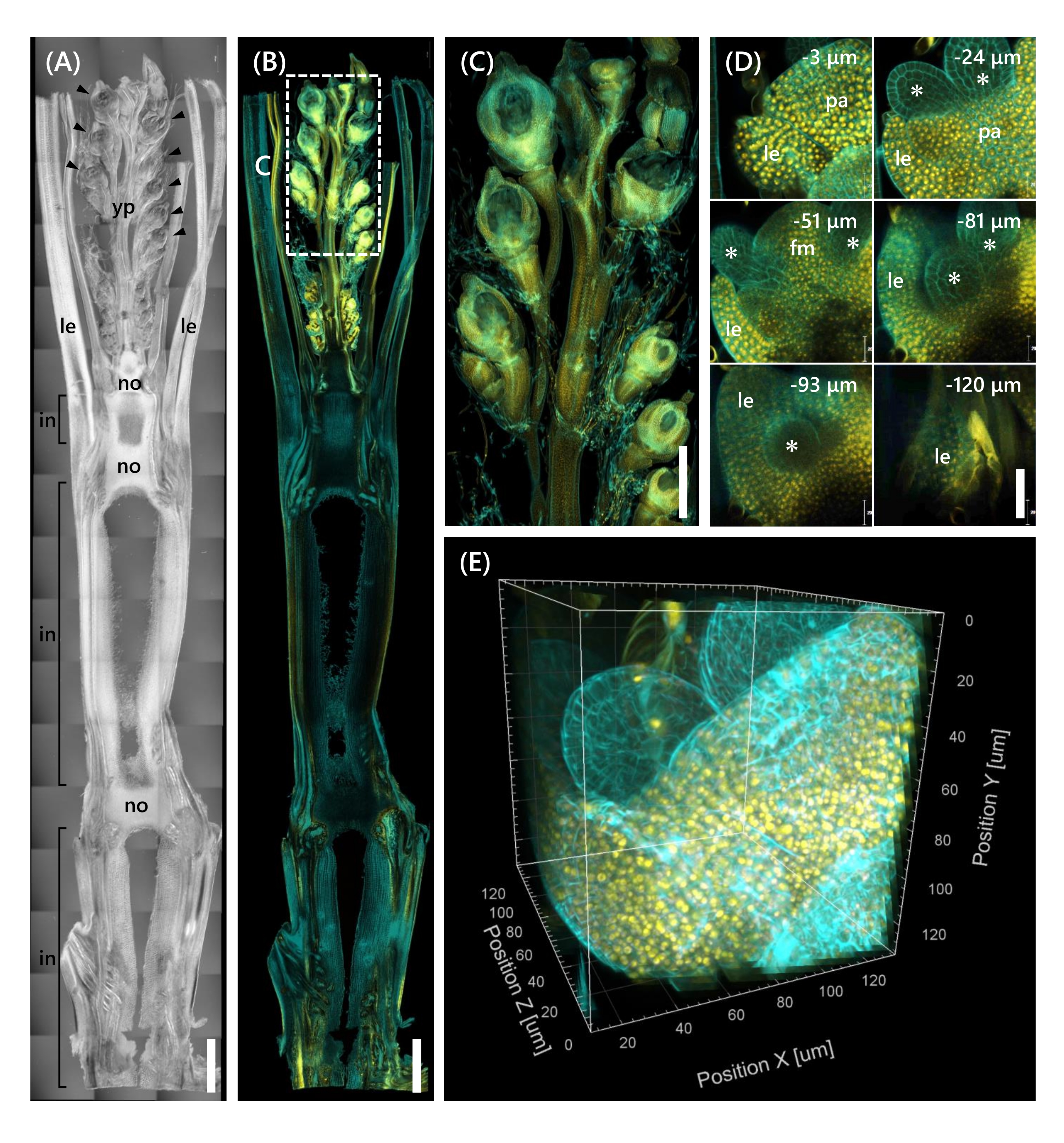
Schließlich wurde die Expression des mit mOrange12 fusionierten Transkriptionsfaktors OsMADS1511 beobachtet, der den Übergang von SAM von der vegetativen in die reproduktive Phasereguliert 13. Gewebeproben aus Nip an der 10 LS mit OsMADS15-mOrange wurden fixiert, auf 130 μm Dicke getrimmt und 2 Wochen lang mit dem CS behandelt. Abbildung 4 zeigt, dass die Clearing-Lösung gleichzeitig verwendet werden konnte, um die fluoreszierenden Proteine (OsMADS15-mOrange) der natürlichen Genexpression und der Fluoreszenzfarbstofffärbung (Calcofluorweiß) zu beobachten. Die gesamte Blüte in Tiefen von 0 bis −130 μm wurde beobachtet, und das 3D-Bild wurde aus 43 z-Stack-Bildern in Abständen von 3 μm rekonstruiert (Abbildung 4E).

Abbildung 1: Probenahmeposition und Verarbeitung. (A) Nip-Sämling bei der 8 LS unter Kurztagesbedingung. (B) Vergrößerte Ansicht der Basis. (C) Ein Probenausschnitt von der SAM/jungen Rispe bis zur Basis. (D) Ein ovaler Querschnitt von C (Einschub). (E) Eine Seite des Sprosses mit einer Klinge dünn abrasieren. (F) Dünn rasierte Oberfläche. (G) Stichprobe mit Internodiendehnung. Der weißliche Abschnitt, der durch die Pfeilspitzen angezeigt wird, ist Knoten. Die Oberseite des oberen Knotens hat die junge Rispe. Maßstabsbalken: (A) 10 cm, (B-C,E-G) 1 cm, (D) 0,5 cm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2: Die Gewebereinigung hängt von der Probendicke und der CS-Behandlungsdauer ab . (A) Unbeschnittene Proben mit partiellen Trieben einschließlich der jungen Rispe. Maximale Dicke: 4 mm. (B) Alle Blätter werden von den Proben abgezogen, um die junge Rispe freizulegen. Maximale Dicke: 2,5 mm. yp: junge Rispe, in: Internodium; Pfeilspitzen zeigen den Knoten an. (C) Proben, die von Hand auf 1 mm Dicke zugeschnitten wurden. (D) Proben, die mit dem vibrierenden Mikroslicer auf eine Dicke von 130 μm zugeschnitten wurden. Maßstabsleiste: 5 mm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 3: Die Tiefe der Fluoreszenzbeobachtung hängt vom Gewebetyp und der CS-Behandlungsdauer ab. Tiefendarstellung von UBQpro::NGCN im (A)-Knoten, (B) Internodium und (C)-Blatt. "Zusammenführen" bedeutet, dass die Bilder mit dem DIC-Bild (Differential Interference Contrast) und allen Z-Stack-Bildern verschmolzen wurden. Bilder, in denen keine Fluoreszenz beobachtet wurde, wurden als "nd" markiert. (nicht erkannt). Einstellungen der konfokalen Lasermikroskopie sind wie folgt: Laser: 488 nm/13%, Objektiv: 20x, Pinhole: 0,93 AE, Mittelwert: 16, Detektorverstärkung: 800, Intervall: 10 μm. Maßstabsbalken: 50 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 4: Tiefenfluoreszenzbeobachtung von OsMADS15-mOrange. (A) Das DIC-Bild von Nip bei 10 LS. yp: junge Rispe, le: Blatt, no: Knoten, in: Internodium; Pfeilspitzen zeigen Röschen an. (B) Fluoreszenzbild von OsMADS15-mOrange. Gelbe Punkte zeigen die fluoreszierenden Proteine von OsMADS15-mOrange am Zellkern an. Die Zellwand wurde mit der weißen Lösung von Calcofluor cyan gefärbt. (C) Vergrößerte Ansicht der jungen Rispe. (D) Tiefe Bildgebung der Blüte. PA: Palea, LE: Lemma, FM: Floral Meristem; Sternchen weisen auf Staubblätter primordia hin. Das konfokale Lasermikroskop wurde wie folgt eingestellt: OsMADS15-mOrange; Laser: 555 nm/13%, Objektiv: 20x, Pinhole: 0,96 AE, Mittelwert: 16, Detektorverstärkung: 800, Intervall: 3 μm. Calcofluor weiß; Laser: 405 nm/3%, Objektiv: 20x, Pinhole: 0,93 AE, Mittelwert: 16, Detektorverstärkung: 470, Intervall: 3 μm. (E) Das 3D-Bild der Abbildung (D), konstruiert aus den Z-Stack-Bildern. Maßstabsbalken: (A-B) 1 mm, (C) 500 μm, (D) 40 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Kritische Schritte des Protokolls
Die kritischen Schritte in diesem Protokoll sind Fixieren und Trimmen. Reissprossen haben harte, dicke oder geschichtete Gewebe, die das Eindringen der Fixierlösung begrenzen. Um die Durchlässigkeit der Fixierlösung zu verbessern, wurde eine Seite des Gewebes bei der Probenahme dünn rasiert, wie in Abbildung 1E-F gezeigt. Darüber hinaus wurden die Vakuumbehandlungen zweimal mit höherem Druck wiederholt. Darüber hinaus wurden die Proben über Nacht bei 4 °C anstelle der üblichen 2 h Fixierung bei 4 °C fixiert.
Der Schlüsselpunkt im Trimmschritt besteht darin, die Dicke der zu erstellenden Gewebe zu bestimmen, um die fluoreszierenden Proteine zu beobachten und gleichzeitig ihre innere Struktur nach einer kurzen CS-Behandlung zu erhalten. Wie in Abbildung 2C gezeigt, wurden die 1 mm dicken Proben, die so dünn wie möglich von Hand geschnitten wurden, auch nach 3 Monaten CS-Behandlung nur in einer begrenzten Anzahl von Geweben transparent. Daher ist der Schnittschritt essentiell für die Tiefenfluoreszenzbeobachtung von erwachsenen Reissprossen. In dieser Studie wurden die Proben auf eine Dicke von 130 μm zugeschnitten, wie in Abbildung 2D gezeigt. Die Dicke von 130 μm ermöglichte es, die Blätter nach 1 Woche CS-Behandlung und die gesamte Probe nach 2 Wochen zu reinigen. Erwachsene Reistriebe bei 9-10 LS wurden in dieser Studie verwendet. Dickes, aber weicheres Gewebe von jüngeren Reistrieben kann mit der CS-Behandlung schneller beseitigt werden. Die Dicke der Proben und die Dauer der CS-Behandlung sollten entsprechend dem Gewebetyp, dem Zustand und der Dicke der zu beobachtenden 3D-Struktur angepasst werden.
Änderungen und Methoden zur Fehlerbehebung
Der CS fiel bei niedrigen Temperaturen leicht aus. Das ausgefällte CS kann die fluoreszierenden Proteine nicht konservieren; Daher ist bei der Lagerung der Proben bei der richtigen Temperatur Vorsicht geboten. Darüber hinaus haben sowohl das CS als auch die fixierende Lösung keine antiseptische Wirkung; Daher werden fluoreszierende Proteine abgebaut, wenn sie kontaminiert werden. Bodenreis ist anfällig für Pilzwachstum; Daher müssen die Probenahme und Handhabung der Proben sorgfältig durchgeführt werden, um eine Kontamination zu vermeiden.
Überschüssige Fluoreszenzfarbstoffe im Puffer können Hintergrundfluoreszenz abgeben und mikroskopische Beobachtungen stören. Zum Beispiel wurde zuvor eine Calcofluor-Weißlösung verwendet, die Evansblau-Farbstoff enthält. Nach Färbung für 1 h und Waschen für 1 h wurden die fluoreszierenden Proteine von OsMADS15-mOrange mit einem 555 nm Laser beobachtet. Fluoreszierende Proteine konnten jedoch aufgrund der Hintergrundfluoreszenz, die von Evans-Blaufarbstoff abgeleitet wurde, nicht beobachtet werden. Diese Hintergrundfluoreszenz wurde durch Waschen der Proben für 2 h fast eliminiert. Darüber hinaus waren die fluoreszierenden Proteine klarer, wenn die Proben über Nacht belassen wurden. Daher wurde in dieser Studie eine reine Calcofluor-Weißlösung verwendet. Die vom Fluoreszenzfarbstoff abgeleitete Hintergrundfluoreszenz sollte vor Beobachtungen mit verschiedenen Laserwellenlängen überprüft werden.
Einschränkungen der Methode
Wie in Abbildung 3 gezeigt, wurden tiefe fluoreszierende Proteine in den Proben beobachtet, die nach 2 Wochen CS-Behandlung 130 μm dick waren. Dies stimmt mit den Ergebnissen in Abbildung 2D überein, wo die 130 μm dicke Probe nach 2 Wochen CS-Behandlung transparent wurde. Wie in Abbildung 3A gezeigt, war die Autofluoreszenz des Zytoplasmas in den Knoten jedoch nach 2 Wochen noch spürbar und wurde erst nach 4 Wochen CS-Behandlung vollständig entfernt. Knoten haben eine hohe Zelldichte und benötigen daher eine längere Zeit, um autofluoreszierende Materialien zu entfernen.
Wie in Abbildung 3C gezeigt, wurden tief fluoreszierende Proteine in den Blättern ohne CS-Behandlung beobachtet, aber die Helligkeit war schwächer als in den Knoten und Internodien in der gleichen Tiefe von 20 μm. Nach 1 Woche CS-Behandlung waren die fluoreszierenden Proteine heller. Chlorophyll ist in Blättern reichlich vorhanden und absorbiert 488 nm Anregungslicht. Sie haben auch eine orange/rote Autofluoreszenz, die die Beobachtung fluoreszierender Proteine mit einem 555-nm-Laser stören kann. Nach 1 Woche CS-Behandlung wurden Chlorophyll und andere autofluoreszierende Materialien entfernt, was zu Bildern mit hohem Signal-Rausch-Verhältnis führte.
Die Tiefen, die nach 2 Wochen und 4 Wochen CS-Behandlung in Geweben beobachtet werden konnten, unterschieden sich nicht signifikant, obwohl die fluoreszierenden Proteine nach 4 Wochen schwächer erschienen (Abbildung 3). Normalerweise schwächt sich die Helligkeit fluoreszierender Proteine und die Autofluoreszenz mit der Zeit ab, was zu einem höheren Signal-Rausch-Verhältnis führt. Daher können fluoreszierende Proteine durch Anpassung der mikroskopischen Bedingungen und Bildverarbeitung deutlicher beobachtet werden. Basierend auf diesen Ergebnissen wurde der Schluss gezogen, dass eine 2-wöchige CS-Behandlung die Beobachtung von tief fluoreszierenden Proteinen unter unseren Probenbedingungen erleichtern könnte. Es werden jedoch 4 Wochen benötigt, um klarere Bilder zu beobachten, die die autofluoreszierenden Materialien vollständig ausschließen.
Strukturen mit starker Autofluoreszenz, wie Gefäßbündel und mehrarmige Zellen, können im CS nicht beseitigt werden. Um diese Strukturen ohne Autofluoreszenz zu beobachten, ist es notwendig, ein Time-Gating-Verfahren12 zu verwenden oder Bilder des Fluoreszenzspektrums durch Spektroskopie zu erhalten. Ein Zwei-Photonen-Mikroskop kann besser für die Beobachtung tieferer Gewebe geeignet sein, wenn dickere Gewebe beobachtet werden.
Bedeutung der Methode in Bezug auf bestehende und alternative Methoden
Im Allgemeinen wurden die inneren Strukturen von Reispflanzen entweder durch Kryostat- oder Vibratomschnitte beobachtet. Ein Kryostat eignet sich zur Vorbereitung von Dünnschliffen, die eine einfachere Beobachtung ermöglichen, aber die Vorbereitung der Proben und der Betrieb der Ausrüstung sind zeitaufwendig. Die ursprüngliche 3D-Struktur aus dünnen Abschnitten zu rekonstruieren, ist ebenfalls schwierig. Das Vibratom ist relativ einfach zu bedienen und für die Herstellung dicker Profile geeignet. Dicke Abschnitte des Zielgewebes erlauben jedoch nur Beobachtungen der Schnittoberfläche und nicht tiefes Gewebe, das Licht nicht erreichen kann. Aus diesen Gründen eignet sich keine der beiden Methoden für Tiefenfluoreszenzbeobachtungen.
Diese Studie befasste sich mit Herausforderungen bei der Tiefenfluoreszenzbeobachtung bei Reistrieben, wie der begrenzten Gewebepenetration des CS und der schlechten Objektauflösung unter einem konfokalen Mikroskop, indem bestehende Methoden kombiniert wurden. Wie in Abbildung 4 gezeigt, beobachteten wir fluoreszierende Proteine (OsMADS15-mOrange), die in den tiefen Geweben erwachsener Reistriebe von der jungen Rispe bis zur Basis exprimiert wurden. Abbildung 4D konzentriert sich auf das Blümchen und zeigt tief fluoreszierende Proteine in Abständen von 3 μm. Gewebe über −130 μm Tiefe wurden nach 2 Wochen CS-Behandlung beobachtet, aber nur die Gewebe innerhalb von −27 μm Tiefe wurden im Blüten in der gleichen Größe und Wachstumsphase ohne CS-Behandlung beobachtet (Daten nicht gezeigt). Das derzeit verbesserte Protokoll ermöglichte nicht nur die Beobachtung der Überexpression von Genen, sondern auch der natürlichen Genexpression in den tiefen Geweben erwachsener Reistriebe.
Bedeutung und Einsatzmöglichkeiten der Methode in spezifischen Forschungsbereichen
Dieses Protokoll, das die Tiefenfluoreszenzbeobachtung von erwachsenen Reissprossen optimiert, ermöglicht die effiziente Reinigung von hartem, dickem oder geschichtetem Gewebe, indem unnötiges Gewebe abgeschnitten und die Durchlässigkeit des CS erhöht wird. Darüber hinaus wurde die Dicke der Proben für die Analyse optimiert, um eine kontinuierliche und strukturelle Tiefenfluoreszenzbeobachtung mit einem konfokalen Lasermikroskop zu ermöglichen, das normalerweise dicke oder undurchsichtige Gewebe nicht auflösen kann.
Es ist schwierig, Reisproben in verschiedenen Wachstumsstadien zu vergleichen, da die fluoreszierenden Proteine im Laufe der Zeit in den Fixier- und PBS-Lösungen abgebaut werden. Die fluoreszierenden Proteine im CS können jedoch länger als 5 Monate gelagert werden1. Die lange Haltbarkeit des CS ist ein großer Vorteil für die Tiefenfluoreszenzbeobachtung in Reis.
In jüngster Zeit wurden viele Clearing-Technologien entwickelt, die es ermöglichen, tiefe Gewebe in 3D zu beobachten und gleichzeitig ihre inneren Strukturen zu erhalten. Diese Technologien haben sich weiterentwickelt und neue Clearing-Lösungen wurden entwickelt. Ein gutes Beispiel ist iTOMEI14, das eine effiziente Entfernung von Chlorophyll und eine hellere Fluoreszenzdetektion ermöglicht. Ein weiteres Beispiel ist ClearSeeAlpha15, das die Bräunung von Geweben während der Clearing-Behandlung verhindert und transparent erscheinen lässt. Die Kombination dieser Clearinglösungen mit der vorliegenden Methode kann ein effizienteres und effektiveres Clearing ermöglichen.
Es wird erwartet, dass die aktuelle Methode dazu beitragen wird, neue Erkenntnisse durch tiefe Bildgebung nicht nur von Reis, sondern auch von anderen Pflanzen zu gewinnen.
Offenlegungen
Die Autoren haben keine Interessenkonflikte zu erklären.
Danksagungen
Wir danken Dr. R. Terada, Dr. Z. Shimatani und Dr. H. Tsuji für die Bereitstellung der OsMADS15-mOrange-Samen; Dr. D. Kurihara für die Bereitstellung der NGCN-Konstruktion; und Dr. R. Shim für die Bearbeitung unseres Manuskripts. Diese Arbeit wurde von JSPS KAKENHI (Förderkennzeichen JP20H05912, 20H05778, 20H05779) und durch das SATREPS-Programm (Nr. JPMJSA1706) der JST und JICA.
Materialien
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tube | BIO-BIK | ST-0150F | |
| 12-multiwell plate | Corning | 353043 | |
| 50 mL conical tube | Corning | 352070 | |
| Calcofluor white solution | Sigma-Aldrich | 910090 | |
| ClearSee | FUJIFILM Wako Pure Chemical | 031-25151 | This can be made or purchased. |
| Confocal laser microscope | Carl Zeiss | LSM700 | |
| Desiccator | SANPLATEC | Custom made of acrylic. 30 cm (L), 30 cm (W), 14.5 cm (H) | |
| Glass coverslip (18 × 18 No.1) | MATSUNAMI | C018181 | |
| HEPES | FUJIFILM Wako Pure Chemical | 342-01375 | |
| Microscope slide (76 × 26) | MATSUNAMI | S2441 | |
| Paraffin film | Bemis | PM-996 | |
| Paraformaldehyde | FUJIFILM Wako Pure Chemical | 162-16065 | |
| Sodium deoxycholate | Tokyo Chemical Industry | C0316 | |
| Sucrose | FUJIFILM Wako Pure Chemical | 190-00013 | |
| UBQpro::NLS-sGFP-nClover3-mNeonGreen (UBQpro::NGCN) | provided by Dr. Kurihara | ||
| Urea | FUJIFILM Wako Pure Chemical | 211-01213 | |
| Vacuum pump | AS One | AS-01 | |
| Vibrating micro-slicer | DOSAKA | DTK-3000 | |
| Xylitol | FUJIFILM Wako Pure Chemical | 248-00545 |
Referenzen
- Kurihara, D., Mizuta, Y., Sato, Y., Higashiyama, T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development. 142 (23), 4168-4179 (2015).
- Hasegawa, J., et al. Three-dimensional imaging of plant organs using a simple and rapid transparency technique. Plant and Cell Physiology. 57 (3), 462-472 (2016).
- Musielak, T. J., Slane, D., Liebig, C., Bayer, M. A versatile optical clearing protocol for deep tissue imaging of fluorescent proteins in Arabidopsis thaliana. PLoS One. 11 (8), 0161107 (2016).
- Pandey, K. B., et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science. 371 (6526), 276-280 (2021).
- Ejaza, M., Bencivenga, S., Tavaresa, R., Busha, M., Sablowski, R. ARABIDOPSIS THALIANA HOMEOBOX GENE 1 controls plant architecture by locally restricting environmental responses. Proceedings of the National Academy of Sciences of the United States of America. 118 (17), (2021).
- Smetana, O., et al. High levels of auxin signalling define the stem-cell organizer of the vascular cambium. Nature. 565 (7740), 485-489 (2019).
- Chu, T. T. H., et al. Sub-cellular markers highlight intracellular dynamics of membrane proteins in response to abiotic treatments in rice. Rice. 11 (1), 23 (2018).
- Nagaki, K., Yamaji, N., Murata, M. ePro-ClearSee: A simple immunohistochemical method that does not require sectioning of plant samples. Scientific Reports. 7, 42203 (2017).
- Sato, M., Akashi, H., Sakamoto, Y., Matsunaga, S., Tsuji, H. Whole-tissue three-dimensional imaging of rice at single-cell resolution. International Journal of Molecular Sciences. 23 (1), 40 (2022).
- Matsuo, T., Hoshikawa, K. Science of the Rice Plant: Morphology. Food and Agriculture Policy Research Center. , (1993).
- Kobayashi, K., et al. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-Like MADS box genes and PAP2, a SEPALLATA MADS box gene. Plant Cell. 24 (5), 1848-1859 (2012).
- Tamaki, S., et al. FT-like proteins induce transposon silencing in the shoot apex during floral induction in rice. Proceedings of the National Academy of Sciences of the United States of America. 112 (8), 901-910 (2015).
- Kodama, Y. Time gating of chloroplast autofluorescence allows clearer fluorescence imaging In Planta. PLoS One. 11 (3), 0152484 (2016).
- Sakamoto, Y., et al. Improved clearing method contributes to deep imaging of plant organs. Communications Biology. 5 (1), 12 (2022).
- Kurihara, D., Mizuta, Y., Nagahara, S., Higashiyama, T. ClearSeeAlpha: Advanced optical clearing for whole-plant imaging. Plant and Cell Physiology. 62 (8), 1302-1310 (2021).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten