
Se requiere una suscripción a JoVE para ver este contenido. Inicie sesión o comience su prueba gratuita.
Method Article
La sedimentación de equilibrio de una proteína de membrana pequeña formador de oligómeros: Efecto de la histidina protonación en pentamérica Estabilidad
En este artículo
Resumen
Sedimentation equilibrium (SE) can be used to study protein-protein interactions in a physiological environment. This manuscript describes the use of this technique to determine the effect of pH on the stability of a homo-pentamer formed by the small hydrophobic (SH) protein encoded by the human syncytial respiratory virus (hRSV).
Resumen
Ultracentrifugación analítica (AUC) se puede utilizar para estudiar las interacciones reversibles entre macromoléculas a través de una amplia gama de fuerzas de interacción y bajo condiciones fisiológicas. Esto hace AUC un método de elección para evaluar cuantitativamente la estequiometría y la termodinámica de homo- y hetero-asociación que son transitorios y reversibles en los procesos bioquímicos. En la modalidad de equilibrio de sedimentación (SE), un equilibrio entre la difusión y sedimentación proporciona un perfil como una función de la distancia radial que depende de un modelo de asociación específica. En la presente memoria, un protocolo detallado SE se describe para determinar el tamaño y el monómero-monómero asociación de energía de un pequeño oligómero proteína de la membrana utilizando una ultracentrífuga analítica. AUC-ES es libre de etiquetas, solamente basado en principios físicos, y se puede utilizar en ambas proteínas solubles y de membrana de agua. Se muestra un ejemplo de este último, la proteína hidrófoba pequeña (SH) en el virus sincitial respiratorio humano (VRSh), Un polipéptido de ácido 65-amino con una sola transmembrana (TM) de dominio α-helicoidal que forma canales iónicos pentaméricas. Datos estructurales a base de RMN muestra que la proteína SH tiene dos protonable residuos de His en su dominio transmembrana que están orientados dirigida a la luz de la canal. SE experimentos han sido diseñados para determinar cómo afecta a pH constante de asociación y el tamaño oligomérico de la proteína SH. Mientras que la forma pentamérica se conservó en todos los casos, su constante asociación se redujo a pH bajo. Estos datos están de acuerdo con una dependencia del pH similares observados para la actividad del canal SH, en consonancia con una orientación lumenal de los dos residuos de His en la proteína SH. Este último puede experimentar la repulsión electrostática y la reducción de la estabilidad oligómero a pH bajo. En resumen, este método es aplicable siempre que la información cuantitativa sobre los cambios de asociación entre proteínas sutiles en condiciones fisiológicas tiene que medir.
Introducción
Ultracentrifugación analítica 1-5 es uno de los métodos más importantes para estudiar las interacciones de macromoléculas en condiciones fisiológicas, siendo accesible tanto a interacciones débiles y fuertes. El método es libre de etiquetas y utiliza la absorción de luz o interferencia, e incluso los sistemas ópticos de fluorescencia se puede utilizar para acceder a los intervalos de concentración en varios órdenes de magnitud 6.
Este método es especialmente útil ya que la mayoría de los procesos bioquímicos dependen de las interacciones reversibles. La estequiometría y la fuerza de estas interacciones tienen que caracterizarse cuantitativamente a entender los procesos biológicos, y un número de métodos existen para este propósito 7, 8. Sin embargo, las interacciones transitorias son difíciles de estudiar 9.
La elección de un método para caracterizar las interacciones macromoleculares depende de su naturaleza estática o dinámica. En el primer caso, Sedim se utiliza la velocidad entación (SV), donde se mide la tasa de transporte radial y complejos se fraccionó sobre la base de diferencias en la masa flotante y forma.
En contraste, las asociaciones dinámicas que son reversibles en la escala de tiempo del experimento no se pueden separar físicamente. En este caso, auto o hetero-interacciones que conducen a interacciones no covalentes están en un equilibrio que depende de la concentración de proteína total. Estas interacciones dinámicas pueden ser estudiados tanto por equilibrio de sedimentación (SE) y la velocidad de sedimentación (SV) 10. Sin embargo, el primer método es más simple de realizar y se describe aquí. En SE, la centrifugación se realiza a una velocidad suficientemente baja para que se alcanza un equilibrio entre la difusión y sedimentación. En este punto, el perfil de equilibrio de una señal óptica (UV-VIS) como una función de la distancia radial, se puede analizar utilizando modelos termodinámicos pre-establecido para asociaciones 11.
ve_content "> En el presente trabajo, un estudio de equilibrio de sedimentación se presenta de la auto-asociación de una proteína de la membrana viral que forma canales iónicos. Debido a su hidrofobicidad, el experimento se ejecuta en presencia de detergente, y en este caso la densidad de disolvente tiene que ser adaptada a la del detergente. Sin embargo, el protocolo descrito sería idéntica en el caso de una proteína soluble en agua, excepto que se requeriría sin concordancia de densidad de disolvente.La proteína utilizada está codificada en el virus respiratorio sincitial humano (VRSh), un neumovirus envuelto en la familia Paramyxoviridae que causa la enfermedad de vías respiratorias bajas en los lactantes, los ancianos y las poblaciones inmunocomprometidas en todo el mundo 12. Hasta 64 millones de casos notificados de infección VRSh y 160.000 muertes ocurren cada año.
El genoma VRSh transcribe 11 proteínas, incluyendo las tres proteínas de la membrana f, g, y una pequeña hidrófoba (SH). Proteína SH está involucradoen la patogénesis de la infección por RSV. RSV carece del gen SH (RSVΔSH) era viable, causó la formación de sincitios y creció así como la de tipo salvaje (WT) virus 13-16. Sin embargo, el virus de RSVΔSH replicado 10 veces menos eficiente que la WT en el tracto respiratorio superior 15, 16. Además, el virus RSVΔSH fue atenuada en vivo en el ratón y modelos de chimpancés 13, 17.
La proteína SH es un (RSV subgrupo A) 64 o 65 (RSV subgrupo B) aminoácidos de longitud tipo II proteína integral de membrana que se acumula principalmente en las membranas de Golgi el compartimiento 18. Proteína SH tiene un solo predijo una hélice transmembrana (TM) de dominio 19 que está altamente conservada 20,21. Los dominios extramembrane C- y N-terminales están orientados lumenally / extracelularmente y el citoplasma, respectivamente.
Tanto dominio sintético TM (residuos 18-43) Y la proteína SH de longitud completa se ha demostrado para formar homopentamers en una variedad de detergentes. La forma homopentamérica es responsable de la actividad del canal en bicapas lipídicas planas 22,23. La orientación correcta de los monómeros TM en la bicapa lipídica se determinó primero utilizando sitio dicroísmo infrarrojos específica 23, que mostró su-22 para estar en un luminal, cerca de inter-helicoidal, orientación. La misma orientación dominio TM fue confirmada por estudios de RMN que reconstruyen la pentamérica a-helicoidal paquete de la proteína de longitud completa en dodecylphosphocholine (DPC) micelas 22. En este modelo de "micelas", un dominio TM helicoidal a- solo estaba flanqueado N-terminal por una a-hélice, y C-terminal por un b-horquilla extendida. Los dos residuos protonables de proteína SH, Su-22 y Su-51, se encuentran en el dominio TM (orientado lumenally), y en la punta de la extramembrane C-terminal de β horquilla (lejos del poro del canal), respectivamente. En un enviro bicellarnment, sin embargo, el TM α-hélice se extiende hasta Su-51, y los dos residuos de His son accesibles para el lumen del canal 24. La estructura de canal adopta una arquitectura similar a un embudo 22, en donde la región más estrecha (Ser-Cys-29 a 45) 22 se alinea con cadenas laterales hidrófobas (ILE-32, Ile-36, Ile-40 y Leu-44), y Ile-36 define el punto más estrecho en el lumen del canal. Su-22 se encuentra en la mayor apertura de este embudo, mientras que su-51 está en la punta de la abertura más pequeña.
En el presente trabajo, centrifugación analítica en un modo de equilibrio de sedimentación se ha utilizado para determinar si su protonación afecta a la estabilidad de la proteína SH pentámero. En este caso, la proteína SH fue solubilizado en detergente C14-betaína, que se ha utilizado previamente para mostrar que las formas de la proteína SH oligómeros pentaméricas 22.
Protocolo
Este protocolo se basa en los siguientes recursos, que han de ser remitidos para más detalles y consideraciones especiales 3, 25-28.
1. Densidad juego de micelas de detergente con 2 H 2 O
Nota: La densidad de la solución tampón debe ser adaptada a la densidad de las micelas de detergente. Agentes de ajuste de densidad comunes incluyen 2 H 2 O, H 2 18 O, 2 H 2 O 18, glicerol y sacarosa 29. H 2 18 O tiene la misma densidad que 2 H 2 O y puede ser una mejor opción si no se desea deuteración de protones intercambiables en la proteína. En este procedimiento, la densidad de 3- -propanesulfonate (C14SB) detergente (N, N-dimethylmyristylammonio) en 50 mM Tris pH 7,3, NaCl 100 mM será comparado con el 2 H 2 O. Como una aproximación inicial las siguientes concentraciones de2 H 2 O se utilizará: 10, 30, y 50% v / v.
1.1. Preparación de la muestra
- Preparar las siguientes soluciones madre y del filtro de esterilización a través de un filtro de jeringa de 0,2 micras: 50 ml de Tris 500 mM pH 7,3 y NaCl 1 M (solución tampón 10X); 1 ml C14SB 250 mM (solución de detergente 50X).
- Preparar 200 l de solución de la muestra por mezcla de 20 l solución tampón 10X, 50X solución de 4 l de detergente, 20 l 2 H 2 O (99,9%) y 156 l H 2 O. desionizada También preparar 200 l de solución de referencia mediante la mezcla de 20 l solución de tampón 10X, 20 l 2 H 2 O (99,9%) y 160 l H 2 O. desionizada
- Repita el paso 1.1.2 para las otras 2 H 2 O concentraciones, es decir, 30% y 50%, ajustando las cantidades 2 H 2 O y H 2 O apropiadamente.
1.2. Asamblea de 6 canales AUC células y carga de la muestra en las células.
Nota: Hay dos tipos de células AUC en función del método de carga de la muestra. Las células sin relleno externo tiene que ser cargado antes de sellar la celda, mientras externa-llenan las células se pueden cargar después de que las células están sellados. Asamblea de una célula AUC-llenar externa se ha descrito previamente 3. En este protocolo, se describe el montaje de una célula AUC de 6 canales sin llenado externo. La principal diferencia es que tiene anillos de los tornillos en ambos lados que deben ser apretados por separado, y no necesita tapones de vivienda (Fig. 1). La diferencia en los pasos de armado, se destacan a continuación.

Figura 1. Vista de una célula AUC de 6 canales sin relleno externo de despiece. Esta cifra ha sido modificado de Beckman Coulter An-50 y An Ti-Ti 60 Analytical Rotor, células, y el manual de usuario de contrapeso.
- Prepare dos montajes de ventana para cada celda AUC con ventana de zafiro en lugar de ventana de cuarzo (Fig. 2). Coloque la junta de la ventana en el soporte de la ventana. Doble ligeramente el revestimiento ventana y colocarlo en el soporte de la ventana de tal manera que la brecha se forma opuesta al chavetero titular ventana. Coloque la ventana de zafiro en el interior del forro ventana, alineando la marca con la ranura titular ventana.
Nota:. La ventana de cuarzo es compresible y por lo tanto producirá la refracción más luz a alta velocidad 28, 30 Por lo tanto para las mediciones de interferencia por encima de 30.000 rpm, como en este experimento coincidente densidad, se utilizan ventanas de zafiro. Una ventana de zafiro es más pesado que la ventana de cuarzo y tiene una "X" grabado en su lado.

Figura 2. Despiece of el conjunto de ventana. Esta cifra ha sido modificado desde Beckman Coulter An-50 y An Ti-Ti 60 Rotor Analítica, células, y el manual de usuario de contrapeso
- Coloque la carcasa de la célula con el número de pieza al revés. Con las ranuras alineadas con la llave de la vivienda, deslizarse en la carcasa de la célula en primer lugar una pieza central 6 del sector con el lado biselado hacia abajo, seguido de un conjunto de ventana con la ventana hacia abajo (Fig. 1, izquierda).
- Cubra ligeramente el anillo de hilos de rosca y anillo roscado lavadora con spinkote. Coloque una arandela anillo de tornillo en la parte superior del conjunto de ventana. Instale el anillo de tornillo en la carcasa ventana con la palabra "OUT" mirando hacia fuera. Apriete a mano el anillo roscado con la función de alineación celular.
- Utilizando la llave de torsión, apriete la rosca a sólo 60 pulgadas-libras.
- Coloque la celda con el número de pieza en posición vertical y se coloca a los 12 del mediodía. Load 120 l referencia en las filas izquierda y 110 l de muestra en las filas correctas. Asegúrese de que cada muestra y la referencia se empareja correctamente.
Nota: volumen de la muestra exacta no es crítica, pero la referencia necesita tener un poco más de volumen que la muestra (5-10 l) de modo que el menisco de la muestra será distinta. - Deslice con cuidado en el montaje de la cubierta celular una ventana con la ventana hacia abajo (Fig. 1, derecha). Tenga cuidado de no molestar a la celda en exceso y derramar el contenido.
- Repita el paso. 1.2 3 y 1.2. 4, apretando el segundo anillo de tornillo a 120 libras por pulgada. Invierta la celda y vuelva a apretar el primer anillo tornillo a 120 libras por pulgada.
- Cargar las células en el rotor, instalar el rotor en la centrífuga e instalar el monocromador de acuerdo con las instrucciones del fabricante 28.
Nota: Los detalles sobre este step también se puede encontrar en esta referencia 3.
1.3. Configuración de medición de interferencia
- Iniciar el software de interfaz de usuario para el instrumento AUC y realizar la configuración y calibración de láser radial para cada celda a 3000 rpm, de acuerdo con las instrucciones del fabricante, que se resumen brevemente en la siguiente etapa.
- Se ha alcanzado 1.3.1.1 Después de vacío suficiente (<100 micras), ejecute el centrifugar a 3000 rpm. Vista previa de la figura de interferencia en el software de interfaz de usuario y ajustar los parámetros del láser para obtener el mayor contraste.
- Crear una nueva creación de archivos (File | New File) especificando "Equilibrio" y "Interferencia" medición. Configuración de un método de equilibrio de sedimentación (botón "Método") funcione a 45.000 rpm o la velocidad máxima prevista para las muestras de proteínas, lo que sea mayor, con la temperatura del plazo a 20 ° C y recoger 1 lectura cada 15 minutos. Monitor equilibrio progreso mediante la apertura de los archivos de datos en HeteroAnalysis y seleccionando la función "Match" después de al menos 12 horas (aproximadamente durante la noche, Fig. 3).
Nota: una función similar también está disponible en SEDFIT (Opciones | Opciones de Carga | Test de aproximación al equilibrio).

Figura 3. El resultado de la función HeteroAnalysis Partido. La función de ajuste puede ser utilizado para monitorear el progreso de equilibrio comparando RMSD entre exploraciones sucesivas y el último análisis. Este ejemplo muestra consecución de equilibrio después de 8 horas como se indica por los valores asintóticos a RMSD eje x.
1.4. Análisis de los datos
- Para cada conjunto de muestras, trazar la pendiente del perfil de distribución radial en contra de la 2 H 2 O concentración.
Nota: La distribución será una muy superficial exponencial que apenfoques linealidad. Intercepción del eje X corresponde a la búsqueda de una concentración de 2 O 2 H. - Para obtener resultados más precisos, realizar el experimento en varias repeticiones. Alternativamente, repetir el experimento con una gama más reducida de 2 H 2 O concentración.
2. La sedimentación de equilibrio de SH en micelas C14SB
2.1. Parámetro de desplazamiento
- Calcular la densidad de tampón y la viscosidad, la proteína de volumen específico parcial y la velocidad de centrifugación mediante el uso de SEDNTERP. Para calcular la densidad de búfer y la viscosidad, seleccione Calcular en la sección "Buffer de datos Seleccione" e introduzca los componentes del tampón en consecuencia, incluyendo el D 2 O concentración.
- 2.1.1.1 Para calcular proteína volumen específico parcial, seleccione Calcular en la sección "V-bar" y escriba la secuencia de aminoácidos de la proteína. Especifique el tamaño oligomérica esperado más alto en "Hacer un oligómero de este monómero: N ="campo, en este caso N = 5. Calcular la velocidad al introducir los valores en el campo de RPM en la ventana principal hasta σ ≈ 1; esta es una regla de oro para garantizar una buena forma exponencial del perfil de distribución radial 25.
Nota: Los valores calculados para este experimento fue la siguiente: ρ = 1,03839 g / ml, η = 1,0267 cP = 0,7569 ml / g, ω 1 = 16.000 rpm. - Calcula las velocidades posteriores a seguir para garantizar la suficiente diferencia entre el perfil de distribución a una velocidad y el siguiente 25.
Nota: Esto también se puede hacer desde la función "Estimar rotor equilibrio acelera función" en SEDFIT, que tiene en cuenta la columna de la solución (volumen de llenado).
2.2. Preparación de las muestras
- Prepare la solución de 1 ml de referencia con 5 mM C14SB y el 32,3% 2 H 2 O como se determina a partir de experimento a juego densidad (sección 1), mediante la mezcla de 100 l SOLUTIO tampón 10Xn (paso 1.1.1), 20 l de 50X solución de detergente (etapa 1.1.1), 323 l 2 H 2 O (99,9%) y 527 l desionizada H 2 O.
- Disolver liofilizada, péptidos SH purificados por HPLC (expresión y purificación descrito previamente 31) en un disolvente apropiado tal como metanol o 50% v / v de acetonitrilo acuoso. Mida A280 de los péptidos disueltos en un microlitro escala espectrofotómetro UV / Vis y alícuota para tres muestras para dar un 280, 12 mm = 0,3, 0,5, y 0,8 (A 280, 10 mm = 0,25, 0,417, y 0,67) cada uno cuando se diluye hasta 130 l. Liofilizar las muestras durante la noche y volver a suspender en 130 l solución de referencia (paso 2.2.1) para dar las soluciones de muestra.
Nota: la proteína SH se puede detectar de UV / Vis absorbancia a 280 nm, ya que contiene residuos de Trp y Tyr. Las proteínas sin residuos aromáticos pueden ser detectados mediante el etiquetado con un cromóforo adecuado, usando un mutante Trp-que contiene, o utilizando interfieremediciones NCE en lugar de absorbancia. - Siga los pasos de la sección 1.2 para montar una célula AUC de 6 canales con ventanas de cuarzo. Cargar la muestra de mayor concentración (A 280, 12 mm = 0,8) en el canal más cercano al centro del rotor y la más baja concentración de la muestra (A 280, 12 mm = 0,3) más alejado del centro del rotor.
2.3. La creación de medidas de absorbancia
- Crear una nueva creación de archivos (File | New File) especificando "Equilibrium" y valoración "de Absorción". Especificar 280 nm como la longitud de onda del detector.
- Realice la calibración radial a 3000 rpm marcando "calibración radial antes de la primera exploración" en Opciones de análisis, especificando la recopilación de datos a baja resolución, por ejemplo, con tamaño de paso radial = 0.01 cm, repeticiones = 3 (baja resolución, rápido), y ejecutar una exploración individual . Una vez finalizada la exploración, desactive la opción.
- Configuración de un método de equilibrio de sedimentación ("methobotón ") d funcione a la primera velocidad calculada en el paso 2.1.3, a 20 ° C, y recoger 1 exploración cada 30 minutos. En "Detalle" 'cada célula, especifique la recopilación de datos a baja resolución como en el paso 2.3.2. Monitorear el progreso de equilibrio mediante la apertura de los archivos de datos en HeteroAnalysis y seleccionando la función "Match" después de al menos 18 horas (durante la noche aproximadamente, Fig. 4).
Nota: logro del equilibrio puede tomar tiempo sustancialmente más largo para la primera velocidad, mientras que las velocidades posteriores tardarán menos tiempo.

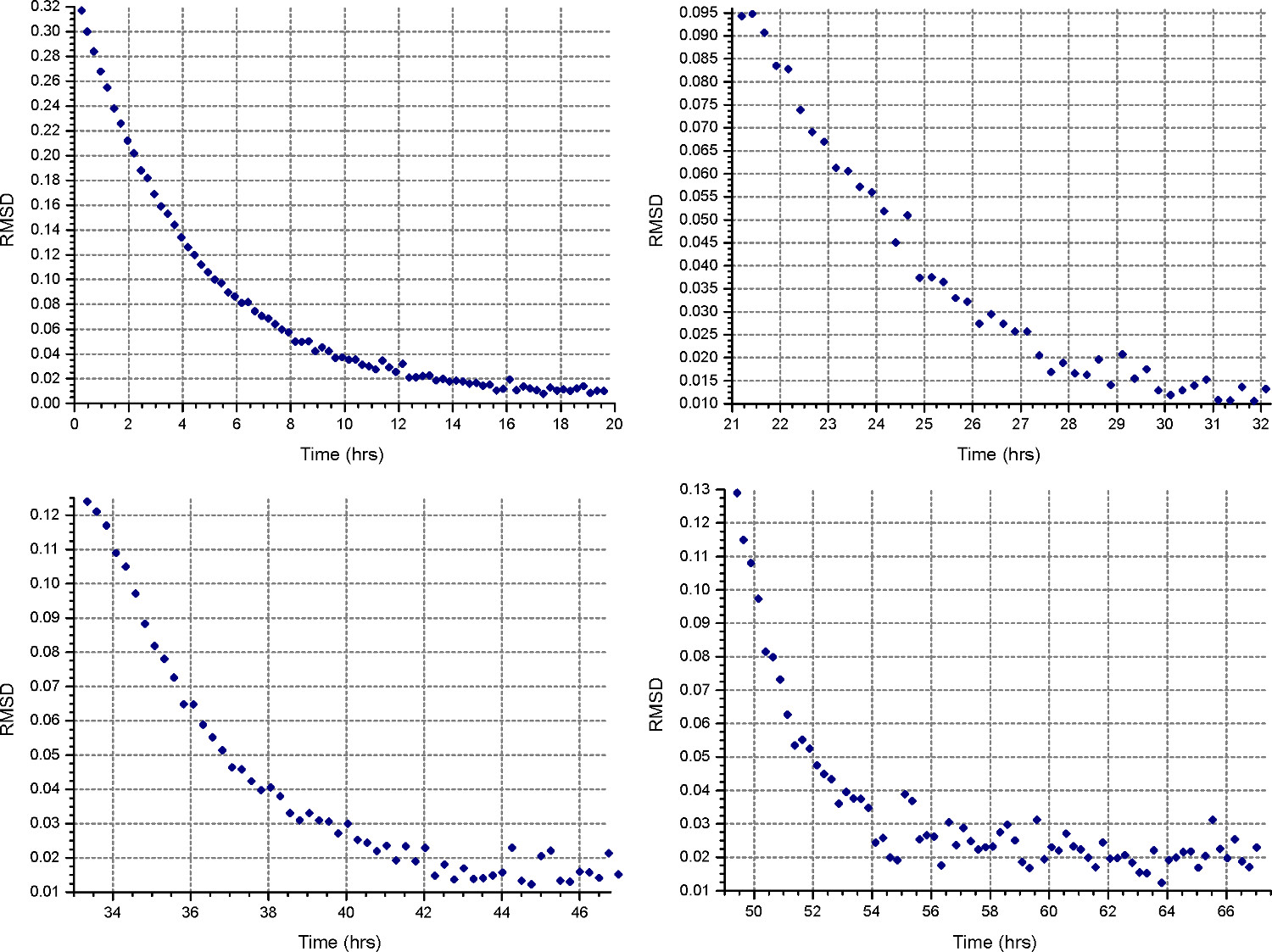
Figura 4. Los resultados de la función HeteroAnalysis Partido. La primera y segunda velocidades (arriba izquierda y derecha) parecen haber alcanzado el equilibrio, pero es mejor esperar unas horas más para estar seguro. En comparación, la tercera y cuarta velocidades (abajo a la izquierda y derecha) claramente han alcanzado el equilibrioen un tiempo más corto. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
- Una vez que se ha alcanzado el equilibrio, se recoge una sola exploración en alta resolución, por ejemplo, con tamaño de paso radial = 0,001 cm, repeticiones = 10 (alta resolución, lento).
- Una vez finalizada la exploración, repita el paso 2.3.3 y 2.3.4 para la siguiente velocidad.
- Opcionalmente, cuando el tiempo requerido para alcanzar el equilibrio para cada velocidad es conocida (calculada o por experiencia), configurar la exploración método de equilibrio de sedimentación para incluir todas las velocidades calculadas en el paso 2.1.3 y recoger 1 scan después de que el tiempo de equilibrio para cada la velocidad. En este caso, especifique la recopilación de datos en alta resolución "Detalle" 'cada célula.
2.4. Análisis de los datos en SEDFIT y SEDPHAT
Nota: Para más detalles y consideraciones en lector de análisis de datos se denomina to la siguiente página web: www.analyticalultracentrifugation.com.
- Exploraciones abiertas de alta resolución en SEDFIT (Datos | datos de equilibrio de sedimentación de carga) y se separaron los datos en 3 canales (estos corresponden a diferentes concentraciones para cada muestra; Opciones | Cargando Opciones | Guardar datos sin procesar de 6 canales en 3 subconjuntos).
- Vuelva a abrir los archivos de datos que pertenecen a la misma muestra y la misma concentración, pero diferentes velocidades en SEDFIT. Ajustar el menisco (línea vertical roja), fondo de la celda (línea vertical azul) y los límites de ajuste (líneas verticales verdes), y exportar los datos para su uso en SEDPHAT (datos | Exportar datos a SEDPHAT). Entrada de los parámetros calculados en el paso 2.1.1, así como del tipo de rotor y el tipo de pieza central a lo solicitado. Repita este paso para cada muestra y la concentración.
- Abre todos los datos de la misma muestra (todas las concentraciones y velocidades) en SEDPHAT y rellenar parámetros del experimento; un ejemplo se muestra en la Fig. 5.
Nota: cuando se añade D 2 O en el búfer, deut ración de protones intercambiables podría alterar significativamente el peso molecular de la proteína, especialmente para las proteínas solubles en agua. Las proteínas de membrana, especialmente los más pequeños, como la proteína SH, se ven menos afectados porque las regiones de membrana embebidos están protegidos de intercambio. Para corregir esto, de entrada el "buffer D fracción mol".
Nota: En este paso se recomienda guardar el conjunto de datos editado por separado seleccionando menú Datos | Copiar todos los datos y Guardar como una nueva configuración. - Seleccione un modelo y rellenar los parámetros globales para ese modelo.
Nota: Como ejemplo, el modelo de "Monómero-n-Mer autoasociación" y sus parámetros se muestran en la Fig. 6.

Figura 5. Un ejemplo de cómo rellenar los parámetros experimentales.
archivos / ftp_upload / 52404 / 52404fig6.jpg "/>
Figura 6. Un ejemplo de cómo rellenar los parámetros globales para el modelo de monómero-n-Mer Auto-Asociación.
- Ejecutar un Fit Global seleccionando menú Fit | Global Fit y esperar hasta que converge en forma. Anote (o tomar una captura de pantalla de) los resultados apropiados, especialmente la reducción de chi-cuadrado global y log K a valores. Extracto de otros datos como los datos de ajuste y los residuos de ajuste de menú Copia y Display | Información de la pantalla termodinámica.
- Regreso a los parámetros globales y comprobar M (1) para ajustarse al peso molecular del monómero y repita el paso 2.4.5. Anote el peso molecular amueblada y reducido chi-cuadrado global.
- Repita el paso 2.4.3 a 2.4.6 para cada modelo a analizar y comparar la calidad apta de cada modelo comparando el valor reducido de chi-cuadrado global, así como los residuos de montaje.
Nota: Los residuos de ajuste Pequeñas y aleatorios generalmente indica un buen ajuste, y el modelo que se adapta mejor haríatener la más pequeña reducción chi-cuadrado global. El peso molecular del monómero equipada y su valor de chi-cuadrado no deben diferir sustancialmente de la de la (teórico) de peso molecular fijo. - Calcular el intervalo de confianza para el registro obtenido Ka por valor seleccionando primero Estadísticas | Crítica chi-cuadrado para proyecciones de superficie error e introducir el intervalo de confianza deseado. A continuación, vaya a la Estadística | Generar proyección superficie de error 1-dimensional y anular la selección de log Ka en el diálogo de los parámetros globales para obtener los valores de chi-cuadrado para log Ka.
Se aconseja a los lectores consultar las siguientes fuentes (http://www.analyticalultracentrifugation.com/ SEDPHAT / statistics.htm) para más detalles sobre el método 32, así como ilustración de este método 33: Nota.
Resultados
El perfil radial de distribución de micelas de detergente C14SB en Tris 50 mM, NaCl 100 mM pH 7,3 formas exponenciales una muy superficial que podría ser instalado en un modelo lineal (Figura 7A). La pendiente de esta distribución está inversamente correlacionada con la concentración de D 2 O (Figura 7B). El punto donde la pendiente es cero, es decir, la correspondencia D 2 O concentración, se encontró que era 32,3%.
Ver...
Discusión
Este documento proporciona un protocolo experimental para la preparación y análisis de oligomerización de una pequeña proteína de membrana en detergente utilizando sedimentación equilibrio muestra. El protocolo descrito es igualmente válido -y simpler- para las proteínas solubles, como no se requiere la etapa de adaptación de la densidad. De hecho, el sistema está constituido por una mezcla de detergente y proteína. Para llevar a cabo estudios de sedimentación, el detergente debe ser invisible al campo gravi...
Divulgaciones
The authors have nothing to disclose.
Agradecimientos
This work has been funded by the National Research Foundation grant NRF-CRP4-2008-02 (J.T.) and Tier 1 grant RG 51/13.
Materiales
| Name | Company | Catalog Number | Comments |
| 3-(N,N-dimethylmyristylammonio)propanesulfonate | Sigma | T0807 | |
| Deuterium oxide 99.8% | Cambridge Isotope | DLM-4-99.8 | |
| An-50 Ti Rotor, Analytical, 8-Place | Beckman Coulter | 363782 | |
| An-60 Ti Rotor, Analytical, 4-Place | Beckman Coulter | 361964 | |
| Cell housing | Beckman Coulter | 334784 | |
| 12 mm six-channel centerpiece, epon charcoal-filled | Beckman Coulter | 331376 | |
| Window holder | Beckman Coulter | 305037 | |
| Window gasket | Beckman Coulter | 327021 | |
| Window liner | Beckman Coulter | 362329 | |
| Sapphire window | Beckman Coulter | 307177 | |
| Quartz window | Beckman Coulter | 301730 | |
| Screw-ring washer | Beckman Coulter | 362328 | |
| Screw ring | Beckman Coulter | 301922 | |
| Spinkote | Beckman Coulter | 306812 | |
| Torque stand assembly | Beckman Coulter | 361318 | |
| Counterbalance | Beckman Coulter | 360219 | |
| Cell alignment tool | Beckman Coulter | 362340 | |
| SEDNTERP | http://bitcwiki.sr.unh.edu/index.php/Main_Page | ||
| HeteroAnalysis | http://www.biotech.uconn.edu/auf/?i=aufftp | ||
| SEDFIT | http://www.analyticalultracentrifugation .com/sedfit.htm | ||
| SEDPHAT | http://www.analyticalultracentrifugation .com/sedphat/default.htm |
Referencias
- Laue, T. M., Stafford, W. F. Modern applications of analytical ultracentrifugation. Annu. Rev. Biophys. Biomol. Struct. 28, 75-100 (1999).
- Lebowitz, J., Lewis, M. S., Schuck, P. Modern analytical ultracentrifugation in protein science: A tutorial review. Protein Sci. 11, 2067-2079 (2002).
- Balbo, A., Zhao, H., Brown, P. H., Schuck, P. . Assembly, Loading, and Alignment of an Analytical Ultracentrifuge Sample Cell. , e1530 (2009).
- Rivas, G., Stafford, W., Minton, A. P. Characterization of heterologous protein-protein interactions using analytical ultracentrifugation. Methods-a Companion to Methods in Enzymology. 19, 194-212 (1999).
- Howlett, G. J., Minton, A. P., Rivas, G. Analytical ultracentrifugation for association and assembly the study of protein. Curr. Opin. Chem. Biol. 10, 430-436 (2006).
- MacGregor, I. K., Anderson, A. L., Laue, T. M. Fluorescence detection for the XLI analytical ultracentrifuge. Biophys. Chem. 108, 165-185 (2004).
- Phizicky, E. M., Fields, S. Protein-Protein Interactions - Methods for Detection and Analysis. Microbiol. Rev. 59, 94-123 (1995).
- Alexandrov, A. A facile method for high-throughput co-expression of protein pairs. Mol. Cell. Proteomics. 3, 934-938 (2004).
- Nooren, I. M. A., Thornton, J. M. Structural characterisation and functional significance of transient protein-protein interactions. J. Mol. Biol. 325, 991-1018 (2003).
- Ebel, C. Sedimentation velocity to characterize surfactants and solubilized membrane proteins. Methods. 54, 56-66 (2011).
- Minton, A. P. Quantitative characterization of reversible macromolecular associations via sedimentation equilibrium: an introduction. Exp. Mol. Med. 32, 1-5 (2000).
- Dowell, S. F. Respiratory syncytial virus is an important cause of community-acquired lower respiratory infection among hospitalized adults. J. Infect. Dis. 174, 456-462 (1996).
- Bukreyev, A., Whitehead, S. S., Murphy, B. R., Collins, P. L. Recombinant respiratory syncytial virus from which the entire SH gene has been deleted grows efficiently in cell culture and exhibits site-specific attenuation in the respiratory tract of the mouse. J. Virol. 71, 8973-8982 (1997).
- Fuentes, S., Tran, K. C., Luthra, P., Teng, M. N., He, B. Function of the respiratory syncytial virus small hydrophobic protein. J. Virol. 81, 8361-8366 (2007).
- Jin, H. Recombinant respiratory syncytial viruses with deletions in the NS1, NS2, SH, and M2-2 genes are attenuated in vitro and in vivo. Virology. 273, 210-218 (2000).
- Karron, R. A. Respiratory syncytial virus (RSV) SH and G proteins are not essential for viral replication in vitro: clinical evaluation and molecular characterization of a cold-passaged, attenuated RSV subgroup B. Proc. Nat. Acad. Sci. USA. 94, 13961-13966 (1997).
- Whitehead, S. S. Recombinant respiratory syncytial virus bearing a deletion of either the NS2 or SH gene is attenuated in chimpanzees. J. Virol. 73, 3438-3442 (1999).
- Rixon, H. W. The small hydrophobic (SH) protein accumulates within lipid-raft structures of the Golgi complex during respiratory syncytial virus infection. J. Gen. Virol. 85, 1153-1165 (2004).
- Collins, P. L., Mottet, G. Membrane orientation and oligomerization of the small hydrophobic protein of human respiratory syncytial virus. J. Gen. Virol. 74, 1445-1450 (1993).
- Collins, P. L., Olmsted, R. A., Johnson, P. R. The small hydrophobic protein of human respiratory syncytial virus: comparison between antigenic subgroups A and B. J. Gen. Virol. 71, 1571-1576 (1990).
- Chen, M. D., Vazquez, M., Buonocore, L., Kahn, J. S. Conservation of the respiratory syncytial virus SH gene. J. Infect. Dis. 182, 1228-1233 (2000).
- Gan, S. W. The small hydrophobic protein of the human respiratory syncytial virus forms pentameric ion channels. J. Biol. Chem. 287, 24671-24689 (2012).
- Gan, S. W., Ng, L., Xin, L., Gong, X., Torres, J. Structure and ion channel activity of the human respiratory syncytial virus (hRSV) small hydrophobic protein transmembrane domain. Protein Sci. 17, 813-820 (2008).
- Li, Y. Inhibition of the Human Respiratory Syncytial Virus Small Hydrophobic Protein and Structural variations in a bicelle environment. J. Virol. 88 (22), 11899-914 (2014).
- Burgess, N. K., Stanley, A. M., Fleming, K. G. Determination of membrane protein molecular weights and association equilibrium constants using sedimentation equilibrium and sedimentation velocity. Meth. Cell. Biol. 84, 181-211 (2008).
- Cole, J. L., Lary, J. W., Moody, T. P., Laue, T. M. Analytical Ultracentrifugation: Sedimentation Velocity and Sedimentation Equilibrium. Meth. Cell. Biol. 84, 143-179 (2008).
- Fleming, K. G. Determination of membrane protein molecular weight using sedimentation equilibrium analytical ultracentrifugation. Curr. Protoc. Prot. Sci. 53, 17.12.11-17.12.13 (2008).
- . . An-50 Ti and An-60 Ti Analytical Rotor, Cells, and Counterbalance. , (2005).
- Mayer, G. Studying membrane proteins in detergent solution by analytical ultracentrifugation: Different methods for density matching. Prog. Colloid Polym. Sci. 113, 176-181 (1999).
- Laue, T. Ch. 20.3. Current Protocols in Protein Science. 20, 20.23.21-20.23.13 (2001).
- Gan, S. W. The Small Hydrophobic Protein Of The Human Respiratory Syncytial Virus Forms Pentameric Ion Channels. J. Biol. Chem. 287, 24671-24689 (2012).
- Bevington, P. R., Robinson, D. K. . Data reduction and error analysis for the physical sciences. 336, (1969).
- Schuck, P., Radu, C. G., Ward, E. S. Sedimentation equilibrium analysis of recombinant mouse FcRn with murine IgG1. Molecular Immunology. 36, 1117-1125 (1999).
- Gan, S. W., Vararattanavech, A., Nordin, N., Eshaghi, S., Torres, J. A cost-effective method for simultaneous homo-oligomeric size determination and monodispersity conditions for membrane proteins. Anal. Biochem. 416, 100-106 (2011).
- Montserret, R. NMR structure and ion channel activity of the p7 protein from hepatitis C virus). J. Biol. Chem. 285, 31446-31461 (2010).
- Stouffer, A. L., DeGrado, W. F., Lear, J. D. Analytical Ultracentrifugation Studies of the Influenza M2 Homotetramerization Equilibrium in Detergent Solutions. Progr Colloid Polym Sci. 131, 108-115 (2006).
- Sorkin, A., von Zastrow, M. Signal transduction and endocytosis: Close encounters of many kinds. Nat. Rev. Mol. Cell Biol. 3, 600-614 (2002).
- Gan, S. W., Ng, L., Lin, X., Gong, X., Torres, J. Structure and ion channel activity of the human respiratory syncytial virus (hRSV) small hydrophobic protein transmembrane domain. Protein science : a publication of the Protein Society. 17, 813-820 (2008).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados