
Method Article
Observación en tiempo real de la reacción de intercambio de filamento de ADN mediada por Rad51
En este artículo
Resumen
Se desarrollaron fluorescencia resonancia energética basada en la transferencia en tiempo real observación sistemas de la DNA del filamento reacción de intercambio mediada por Rad51. Utilizando los protocolos aquí presentados, que son capaces de detectar la formación de intermedios de reacción y su conversión en productos, mientras que analizar también la cinética enzimática de la reacción.
Resumen
La reacción de intercambio de filamento de ADN mediada por Rad51 es un paso crítico de recombinación homóloga. En esta reacción, Rad51 forma un filamento de nucleoproteína de ADN monocatenario (ssDNA) y capta ADN de doble cadena (dsDNA) no específicamente para interrogar para una secuencia homóloga. Después de encontrar homología, Rad51 cataliza el intercambio de filamento de ADN para mediar el emparejamiento de la ssDNA con el filamento complementario de la dsDNA. Esta reacción es altamente regulada por numerosas proteínas cómplice en vivo. Aunque ensayos bioquímicos convencionales han sido utilizados con éxito para examinar el papel de estas proteínas accesorias en vitro, análisis cinético de la formación intermedia y su progresión en un producto final ha demostrado ser un reto debido a la inestable y transitoria naturaleza de los intermedios de reacción. Para observar estos pasos de la reacción en solución, transferencia de energía de resonancia de fluorescencia (FRET)-basado en la observación en tiempo real se establecieron sistemas de esta reacción. Análisis cinético de las observaciones en tiempo real muestran que la reacción de intercambio de filamento de ADN mediada por Rad51 obedece a un modelo de tres pasos de reacción que implica la formación de un ADN de tres-filamento intermedio, la maduración de este intermedio y la liberación de ssDNA de el intermedio maduro. El complejo de Swi5-Sfr1, una proteína accessary conservado en eucariotas, aumenta fuertemente los pasos segundo y terceros de esta reacción. El traste-ensayos aquí presentados nos permiten descubrir los mecanismos moleculares a través de la cual recombinación accessary proteínas estimulan la actividad de intercambio del filamento de ADN de Rad51. El objetivo principal de este protocolo es mejorar el repertorio de técnicas disponibles para los investigadores en el campo de la recombinación homóloga, particularmente aquellos que trabajan con proteínas de especies distintas de Schizosaccharomyces pombe, para que el conservación evolutiva de los resultados presentados en este documento puede ser determinada.
Introducción
Recombinación homóloga (HR) facilita la mezcla de información genética entre dos moléculas diferentes de ADN. Recursos humanos es esencial para dos fenómenos biológicos fundamentales: la generación de diversidad genética durante la gametogénesis1 y la reparación de ADN de doble cadena rompe (distritales)2 durante la mitosis. Distritales son la forma más severa de lesión del ADN y constituyen una rotura en el cromosoma. Incorrecta reparación de distritales puede causar extensos cambios cromosómicos e inestabilidad genómica, que son dos características distintivas del cáncer3.
La reacción de intercambio del filamento de ADN es la fase central de recursos humanos. La proteína Rad51, que es miembro de la familia altamente conservado de RecA-tipo de recombinasas, es la proteína clave que cataliza esta reacción en eucariotas4,5. En esta reacción, Rad51 se une al ADN monocatenario (ssDNA) generado por el proceso de nucleolytic del fin OSD y formas un complejo de nucleoproteína helicoidal denominado filamento presináptico. Este filamento coge ADN intacto de la doble cadena (dsDNA) producir para buscar para una secuencia homóloga. Cuando el filamento encuentra una secuencia homóloga, se forma una reacción intermedia que contiene tres ADN y el filamento de Rad51 media intercambio del filamento dentro de esta estructura6,7,8. Para lograr esta reacción eficientemente, Rad51 requiere varias clases de proteínas accesorias como BRCA1 y BRCA2, los productos de mama cáncer susceptibilidad genes9,10.
Comprensión cómo factores accesorios regulan Rad51 es un paso integral en el descubrimiento de las causas de la inestabilidad genómica durante tumorigenesis. Aunque mucha investigación se refiere a los efectos de estos factores en la formación del filamento presináptico y estabilidad11,12,13,14,15,16, el contribución de estos factores a la formación del intermedio de tres hebras y su transformación en el producto final sigue siendo confuso. Observando estos pasos de reacción a través de experimentos bioquímicos convencionales es muy difícil porque el tres-filamento intermedio es inestable y propenso a colapsar por manipulaciones experimentales comunes como desproteinización de muestras o electroforesis.
Para superar este problema, adaptamos dos sistemas anteriormente desarrollados observación en tiempo real de la reacción de intercambio de filamento de ADN mediante la transferencia de energía de resonancia de fluorescencia (FRET): emparejamiento de filamento de ADN y DNA filamento desplazamiento ensayos17, 18 (figura 1). En el filamento de la DNA emparejamiento ensayo, Rad51 forma un filamento presináptico con fluoresceína amidite (FAM) - etiquetado ssDNA y entonces homólogo carboxi-x-rodamina (ROX) - etiquetado dsDNA se agrega para iniciar la reacción de intercambio del filamento. Cuando el filamento coge el dsDNA marca ROX y forma el intermedio de tres hebras, las dos fluoróforos en proximidad cercana y emisión de fluorescencia de FAM se apaga por ROX (figura 1A). En el análisis de desplazamiento de hebra de ADN, un filamento presináptico en sin etiqueta ssDNA se incuba con ROX y FAM dsDNA doble etiquetado. Cuando cambio de cadena se ha completado y la FAM etiquetado ssDNA se libera de la línea de tres intermedios, la emisión de FAM aumenta porque la FAM ya no está en proximidad cercana a ROX (figura 1B). Estos ensayos nos permiten observar la formación de productos intermedios de tres hebras y su transformación en productos finales en tiempo real sin interferencias a la reacción.
Usando este sistema de observación en tiempo real, encontramos que la reacción de intercambio de filamento de ADN mediada por Rad51 procede en tres pasos incluyendo la formación de la primera reacción intermedia (C1), transición de la primera intermedia hacia un segundo intermedio (C2) y liberación de ssDNA de C219. También encontramos esa levadura de la fisión (S. pombe) Swi5-Sfr1, que es un evolutivamente conservado Rad51 proteína accesorio complejo13,16,20,21,22, estimula la transición de C1-C2 y liberación de ssDNA de C2 de una manera que depende de la hidrólisis de ATP por Rad5119.
Se desconoce si estos resultados son conservados evolutivamente. Este protocolo se proporciona con la esperanza de que los investigadores en el campo de recursos humanos, especialmente aquellos que trabajan con proteínas de microorganismos distintos de S. pombe, pueden aplicar estas técnicas para determinar la medida en que el mecanismo molecular de Rad51 conducido intercambio del filamento se conserva. Además, estas técnicas han probado altamente acertadas en la determinación de la función de Swi5-Sfr1 de S. pombe . Por lo tanto, es una predicción racional que estas técnicas serán invaluables en el descubrimiento de los roles precisa de otros factores accesorios de HR.
Protocolo
1. preparación de sustratos de DNA y proteínas
- Purificar proteínas S. pombe Rad51 y Swi5-Sfr1 a la homogeneidad (juzgado mediante tinción de Coomassie), divulgado previamente13,21.
- Preparación de sustratos de DNA oligonucleotide enumerados en cuadro 118.
Nota: Los oligonucleótidos fueron adquiridos (véase Tabla de materiales) y sintetizado en grado HPLC. Para el filamento de la DNA reacción, oligonucleótidos de emparejamiento 16FA(-), 16A (-) _40bp y 16AR (+) _40bp se requiere. Para el análisis de desplazamiento de ADN, oligonucleótidos 16A(-), 16FA _40bp (-) y 16AR (+) _40bp son necesarios (figura 1 y tabla 1). Todas las concentraciones de ADN en este protocolo se refieren a concentraciones en comparación con las concentraciones de nucleótidos del fragmento. - Para formar el donante dsDNA, mezclan cantidades equimolares de hebras complementarias en un tubo PCR de paredes delgadas con recocido buffer (10 mM Tris-HCl, pH 7,5, 100 mM NaCl, 10 mM de MgCl2), asegurando un volumen total superior a 20 μl. realizar esta mezcla en un estante del metal del prechilled en hielo (entre 2 ° C y 4 ° C).
Nota: La combinación de oligonucleótidos para el análisis de emparejamiento son 16A _40bp (-) y 16AR (+) _40bp. Para el análisis de desplazamiento, Recueza 16FA oligonucleótidos (-) _40bp y 16AR (+) _40bp. - Calentar la mezcla recocida a 90 ° C durante 5 minutos y enfriar durante 3 h a 30 ° C utilizando una máquina PCR. Tienda el recocido ADN a-20 ° C.
2. ADN hebra emparejamiento y ensayos de desplazamiento
- Realizar análisis de emparejamiento del filamento de la DNA.
- Preparar 1,6 mL de tampón de reacción A (30 mM HEPES-KOH de pH 7,5, 1 mM Ditiotreitol [TDT], 15 mM MgCl2, 0.25 mM ATP, 0,1 mg/mL albúmina de suero bovino [BSA] y 0.0075% polyoxyethylenesorbitan monolaurato) que contiene 36 nM 16FA(-) en una microcentrífuga de 2,0 mL tubo de plástico (polipropileno) y previamente lo incube a 37 ° C durante 5 minutos.
- Para formar filamentos de ssDNA Rad51, añadir la proteína Rad51 a una concentración final de 1.5 μm en el tampón de reacción previamente incubados e incubar a 37 ° C durante 5 minutos.
- Añadir proteínas de Swi5-Sfr1 a la mezcla a una concentración final de 0,15 μm e incubar a 37 ° C por un 5 minutos más.
- Tomar 1,5 mL de la mezcla y transferencia en una cubeta de cuarzo de 1.0 x 1.0 cm que contiene un agitador magnético y coloque la cubeta en un spectrofluorometer. Configurar el controlador de temperatura peltier del espectrofotómetro a 37 ° C y el agitador magnético a 450 rpm para asegurar una mezcla rápida de la muestra inyectada.
- Comienza la medición de la emisión de fluorescencia de FAM en 525 nm (ancho de banda: 20 nm) sobre excitación en 493 nm (ancho de banda: 1 nm). Recolectar datos cada segundo.
- Después de comenzar la medición de 100 s, inyectar dsDNA ROX etiquetado donante a una concentración final de 36 nM en la mezcla utilizando una jeringa y medir el cambio en emisiones a intervalos de 1 s por un 30 min más.
- Realizar análisis de desplazamiento de hebra de ADN.
- Preparar 1,6 mL de tampón de reacción A que contiene 36 nM 16A(-) en un plástico de microcentrífuga de 2.0 mL tubo y previamente lo incube a 37 ° C durante 5 minutos.
- Forman filamentos de ssDNA de Rad51 en presencia de Swi5-Sfr1 a 37 ° C, como se describe en pasos 2.1.2. y 2.1.3.
- Tomar 1,5 mL de la mezcla y transferir a la cubeta de cuarzo que contiene un agitador magnético y coloque la cubeta en el espectrofotómetro, tal como se describe en el paso 2.1.4.
- Comienza la medición de la emisión de fluorescencia y después 100 s, inyectar FAM y etiquetados como ROX dsDNA de donante como se describe en el paso 2.1.6. Medir el cambio en la emisión de fluorescencia a intervalos de 1 s por un 30 min más.
3. Análisis de datos experimentales de la sincronización y análisis de desplazamiento
- Estimar la máxima eficiencia de traste.
- Preparar 16FA(-) recocido con 16AR (+) _40bp y 16FA (-) _40bp recocido con 16AR (+) _40bp con el mismo procedimiento descrito en los pasos 1.3 y 1.4.
- Preparar 130 μL de tampón de reacción A que contiene 36 nM de cualquiera de los dos 16FA(-), 16FA(-) recocido con 16AR (+) _40bp, 16FA (-) _40bp recocido con 16AR (+) _40bp o 16FA _40bp (-) en una cubeta de cuarzo de 0.2 x 1.0 cm.
- Coloque la cubeta en el spectrofluorometer e incubar a 37 ° C durante 5 minutos.
- Medir espectros de fluorescencia de 500 a 600 nm con excitación a 493 nm.
- Para probar el efecto de Rad51 en la emisión de FAM y amortiguamiento de FAM por ROX, añadir Rad51 a una concentración final de 1.5 μM a la mezcla e incubar a 37 ° C durante 5 minutos.
- Medir espectros de fluorescencia de 500 a 600 nm con excitación a 493 nm.
- Calcular la máxima eficiencia de traste (Emáxima) mediante la ecuación descrita a continuación:
Emáxima = (intensidad de fluorescencia a 525 nm de dsDNA con ROX y FAM) / (intensidad de fluorescencia en 525 nm FAM etiquetado ssDNA)
- Analizar los datos experimentales de la prueba de desplazamiento.
- Para convertir el cambio en la fluorescencia observada en el ensayo de desplazamiento para el cambio en la cantidad de producto final, normalizar los datos experimentales obtenidos de este ensayo utilizando la ecuación descrita a continuación, donde Fmateria prima es la intensidad de la fluorescencia de datos en bruto y Fnormalizada es el cambio en la fluorescencia Indicador calculado por la siguiente ecuación.
Fnormalizada = ([Fmateria prima al tiempo x]-[Fraw a tiempo 0]) / (([Fraw a tiempo 0] / Emáxima)-[Fraw a tiempo 0])
Fmateria prima a tiempo 0 es la fluorescencia promedio durante los primeros 5 s después del tiempo muerto (es decir, el tiempo requerido para la mezcla después de una injectant se introduce en la cubeta). - Para excluir los efectos de fotoblanqueo y desplazamiento espontáneo, restar Fnormalizada sin la proteína de Fnormalizada de la muestra para obtener FD, que es el cambio en la cantidad de producto final en este ensayo.
FD = [Fnormalizada de muestra] - [Fnormalizada sin proteína]
- Para convertir el cambio en la fluorescencia observada en el ensayo de desplazamiento para el cambio en la cantidad de producto final, normalizar los datos experimentales obtenidos de este ensayo utilizando la ecuación descrita a continuación, donde Fmateria prima es la intensidad de la fluorescencia de datos en bruto y Fnormalizada es el cambio en la fluorescencia Indicador calculado por la siguiente ecuación.
- Analizar los datos experimentales de la prueba enlace.
- Normalizar datos experimentales obtenidos el análisis apareado mediante la ecuación descrita a continuación, donde Fcruda intensidad de fluorescencia de datos brutos y Fnormalizada es el cambio en la fluorescencia Indicador calculado por la siguiente ecuación.
Normalizado de F = (Fmateria prima al tiempo x) / (Fraw a tiempo 0)
Fmateria prima a tiempo 0 es la fluorescencia promedio durante los últimos 20 s antes de iniciar la reacción mediante la inyección del sustrato de dsDNA. - Para convertir el cambio en la fluorescencia en cambio en la cantidad de sustrato y excluir los efectos de fotoblanqueo y sincronización espontánea, normalizar Fnormalizada de muestra utilizando la ecuación descrita a continuación, donde FP es el cambio en la cantidad de sustrato en este ensayo.
FP = 1 - (([Fnormalizada sin proteína] - [Fnormalizada de muestra]) / [1-Emáxima]) - Para estudiar la cinética de la reacción de intercambio del filamento de ADN, realizar análisis de regresión no lineal de la última plaza de la reacción de sincronización utilizando el programa análisis del23 (véase Tabla de materiales).
- Preparar un archivo en formato .txt que contiene los datos del curso tiempo de FP.
- Inicie el programa y pegar el script en el Archivo de código suplementario en una ventana del programa:
- Iniciar el análisis de regresión no lineal de la última plaza. Los resultados de este análisis se mostrará en la misma ventana.
- Normalizar datos experimentales obtenidos el análisis apareado mediante la ecuación descrita a continuación, donde Fcruda intensidad de fluorescencia de datos brutos y Fnormalizada es el cambio en la fluorescencia Indicador calculado por la siguiente ecuación.
Resultados
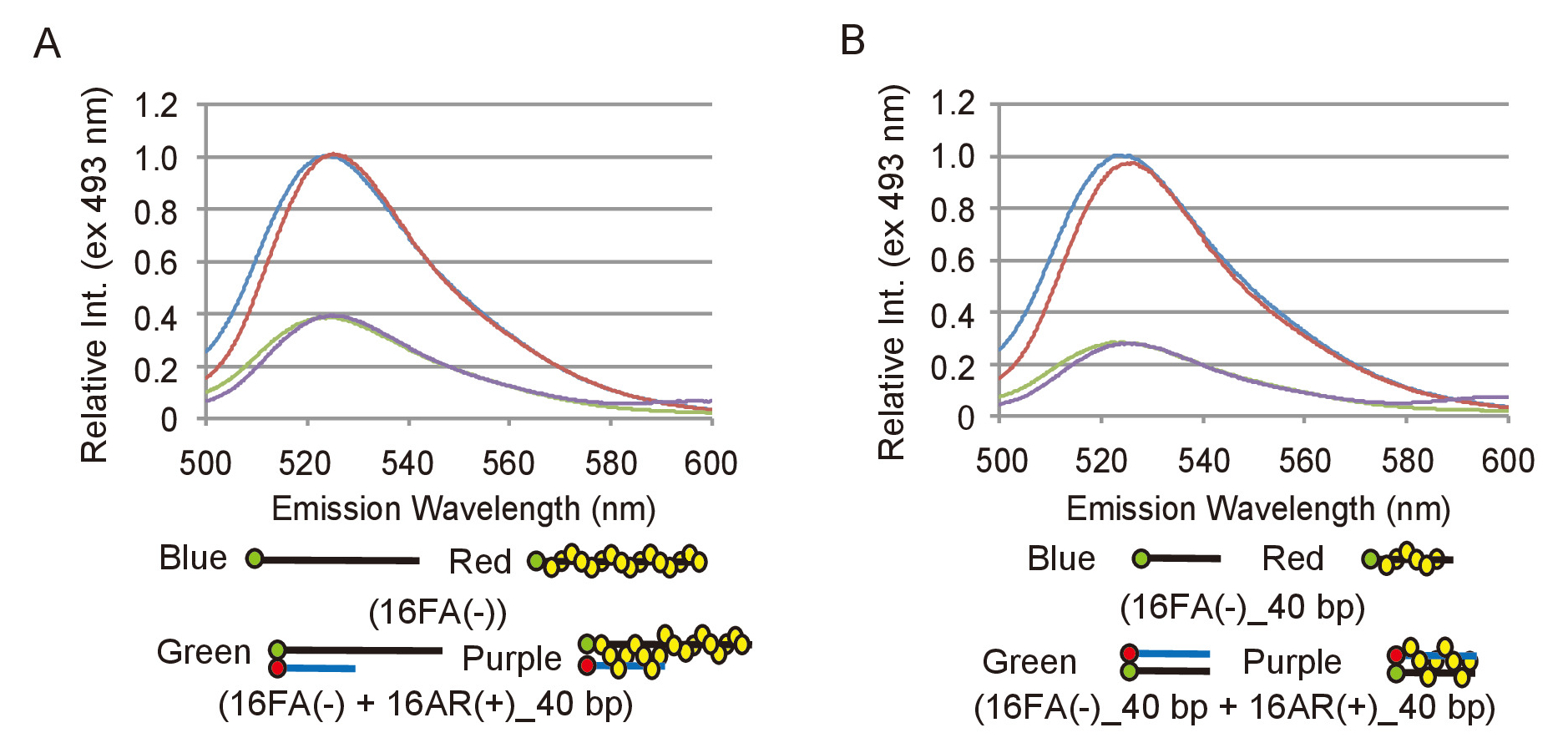
Con el fin de analizar con eficacia los datos experimentales de los ensayos de emparejamiento y el desplazamiento, es necesario definir cómo un cambio en la emisión de fluorescencia de FAM corresponde a una conversión de sustratos de ADN en los productos. Para lograr esto, debe determinarse la gama relevante de intensidad de fluorescencia. Para el análisis de asociación, la emisión de fluorescencia de 16FA(-), que corresponde al sustrato de ssDNA, se compara con la emisión de 16FA(-) recocido con 16AR (+) _40bp, que corresponde a los productos finales de esta reacción (figura 2A). Esto equivale a la máxima eficiencia de trastes y por lo tanto, la máxima reducción en la intensidad de la fluorescencia que sería esperar si todo substrato de ssDNA se convirtió en el producto de dsDNA. Para el análisis de desplazamiento, la emisión de 16FA (-) _40bp recocido con 16AR (+) _40bp, que corresponde al sustrato, se compara con la emisión de 16FA _40bp (-), que corresponde al producto final (figura 2B). En este caso, el aumento máximo de intensidad de fluorescencia de FAM transmite un escenario en el que todo el sustrato de dsDNA se convierte en producto de ssDNA. S. pombe Rad51 no afectó la emisión de fluorescencia de la FAM o amortiguamiento eficiencia de FAM por ROX en ambos ensayos (figura 2). La eficiencia máxima del traste debe ser a medida con cada nueva preparación de oligonucleótidos es dependiente en la eficacia del etiquetado de los oligonucleótidos.
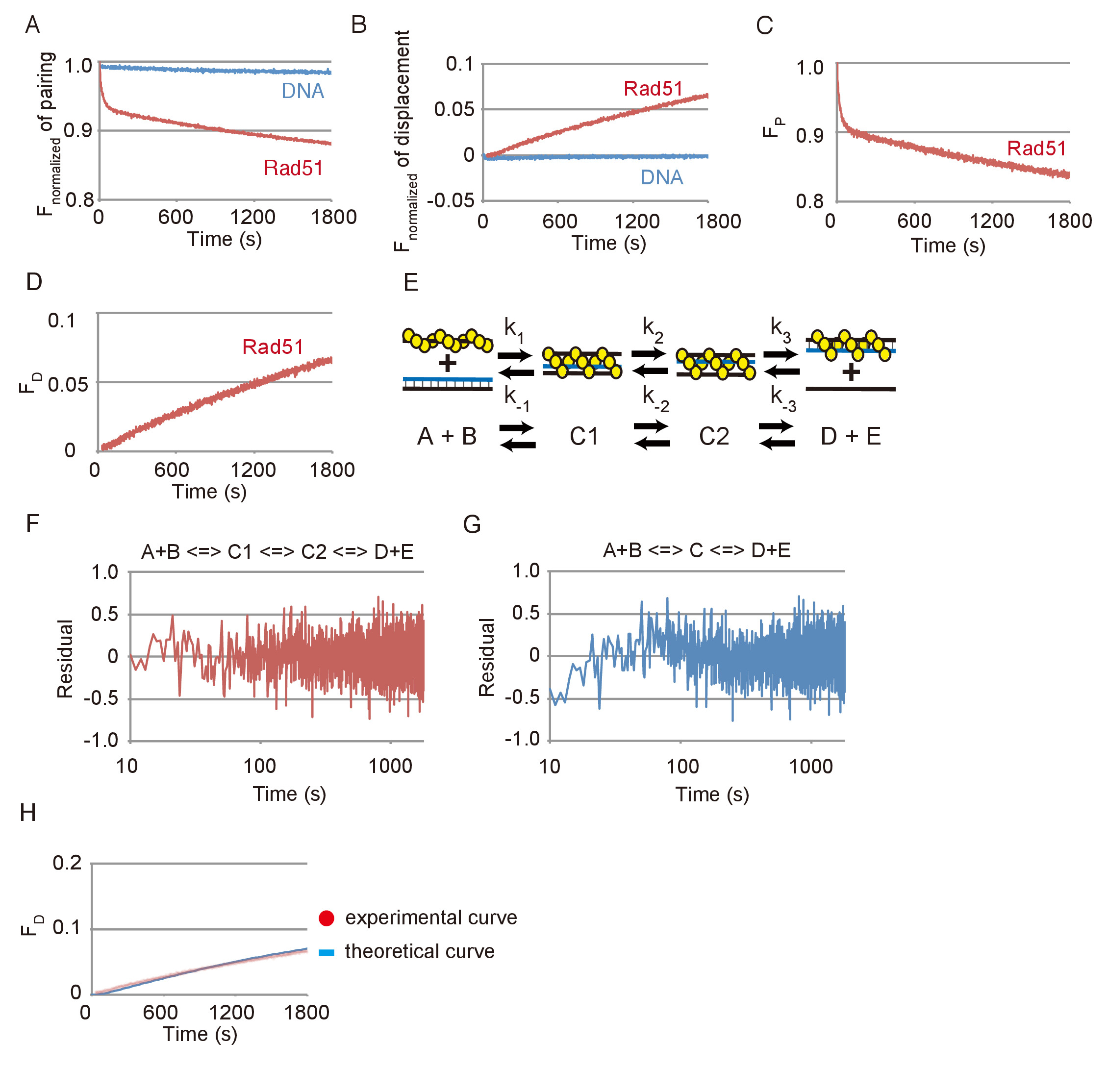
Datos representativos de las reacciones de asociación y desplazamiento de filamento ADN se muestran en la figura 3. Los efectos de reacciones espontáneas entre DNAs de sustrato y photobleaching eran pequeños en ambos ensayos, según lo revelado por los insignificantes cambios considerados en la emisión de FAM sin Rad51 en comparación con los cambios substanciales con Rad51 (Figura 3A y Figura 3B). Base de datos que se muestra en la Figura 3A o 3B de la figura, el cambio en la fluorescencia fue convertido en el cambio en la cantidad de sustrato (FP) o el producto final (FD), respectivamente, mediante las ecuaciones descritas en (pasos) 3.2.2 o 3.3.2 Figura 3 y figura 3D).
La reacción de emparejamiento fue simulada utilizando un modelo secuencial de tres pasos de reacción, que consiste en la formación de los primeros tres hebras intermedios (C1), la transición de la primera intermedia en el segundo intermedio (C1 a C2) y la liberación de ssDNA desde el segundo intermedio para formar los dos productos (D + E) (figura 3E). Para probar si los residuos entre los datos experimentales de la hebra de DNA emparejamiento ensayo y una curva teórica obtenida por la simulación de la simulación mediante un ajuste del modelo secuencial de tres pasos de reacción los datos experimentales, fueron calculados (figura 3F). Además, residuos entre la reacción de emparejamiento y una curva teórica generada utilizando un modelo de reacción de dos etapas secuenciales fueron también calculados (figura 3). Los residuos de la reacción de emparejamiento y el modelo de dos etapas muestran una desviación sistemática en la primera etapa, mientras que los residuos de la reacción de emparejamiento y el modelo de tres pasos no muestran tal desviación. Esto indica que el modelo de tres pasos es un mejor ajuste que el modelo de dos etapas para simular la reacción de enlace.
Para probar si el modelo de tres pasos es consistente con la reacción de desplazamiento que detecta el paso finales de intercambio del filamento de ADN, nos genera una curva teórica de la reacción de desplazamiento utilizando los parámetros cinéticos obtenidos de la simulación de la Asociación la reacción se muestra en la figura 3 y la compararon con los datos experimentales de la reacción de desplazamiento se muestra en la figura 3D (figura 3 H). La curva teórica ajustar los datos experimentales de la prueba de desplazamiento. De estos resultados, concluimos que la simulación utilizando el modelo de tres pasos es evaluar razonablemente la reacción de intercambio de filamento de ADN mediada por Rad51.
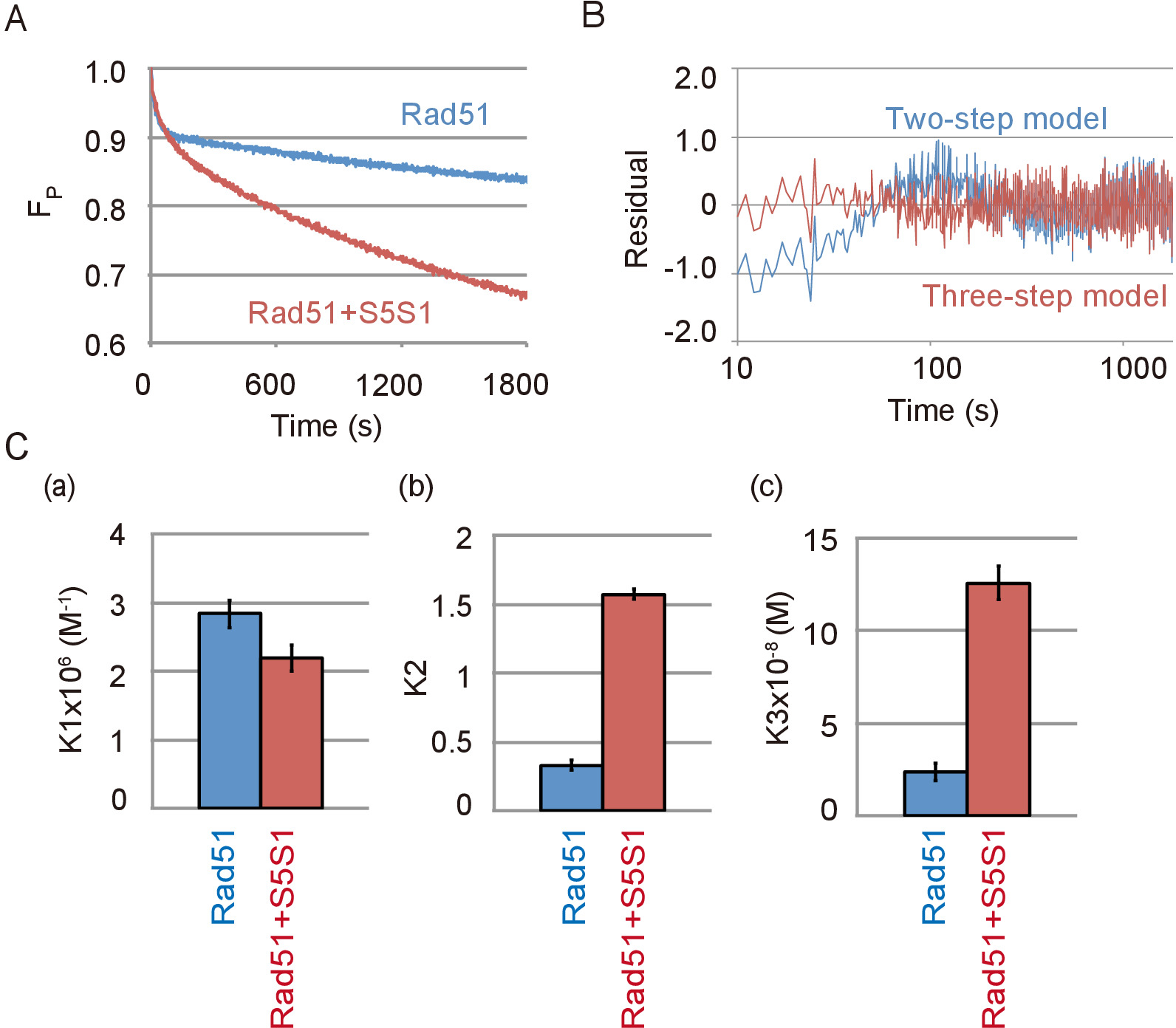
Datos representativos de la cadena de ADN maridaje con Rad51 y el complejo de Swi5-Sfr1, una proteína del accessary de Rad51, la reacción se muestran en la Figura 4A. El complejo de Swi5-Sfr1 había estimulado fuertemente la actividad Asociación de Rad51. Como se ha visto en la ausencia de Swi5-Sfr1, la reacción maridaje mejor ajuste el modelo de tres pasos que el modelo de dos etapas en la presencia de Swi5-Sfr1 (Figura 4B). A través de la simulación de la reacción utilizando el modelo de tres pasos, calcularon las constantes de equilibrio de la reacción de cada paso de la reacción con o sin Swi5-Sfr1. Las constantes de equilibrio de la reacción indicaron que el complejo de Swi5-Sfr1 no estimula el primer paso de reacción (figura 4, panel de un), en el que un intermedio de tres hebras está formado, pero estimula fuertemente la transición de C1-C2 ( Figura 4, panel b) y la liberación de ssDNA del intermedio C2 (figura 4, panel c).

Figura 1: diseño Experimental de la DNA del filamento ensayos de emparejamiento y el desplazamiento. Esquemas de la DNA del filamento emparejamiento (A) y desplazamiento (B) ensayos. Círculos amarillos representan monómeros Rad51. Verde los círculos que contienen "F" y círculos rojos que contienen la "R" representan fluoresceína amidite (FAM) y carboxi-X-rodamina (ROX), respectivamente. Filamentos de la DNA de negro son idénticas en secuencia y complementario al azul filamentos de la DNA. Delgadas líneas negras con puntas de flecha señalan el nombre de cada oligonucleótido, como se muestra en la tabla 1. Esta figura ha sido adaptada de Ito et al. 19 y modificado. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: medición de la eficiencia máxima del traste de los ensayos de emparejamiento y el desplazamiento. (A) comparación de espectros de fluorescencia entre el substrato de ssDNA, 16FA(-) y el producto de dsDNA, 16FA(-) junto con 16AR (+) _40bp, de la prueba de emparejamiento. Líneas azules y rojas representan los espectros de fluorescencia del substrato sin y con Rad51, respectivamente. Verdes y púrpura líneas muestran los espectros de fluorescencia del producto final sin y con Rad51, respectivamente. (B) comparación de espectros de fluorescencia entre el sustrato del dsDNA, 16FA _40bp (-) junto con 16AR (+) _40bp y producto de ssDNA, 16FA _40bp (-), el ensayo de desplazamiento. Líneas azules y rojas representan los espectros de fluorescencia del producto final sin y con Rad51, respectivamente. Verdes y púrpura líneas muestran los espectros de fluorescencia del substrato sin y con Rad51, respectivamente. Esta figura es una adaptación de Ito et al. 19 y modificado. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: reacciones de emparejamiento y desplazamiento de filamento ADN mediaron por Rad51. (A) curso temporal de la fluorescencia normalizada de la reacción de emparejamiento con o sin Rad51. (B) curso temporal de la fluorescencia normalizada de la reacción de desplazamiento con o sin Rad51. (C) Curso del tiempo el cambio en la cantidad de sustrato en la reacción de emparejamiento con Rad51. (D) curso del tiempo el cambio en la cantidad de sustrato en la reacción de desplazamiento con Rad51. (E) A diagrama esquemático del modelo secuencial de tres etapas de reacción. A y B corresponden a la presináptica dsDNA de filamento y de los donantes. C1 corresponde al primer intermedio de tres hebras (inmaduro). C2 se corresponde con el segundo intermedio de tres hebras (maduro). D y E corresponden a del heterodúplex y ssDNA de C2. (F y G) residuos entre los datos experimentales de la hebra de DNA emparejamiento ensayo y una curva teórica obtienen por simulación utilizando el modelo de dos etapas (G) o (F) de tres pasos. (H) puntos rojos indican datos experimentales de la reacción de desplazamiento con Rad51 se muestra en el panel azul de D. línea indica la curva teórica de los productos finales. La curva teórica se generó por simulación utilizando las constantes de tarifa de reacción obtenidas el ensayo de sincronización que se muestra en el panel C. Esta figura es una adaptación de Ito et al. 19 y modificado. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Swi5-Sfr1 estimula la segunda y tercera etapas de la DNA del filamento reacción de intercambio. (A) curso del tiempo el cambio en la cantidad de sustrato en la reacción de emparejamiento con o sin Swi5-Sfr1 (S5S1). (B) residuos entre los datos experimentales de la sincronización de filamento de ADN ensayo con Swi5-Sfr1 y una curva teórica obtenida por simulación utilizando el modelo de tres pasos (línea roja) o dos etapas (línea azul). (C) la combinación de la reacción se muestra en la Figura 4A fue simulada por el modelo de tres pasos con el programa de análisis23 (véase Tabla de materiales). Las constantes de equilibrio de la reacción de cada paso de la reacción, K1 (un), K2 (b) y K3 (c), se obtuvieron de la simulación. Esta figura es una adaptación de Ito et al. 19 y modificado. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Oligonucleótidos para filamento de la DNA Análisis de emparejamiento | ||||
| 16FA(-) | 5'-[FAM] - AAATGAACATAAAGTAAATAAGTATAAGGATAATACA AAATAAGTAAATGAATAAACATAGAAAATAAAGTAAAGGATAT AAA -3 ' | |||
| 16A (-) _40bp | 5'-AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAA-3' | |||
| 16AR (+) _40bp | 5 '- TTTTGTATTATCCTTATACTTATTTACTTTATGTTCATTT-[ROX] -3' | |||
| Oligonucleótidos para análisis de desplazamiento de hebra de ADN | ||||
| 16A(-) | 5'-AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAATA AGTAAATGAATAAACATAGAAAATAAAGTAAAGGATAT AAA -3 ' | |||
| 16FA (-) _40bp | 5'-[FAM] - AAATGAACATAAAGTAAATAAGTATAAGGATAATACAAAA-3' | |||
| 16AR (+) _40bp | 5 '- TTTTGTATTATCCTTATACTTATTTACTTTATGTTCATTT-[ROX] -3' | |||
Tabla 1: lista de los oligonucleótidos utilizados en los ensayos de emparejamiento y el desplazamiento de filamento de ADN. En su caso, las posiciones de fluoróforos (fluoresceína amidite, FAM, carboxi-x-rodamina, ROX) se indican en paréntesis la Plaza.
Discusión
Aquí, hemos descrito un protocolo detallado que utiliza traste para medir cambio de filamento de ADN basada en Rad51 en tiempo real. Lo importante, estas medidas permiten la determinación de la cinética de la reacción. Mientras que las descripciones anteriormente indicadas son suficientes para reproducir los resultados publicados, hay varios puntos críticos que se describirán en esta sección. Además, las ventajas y desventajas de las metodologías basadas en el traste para estudiar cambio de filamento de ADN se analizará, junto con la aplicación de estas técnicas para el estudio de otros aspectos del metabolismo de ADN.
Como reconstrucciones bioquímicos todos, asegurando que todos los sustratos de la reacción son de alta pureza es esencial. Es negligente para asumir la ausencia de actividades contaminantes basados únicamente en la aparente pureza de una preparación de proteínas juzgada mediante tinción de Coomassie. En particular, la presencia de trazas de nucleasas o helicasas puede afectar drásticamente los resultados de los ensayos de emparejamiento y el desplazamiento. Por lo tanto, le recomendamos pruebas de este tipo de actividades cada vez que un nuevo lote de proteína es purificado. Además, es prudente verificar la pureza de substratos DNA sintetizados por electroforesis en geles de poliacrilamida nativa. A pesar de muchas empresas garantizando la pureza de los oligonucleótidos, hemos encontrado a menudo a través de nuestras propias pruebas que la pureza de la DNA sintetizada puede variar entre lotes.
Es importante considerar los siguientes dos puntos al llevar a cabo experimentos con cubetas de cuarzo. En primer lugar, algunas proteínas son propensas a enlazar un cubetas de cuarzo. Para contrarrestar esto, monolaurato de BSA y polyoxyethylenesorbitan están incluidos en los almacenadores intermediarios de reacción. En segundo lugar, la temperatura tiene un efecto drástico sobre la velocidad y fluorescencia la intensidad de la reacción. Para minimizar este efecto, la cubeta de cuarzo debe ser previamente incubada a 37 ° C antes de usar.
Aunque los ensayos bioquímicos convencionales han sido muy útiles en el estudio de cambio de filamento de ADN, tienen varios inconvenientes. En un experimento típico del curso del tiempo, una reacción se incuba a una cierta temperatura y alícuotas son retiradas en los puntos de tiempo deseados y desproteinizadas mediante tratamiento con detergentes y proteasa para terminar la reacción. Al finalizar el curso del tiempo, las muestras entonces se someten a electroforesis para separar sustratos de ADN de los productos. La ventaja principal del método descrito aquí es que permite la observación en tiempo real de la reacción sin ningún disturbio. Cualquier punto durante la reacción puede ser inspeccionado sin interrupción de la reacción sí mismo y no hay que deproteinize muestras o someter a las fuerzas potencialmente disruptivas de la electroforesis. Esto es especialmente relevante cuando supervisión de estructuras de ADN inestable.
A pesar de estas ventajas sobre ensayos convencionales, el método descrito aquí tiene algunas desventajas. Mientras que la utilización de oligonucleótidos ADN sustratos para intercambio del filamento simplifica la interpretación de los resultados, es importante recordar que estos sustratos no se parecen a los sustratos de ADN involucrados en recursos humanos en la célula. Algunos ensayos convencionales utilizan substratos de ADN plásmido tamaño, que son más propensos a reflejar el número de pares de bases que son intercambiados en vivo. Además, la utilización de sustratos de dsDNA circular topológicamente restringida en un subconjunto de pruebas convencionales puede recrear al menos parcialmente las tensiones en el ADN fisiológico.
La aplicación del método descrito aquí ha comenzado a desentrañar los mecanismos moleculares de intercambio de filamento de ADN basada en Rad51. Sin embargo, hay muchas preguntas interesantes que quedan por contestar. Es evidente que la HR durante meiosis requiere Rad51 y Dmc1, el recombinase de RecA-tipo específicos de la meiosis en eucariotas24. Sin embargo, la falta de grandes diferencias bioquímicas entre estos dos recombinasas ha desconcertado a investigadores en el campo durante años. Por otra parte, los papeles de numerosos grupos distintos de factores accesorios de la recombinación ha sido un tema focal de la investigación en el ámbito de los RRHH. Además de dilucidar las diferencias bioquímicas entre Rad51 y Dmc1, nuestro objetivo es investigar y comparar los efectos de factores accesorios diferentes de la recombinación en la cinética de intercambio del filamento de ADN en el futuro inmediato. Finalmente, es importante destacar que la metodología basada en el traste que se describe aquí no se limita al estudio del intercambio del filamento de ADN. Con modificaciones relativamente de menor importancia, tenemos la visión de muchos tipos de aplicaciones para esta técnica en la investigación de funcionalmente diversas proteínas implicadas en ADN metabolismo25,26,27,28. Esperamos que los acontecimientos descritos aquí ofrecerá más opciones a los investigadores pertenecientes a muchas disciplinas diferentes.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Este trabajo fue financiado por subvenciones para la investigación científica (A) (18H 03985) y en áreas innovadoras (15H 05974) HI, para jóvenes científicos (B) (17K 15061) a BA y para científico investigación (B) (18H 02371) HT de la sociedad japonesa para la promoción de la ciencia ( PÁGINAS JSP).
Materiales
| Name | Company | Catalog Number | Comments |
| 0.2 x 1.0 cm quartz cuvette | Hellma Analytics | 105-250-15-40 | |
| 1.0 x 1.0 cm quartz cuvette | Hellma Analytics | 101-10-40 | |
| adenosine triphosphate (ATP) | Sigma | A2383 | |
| DynaFit | BioKin, Ldt. | DynaFit is a program to analyze kinetics of biochemical reactions. | |
| Fluorescent labeled and non-labeled oligonucleotides | Eurofins Genomics | The sequences of oligos are listed in Table. 1. | |
| Magnetic stirrer | Aisis (Japan) | CM1609 | |
| PCR machine | TAKARA (Japan) | TP600 | TAKARA PCR Thermal Cycler Dice |
| Spectrofluorometer | JASCO | FP8300 | Contains a peltier temperature controller and magnetic stirrer system |
| Syringe | HAMILTON | 1702RN 25ul SYR (22s/2"/2) |
Referencias
- Camerini-Otero, R. D., Hsieh, P. Homologous recombination proteins in prokaryotes and eukaryotes. Annual Review of Genetics. 29 (1), 509-552 (1995).
- Cromie, G. A., Connelly, J. C., Leach, D. R. Recombination at double-strand breaks and DNA ends: conserved mechanisms from phage to humans. Molecular Cell. 8 (6), 1163-1174 (2001).
- Pierce, A. J., et al. Double-strand breaks and tumorigenesis. Trends in cell biology. 11 (11), S52-S59 (2001).
- Haber, J. E. Genome Stability. , Garland Science. (2013).
- Kowalczykowski, S. C. An Overview of the Molecular Mechanisms of Recombinational DNA Repair. Cold Spring Harbor perspectives in biology. 7 (11), a016410(2015).
- Bianco, P. R., Tracy, R. B., Kowalczykowski, S. C. DNA strand exchange proteins: a biochemical and physical comparison. Frontiers in bioscience: a journal and virtual library. 3, D570-D603 (1998).
- Renkawitz, J., Lademann, C. A., Jentsch, S. Mechanisms and principles of homology search during recombination. Nature Reviews Molecular Cell Biology. 15 (6), 369-383 (2014).
- Greene, E. C. DNA Sequence Alignment during Homologous Recombination. The Journal of biological chemistry. 291 (22), 11572-11580 (2016).
- Sung, P., Krejci, L., Van Komen, S., Sehorn, M. G. Rad51 recombinase and recombination mediators. Journal of Biological Chemistry. 278 (44), 42729-42732 (2003).
- Prakash, R., Zhang, Y., Feng, W., Jasin, M. Homologous recombination and human health: the roles of BRCA1, BRCA2, and associated proteins. Cold Spring Harbor perspectives in biology. 7 (4), a016600(2015).
- Sung, P. Function of yeast Rad52 protein as a mediator between replication protein A and the Rad51 recombinase. Journal of Biological Chemistry. 272 (45), 28194-28197 (1997).
- Sung, P. Yeast Rad55 and Rad57 proteins form a heterodimer that functions with replication protein A to promote DNA strand exchange by Rad51 recombinase. Genes & Development. 11 (9), 1111-1121 (1997).
- Kurokawa, Y., Murayama, Y., Haruta-Takahashi, N., Urabe, I., Iwasaki, H. Reconstitution of DNA strand exchange mediated by Rhp51 recombinase and two mediators. PLoS biology. 6 (4), e88(2008).
- Jensen, R. B., Carreira, A., Kowalczykowski, S. C. Purified human BRCA2 stimulates RAD51-mediated recombination. Nature. 467 (7316), 678-683 (2010).
- Liu, J., et al. Rad51 paralogues Rad55-Rad57 balance the antirecombinase Srs2 in Rad51 filament formation. Nature. 479 (7372), 245-248 (2011).
- Lu, C. -H., et al. Swi5-Sfr1 stimulates Rad51 recombinase filament assembly by modulating Rad51 dissociation. Proceedings of the National Academy of Sciences of the United States of America. , (2018).
- Bazemore, L. R., Takahashi, M., Radding, C. M. Kinetic analysis of pairing and strand exchange catalyzed by RecA. Detection by fluorescence energy transfer. Journal of Biological Chemistry. 272 (23), 14672-14682 (1997).
- Gupta, R. C., Bazemore, L. R., Golub, E. I., Radding, C. M. Activities of human recombination protein Rad51. Proceedings of the National Academy of Sciences. 94 (2), 463-468 (1997).
- Ito, K., Murayama, Y., Takahashi, M., Iwasaki, H. Two three-strand intermediates are processed during Rad51-driven DNA strand exchange. Nature Structural & Molecular Biology. 25 (1), 29-36 (2018).
- Akamatsu, Y., Dziadkowiec, D., Ikeguchi, M., Shinagawa, H., Iwasaki, H. Two different Swi5-containing protein complexes are involved in mating-type switching and recombination repair in fission yeast. Proceedings of the National Academy of Sciences. 100 (26), 15770-15775 (2003).
- Haruta, N., et al. The Swi5-Sfr1 complex stimulates Rhp51/Rad51- and Dmc1-mediated DNA strand exchange in vitro. Nature Structural & Molecular Biology. 13 (9), 823-830 (2006).
- Argunhan, B., Murayama, Y., Iwasaki, H. The differentiated and conserved roles of Swi5-Sfr1 in homologous recombination. FEBS Letters. 591 (14), 2035-2047 (2017).
- Kuzmic, P. Program DYNAFIT for the analysis of enzyme kinetic data: application to HIV proteinase. Analytical biochemistry. 237 (2), 260-273 (1996).
- Brown, M. S., Bishop, D. K. DNA strand exchange and RecA homologs in meiosis. Cold Spring Harbor perspectives in biology. 7 (1), a016659(2014).
- Rudert, W. A., et al. Double-labeled fluorescent probes for 5' nuclease assays: purification and performance evaluation. BioTechniques. 22 (6), 1140-1145 (1997).
- Xiao, J., Singleton, S. F. Elucidating a key intermediate in homologous DNA strand exchange: structural characterization of the RecA-triple-stranded DNA complex using fluorescence resonance energy transfer. Journal of Molecular Biology. 320 (3), 529-558 (2002).
- Grimme, J. M., et al. Human Rad52 binds and wraps single-stranded DNA and mediates annealing via two hRad52-ssDNA complexes. Nucleic Acids Research. 38 (9), 2917-2930 (2010).
- Algasaier, S. I., et al. DNA and Protein Requirements for Substrate Conformational Changes Necessary for Human Flap Endonuclease-1-catalyzed Reaction. The Journal of biological chemistry. 291 (15), 8258-8268 (2016).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados