
Method Article
Lorsque vous coupez des questions : Une Dissection et Guide de l’analyse de l’Orientation spatiale de la rétine de souris de repères oculaires
Dans cet article
Résumé
Ce protocole fournit un guide complet de la dissection et l’analyse de l’utilisation de repères profond oculaires, s-opsine immunohistochimie, Retistruct et du code personnalisé avec précision et fiabilité orienter la rétine de souris isolées dans l’espace anatomique.
Résumé
Avec précision et fiabilité identifier l’orientation spatiale de la rétine de souris isolé est important pour de nombreuses études en neuroscience visuelle, y compris l’analyse de la densité et les gradients de la taille des types de cellules rétiniennes, l’accordage de sens de direction-sélectif cellules ganglionnaires et l’examen des profils topographiques dégénérescence dans certaines maladies de la rétine. Cependant, il existe de nombreuses méthodes de dissection oculaires différents rapportés dans la littérature qui sont utilisés pour identifier et étiqueter rétinienne orientation dans la rétine de souris. Alors que la méthode d’orientation utilisée dans ces études est souvent négligée, orientation comment rétinienne pas considérée est déterminée peut causer écarts dans la littérature et de la confusion lorsque vous essayez de comparer les données entre les études. Des monuments oculaires superficielles comme cornéens brûlures sont couramment utilisées, mais ont récemment été démontrés être moins fiables que les monuments plus profondes comme les muscles rectus, la fissure choroïde ou le gradient de s-opsine. Ici, nous fournissons un guide complet pour l’utilisation des points de repère profond oculaires de disséquer avec précision et de documenter l’orientation spatiale d’une rétine de souris isolé. Nous avons aussi comparé l’efficacité des deux anticorps s-opsine et inclus un protocole pour immunohistochemistry s-opsine. Parce que l’orientation de la rétine selon le gradient de s-opsine nécessite une reconstruction rétinienne avec le logiciel Retistruct et rotation avec un code personnalisé, nous avons présenté les mesures importantes nécessaires pour utiliser ces deux programmes. Dans l’ensemble, le but du présent protocole est de livrer un ensemble fiable et reproductible des méthodes pour l’orientation rétinienne précise qui s’adapte aux protocoles plus expérimentales. Des grands objectifs de ce travail consiste à normaliser les méthodes d’orientation rétinienne pour de futures études.
Introduction
Un aspect important et parfois négligé des neurosciences rétinienne est l’orientation correcte et l’analyse de la rétine tout montage isolée, que ce soit l’orientation de la rétine dans une chambre d’enregistrement électrophysiologie ou sur une lame histologique. Cela est particulièrement important pour les études concernant la rétine de souris, qui est actuellement le modèle plus répandu pour les enquêtes du système visuel chez les mammifères. Des découvertes récentes révèlent que la rétine de souris n’est pas uniforme dans l’espace, mais a des gradients de densité et la taille des types de cellules rétiniennes fonctionnellement distinctes, telles que cellules ganglionnaires melanopsin et les cellules ganglionnaires OFF-alpha transitoires opsines des cônes1,2 ,3,4,5. En conséquence, la méthode utilisée pour déterminer l’orientation de la rétine peut influencer les résultats expérimentaux impliquant les cellules type ou opsine distributions2,3,6, direction réglage de direction sélective ganglion cellules7,8,9et les modèles topographiques de dégénérescence rétinienne10,11,12,13,14 . En fait, le pas considérée orientation comment rétinienne est rapportée peut causer divergences dans la littérature et de la confusion lorsque vous essayez de comparer les données entre les études. Il est donc essentiel que les chercheurs déclarent la méthode pour déterminer l’orientation de la rétine afin que les résultats de ces études peuvent être correctement interprétés.
Orientation rétinienne est couramment identifiée en marquant la cornée dorsale, ventrale, nasale ou temporelle avant énucléation oculaire1,3,12,15,16,17 ,18,19 ou par coupure ou coloration repères oeil profond anatomiques tels que les muscles extraoculaires6,7, la choroïde fissure20,21, ou le s-opsine dégradé2,3. Les muscles rectus peuvent servir à identifier le nasal dorsal, ventral, et la rétine temporale en faisant une coupe profonde relaxation qui divise l’attachement de l’autre le rectus supérieur, rectus inférieur, rectus médial ou muscle latéral droit, respectivement. Cependant, pour la plupart des expériences, à l’aide d’un muscle droit est suffisant pour orienter la rétine22. La fissure choroïde, qui est un vestige du développement de le œil, peut être considérée comme une légère ligne horizontale à l’arrière de le œil. Chaque extrémité de cette ligne se termine à la nasale ou le pôle temporal du globe23. Enfin, expression s-opsine est distribuée asymétrique à la rétine ventrale chez les souris, et s-opsine anticorps peuvent être utilisés pour révéler la rétine ventrale dans immunohistochemical expériences1.
Travaux récents de Stabio, et al. 22 a démontré que des monuments oculaires superficielles comme cornéens brûlures sont une méthode moins fiable pour orienter la rétine dans l’espace anatomique, probablement en raison de l’erreur humaine et la variabilité dans la fabrication de la brûlure cornéenne lorsque vous utilisez le temporel et médial canthi comme points de référence. En revanche, monuments profondes, comme le muscle droit supérieur, fissure choroïde et le gradient de s-opsine, ont démontré d’être des repères plus fiables et précises pour orienter la rétine22. Cependant, l’identification de ces repères anatomiques nécessite des étapes de la dissection unique qui ne sont pas décrites en détail dans la littérature. Ainsi, l’objectif du présent protocole est de fournir un tutoriel complet sur la façon d’utiliser le muscle droit supérieur, fissure choroïde et gradient de s-opsine d’identifier avec précision l’orientation spatiale de la rétine de souris. En outre, nous avons inclus une comparaison de l’efficacité des deux anticorps s-opsine, mais aussi un protocole pour immunohistochemistry s-opsine.
Un défi supplémentaire aux études s’appuyant sur l’orientation précise rétinienne est la grosse pièce relève pour aplatir les rétines wholemount sur un plat, chambre d’enregistrement ou une diapositive. Cela peut introduire des défis pour l’analyse de ce qui est naturellement une structure tridimensionnelle lorsqu’il est imagé comme une structure plane à deux dimensions. Un programme appelé Retistruct24 peut servir pour renvoyer un plat wholemount de la rétine à sa structure tridimensionnelle avant que les données recueillies auprès d’elle sont analysées. Ainsi, une section du présent protocole est dédiée à mettre en évidence les étapes qui sont nécessaires pour utiliser le logiciel Retistruct pour reconstruire la rétine de souris s-opsine immunomarquage. Nous avons également inclus une section du protocole pour l’utilisation de notre script personnalisé de MATLAB, qui a été développé à la rétine de souris de précisément faire pivoter et orienter colorées avec s-opsine.
Protocole
Toutes les méthodes décrites ici ont été approuvés par l’animalier institutionnel et utilisation Comité (IACUC) de l’Université d’Akron.
1. en utilisant le point de repère Muscle Rectus supérieur pour identifier l’Orientation rétinienne
Remarque : Le muscle droit supérieur est un point de repère pour la dorsale de la rétine (tableau 1). Si l’expérience ne nécessite pas le marquage de la rétine dorsale, ignorez l’étape 1 et passez à l’étape 2.
- Suivre votre protocole institutionnel animalier et Comité d’urbanisme approuvé d’euthanasie de la souris.
- Pour identifier l’orientation générale du globe, assurez-vous une brûlure marquer sur la cornée dorsale directement entre les canthi nasale et temporelles près de la frontière de la cornée-sclérotique immédiatement après l’euthanasie (Figure 1A). Faire la brûlure marque par chauffage un stylo de cautérisation pendant dix secondes et ensuite toucher le bout du stylo à la cornée dorsale pour moins d’une seconde.
NOTE : Tenue de que la plume de cautérisation de la cornée pour trop longtemps provoquera le globe à la perforation.
NOTE : Tandis que des stylos de cautérisation émettre de la lumière, la plume de cautérisation énumérée dans la Table des matières n’émet pas de lumière lorsqu’il est chauffé, ce qui en fait une option sûre pour expériences adapté à l’obscurité. - Pour l’énucléation, utiliser forceps courbe doucement pousser le œil de son support et poignée du globe des dessous. Ne pas couper le nerf optique pour enlever le globe ; au lieu de cela, soulever doucement le globe de sa douille tout en simultanément déplacez-la doucement de gauche à droite jusqu'à ce que le monde est sorti de la prise.
NOTE : La présente proposition permettra les muscles rectus reste attachée à la planète quand le globe est finalement supprimé complètement de la prise. Le nerf optique aussi souvent reste attaché au monde entier. - Transférer le globe avec les muscles rectus attaché dans une boîte de Pétri contenant le milieu de la dissection. Assurez-vous de garder une trace de que l’oeil est le œil gauche et le œil droit.
Remarque : Le dissecteur doit utiliser un support approprié de dissection qui s’aligne avec leur protocole expérimental. - Dans le cadre de la dissection, localiser visuellement la brûlure cornéenne dorsale et identifier le muscle droit supérieur avec auxquels il est associé (Figure 1A).
- À l’aide de ciseaux de dissection ou un 20 G (0,9 mm x 25 mm) aiguille de ponction (voir Table des matières), la cornée à la marque de brûlure. Faire un profond soulagement coupe dans le monde vers le nerf optique à coupent le muscle supérieur. Une rétine isolée et reconstituée avec cette coupe est montrée dans les Figures 1 b et 1C.
- Commencez à isoler la rétine à l’aide de deux paires de pinces (voir Table des matières) à déchirer délicatement le trou fait avec le pneu crevé à l’étape 1.6 jusqu'à ce que la partie de la rétine est exposée.
Remarque : Il est important de le faire doucement, comme déchirement avec trop de force peut causer la coupe relève à déchirer davantage. - Forceps permet de distinguer la rétine de la sclère, jusqu'à ce que la sclérotique a été complètement enlevée. Supprimer l’iris, cristallin, vitreux et toutes les structures restantes avec une pince jusqu'à ce que la rétine est complètement isolée.
Remarque : Le protocole peut être suspendu ici. Si le tissu va être fixe pour s-opsine immunohistochemistry, passez à l’étape 3.5.
2. en utilisant le point de repère Fissure choroïde pour identifier l’Orientation rétinienne
Remarque : La fissure choroïde est présente sur la sclérotique à l’arrière de le œil et s’étend du pôle temporal au pôle nasal (Figures 2 b et 2C; Tableau 1).
- Suivre protocole institutionnel animalier et Comité d’urbanisme approuvé d’euthanasie de la souris.
- Pour identifier l’orientation générale du globe, assurez-vous une brûlure marquer sur la cornée dorsale directement entre les canthi nasale et temporelles près de la frontière de la cornée-sclérotique immédiatement après l’euthanasie (Figure 2A). Faire la brûlure marque par chauffage un stylo de cautérisation pendant dix secondes et ensuite toucher le bout du stylo à la cornée dorsale pour moins d’une seconde.
NOTE : Tenue de que la plume de cautérisation de la cornée pour trop longtemps provoquera le globe à la perforation. - Enucleate le œil et le transfert du globe dans une boîte de Pétri contenant le milieu de la dissection. Assurez-vous de garder une trace de que l’oeil est le œil gauche et le œil droit.
Remarque : Le dissecteur doit utiliser un support approprié de dissection qui s’aligne avec leur protocole expérimental. - Visuellement, localiser et identifier la fissure choroïde à l’arrière de le œil (Figure 2B, 2C).
Remarque : La fissure choroïde est également visible à l’intérieur de le œilleton sous lumière infra-rouge20. - Orienter le monde dans la boîte de Pétri afin que la brûlure dorsale est située au pôle supérieur, tel qu’il serait si le œil était toujours chez la souris.
Remarque : La présence de la partie dorsale burn permet d’identifier la côté nasale et temporelle de la planète, tant que ce soit un oeil droit ou gauche a été documentée : si c’est un œil droit, la fissure choroïde nasale sera à droite de la brûlure et la tempora fissure choroïde l sera à gauche de la brûlure. Si c’est un œil gauche, la fissure choroïde temporelle sera à droite de la brûlure et la fissure choroïde nasale sera à gauche de la brûlure.
À l’aide de ciseaux de dissection ou un 20 G (0,9 x 25 mm) avec une aiguille (voir Table des matières), faire une ponction dans le monde où se trouve la brûlure dorsale. - Effectuer un soulagement peu profond vers le nerf optique où se trouve la brûlure cornéenne dorsale. Cette réduction sera perpendiculaire à la fissure choroïde, permettant une identification de la rétine dorsale après isolement (Figure 2D).
- Faire les suivantes deux incisions profondes relève vers le nerf optique : un en alignant les lames de la dissection ciseaux vers le haut à la ligne de fissure choroïde temporelle à l’arrière de le œil, et une en alignant les lames de la dissection ciseaux vers le haut à la choroïde nasale ligne de fissure à l’arrière de le œil. Ces réductions sont indiquées sur une rétine isolée et reconstituée en Figure 2D et 2E.
NOTE : Vous pouvez également un profond sillon peut être fait à la fissure choroïde temporelle et une coupe peu profonde peut être faite à la fissure choroïde nasale, rendant la brûlure cornéenne dorsale couper inutiles. Cela permet l’orientation exacte de la rétine avec des réductions moins soulager. - Commencez à isoler la rétine à l’aide de deux paires de pinces (voir Table des matières) à déchirer délicatement le trou fait avec le pneu crevé en étapes 2.7 et 2.8, jusqu'à ce que la partie de la rétine est exposée.
Remarque : Il est important de le faire doucement, comme déchirement avec trop de force peut causer le cliché (s) relève à déchirer davantage. - Forceps permet de distinguer la rétine de la sclère, jusqu'à ce que la sclérotique a été complètement enlevée. Supprimer l’iris, cristallin, vitreux et toutes les structures restantes avec une pince jusqu'à ce que la rétine est complètement isolée.
Remarque : Le protocole peut être suspendu ici. Si le tissu va être fixe pour s-opsine immunohistochemistry, passez à l’étape 3.5.
3. étiquetage du Gradient de S-opsine dans la rétine de souris
NOTE : L’expression de photopigment s-opsine est asymétrique distribuée à la rétine ventrale1, ce qui en fait un excellent marqueur de la partie ventrale de la rétine. Cette méthode n’est utile que pour les fixes et les tissus immunostained (tableau 1). Les étapes suivantes peuvent être appliquées à la rétine qui ont été disséqués en utilisant une des méthodes ci-dessus.
- Suivre protocole institutionnel animalier et Comité d’urbanisme approuvé d’euthanasie de la souris.
- Immédiatement après l’euthanasie, enucleate le œil et placer le globe dans une boîte de Pétri avec milieu de dissection. Assurez-vous de garder une trace de que l’oeil est le œil gauche et le œil droit afin d’identifier l’orientation rétinienne après que la rétine est disséquée.
Remarque : Le dissecteur doit utiliser un support approprié de dissection qui s’aligne avec leur protocole expérimental. -
Commencez à isoler la rétine à l’aide de deux paires de pinces (Table des matières) à déchirer délicatement un trou dans la cornée jusqu'à ce que la partie de la rétine est exposée.
Remarque : Il est important de le faire doucement, comme déchirement avec trop de force peut causer la rétine se déchirer. - Forceps permet de distinguer la rétine de la sclère, jusqu'à ce que la sclérotique a été complètement enlevée. Supprimer l’iris, cristallin, vitreux et toutes les structures restantes avec une pince jusqu'à ce que la rétine est complètement isolée.
Remarque : Le protocole peut être suspendu ici. Si à l’aide de la rétine pour un ex vivo expérimenter, faire l’expérience avant d’effectuer les étapes suivantes. - À l’aide de ciseaux de dissection, faire quatre coupes relève dans la rétine afin qu’il se sont posées à plat. Montez la rétine ganglion cell-côté vers le haut sur une membrane de nitrocellulose (Table des matières) en appuyant doucement sur chaque coin de la rétine sur la membrane avec une pince.
Remarque : L’emplacement des découpes relève peut être arbitraire, lorsqu’on utilise le gradient de s-opsine pour orientation rétinienne. - Transfert la rétine montée dans le premier puits dans une plaque 24 puits (Table des matières) à l’aide de pinces, contenant 1 mL de paraformaldéhyde à 4 % (Table des matières) pour la fixation. Placer la plaque 24 puits dans un agitateur orbital à température ambiante (Table des matières) et fixer la rétine pendant 40 min exactement.
Remarque : Toutes les étapes de lavage et d’incubation suivants doivent être établie avec la plaque 24 puits dans un agitateur orbital. - Laver la rétine pendant 15 min à température ambiante en le transférant à la seconde bien remplie avec 1 mL de PBS de 0,1 M. Répétez cette opération deux fois en séquentiellement transférant la rétine dans les puits 0,1 de troisième et quatrième remplis de PBS de M.
- Transférer la rétine montée au cinquième bien contenant 1 mL de solution (1,7 % Triton X-100 et 5,2 % âne sérum normal dans du PBS de 0,1 M ; Voir le Tableau des matériaux) de blocage et incuber une nuit à 4 ° C.
- Ajoutez l’anticorps primaire de lapin anti-s-opsine (voir Table des matières) à la solution de blocage à une concentration de 1/500 et incuber pendant trois jours à 4 ° C.
- Laver l’excès d’anticorps primaire de la rétine six fois en le plaçant dans l’ordre dans six puits contenant 1 mL de PBS de 0,1 M pendant 10 minutes chacun, à la température ambiante.
- Placez la rétine dans un puits avec la solution de blocage fraîchement (1,7 % Triton X-100 et 5,2 % âne sérum normal dans du PBS de 0,1 M) et ajoutez âne anti-lapin Alexa-594 l’anticorps secondaire (voir Table des matières). Incuber la rétine avec l’anticorps secondaire durant la nuit à 4 ° C.
- Laver l’anticorps secondaire excédentaire de la rétine six fois en le plaçant dans l’ordre dans six puits contenant 1 mL de PBS frais de 0,1 M de 10 min chacun, à la température ambiante.
- À l’aide de pinces, transférer la rétine montée à une boîte de Pétri contenant 0,1 M PBS. Communiqué de la rétine de la membrane de nitrocellulose en insérant doucement l’extrémité des pinces entre la rétine et la membrane jusqu'à ce que la rétine est n’est plus attachée.
- Montez la rétine sur une lame de microscope de verre doucement il aiguillon avec une pince jusqu'à ce que les bâtonnets de la rétine au verre et retirer la lame de la boîte de Pétri.
- Couvrir la rétine sur la diapositive avec Aquamount et recouvrir d’un lamelle couvre-objet #1.5. Mettre la lame dans un bac de diapositive (voir Table des matières) et laisser reposer à température ambiante pendant une heure.
- Retourner la lame pour le réfrigérateur et le magasin dans un bac de diapositive (voir Table des matières) à 4 ° C quand pas en service. Après que la diapositive a été couvertes pendant 24 heures, utiliser le vernis à ongles pour sceller les côtés de la lame pour éviter la dessiccation.
Remarque : Le protocole peut être suspendu ici.
4. utilisation reconstruit rétines immunomarquage avec S-opsine pour identifier l’Orientation rétinienne
- Visualiser le gradient de s-opsine avec un microscope confocal ou un microscope à épifluorescence avec un accessoire de caméra (voir Table des matières) et l’image de la rétine pour que la rétine entière est visible en une seule image (Figures 1 b, 2D, 3 a et 3D). cela peut être fait par l’imagerie de la rétine dans les sections à faible grossissement et puis assembler les images.
- Nommez les rétines afin qu’ils soient identifiables. Par exemple, nommez la rétine premier à être reconstruit « Retina1 ».
- Téléchargez et installez ImageJ à https://imagej.nih.gov/ij/download.html.
- Créez un dossier individuel pour chaque rétine qui doit être reconstruit, mais laissez les dossiers vides. Par exemple, créez un dossier intitulé « Retina1 ». Tous les fichiers nécessaires pour reconstruire cette rétine seront placés dans ce dossier dans les étapes ultérieures.
Remarque : Les seuls fichiers de que ces dossiers doivent contenir sont les fichiers qui doivent être analysés par Retistruct. Tous les fichiers autres que celles détaillées ci-dessous fera la rétine impossible d’être ouvert avec le logiciel Retistruct. - Ouvrez l’image de la rétine dans ImageJ en choisissant Fichier → Ouvrir, puis en choisissant « Retina1 ».
- Sans apporter de modifications à l’image, enregistrer en tant que « image.png » dans le dossier intitulé « rétine 1" en choisissant Fichier → Enregistrer sous → PNG.
Remarque : Le fichier doit être nommé « image.png » afin que le logiciel de Retistruct reconnaître le fichier comme une rétine pour la reconstruction. - Utilisez l’outil de ligne de segmenté d’esquisser les contours de la rétine. En cliquant sur deux points adjacents à la frontière de la rétine, l’outil courbe segmentée sera essentiellement « relier les points » entre les deux places adjacentes, création d’un plan. Répétez jusqu'à ce que la rétine entière a été soulignée. Enregistrer le contour de la rétine comme « outline.roi » dans le dossier intitulé « Retina1 » en sélectionnant Analyze → outils → ROI gestionnaire → ajouter [t] → plus → enregistrer.
Remarque : Le bord de la rétine peut être identifié où la coloration de la s-opsine passe à l’arrière-plan. - Utilisez l’outil de ligne segmentée en suivant les instructions à l’étape 4.7 d’esquisser la frontière du disque optique. Enregistrer le contour du disque optique comme « od.roi » dans le dossier intitulé « Retina1 » en sélectionnant Analyze → outils → ROI gestionnaire → ajouter [t] → plus → enregistrer.
Remarque : Le disque optique est identifié comme le petit trou au milieu de la rétine et variera selon la qualité de la dissection.
Remarque : Tous les fichiers nécessaires pour la reconstruction de Retistruct (« image.png », « outline.roi » et « od.roi ») devraient maintenant être enregistré le dossier « Retina1 ». - Pour télécharger, installer et ouvrez le programme Retistruct, suivez les instructions décrites dans le guide de l’utilisateur Retistruct dans la section documents complémentaires de Sterratt, et al. 24
- Une fois le Retistruct fenêtre apparait, cliquez sur l’icône « Ouvrir » en haut à gauche de la fenêtre et sélectionnez le répertoire « Retina1 ».
- Une fenêtre d’image apparaîtra indiquant qu’aucune barre d’échelle n’existe. Cliquez sur « Fermer » et une image de la rétine apparaît dans la boîte. Visualiser le contour de la rétine en cliquant sur le bouton « Propriétés » en haut à droite de la fenêtre et changement de couleur du contour une couleur visible (Figure 5A).
- IMPORTANT : Préciser si la rétine est d’un œil droit ou le œil gauche dans le panneau sur la gauche (Figure 5A).
- Cliquez sur le bouton « Ajouter Tear » sur la gauche et spécifiez l’emplacement une larme ou la coupe de la rétine en cliquant sur les trois sommets de la déchirure (Figure 5A). Cela créera des lignes qui relient les trois sommets de la coupe. Répéter pour toutes les coupes dans la rétine.
- Spécifiez la rétine dorsale en cliquant sur un point arbitraire de l’esquisse de la rétine. Une lettre majuscule « D » s’affiche à ce moment-là sur le contour (Figure 5B).
Remarque : La rétine dorsale sera la moitié sombre de la rétine, à l’opposé du gradient de s-opsine. Cependant, marquant la rétine dorsale dans Retistruct n’est pas une méthode fiable pour identifier la moitié dorsale de la rétine, le marquage des « dorsale » peut donc être arbitraire dans cette étape. - Reconstruire la rétine en cliquant sur le bouton de « Reconstruire la rétine » en haut à gauche de l’écran (Figure 5B). Un tracé polaire de la rétine reconstituée apparaîtra avec les coupures visibles dans la même couleur que le contour (Figure 5C).
- Cliquez sur le bouton « Enregistrer » sur la droite de l’écran pour que la rétine reconstituée et toutes les données qui lui sont associées est enregistré dans le répertoire du dossier « Retina1 » (Figure 5D).
- Enregistrez la rétine reconstituée en cliquant sur le bouton « PDF » dans le panneau de droite (Figure 5D). Une boîte apparaît pour vous demande de spécification de la taille. La taille par défaut est acceptable pour les étapes suivantes. Cette action permettra d’économiser la rétine reconstruite comme « image.polar.pdf » dans le répertoire du dossier « Retina1 ».
- Ouvrez « image.polar.pdf » dans un programme de peinture (ou autre programme de manipulation d’image) et utiliser l’outil « Pot de peinture » (ou similaire) pour changer le fond de la rétine reconstruit au noir. Enregistrez la rétine reconstruite sous un fichier .tif, tels que « Retina1_reconstructed.tif » dans le répertoire du dossier « Retina1 ».
Remarque : Le protocole peut être suspendu ici. - Télécharger le code MATLAB pour la rotation de la rétine appelée « Retina_Rotator.m » (voir documents complémentaires). Placez le fichier de code dans son propre dossier avec aucun autre fichier dans le dossier.
- Ouvrir MATLAB, version 2007 b ou version ultérieure. Double-cliquez sur le fichier de code pour l’ouvrir dans MATLAB. Dans la fenêtre commande, tapez « Retina_Rotator » et puis appuyez sur la touche entrée. Une fenêtre de recherche s’affiche.
Remarque : Le code est spécifique pour les fichiers .tif. Si le fichier doit pivoter n’est pas au format correct, le code ne tourne pas correctement la rétine. Reportez-vous aux étapes 4.17 et 4.18. pour sauver la rétine reconstituée dans le format approprié. - Ouvrez le fichier à être tourné. Par exemple, choisissez « Retina1_reconstructed.tif ». Le code va alors analyser la rétine reconstituée et enregistrera automatiquement la rétine pivotée comme « Retina1_reconstructed_rotated.tif » dans le dossier où se trouve le fichier d’origine.
- A la fin le code d’analyse de la rétine, une fenêtre apparaîtra également montrer les images de la rétine avant et après rotation pour comparaison (Figures 3 b et 3C ; 3E les chiffres et 3F).
Remarque : Ce code tourne la rétine reconstruite pour que la moitié ventrale (plus brillante) soit sur le fond et la moitié dorsale (luminositée) est sur le dessus, donc précisément orienter la rétine selon le gradient de s-opsine1. Si la rétine est d’un bon œil, ou œil gauche a été documentée, l’emplacement des poteaux nasales et temporelles aussi peut être extrapolée à partir de cette méthode d’orientation (Figure 3).
Résultats
Une coupe relève qui traverse le muscle droit supérieur avec précision et fiabilité identifie la rétine dorsale (Figure 1). La fissure choroïde avec précision et fiabilité identifie la rétine nasale et temporelle avec des coupes de relaxation profondes le long de la fissure choroïde temporelle et nasale (Figure 2). Dans cet exemple, une coupe relève également a été déposée dans la rétine dorsale afin d’identifier l’axe de la dorsale/ventrales de la rétine (Figure 2D, flèche verticale). Les étapes de ces processus sont indiqués à des fins de réplication par dissecteurs futures. Une combinaison d’immunohistochimie s-opsine (Figure 3 a et 3D), reconstruction avec le logiciel Retistruct (3 b, 3E) et une rotation précise avec un code personnalisé de MATLAB (3C, 3F) permettant la identification des moitiés ventrales et dorsales de la rétine, mais aussi les pôles nasales et temporelles si elle ne sait pas si la rétine est d’un œil droit ou gauche (Figure 3). Nous avons également comparé les deux anticorps primaires s-opsine couramment utilisés pour l’efficacité dans l’étiquetage des cônes s-opsine (Figure 4A-D) : les deux la chèvre anti-s-opsine anticorps primaire et l’anticorps primaire de lapin anti-s-opsine efficacement l’étiquette cônes s-opsine (Figure 4E) dans la même souris.
Soulager coupures ont été identifiés sur les rétines immunomarquage reconstruit s-opsine et leurs emplacements ont été comparés à l’orientation déterminée par le gradient de s-opsine. En utilisant notre personnalisé MATLAB code rétines (voir Matériaux supplémentaires), ont été tournés avec précision afin que la concentration la plus élevée de s-opsine coloration est située sur le ventre, plaçant ainsi le vrai dorsale à 90 ° (pour rectus supérieur), véritable nasale à 0 ° (pour les voies nasales fissure choroïde) et vrai temporelle à 180 ° (pour fissure choroïde temporelle). La valeur de chaque individu relève coupée l’angle a été déterminée à l’aide de l’outil angle dans ImageJ après que rétines ont redressé selon le gradient de s-opsine. Un angle moyen a été calculé pour chaque relève coupé type et la valeur moyenne de chaque type de coupe relève a été ensuite tracée sur un terrain polaire (Figure 6). En moyenne, coupes de muscle rectus supérieur identifiés le pôle dorsal à 96,3 ± 4,3 ° (n = 11) (Figure 6). La fissure choroïde nasale identifié le pôle nasal à 6,7 ± 5,8 ° et la fissure choroïde temporelle du pôle temporal à 172,0 ± 4,4 ° (n = 9 ; La figure 6).

Figure 1 : à l’aide du muscle droit supérieur afin d’identifier précisément la rétine dorsale d’un bon oeil. (A) un exemple d’une brûlure cornéenne dorsale près de la frontière cornée sclère faite avec un feutre cautérisation (flèche blanche). Le muscle droit supérieur est également visible dans ce point de vue (flèche blanche). (B) un exemple d’un ensemble monté rétine avec un soulagement coupe faite dans la rétine dorsale en coupant le muscle droit supérieur. Flèche représente le profond soulagement coupe opérée dans la rétine dorsale en coupant le muscle droit supérieur. La rétine est colorée avec l’anticorps primaire chèvre anti-s-opsine (voir Table des matières) et anticorps secondaire âne anti-chèvre Alexa 594 (voir Table des matières; excitation : 590 nm ; émission : 620 nm) (cyan). Rétine a été photographié avec un microscope à épifluorescence avec un filtre de Texas Red (595 nm). (C) une rétine reconstruit en Retistruct et tourné avec un code MATLAB personnalisé (voir la Documentation supplémentaire) avec le rectus supérieur muscle soulager coupé visible (flèche blanche). D: dorsale, V: ventrale, T: temporelle, N: nasale. Barreaux de l’échelle = 1 mm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
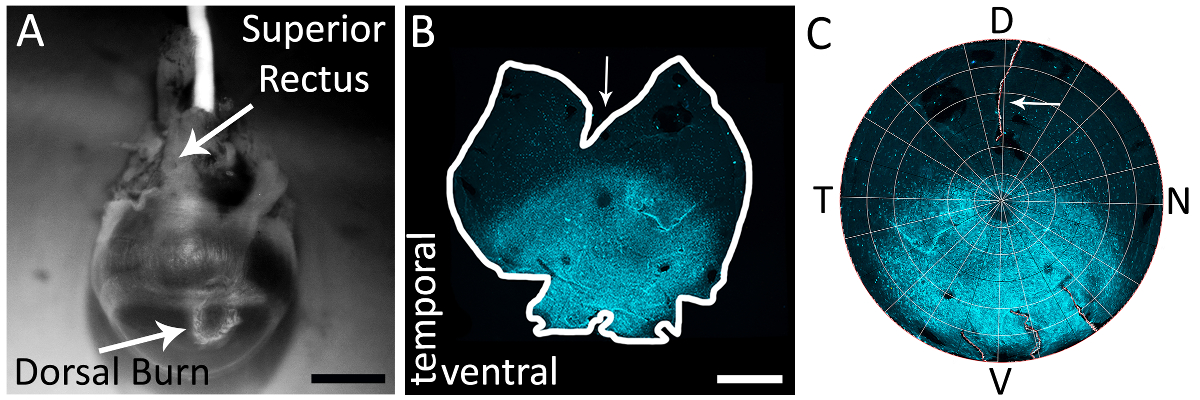{kind=link}

Figure 2 : à l’aide de la fissure choroïde afin d’identifier précisément les pôles nasales et temporale de la rétine d’un œil droit. (A) un exemple d’une brûlure cornéenne dorsale près de la frontière cornée sclère faite avec un feutre cautérisation. (B) la choroïde fissure visible à l’arrière de le œil sur la sclère (flèche blanche). La brûlure cornéenne dorsale est également visible dans ce point de vue, situé à environ 90° de la fissure choroïde temporelle. (C) la choroïde fissure visible à l’arrière de le œil sur la sclère, voyageant du nerf optique jusqu'à la frontière cornée sclère. (D) une rétine tachée de chèvre anti-s-opsine (voir Table des matières) et anticorps secondaire âne anti-chèvre Alexa 594 (voir Table des matières; excitation : 590 nm ; émission : 620 nm) (cyan) avec des coupes de fissure choroïde (horizontal les flèches) et la relaxation dorsale coupe (flèche verticale). Rétine a été photographié avec un microscope à épifluorescence avec un filtre de Texas Red (595 nm). E une rétine reconstruit en Retistruct et tourné avec un code personnalisé de MATLAB (voir matériaux supplémentaires) avec la coupe de relaxation dorsale et les coupes nasale et temporelle fissure choroïde visibles (flèches blanches). D: dorsale, V: ventrale, T: temporelle, N: nasale. Barreaux de l’échelle = 1 mm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
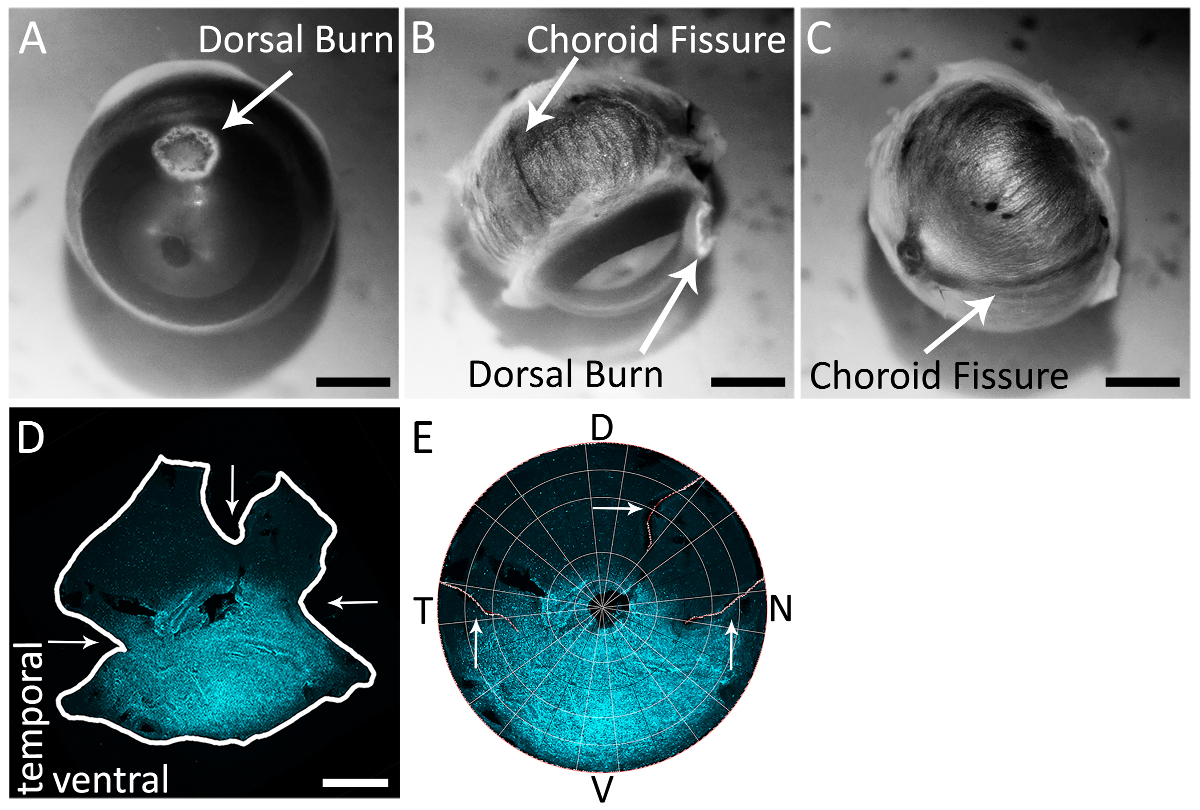{kind=link}

Figure 3 : en utilisant le gradient de s-opsine d’identifier tous les quatre pôles de la rétine. (A) un exemple d’une rétine dissection d’un œil droit qui a été immunomarquées à étiqueter s-opsine et photographié avec un microscope à épifluorescence avec un filtre de Texas Red (595 nm). Les réductions de cette rétine sont arbitraires puisque l’orientation topographique est déterminée par le gradient de s-opsine. (B) les résultats de la reconstruction de la rétine chez les A avec Retistruct. Notez que le gradient de s-opsine n’est pas aligné correctement parce que la rétine n’a pas été exécutée par le code MATLAB personnalisé (voir Matériaux supplémentaires). (C) les résultats de la rotation de la rétine dans A avec le code personnalisé. La rétine a été tournée afin que la concentration la plus élevée de s-opsine coloration se trouve en bas et identifiée comme la rétine ventrale. Parce que la rétine est un œil droit, le pôle temporal est situé à 90° dans le sens inverse du pôle dorsal et le pôle nasal est situé à 90° vers la droite du pôle dorsal. (D) un exemple d’une rétine dissection d’un œil gauche qui a été immunomarquées à étiqueter s-opsine et photographié avec un filtre de Texas Red (595 nm). Les réductions de cette rétine sont arbitraires puisque l’orientation topographique est déterminée par le gradient de s-opsine. (E) les résultats de reconstituer numériquement la rétine en ré avec Retistruct. Notez que le gradient de s-opsine n’est pas aligné correctement parce que la rétine n’a pas subi une rotation par le code personnalisé. (F) les résultats de la rotation de la rétine en ré avec le code personnalisé. La rétine a été tournée afin que la concentration la plus élevée de s-opsine coloration se trouve en bas et identifiée comme la rétine ventrale. Parce que la rétine est un œil gauche, le pôle nasal est situé à 90° dans le sens inverse du pôle dorsal et le pôle temporal est situé à 90° vers la droite du pôle dorsal. D: dorsale, V: ventrale, T: temporelle, N: nasale. Barreaux de l’échelle = 1 mm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
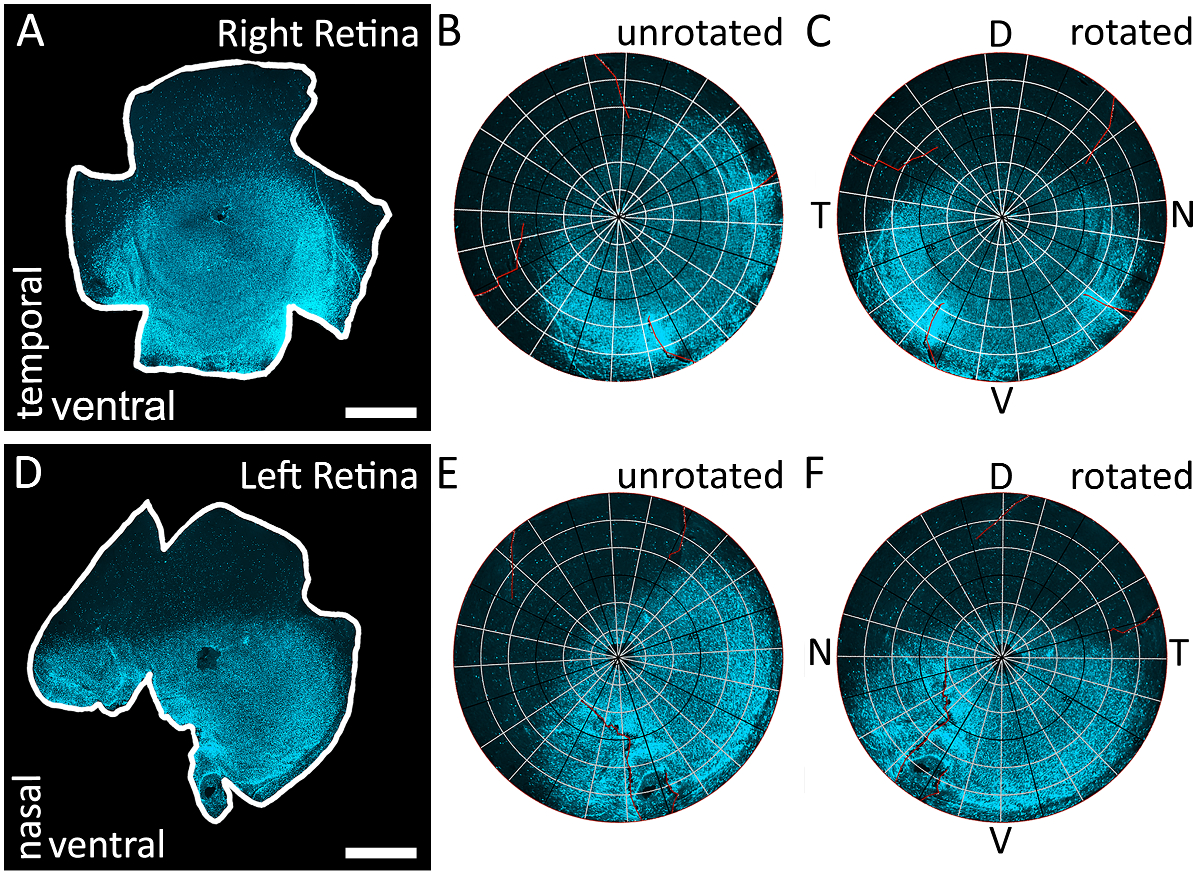{kind=link}

Figure 4 : Comparaison de deux anticorps primaires s-opsine dans l’étiquetage des cônes s-opsine. (A) A rétine colorées avec l’anticorps primaire de chèvre anti-s-opsine (voir Table des matières). (B) autre la rétine de souris même colorées avec l’anticorps primaire de lapin anti-s-opsine (voir Table des matières). (C), une région représentative (0,1 x 0,1 mm2) de la rétine colorées avec l’anticorps primaire de chèvre anti-s-opsine. Image prise sur un microscope à épifluorescence à un grossissement de 40 X. (D), une région représentative (0,1 x 0,1 mm2) de la rétine tachée de lapin anti-s-opsine (voir Table des matières), une alternative de l’anticorps primaire. Image a été prise sur un microscope à épifluorescence à un grossissement de 40 X. (E) les deux anticorps étiqueter le même nombre de segments externes s-cône car il n’y a pas de différence significative du nombre d’immunopositifs s-cônes qui sont souillés de chèvre anti-s-opsine et de lapin anti-s-opsine à tout de la rétine testé excentricités (n = 2 ; Analyse de la variance avec test de Bonferroni post hoc ; p > 0,05). Barreaux de l’échelle = 1 mm (A-B) ; 25 µm (C-D). S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
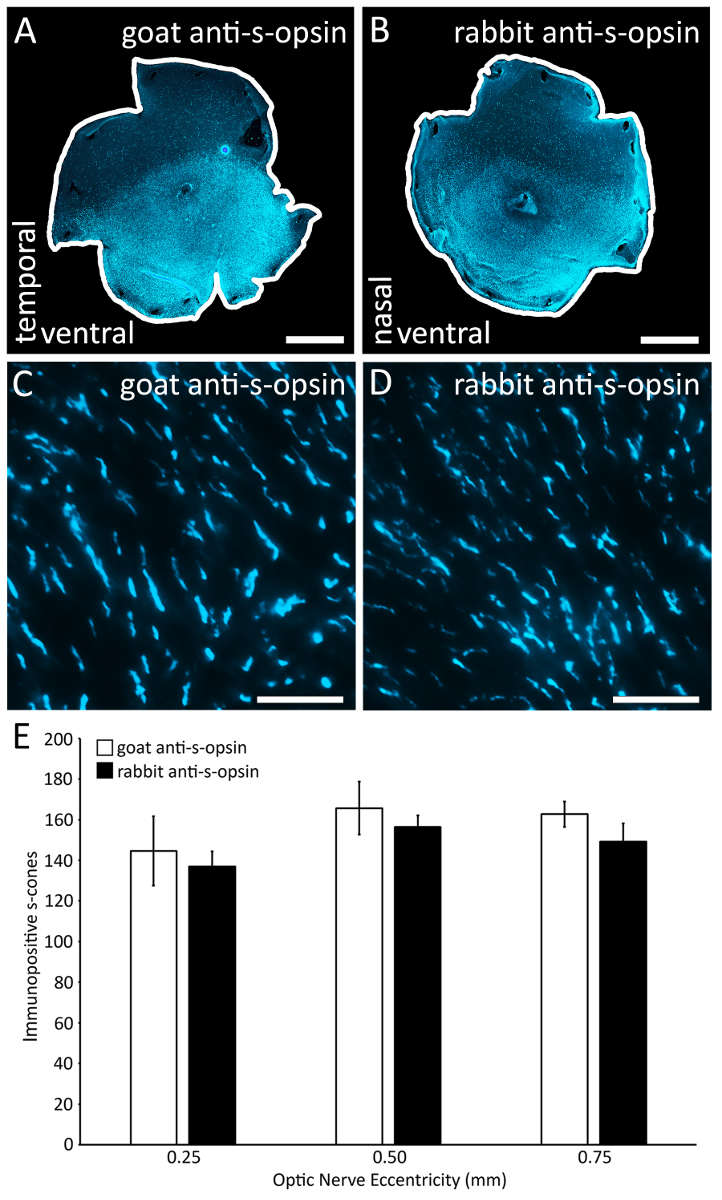{kind=link}

Figure 5 : un guide visuel pour utiliser le logiciel de Retistruct pour reconstruire les rétines immunomarquage avec s-opsine. Rétine (A), A ouvert en Retistruct dont le contour est visible et une « larme » ajouté. Points de la « déchirure » sont indiquées par des flèches blanches superposées. Toutes les découpes de cette rétine sont arbitraires, car aucun point de repère particulier ne servait à marquer l’orientation rétinienne au cours de la dissection. Les touches importantes sont encadrés en rouge. (B) une rétine avec tous les « larmes » ajouté et la rétine dorsale identifié avec « D » sur le bord de la rétine. Notez que le bouton de « Reconstruire la rétine » est maintenant visible. Les touches importantes sont encadrés en rouge. (C) le processus de reconstruction de la rétine. L’intrigue polaire de la rétine reconstituée s’affiche sur la droite, montrant que le soulager des coupes cyan (flèches bleues superposées afin de préciser les endroits coupés). (D) le résultat final de l’exécution de la rétine par l’intermédiaire de Retistruct. La rétine wholemount original reste sur la gauche et la rétine reconstituée apparaît sur la droite. Les coupes relève sont visibles en cyan (flèches blanches superposées afin de préciser les endroits coupés). S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 6 : la fissure de muscle et de la choroïde rectus supérieur peut être utilisée pour orienter avec précision la rétine de souris. Un tracé polaire des angles obtenus de chaque muscle droit supérieur soulager coupures ou fissure choroïde coupe dans la rétine qui ont été reconstruits avec Retistruct. Soulager coupures ont été identifiés sur les rétines immunomarquage reconstruit s-opsine et leurs emplacements ont été comparés à l’emplacement du dégradé s-opsine. En utilisant le code MATLAB personnalisé à tourner avec précision les rétines, afin que la concentration la plus élevée de s-opsine coloration est situé sur le ventre, la vraie dorsale (90° pour rectus supérieur), vrai nasal (0° pour fissure choroïde nasale) et vrai temporelle (180° pour la choroïde temporelle fissure) ont été déterminés pour chaque rétine. La valeur de chaque relaxation individuelle, angle de coupe a été déterminée dans ImageJ et un angle moyen a été calculée pour chaque relève coupé type. Coupes de muscle rectus supérieur identifiés le pôle dorsal à 96,3 ± 4,3 ° (n = 11). La fissure choroïde nasale identifié le pôle nasal à 6,7 ± 5,8 ° et la fissure choroïde temporelle du pôle temporal à 172,0 ± 4,5 ° (n = 9). S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
| Landmark profonde | Localisation de la brûlure cornéenne | Pôle de rétine identifié | Application à titre expérimental |
| Rectus supérieur | Dorsale | Dorsale | Webcam live ou fixe |
| Fissure choroïde nasale | Dorsale | Nasal | Webcam live ou fixe |
| Fissure choroïde temporelle | Dorsale | Temporelle | Webcam live ou fixe |
| S-opsine Gradient | Aucun | Dorsale, ventrale, nasale, temporelle | Fixe |
Tableau 1 : Repères profondes, le pôle de la rétine, ils s’identifient, et qu’ils peuvent être utilisés pour l’application de tissus vivants ou fixe.
Discussion
Il n’y a eu aucun protocole exhaustif et normalisé pour la détermination et l’orientation de la rétine de souris isolées dans l’espace anatomique d’étiquetage. Le protocole ici tente de combler cette lacune en normalisant et détaillant l’utilisation de repères anatomiques profondes comme points de référence fiable identifier l’orientation rétinienne. Il a été démontré que les repères anatomiques profondes dans ce protocole fournissent une méthode plus précise et fiable pour orienter la rétine de souris que superficielles monuments tels que brûlure cornéenne22. Ainsi, les études qui sont sont appuyés sur la cornée brûlures rétiniennes orientation ont pu plus erreurs dans l’orientation que les études qui se sont appuyés sur des sites tels que les muscles rectus et fissure choroïde. Cette différence met en évidence la nécessité et l’importance du présent protocole standardisé en ce qui concerne l’interprétation des résultats et de faire des comparaisons entre les études qui dépendent de l’orientation exacte rétinienne. Dans l’ensemble, un protocole standardisé fournira une méthode commune pour les chercheurs de la vision à suivre, ce qui évite la présence d’une variable confusionnelle en acquisition de données qui peut-être se produire avec l’utilisation de méthodes non normalisées pour l’identification rétinienne orientation.
Les méthodes présentées ici sont facilement reproductibles et applicable à de nombreux types de protocoles expérimentaux. En fait, un des grands avantages de ce protocole est sa capacité d’adaptation. Parce que la fissure choroïde, l’expression de s-opsine et points de repère rectus muscle ont été jugées pour identifier de façon fiable l’orientation rétinienne22 le point de repère qui adapte les paramètres expérimentaux peut être choisi pour optimiser l’acquisition de données (Table 1). en outre, les méthodes de dissection peuvent être combinés afin de clarifier davantage l’orientation de la rétine. Par exemple, fissure choroïde coupes peuvent être combinés avec s-opsine immunohistochemistry pour orienter tous les quatre pôles de la rétine : hémisphères nasales et temporelles peuvent être identifiés par les réductions de fissure choroïde, et s-opsine immunohistochimie permet d’identifier hémisphères ventrales et dorsales. Pourtant, la capacité d’adaptation du présent protocole peut-être être limitée par la nature sensible au temps des expériences de physiologie. Parce que le temps que nécessaire pour identifier un point de repère, établir une brûlure cornéenne et exécuter une coupe relève pourrait entraîner la mort tissulaire importante dans les expériences de ex vivo , certaines de ces méthodes de dissection peuvent être moins qu’optimales. Heureusement, une fois un dissecteur s’est familiarisé avec la fissure choroïde ou méthode de dissection de muscle rectus supérieur, identifier les repères profondes et la soulager coupures rapidement devient une partie de la routine de dissection et n’ajoutent pas de manière significative à la longueur de la dissection. Même si nous reconnaissons que les étapes détaillées ici peuvent ajouter ponctuellement à des expériences extrêmement sensibles au temps, nous conseillons d’utiliser le gradient de s-opsine pour post hoc rétinienne orientation lorsque la viabilité du tissu n’est plus un problème (Figure 3 ). La coloration de la rétine pour s-opsine est un moyen efficace d’orienter la rétine, comme il peut identifier tous les quatre pôles : s-opsine coloration divise pôles dorsales et ventrales de la rétine et permet pour identification des nasales et temporelle pôles selon que la rétine est d’un œil droit ou gauche (Figure 3). Par conséquent, nous croyons que ce protocole offre un ensemble fiable et reproductible des méthodes pour l’orientation exacte rétinienne qui peut s’acquitter de tous les paramètres expérimentaux.
Comme avec toute dissection rétinienne modifiée, la validité de la méthode de dissection est limitée par l’exactitude de la dissecteur et la qualité du tissu qui a été isolé. Si n’importe quel tissu est perdu au cours de la dissection ou la rétine est trop tronquée pour reconstruction exacte, Retistruct et le programme MATLAB sera pas en mesure de manière fiable reconstruire ou orienter la rétine. Il est donc important de pratiquer la méthode de dissection avant de l’utiliser pour la collecte de données des expériences. Tandis que les types de dissections expliquées ici ne sont pas difficiles, ils doivent être pratiqués afin d’assurer la répétabilité d’identification rétinienne orientation avec un point de repère particulier. En outre, il est essentiel que la pratique de dissecteur identifier visuellement les repères anatomiques avant de commencer la collecte de données pour s’assurer que le point de repère correct est utilisée. Une façon de vérifier l’exactitude d’un dissecteur particulière est de faire soit fissure choroïde coupes ou muscle droit supérieur coupe et ensuite comparer l’emplacement des incisions au dégradé s-opsine, puisqu’elle est un repère fixe et n’est donc pas tributaire de l’exactitude des dissectio n. dissecteurs potentiels peuvent également comparer leurs rétines reconstruits aux exemples des rétines reconstituées avec point de repère précis coupes sont indiquées à la Figure 1 et la Figure 2. Essentiellement, un dissecteur potentiel doit effectuer les étapes décrites dans le présent protocole pour un type particulier de dissection, qu’il s’agisse du muscle droit supérieur ou choroïde fissure méthode et comparer les résultats au dégradé s-opsine pour établir la validité d’un dissecteur particulière. Car si le dissecteur est incertain au sujet de l’emplacement de l’historique, il peut se produire dans une orientation erronée de la rétine qui permettront, par défaut, affecte interprétation et collecte de données.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous tenons à remercier un jour Bretagne, Jessica Onyak pour leur assistance technique et le Dr Liu de bien vouloir nous laisser utiliser son microscope à épifluorescence. Remerciements de soutien : R15EY026255 NIH-01 et la Fondation de Kirchgessner Karl.
matériels
| Name | Company | Catalog Number | Comments |
| 0.1 M Phosphate Buffered Saline | Sigma-Aldrich | P5244 | |
| Axioplan2 Epifluorescent Microscope | Zeiss | N/A | |
| Clear Nailpolish | N/A | N/A | |
| Corning LSE Low Speed Orbital Shaker | Sigma-Aldrich | CLS6780FP | |
| Costar TC-Treated 24-well Plates | Sigma-Aldrich | CLS3524 | |
| Dissection Microscope | Olympus | SZ51 | |
| Donkey anti-Goat Alexa 594 | Life Technologies | A11058 | |
| Donkey anti-Rabbit Alexa 594 | Life Technologies | A21207 | |
| Donkey Normal Serum | Millipore | 566460 | Use at 5.2% (52 μL with 86 μL of 20% Triton X-100 and 863 μL of 0.1M PBS for 1 mL of blocking solution) |
| Fisherbrand Superfrost Plus Microscope Slides | Fisher Scientific | 12-550-15 | |
| Goat anti-s-opsin | Santa Cruz Biotechnologies | sc-14363 | Not commerically available as of 2017 |
| Graefe Curved Forceps | Fine Science Tools | 11052-10 | |
| ImageJ or FIJI | National Institute of Health | N/A | Freely available software |
| Low Temperature Cautery Ophthalmic Fine Tip Cauterizer | Bovie Medical Corporation | AA00 | |
| MATLAB | MathWorks | N/A | At least version 2007b or later |
| Micro Cover Glasses | VWR International | 48393-241 | |
| Micro Slide Trays | VWR International | 82020-913 | |
| Moira Ultra Fine Forceps | Fine Science Tools | 11370-40 | |
| Nitrocellulose membrane | Millipore | HAWP04700 | |
| Paraformaldehyde | Electron Microscopy Sciences | 15714-S | Use at 4% (25 μL and 875 μL of 0.1 M PBS for 1 mL of fixative) |
| PrecisionGlide Needle 20G (0.90 mm x 25 mm) | BD PrecisionGlide | 305175 | |
| Pyrex Glass Petri Dish | Sigma-Aldrich | CLS3160152 | |
| R | The R Project for Statistical Computing | N/A | Freely available software; version 3.4.3 or later |
| Rabbit anti-s-opsin | Millipore | ABN1660 | |
| Retiga R3 Microscope Camera | Qimaging | 01-RET-R3-R-CLR-14-C | |
| Retistruct | N/A | N/A | Freely available software compatiable with Windows 7 or Windows 10 |
| Shandon Aqua-Mount Slide Mounting Media | Fisher Scientific | 14-390-5 | |
| Triton X-100 | Sigma-Aldrich | T8787 | Use 1.7% (86 μL of 20% Triton-X with 52 μL of Donkey Normal Serum and 863 μL of 0.1 M PBS for 1 mL of blocking solution) |
| Vannas Spring Dissection Scissors | Fine Science Tools | 15000-03 | |
| 5MP USB Microscope Digital Camera | AmScope | MU500 | To be used with the Olympus Dissection Microscope |
Références
- Applebury, M. L., et al. The murine cone photoreceptor: A single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 27 (3), 513-523 (2000).
- Hughes, S., Watson, T. S., Foster, R. G., Peirson, S. N., Hankins, M. W. Nonuniform distribution and spectral tuning of photosensitive retinal ganglion cells of the mouse retina. Curr Biol. 23 (17), 1696-1701 (2013).
- Sondereker, K. B., Onyak, J. R., Islam, S. W., Ross, C. L., Renna, J. M. Melanopsin ganglion cell outer retinal dendrites: Morphologically distinct and asymmetrically distributed in the mouse retina. J Comp Neurol. 525 (17), 3653-3665 (2017).
- Bleckert, A., Schwartz, G. W., Turner, M. H., Rieke, F., Wong, R. O. L. Visual space is represented by nonmatching topographies of distinct mouse retinal ganglion cell types. Current Biology. 24 (3), 310-315 (2014).
- Warwick, R. A., Kaushansky, N., Sarid, N., Golan, A., Rivlin-Etzion, M. Inhomogeneous Encoding of the Visual Field in the Mouse Retina. Curr Biol. 28 (5), 655-665 (2018).
- Valiente-Soriano, F. J., et al. Distribution of melanopsin positive neurons in pigmented and albino mice: evidence for melanopsin interneurons in the mouse retina. Front Neuroanat. 8, 131(2014).
- Sabbah, S., et al. A retinal code for motion along the gravitational and body axes. Nature. 546 (7659), 492-497 (2017).
- Vaney, D. I., Sivyer, B., Taylor, W. R. Direction selectivity in the retina: Symmetry and asymmetry in structure and function. Nat Rev Neurosci. 13 (3), 194-208 (2012).
- Huberman, A. D., et al. Genetic identification of an On-Off direction-selective retinal ganglion cell subtype reveals a layer-specific subcortical map of posterior motion. Neuron. 62 (3), 327-334 (2009).
- Ueki, Y., Ramirez, G., Salcedo, E., Stabio, M. E., Lefcort, F. Loss of Ikbkap causes slow, progressive retinal degeneration in a mouse model of familial dysautonomia. eNeuro. 3 (5), (2016).
- Maiorano, N. A., Hindges, R. Restricted perinatal retinal degeneration induces retina reshaping and correlated structural rearrangement of the retinotopic map. Nat Commun. 4, 1938(2013).
- Hadj-Said, W., et al. Quantitative and topographical analysis of the losses of cone photoreceptors and retinal ganglion cells under taurine depletion. Invest Ophthalmol Vis Sci. 57 (11), 4692-4703 (2016).
- Tao, Y., et al. The temporal topography of the N-Methyl- N-nitrosourea induced photoreceptor degeneration in mouse retina. Sci Rep. 5, 18612(2015).
- Risner, M. L., Pasini, S., Cooper, M. L., Lambert, W. S., Calkins, D. J. Axogenic mechanism enhances retinal ganglion cell excitability during early progression in glaucoma. Proc Natl Acad Sci U S A. , (2018).
- Estevez, M. E., et al. Form and function of the M4 cell, an intrinsically photosensitive retinal ganglion cell type contributing to geniculocortical vision. J Neurosci. 32 (39), 13608-13620 (2012).
- Kolesnikov, A. V., Kefalov, V. J. Transretinal ERG recordings from mouse retina: Rod and cone photoresponses. J Vis Exp. (61), (2012).
- Lin, B., Wang, S. W., Masland, R. H. Retinal ganglion cell type, size, and spacing can be specified independent of homotypic dendritic contacts. Neuron. 43 (4), 475-485 (2004).
- Ortin-Martinez, A., et al. Number and distribution of mouse retinal cone photoreceptors: differences between an albino (Swiss) and a pigmented (C57/BL6) strain. PLoS One. 9 (7), 102392(2014).
- Zhang, H., et al. The degeneration and apoptosis patterns of cone photoreceptors in rd11 Mice. J Ophthalmol. 2017, 9721362(2017).
- Wei, W., Elstrott, J., Feller, M. B. Two-photon targeted recording of GFP-expressing neurons for light responses and live-cell imaging in the mouse retina. Nat Protoc. 5 (7), 1347-1352 (2010).
- Wang, J., et al. Anatomy and spatial organization of Muller glia in mouse retina. J Comp Neurol. 525 (8), 1759-1777 (2017).
- Stabio, M. E., et al. A novel map of the mouse eye for orienting retinal topography in anatomical space. J Comp Neurol. 526 (11), (2018).
- Lamb, T. D., Collin, S. P., Pugh, E. N. Evolution of the vertebrate eye: Opsins, photoreceptors, retina and eye cup. Nat Rev Neurosci. 8 (12), 960-976 (2007).
- Sterratt, D. C., Lyngholm, D., Willshaw, D. J., Thompson, I. D. Standard anatomical and visual space for the mouse retina: Computational reconstruction and transformation of flattened retinae with the Retistruct package. PLoS Comput Biol. 9 (2), 1002921(2013).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.