
Method Article
Purification de H3 et H4 histones et la Quantification des histones acétylées marques dans les cellules et les tissus cérébraux
Dans cet article
Résumé
Le but de cet article est de fournir un guide complet et systématique à l’épuration efficace des histones H3 et H4 et la quantification des résidus d’histones acétylées.
Résumé
Dans tous les organismes eucaryotes, chromatine, le modèle physiologique de l’information tout génétique, est essentielle pour l’hérédité. La chromatine est soumises à un ensemble de diverses modifications post-traductionnelles (MPT) pour la plupart se produisent dans les terminus aminés d’histones (c.-à-d., histone queue) et de réguler l’accessibilité et l’état fonctionnel de l’ADN sous-jacente. Queues d’histone s’étendent depuis le cœur du nucléosome et sont sous réserve de l’ajout de groupements acétyles par histones acétyltransférases (chapeaux) et la suppression des groupes d’acétyle par les histones désacétylases (HDAC) au cours de la croissance cellulaire et la différenciation. Patrons d’acétylation spécifiques sur les résidus de lysine (K) sur des queues d’histone déterminer une homéostasie dynamique entre chromatine transcriptionnellement active ou transcriptionnellement réprimée par (1) qui influencent l’assembly d’histone de base et (2) recruter des synergique ou protéines associées à la chromatine antagonistes sur le site de transcription. Le mécanisme de régulation fondamental de la nature complexe de la queue de l’histone SPTM influe sur la plupart des processus basés sur les modèles de la chromatine et entraîne des changements dans la maturation cellulaire et la différenciation dans le développement normal et pathologique. L’objectif du présent rapport est de fournir une méthode efficace pour purifier les histones de noyau de cellules et de tissus du cerveau et de quantifier fiablement marques acétylation des histones H3 et H4 de novices.
Introduction
L’épigénétique de l’expression fait référence aux changements héréditaires dans l’activité des gènes qui se produisent indépendamment des variations de l’ADN de séquence1,2. La répression et la transcription de gènes sont déterminés par (1) l’accessibilité de l’ADN chromosomique enroulé autour d’un octamère d’histones de noyau (deux exemplaires, chacun des H2A, H2B, H3 et H4) et (2) la disponibilité des facteurs de transcription et protéines d’échafaudage recruté au promoteur spécifique sites3,4. La transcription de gènes est réglementée par l’enzyme-négociée des modifications des sites du promoteur ADN spécifiques et à la SPTM histone queues5,6,7. Les extrémités N-terminales des histones H3 et H4 sont parmi les plus hautement conservées séquences connues dans les organismes eucaryotes,3, et leurs modifications post-traductionnelles ont été largement documentées pour jouer un rôle central dans la détermination de structure de la chromatine et fonction8,9. SPTM à des queues d’histone (c.-à-d., acétylation, méthylation, la phosphorylation et l’ubiquitination) change le potentiel d’interaction des queues, influence l’état structural et pliage de la fibre de chromatine et par conséquent, réglemente l’accessibilité de l’ADN ainsi que4,10,11,12. Groupes d’acétyle sont ajoutés et supprimés des résidus K sur les queues d’histone par un ensemble d’interaction histones épigénétique des enzymes particulières, nommément chapeaux et HDACs, respectivement13. Par exemple, l’acétylation de l’histone H4 sur la lysine 12 (H4K12ac) a été précédemment démontrée pour activer la transcription des gènes associés à mémoire acquisition et consolidation des14. En outre, plusieurs éléments de preuve indiquent que le contrôle épigénétique induite par l’enzyme de la transcription du gène est un aspect crucial de santé cellulaire croissance et différenciation6,15. Alternance dans la régulation épigénétique de l’expression des gènes, par une mutation de l’épigénétiques enzymes elles-mêmes, ou par des modifications épigénétiques de l’ADN s’est avéré être la dysrégulation dans les maladies humaines où le changement dans un gène particulier activité est une caractéristique de la pathologie (par exemple, cancer)6,16,17. Ainsi, l’évaluation des changements dans les histones de noyau SPTM émerge comme une cible de grande valeur pour des interventions thérapeutiques potentielles. Toutefois, déterminer l’abondance, partenaires qui interagissent et les rôles spécifiques des histones SPTM ont prouvé difficile18.
Dans le présent rapport, une stratégie optimisée, débit moyen pour purifier les histones de noyau de cellules et de tissus cérébraux dans une seule fraction et un protocole complet pour la quantification des histones H3 et H4 SPTM est décrite. À noter, bien qu’actuellement publiée à base d’acide techniques de purification et de stratégies de détection des anticorps histone ont été largement adoptés pour la caractérisation de l’histone, ils manquent de détails descriptifs concernant les étapes critiques de la procédure, ainsi entraver la quantification et l’extraction de l’histone rapide et reproductible. Par exemple, extraire de la transformation des cellules et des biopsies tissulaires nécessite différents outils et technologies d’extraction réussie. En outre, le protocole optimisé présenté dans le manuscrit actuel montre une approche pratique, débit moyen. Core-histones sont extraits sous forme de fraction simple, pure, qui permet la détection humoral en aval fiable de PTM sans aucune interférence des impuretés. En outre, dans le manuscrit actuel, les défis concernant la détection des histones en raison de leur faible poids moléculaire ont été contournées. En règle générale, le manque de compatibilité entre purification, quantification et protocoles d’électrophorèse de gel d’entraver scientifiques d’obtenir des résultats reproductibles et péremptoire. Est présenté ici, un flux de travail optimisé pour purifier les histones de noyau de cellules et de tissus et les préparer pour en aval PTM analyse par transfert western.
Le protocole actuel permet la purification des protéines histones de noyau tout en préservant leurs modifications post-traductionnelles indigènes (c.-à-d., acétylation, méthylation et phosphorylation). La figure 1 illustre la chronologie du protocole de purification histone.
Protocole
Toutes les souris étaient logés dans une humidité et température-contrôlée, accrédité par AAALAC animalerie à l’Université de Miami Miller School of Medicine. Toutes les expériences ont été approuvés par l’Université de Miami Miller School of Medicine institutionnels Animal Care et le Comité de l’emploi (IACUC) et réalisées selon les spécifications des NIH.
1. préparation de l’extrait échantillon
-
Cellules adhérentes
- Cellules de plaque dans les plats de 10 cm dans le milieux de culture appropriés (1 x 106 à 1 x 109 cellules par plat pour les lignées cellulaires, tels que BV2, HEK-293 et SH-SY5Y, mais ~ 1 x 1015 cellules par plat pour éléments primaires, tels que les neurones du cortex primaires). S’assurer que les cellules sont également répartis sur toute la surface de la plaque et laisser les cellules à se développer pendant 48 h atteindre le confluent de ~ 100 % (37 ° C, 5 % de CO2).
- Lorsque les cellules atteignent la confluence désiré, délicatement aspirer les milieux de culture et laver les cellules 2 x avec préchauffée médias sans sérum sous une hotte de culture de tissus.
- Aspirer les médias sans sérum du plat et ajouter 1 mL de tampon d’extraction glacee (0,4 M, acide sulfurique, 1 mM KCl, 1 mM MgCl2, 50 mM Tris-HCl [pH 8.0] et 1 x cocktail inhibiteur de protéase) à chaque plat.
- Utilisez un grattoir de cellules en plastique pour recueillir toutes les cellules dans le tampon d’extraction (en grattant) et les transférer à un 1,5 mL étiqueté tube avec une pipette 1 000 µL. Pipette les cellules haut et bas 3 x pour faciliter l’homogénéisation.
- Fermez tous les tubes et les mettre immédiatement sur la glace.
-
Tissu cérébral
- Si les tissus congelés sont utilisé, placer le tissu dans un tube de prérefroidies 1,5 mL et Décongelez-le brièvement sur la glace. Si on utilise les tissus frais, passer immédiatement au point 1.2.2.
Remarque : Le protocole actuel décrit les procédures à l’aide de cerveau de souris congelées et échantillons de cortex préfrontal de souris. - Homogénéiser les tissus à l’aide d’un ordinateur de poche homogénéisateur Dounce en utilisant la quantité appropriée de tampon d’extraction et le nombre recommandé d’accidents vasculaires cérébraux (tableau 1). Pour éviter le déchiquetage de chromatine excessive, ne pas dépasser le nombre de traits recommandées.
- À l’aide d’une pipette µL monocanal 1 000, transférer l’homogénat dans un tube de prérefroidies 1,5 mL. Fermez tous les tubes et les mettre immédiatement sur la glace.
- Si les tissus congelés sont utilisé, placer le tissu dans un tube de prérefroidies 1,5 mL et Décongelez-le brièvement sur la glace. Si on utilise les tissus frais, passer immédiatement au point 1.2.2.
2. préparation de l’extrait brut Histone
- Placer les tubes de 1,5 mL contenant les cellules ou tissus en suspension dans le tampon d’extraction sur une plate-forme tournante et faire tourner à 15 t/min à 4 ° C pour permettre l’extraction de pétrole bruts histones.
NOTE : Le temps d’extraction de cellule différente et les types de tissus peut être différente et doit être optimisé pour chaque procédure. Le protocole actuel présente des résultats obtenus après 15 min, 2 h et 24h d’extraction (Figure 2, Figure 3, Figure 4et Figure 5). - Utiliser son au préalable la microcentrifugeuse à 4 ° C. Après l’extraction désirée le temps a passé, centrifuger les tubes à vitesse maximale pendant 10 min à 4 ° C.
- Transférer le surnageant, y compris les histones bruts pour un tube neuf, prérefroidies 1,5 mL. Jeter le culot.
- Stocker le surnageant à-80 ° C (l’extraction peut être arrêtée à cette étape, voir « Étape Stop » à la Figure 1) ou procéder immédiatement à l’étape suivante.
- Neutraliser les histones bruts avec un volume de 1/4 de 5 x tampon de neutralisation (par exemple, ajouter 250 µL de 5 x tampon de neutralisation dans 1 mL d’histones bruts). Mélangez bien en pipettant également, haut et bas x 6.
- Vérifier le pH du mélange avec des bandelettes de pH. Ajuster en conséquence en ajoutant plus tampon de neutralisation pour atteindre un pH de 7.
- Évaluer la présence de l’histone et protéines non-histones dans l’histone brut extrait comme suit (Figure 2).
- Ajouter 37,5 µL de l’échantillon à 12,5 µL de 4 x (Laemmli) solution tampon et dénaturer pendant 10 min à 99 ° C.
- Charger l’échantillon sur un gel SDS-PAGE et exécutez le gel pendant 1 h à 100 V.
- Souillez le gel durant la nuit avec la solution colorante R-250 brillant bleu de Coomassie et décolorer pendant trois lavages consécutifs (1 h/lavage) avec la solution de décoloration bleu de Coomassie brillant bleu R-250.
NOTE : Histones bruts (Figure 2) peuvent être comparés avec les histones éluées et purifiées (c'est-à-direentrée de colonne [Figure 5]).
3. purification des Histones principales
-
Rotation d’équilibration de la colonne
- Ajouter 500 µL de tampon de l’équilibration à chaque colonne utilisée. Ne touchez pas la membrane de la colonne.
- Centrifuger à 4 ° C pendant 3 min à 800 g. Jeter le cheminement. Répéter 1 x.
-
Purification de l’histone
- Ajouter 500 µL de l’échantillon de l’étape 2.6 à la colonne. Centrifuger à 4 ° C pendant 3 min à 800 g. Recueillir le cheminement.
- Répétez l’étape précédente autant de fois que nécessaire pour charger la totalité de l’échantillon sur la colonne. Ne pas surcharger la colonne.
- Combiner le cheminement de chaque étape de centrifugation afin d’analyser l’efficacité de la liaison de colonne (Figure 3).
- Suivez l’étape 2.7 pour analyser la colonne intermédiaire.
-
Lavage de colonne
- Ajouter 500 µL de la mémoire tampon de lavage à chaque colonne. Centrifuger à 4 ° C pendant 3 min à 800 g. Recueillir le cheminement lavage (#1).
- Répétez l’étape 3.3.1 pour un total de trois lavages. Recueillir les lavages intermédiaires #2 et #3. Piscine pas les colonne consécutive accréditives lavages.
- Pour continuer à évaluer l’efficacité de la colonne histone-liaison, analyser les lavages de trois colonnes en suivant l’étape 2.7 (Figure 4).
-
Élution de l’histone
- Transférer la colonne dans un nouveau tube marqué 1,5 mL.
- Ajouter 50 µL de tampon d’élution de l’histone. Centrifuger à 4 ° C pendant 3 min à 800 g. Sauver les cheminement contenant histones.
- Pour une élution supplémentaire, répétez l’étape 3.4.2. Ne pas combiner les premiers et deuxième cheminement éluats elles diffèrent dans la pureté et la quantité des histones.
4. la précipitation des Histones principales
- Ajouter de l’acide perchlorique (PCA) aux histones purifiées à une concentration finale de 4 % PCA (p. ex., ajouter 3 µL de 70 % PCA à 50 µL d’histones purifiées de l’étape 3.4.2.
- Centrifuger pendant 3 s pour recueillir tout le liquide résiduel de la paroi du tube. La composition de pipetage et descendre 6 x.
- Placer les tubes dans un rack et incuber pendant 24 h à 4 ° C.
- Le lendemain, utiliser son au préalable un microcentrifuge à 4 ° C et centrifuger les échantillons pendant 75 minutes à la vitesse maximale à 4 ° C.
- Après que la centrifugation est terminée, une petite pastille blanche contenant des histones précipités sera visible sur le fond du tube. Ne pas vortexer l’échantillon.
- Aspirer le surnageant avec précaution et, sans déranger le culot, ajouter 500 µL de glacé 4 % PCA à l’échantillon.
- Centrifuger à 4 ° C pendant 10 min à vitesse maximale. Soigneusement aspirer le surnageant.
- Répétez l’étape 4,7 2 x.
- Sans déranger le culot, ajouter 500 µL d’acétone glacée. Centrifuger à 4 ° C pendant 10 min à vitesse maximale. Soigneusement aspirer le surnageant.
- Répétez l’étape 4,9 2 x.
- Soigneusement aspirer le surnageant, laisser les tubes non écrêtées et laisser l’échantillon sec sur la glace pendant 30 min. vérifier si tous les acétone résiduel se soit évaporée.
- Laisser les tubes non écrêtée et laisser l’échantillon sécher à température ambiante (RT) pendant 5 min.
- Resuspendre le culot dans 30 µL d’eau stérile. Ne pas pipeter de haut en bas. Mettez le tube doucement avec un doigt.
- Bouchez tous les tubes et permettre les histones de reconstituer sur la glace pendant 30 à 50 min, selon la taille de granule. Vérifiez si le culot est remis en suspension.
- Bouchez tous les tubes et permettre le culot remettre en suspension plus à ta pendant 5 min.
NOTE : Cette solution (élution de premier et deuxième de l’étape 3.4.3) est constituée des histones purifiées et morue dessalées et peut être utilisée pour une analyse plus poussée quantification et histone acétylation.
5. quantification des protéines histones éluées
- Un spectrophotomètre selon le protocole du fabricant permet de quantifier les protéines histones total obtenus après l’élution finale à l’étape 4.15. Mesurer l’absorbance à 230 nm. Enregistrer le ratio A260/A280 indicatif de la contamination de l’échantillon avec de l’acide nucléique.
- Utilisez la formule suivante pour calculer la concentration d’histone (x) :

Ici, l’OD est la densité optique mesurée à A230 nm. - Une concentration d’histone de ~1.5 mg/mL est considéré comme un rendement moyen pour les lignées cellulaires, alors qu’une concentration d’histone de ~5.0 mg/mL est considéré comme un rendement moyen de 30 mg de tissu.
6. immunobuvardage
- Ajuster l’histone purifiée et éluée d’étape 4.15 à ~ 10 µg d’histone protéines/échantillon.
- Ajouter le volume approprié d’eau et 4 x Laemmli solution tampon pour ajuster les volumes de chargement.
- Dénaturer les échantillons pendant 10 min à 99 ° C. Refroidir sur glace. Centrifuger pendant 3 s pour recueillir tout le liquide résiduel et la condensation de la paroi du tube.
- Charger les échantillons sur un gel SDS-PAGE et exécutez le gel pendant 1 h à 100 V.
- Pour visualiser les protéines histones totales, souiller le gel durant la nuit avec la solution colorante R-250 brillant bleu de Coomassie et décolorer pendant trois lavages consécutifs (1 h/lavage) avec la solution de décoloration bleu de Coomassie brillant bleu R-250.
Remarque : La première élution des protéines histones contient qualité histones (Figure 5 a) tandis que l’élution deuxième contient peu à aucune concentration d’histones (Figure 5 b). - Afin de quantifier l’histone SPTM, utiliser un système de transfert (voir Table des matières) pour transférer la protéine histone du gel SDS-PAGE (étape 6.4) sur une membrane PVDF.
- Pour assembler le "sandwich" de transfert, ouvrir la cassette de système de transfert et placez la pile de membrane PVDF (étiquetée comme fond +) sur le fond de la cassette avec la membrane vers le haut. Rouler la membrane doucement avec un rouleau de la tache d’enlever tout l’air entre la pile et la membrane.
- Posez le gel sur le dessus de la membrane, rouler le gel doucement avec un rouleau de la tache d’enlever tout l’air entre la membrane et le gel et déposer la pile supérieure sur le gel. Rouler doucement à nouveau et placer le couvercle sur le dessus de la "sandwich", appuyer fermement et tourner le nob vers la droite pour verrouiller.
- Insérez la cassette à l’emplacement de système de transfert. Sur l’écran de l’appareil, sélectionnez Protocole Turbo. Utiliser un protocole de 3 min pour un simple gel mini ou un protocole de 7 min pour plus de deux minis gels.
- Colorent la membrane avec la tache de Ponceau S pendant 5 min et visualiser les histones totales.
- Lavez-les dans 1 x Tris salin (TBS) avec 0,1 % Tween-20 à 2 h et bloc en 5 % de lait pendant 1 h. Incuber avec des anticorps primaires et secondaires (une nuit à 4 ° C ou 1 h à RT, respectivement) ou selon un protocole préalablement optimisé.
Remarque : dans le protocole actuel, les anticorps contre les histones acétylées H4K12 (Figure 6 et Figure 7) et les H3K27 (Figure 8) ont été utilisés.
Résultats
Pour illustrer la progression de la protocole de purification des histones et la composition de toutes les fractions analysées, nous avons évalué l’histone différents extraits de cellules microgliales humaines BV2. Pour illustrer la quantification des histones H3 et H4 SPTM (c.-à-d., acétylation), nous avons utilisé les lysats de tissu de cerveau.
Les cellules BV2 étaient plaqués à 5 x 106 cellules par plat dans les plats traitée pour la culture de tissu de 10 cm et a permis de croître à confluence pendant 48 h. cellules ont été ensuite recueillis et histones ont été libérés de la chromatine par une incubation dans le tampon d’extraction contenant 0,4 M l’acide sulfurique, KCl 1 mM, 1 mM MgCl2, 50 mM Tris-HCl (pH 8,0) et 1 x inhibiteur de la protéase. Le temps d’extraction, entre 15 minutes et 24h, n’a pas affecté la composition globale des extraits bruts histone tel que déterminé par le bleu de Coomassie brillant bleu coloration (Figure 2). Les histones bruts, prochaines étaient insérées dans les colonnes de l’histone équilibré et le cheminement a été analysée. Rendement élevé de liaison de la colonne est déterminé par l’absence de protéine histone dans le cheminement lors de l’analyse par coloration au bleu de Coomassie brillant bleu. Nous avons déterminé l’efficacité de la liaison de colonne à 100 % car il n’y a aucune protéine histone détectables présentes dans l’analyse intermédiaire (Figure 3). Toutes les membranes avec des histones liées ont été lavés puis trois fois avec le tampon de lavage pour éliminer toutes les impuretés restantes, laissant seule histone protéines liées au gel de silice. Nous avons déterminé que, pour l’extraction de histone tous les temps (c.-à-d., 15 min, 2 h et 24h), le premier lavage de membrane a été le plus important enlever des contaminations non histones des colonnes, tandis que les deuxième et troisième lavages n’influencent pas la pureté de l’échantillon. Ainsi, selon le type de l’échantillon, les deux derniers lavages peuvent être omis. Après l’élution première des protéines histones de la colonne (à l’aide du tampon d’élution contenant 1mM NaCl et EDTA), histones ont été précipités du jour au lendemain avec 4 % d’acide perchlorique puis granulées, lavés et analysés pour l’enrichissement des histones purifiées H3 et H4. Nous avons observé que 24h d’extraction temps augmente le montant des histones H3 et H4 dans la fraction purifiée comparé à 15 min et 2 h de temps d’extraction (Figure 5 a). La deuxième élution de la colonne n’a pas entraîné les histones qualité ou haute-quantité (Figure 5 b).
Ensuite, nous avons utilisé des homogénats de tissu de cerveau de quantifier des histones H3 et H4 SPTM, à savoir l’acétylation. Sauvage (C57BL6/J) souris mâles ont administré un large agissant inhibiteur d’HDAC (tributyrine) à une dose de 5 g/kg par voie orale pendant 3 tissus d. ensemble du cerveau ont été recueilli le jour 4 et bruts histones ont été extraites selon le protocole décrit. À l’aide d’un non-appariés t-test, nous avons déterminé que tributyrine augmente l’acétylation des histones dans l’extrait brut (t(6) = 6.184, P = 0,0004) ; Toutefois, les impuretés sont détectées dans l’extrait (les bandes d’histones ne sont pas clairement définis). Ainsi, les anticorps H4K12ac n’a pas une grande spécificité (Figure 6). Pour continuer à évaluer l’applicabilité du protocole présenté à de petites sections de tissus, nous avons recueilli le cortex préfrontal de triple transgéniques la maladie d’Alzheimer (3 x Tg-AD) souris traitées tous les jours à 10 mg/kg M344, une classe I et inhibiteur d’HDAC IIb, pour quatre mois. Précipitations et purification de l’histone a été réalisée selon le protocole décrit dans les présentes. Utilisant la purifiée histone H3 et H4 fraction, nous avons déterminé que M344 augmente l’acétylation H4K12 2,4 fois (t(6) = 13.03, P < 0,0001), avec une grande spécificité de l’anticorps H4K12ac (Figure 7). De même, nous avons observé une augmentation de l’acétylation des histones H3 BV2 cellules en réponse à un autre inhibiteur d’HDAC, à savoir l’inhibiteur sélectif de la HDAC3, RGFP-966. 10 µM des causes RGFP-966 une environ deux fois plus d’acétylation à histone H3K27 après 24 h de traitement. Étudiant de non appariés t-test a été utilisé pour comparer des hématies par rapport aux traités.

Figure 1 : chronologie du protocole de purification histone. Toutes les mesures pour les analyses de l’histone figurent ci-dessous ainsi que le temps estimé nécessaire pour chaque étape. Les figures illustrant le résultat des mesures particulières et présenté dans le manuscrit figurent entre parenthèses. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 : gel de Coomassie brillant bleu teinté représentative démontrant des histones brutes extraites des cellules BV2. Les cellules BV2 ont été cultivés pendant 48 h avant que ne commence le protocole d’extraction d’histone. Les histones bruts ont été extraits pour 15 min, 2 h et 24h, avec trois répétitions pour chaque point dans le temps (c’est également le cas dans la Figure 3, Figure 4et Figure 5). Des protéines non histones et l’histone sont présents dans l’extrait brut d’histone. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
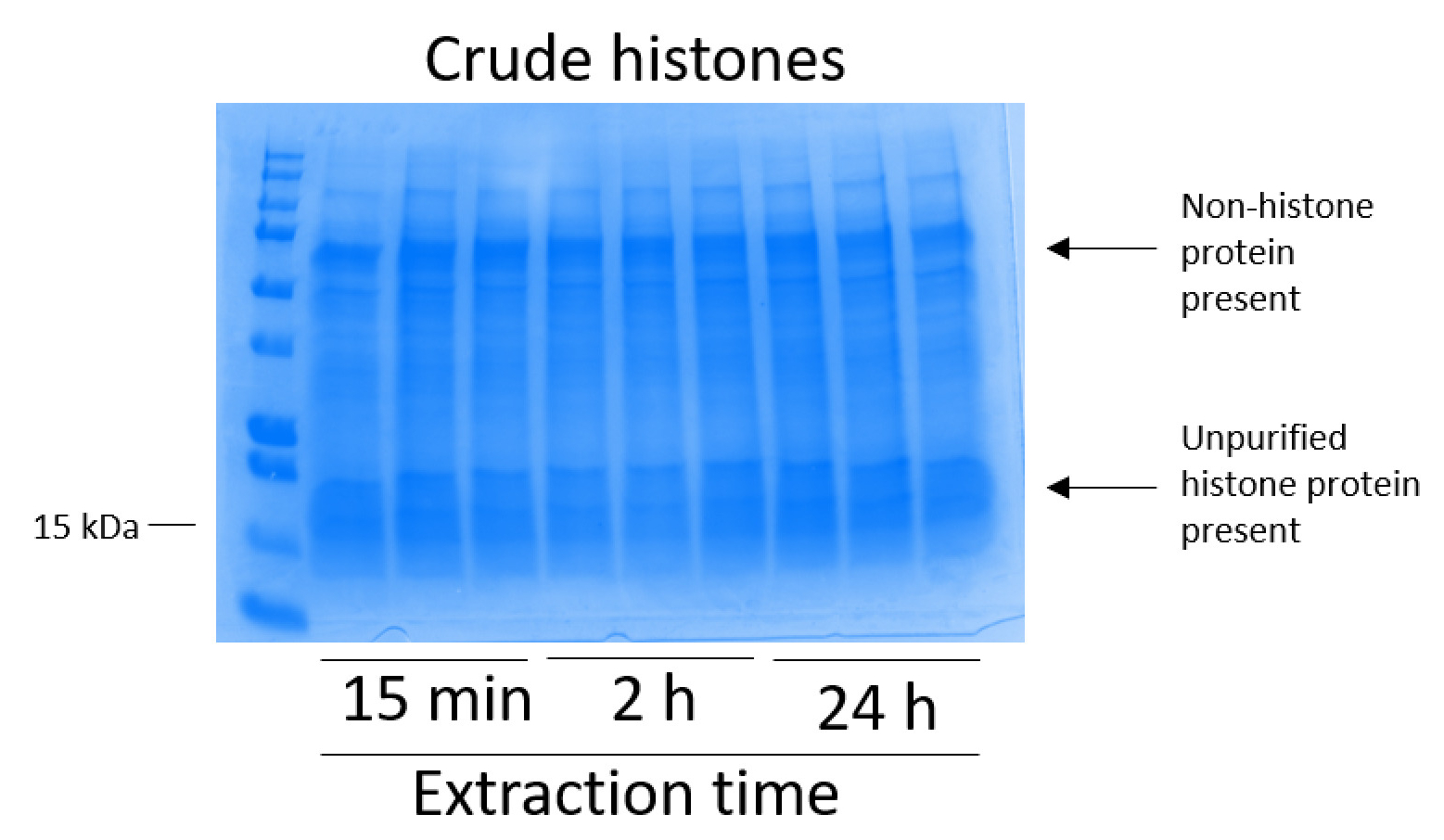{kind=link}

Figure 3 : gel de Coomassie brillant bleu teinté représentative démontrant la colonne intermédiaire suivant histone purification pas de cellules BV2. Suite d’histone brut en passant par la colonne de liaison d’histone, seulement des protéines non histones sont présents dans le cheminement. Histones sont absents dans cette fraction. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 4 : représentant gel teinté bleu de Coomassie brillant bleu, démontrant un lavage de colonne suite à une étape de purification histone de cellules BV2. Quel que soit les temps d’extraction histone, qui étaient (A) 15 min, (B), 2 h, ou (C), 24h, de faibles quantités de protéines non histones sont seulement présents dans la colonne de la première-lavage histonebinding. Histones étaient absents dans tous les lavages. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
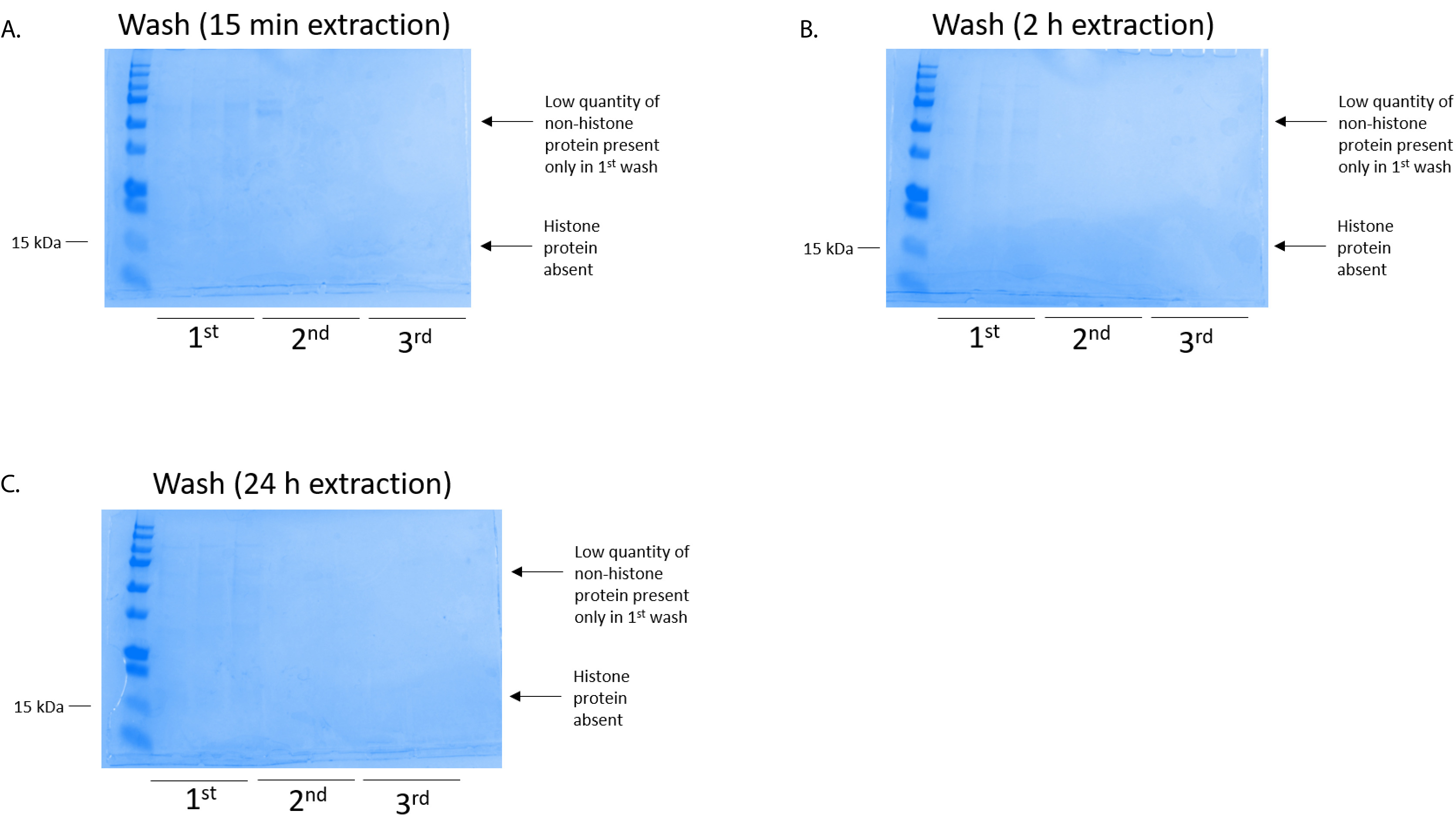{kind=link}

Figure 5 : gel de Coomassie brillant bleu teinté représentative démontrant elutions suite à une étape de purification histone de cellules BV2. (A) qualité purifiée et dessalé histones H3 et H4 sont détectés après l’élution de première de la colonne de purification des histones. Colonne (B) la deuxième élution de la purification de l’histone n’a donné une haute qualité ou quantité des histones H3 ou H4. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
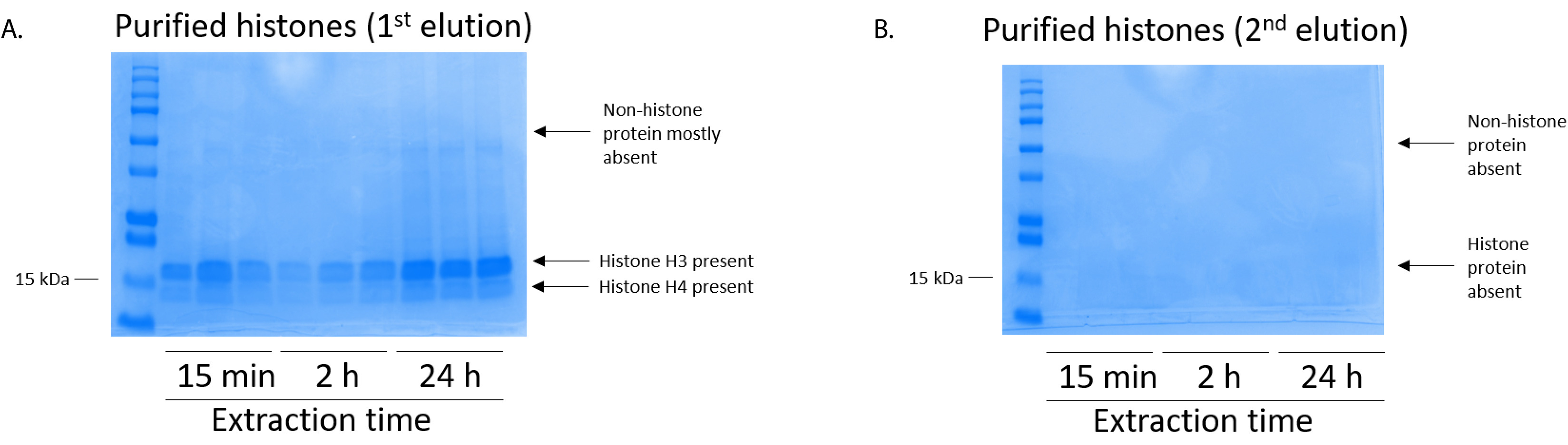{kind=link}

Figure 6 : l’agissant largement HDAC inhibiteur, tributyrine, augmente l’acétylation H4K12 dans tout le cerveau de souris de type sauvage. (A) ce panneau montre un représentant western blot, représentant une augmentation de H4K12 l’acétylation de l’histone brut extrait prélevés dans tout le cerveau de souris de type sauvage en réponse à l’HDAC largement par intérim inhibiteur, tributyrine. (B), ce panneau présente la quantification de l’augmentation H4K12 acétylation in vivo. Non appariés t-test a été utilisé pour comparer des groupes (t(6) = 6.076, P = 0,0005). Les barres représentent la moyenne ± l’écart-type de la moyenne (SEM). N = 8. P < 0,0001. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 7 : la classe I et inhibiteur d’HDAC IIb, M344, augmente l’acétylation H4K12 dans le cortex préfrontal triple transgénique la maladie d’Alzheimer (3 x Tg-AD) souris. (A) ce panneau montre un représentant western blot représentant une augmentation de H4K12 acétylation dans purifiés et extrait de morue dessalée histone récoltées dans le cortex préfrontal du 3 x souris Tg-AD en réponse à l’inhibition des HDACs par M344. (B) ce panneau montre la quantification de l’augmentation de l’acétylation H4K12 en réponse à M344 administré à une dose quotidienne de 10 mg/kg pendant quatre mois. Non appariés t-test a été utilisé pour comparer des groupes (t(6) = 13 h 30, P < 0,0001). Les barres représentent la moyenne ± SEM. N = 8. P < 0,00001. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 8 : l’inhibiteur sélectif de la HDAC3, RGFP-966, augmente l’acétylation H3K27 dans les cellules microgliales BV2. (A), ce tableau montre une tache occidentale représentante, représentant une augmentation de l’acétylation des H3K27 dans l’extrait purifié et morue dessalée histone prélevée BV2 cellules en réponse à l’inhibition de la HDAC3 de RGFP-966. (B) RGFP-966 provoque une environ deux fois plus d’acétylation de l’histone H3 et lysine (K) 27 après 24 h de traitement. Non appariés t-test a été utilisé pour comparer par rapport aux traités hématies (t(4) = 5.981, P = 0,002). Les barres représentent la moyenne ± SEM. N = 6. P < 0,01. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
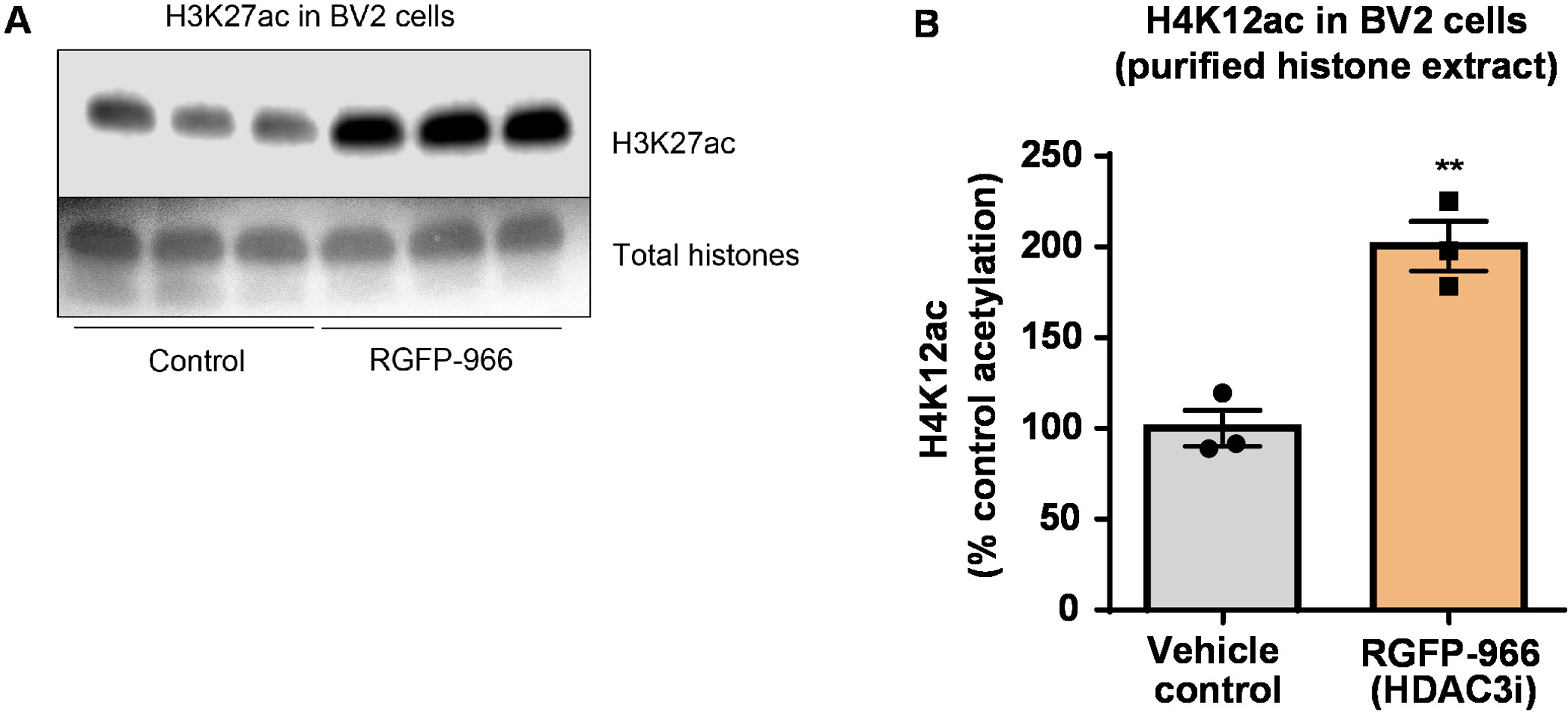{kind=link}
| Tissu représentant (un hémisphère) * | Tissu moyen poids (mg) | Tampon d’extraction (mL) | Nombre de traits |
| Cervelet de souris | 40 | 1 | 40 |
| Cortex frontal de souris | 30 | 0,3 | 20 |
| Hippocampe de souris | 27 | 0,3 | 18 |
| Cortex entorhinal souris | 19 | 0,3 | 17 |
| * Toutes les expériences ont été effectuées sur des souris mâles adultes. | |||
| Âge moyen : 16 mois. Poids moyen : 30 grammes. | |||
Tableau 1 : Condition optimisée pour l’homogénéisation de tissu de cerveau.
Discussion
Dans les travaux en cours, nous avons démontré une méthode optimisée pour purifier les histones core et quantifier des histones H3 et H4 SPTM (p. ex., acétylation). Le protocole présenté est un workflow complet qui intègre optimisé des procédures concernant les cellules et préparation de tissu de cerveau, élution de purification de l’histone et la précipitation de l’histone détaillée, brute et la quantification, qui sont suivis électrophorèse de l’histone et quantification de PTM histone robuste. La grande quantité de détails fournis ici permet une génération réplicable de données de grande qualité, malgré la nécessité de longues manipulations des échantillons histone.
De nombreux protocoles actuellement publiés nécessitent l’utilisation de la CLHP pour isoler les fractions pures des histones H3 et H419. Bien que l’HPLC est une technique puissante, sa complexité et son faible débit dissuader la plupart des biologistes moléculaires et des profanes de son utilisation fréquente. En effet, HPLC n’est pas disponible pour nombreux laboratoires et du personnel hautement qualifié est nécessaire pour faire fonctionner l’instrument. HPLC est souvent fastidieux, coûteux et potentiellement dangereux. Présenté ici est une stratégie peu coûteuse, débit moyen d’obtenir des résultats de qualité similaire qui contourne la CLHP. La stratégie signalée est également plus pratique et utilisable dans presque n’importe quel laboratoire car il utilise une approche de colonne essorage simple qui ne nécessite pas de compétences opération instrument spécialisé. En outre, tétramère de histone H3/H4 est extrait comme un simple, pure et la fraction abondante, ce qui permet une quantification fiable des conserves SPTM sur chacune des protéines.
SPTM est extrêmement sensibles aux changements dans le stress oxydatif et les changements de pH20,21. Ainsi, contrairement aux méthodes publiées antérieurement18, nous rapportons une stratégie efficace de rincer les cellules dans des milieux sans sérum pour assurer un minimum de remaniement métabolique des cellules et pour éviter l’interférence de la SPTM natif avec composants de sérum. Le protocole actuel non seulement permet d’éviter l’isolement des noyaux traditionnels, mais fournit également des temps optimaux pour lyse cellulaire et la procédure d’homogénéisation tissu exact qui permet la préservation de l’enveloppe nucléaire, tout en évitant l’agrégation nucléaire. Bien que le temps d’extraction peuvent être manipulé selon le nombre de cellules, le type de cellule utilisé, la taille de tissu, etc., lyse prolongée n’est pas souhaitable, car cela pourrait conduire à la lyse des noyaux et libération d’ADN, ce qui rend l’échantillon difficile à gérer. Ce qui est important, plusieurs points de contrôle au sein du protocole existent pour la validation de la purification de l’histone réussie (par exemple, les étapes 2,7 et 3.2.3). Cette stratégie facilite également tout au long de la longue procédure de dépannage.
Une autre caractéristique unique et importante du protocole présenté est sa compatibilité totale avec les outils d’analyse par transfert western en aval et d’autres si vous le souhaitez. Histones sont détectés à ~ 15 kDa13,22,23 et, de même à d’autres protéines de faible poids moléculaire, sont sont avérées difficiles à détecter par les techniques d’immunotransfert standard. L’utilisation d’un système de haute performance et haut débit de transfert en combinaison avec des gels de protéines résolution optimale permet le maintien de confirmation natif de protéine (en l’absence de la SDD) et de l’activité en l’absence de SDS et l’efficacité de transfert élevé des protéines de faible poids moléculaire histone, assurant ainsi une quantification de PTM histone fiable.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Les auteurs expriment leur gratitude à la Floride Ministère de la santé Ed et Ethel Moore Alzheimer Research Programme (subventions 6AZ08 et 7AZ26), le NIH-NIAAA (subvention 5R01AA023781-03) et l’American Heart Association (17PRE33660831 de concession).
matériels
| Name | Company | Catalog Number | Comments |
| 1.5 mL microcentrifuge tubes | ThermoFiser Scientific | 05-408-129 | or equivalent from other sources |
| Sterile water | Gibco | 15-230-204 | or equivalent from other sources |
| 70% perchloric acid | Sigma Aldrich | 311421 | or equivalent from other sources |
| 100% acetone | Sigma Aldrich | 270725 | or equivalent from other sources |
| pH-indicator strips, non-bleeding | Milliipore Sigma | 1095310001 | |
| 4x SDS sample buffer | BIO-RAD | 161-0747 | |
| Benchtop rotor | Cole-Parmer | UX-04397-34 | or equivalent from other sources |
| 1.5 mL tube rack | ThermoFiser Scientific | 05-541 | or equivalent from other sources |
| Histone purification mini kit | Active Motif | 40026 | spin columns included in the kit |
| Protease Inhibitor Cocktail | ThermoFiser Scientific | 78430 | or equivalent from other sources |
| Nanodrop instrument | ThermoFiser Scientific | ND-2000 | |
| Tissue culture dishes | VWR | 10062-880 | required for histone extraction from cultured cells |
| Tissue culute media | varies based on cell line used | varies based on cell line used | required for histone extraction from cultured cells |
| Low-serum media | ThermoFiser Scientific | 51985091 | required for histone extraction from cultured cells |
| Plastic cell scraper | Falcon | 353086 | required for histone extraction from cultured cells |
| SDS-PAGE gradient gel | BIO-RAD | 456-9035 | required for histone extraction from cultured cells |
| Coomassie Brilliant Blue R-250 Staining Solution | BIO-RAD | 1610436 | required for histone extraction from cultured cells |
| Coomassie Brilliant Blue R-250 Destaining Solution | BIO-RAD | 1610438 | required for histone extraction from cultured cells |
| Trans-Blot Turbo Mini PVDF Transfer Packs | BIO-RAD | 1704156 | required for histone extraction from cultured cells |
| Trans-Blot Turbo Transfer System | RIO-RAD | 1704150 | required for histone extraction from cultured cells |
| Ponceau S stain | CellSignalling | 59803S | required for histone extraction from cultured cells |
| Dounce homogenizer (size/cap sc 7mL) with a small size clearance | Kimble Chase | 885302-0007 | required for histone extraction from tissues |
| 100% bleach | Clorox | 68973 | required for histone extraction from tissues |
| H4K12ac antibody | Active Motif | 39166 | required for PTMs quantification via WB |
| H3K27ac antibody | Active Motif | 39134 | required for PTMs quantification via WB |
Références
- Holliday, R. Is there an Epigenetic Component in Long-term Memory? Journal of Theoretical Biology. 200, 339-341 (1999).
- DeWoskin, V. A., Million, R. P. The epigenetics pipeline. Nature Reviews Drug Discovery. 12, 661-662 (2013).
- Eberharter, A., Becker, P. B. Histone acetylation: a switch between repressive and permissive chromatin. EMBO Reports. 3, 224-229 (2002).
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature. 389, 349-352 (1997).
- Sartor, G. C., Powell, S. K., Brothers, S. P., Wahlestedt, C. Epigenetic Readers of Lysine Acetylation Regulate Cocaine-Induced Plasticity. The Journal of Neuroscience. 35, 15062-15072 (2015).
- Komatsu, N., et al. SAHA, a HDAC inhibitor, has profound anti-growth activity against non-small cell lung cancer cells. Oncology Reports. 15, 187-191 (2006).
- Bahari-Javan, S., Sananbenesi, F., Fischer, A. Histone-acetylation: a link between Alzheimer's disease and post-traumatic stress disorder. Frontiers in Neuroscience. 8, 160(2014).
- Roh, T. -Y., Cuddapah, S., Zhao, K. Active chromatin domains are defined by acetylation islands revealed by genome-wide mapping. Genes & Development. 19, 542-552 (2005).
- Mutskov, V., Felsenfeld, G. Silencing of transgene transcription precedes methylation of promoter DNA and histone H3 lysine 9. The EMBO Journal. 23, 138-149 (2004).
- Howe, L., Brown, C. E., Lechner, T., Workman, J. L. Histone acetyltransferase complexes and their link to transcription. Critical Reviews in Eukaryotic Gene Expression. 9, 231-243 (1999).
- Jenuwein, T., Allis, C. D. Translating the histone code. Science. 293, 1074-1080 (2001).
- Bowman, G. D., Poirier, M. G. Post-Translational Modifications of Histones That Influence Nucleosome Dynamics. Chemical Reviews. 115, 2274-2295 (2015).
- Volmar, C. -H., Wahlestedt, C. Histone deacetylases (HDACs) and brain function. Neuroepigenetics. 1, 20-27 (2015).
- Plagg, B., Ehrlich, D., Kniewallner, K. M., Marksteiner, J., Humpel, C. Increased Acetylation of Histone H4 at Lysine 12 (H4K12) in Monocytes of Transgenic Alzheimer's Mice and in Human Patients. Current Alzheimer Research. 12, 752-760 (2015).
- Bhaskara, S., et al. Hdac3 is essential for the maintenance of chromatin structure and genome stability. Cancer Cell. 18, 436-447 (2010).
- Mottamal, M., Zheng, S., Huang, T. L., Wang, G. Histone Deacetylase Inhibitors in Clinical Studies as Templates for New Anticancer Agents. Molecules. 20, 3898-3941 (2015).
- Ramakrishnan, S., et al. HDAC 1 and 6 modulate cell invasion and migration in clear cell renal cell carcinoma. BMC Cancer. 16, 617(2016).
- Wapenaar, H., Dekker, F. J. Histone acetyltransferases: challenges in targeting bi-substrate enzymes. Clinical Epigenetics. 8, 59(2016).
- Klinker, H., Haas, C., Harrer, N., Becker, P. B., Mueller-Planitz, F. Rapid Purification of Recombinant Histones. PLoS ONE. 9, e104029(2014).
- Chen, K., et al. Neurodegenerative Disease Proteinopathies Are Connected to Distinct Histone Post-translational Modification Landscapes. ACS Chemical Neuroscience. 9, 838-848 (2018).
- Simithy, J., Sidoli, S., Garcia, B. A. Integrating Proteomics and Targeted Metabolomics to Understand Global Changes in Histone Modifications. Proteomics. , e1700309 (2018).
- Volmar, C. -H., et al. An Epigenetic Approach for the Modulation of Amyloid Precursor Protein (APP) Processing and Improvement of Memory in Alzheimer's Disease. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 40, S470(2015).
- Volmar, C. -H., et al. M344 promotes nonamyloidogenic amyloid precursor protein processing while normalizing Alzheimer’s disease genes and improving memory. Proceedings of the National Academy of Sciences. 114 (43), E9135-E9144 (2017).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.