
Method Article
Cartographie du site de démarrage de la transcription à l'aide d'un transporteur-CAGE super-faible
Dans cet article
Résumé
Cap Analysis of Gene Expression (CAGE) est une méthode de cartographie quantitative à l'échelle du génome de l'ARNm 5'ends pour capturer les sites de transcription de la polymérase II de l'ARN à une résolution à un seul nucléotide. Ce travail décrit un protocole à faible apport (SLIC-CAGE) pour la génération de bibliothèques de haute qualité utilisant des quantités de nanogrammes d'ARN total.
Résumé
L'analyse du capuchon de l'expression génique (CAGE) est une méthode utilisée pour la détection de résolution à un nucléotide des sites de transcription de la polymérase II de l'ARN (TSS). La détection précise des SST améliore l'identification et la découverte des principaux promoteurs. En outre, les exhausteurs actifs peuvent être détectés par des signatures d'initiation de transcription bidirectionnelle. Décrit ici est un protocole pour effectuer super-faible porteur d'entrée-CAGE (SLIC-CAGE). Cette adaptation SLIC du protocole CAGE minimise les pertes d'ARN en augmentant artificiellement la quantité d'ARN grâce à l'utilisation d'un mélange de porteurs d'ARN transcrit in vitro qui est ajouté à l'échantillon d'intérêt, permettant ainsi la préparation de la bibliothèque à partir de nanogrammes-montants de total L'ARN (c.-à-d. des milliers de cellules). Le transporteur imite la distribution prévue de la longueur des fragments de la bibliothèque d'ADN, éliminant ainsi les biais qui pourraient être causés par l'abondance d'un transporteur homogène. Dans les dernières étapes du protocole, le transporteur est retiré par dégradation avec des endonucas homing et la bibliothèque cible est amplifiée. La bibliothèque d'échantillons cibles est protégée contre la dégradation, car les sites de reconnaissance de l'endouclène sont longs (entre 18 et 27 pb), ce qui rend la probabilité de leur existence dans les génomes eucaryotes très faible. Le résultat final est une bibliothèque d'ADN prête pour le séquençage de la prochaine génération. Toutes les étapes du protocole, jusqu'au séquençage, peuvent être complétées dans les 6 jours. La préparation du transporteur nécessite une journée de travail complète; cependant, il peut être préparé en grandes quantités et conservé congelé à -80 oC. Une fois séquencées, les lectures peuvent être traitées pour obtenir des TSS à résolution mononucléotide à l'échelle du génome. Les SST peuvent être utilisés pour la découverte de promoteurs ou d'améliorateurs de base, ce qui donne un aperçu de la régulation des gènes. Une fois agrégées aux promoteurs, les données peuvent également être utilisées pour le profilage d'expression centré sur 5'centrage.
Introduction
L'analyse du capuchon de l'expression génique (CAGE) est une méthode utilisée pour la cartographie à un seul nucléotide à l'échelle du génome des sites de transcription de la polymérase II de l'ARN (TSS)1. Sa nature quantitative permet également le profilage d'expression centré de 5'-fin. Les régions entourant les SST (environ 40 pb en amont et en aval) sont des promoteurs principaux et représentent l'emplacement physique où l'ARN polymérase II et les facteurs de transcription générale se lient (révisé s'il y a2,3). L'information sur l'emplacement exact des SST peut être utilisée pour la découverte des promoteurs de base et pour surveiller la dynamique des promoteurs. En outre, en tant que rehausseurs actifs présentent des signatures de transcription bidirectionnelle, les données CAGE peuvent également être utilisées pour la découverte et la surveillance de la dynamique des exhausteurs4. La méthodologie CAGE a récemment gagné en popularité en raison de son application et de son utilisation dans des projets de recherche de grande envergure tels que ENCODE5, modENCODE6, et les projets FANTOM7. En outre, l'information tSS s'avère également importante pour distinguer les tissus sains et malades, car les SST spécifiques à la maladie peuvent être utilisés à des fins diagnostiques8.
Même si plusieurs méthodes de cartographie TSS sont disponibles (CAGE, RAMPAGE, STRT, nanoCAGE, nanoCAGE-XL, oligo-capping), nous et d'autres avons récemment montré que CAGE est la méthode la plus impartiale pour capturer les vrais TSS avec le moins de faux positifs9 , 10. Le protocole CAGE récent, nAnT-iCAGE11, est le protocole le plus impartial pour le profilage TSS, car il évite de couper les fragments à des étiquettes courtes en utilisant des enzymes de restriction et n'utilise pas l'amplification PCR. Une limitation du protocole nAnT-iCAGE est l'exigence d'une grande quantité de matériel de démarrage (p. ex., 5 g d'ARN total pour chaque échantillon). Pour répondre à des questions spécifiques et pertinentes sur le plan biologique, il est souvent impossible d'obtenir des quantités aussi élevées de matériel de démarrage (p. ex., pour les cellules triées au FACS ou les stades embryonnaires précoces). Enfin, si nAnT-iCAGE réussit, seulement 1-2 ng de matériel de bibliothèque d'ADN est disponible de chaque échantillon, limitant ainsi la profondeur de séquençage réalisable.
Pour permettre le profilage TSS en utilisant seulement des nanogrammes d'ARN total, nous avons récemment développé Super-low Input Carrier-CAGE10 (SLIC-CAGE, Figure 1). SLIC-CAGE n'a besoin que de 10 ng d'ARN total pour obtenir des bibliothèques à haute complexité. Notre protocole s'appuie sur le porteur d'ARN synthétique soigneusement conçu ajouté à l'ARN d'intérêt pour atteindre un total de 5 g de matériel d'ARN. Le porteur synthétique imite la bibliothèque d'ADN cible dans la distribution de longueur pour éviter les biais potentiels qui pourraient être causés par des molécules homogènes en excès. La séquence du porteur est basée sur la séquence du gène de synthetase de leucyl-tRNA d'Escherichia coli (tableau 1) pour deux raisons. Tout d'abord, tout reste du porteur dans la bibliothèque finale, même s'il est séquencé, ne sera pas cartographié à un génome eucaryotique. Deuxièmement, comme E. coli est une espèce mésophile, ses gènes d'entretien ménager sont optimisés pour la plage de température appropriée pour SLIC-CAGE. La séquence de porteur est également incorporée avec des sites de reconnaissance d'endonuclease de homing pour permettre la dégradation spécifique de l'ADN dérivé des molécules d'ARN de porteur. La bibliothèque cible dérivée de l'échantillon demeure intacte, car les sites de reconnaissance de l'endonucléase sont longs (I-CeuI 27 pb; I-SceI 18 pb) et statistiquement peu susceptible d'être trouvé dans les génomes eucaryotes. Après la dégradation spécifique du transporteur et l'enlèvement des fragments par exclusion de taille, la bibliothèque cible est amplifiée pcR et prête pour le séquençage de la prochaine génération. Selon la quantité d'ARN de départ (1-100 ng), entre 13-18 cycles d'amplification DE PCR devraient être exigés. La quantité finale d'ADN par échantillon varie entre 5-50 ng, ce qui donne assez de matière pour un séquençage très profond. Lors de l'utilisation de seulement 1-2 ng de l'ARN total, les vrais TSS peuvent être détectés; cependant, on s'attend à ce que les bibliothèques soient moins complexes. Enfin, comme SLIC-CAGE est basé sur le protocole nAnT-iCAGE11, il permet le multiplexage de jusqu'à huit échantillons avant le séquençage.
Protocole
1. Préparation du transporteur
-
Préparation de modèles d'ADN pour la transcription in vitro
- Préparer le mix PCR pour chaque modèle PCR en combinant 41 L d'eau, 20 L de tampon HF de 5x, 8 L de dNTP de 2,5 mL, 10 'L de 10 'M amorce avant unique (PCR'GN5-f1, tableau 2; amorces sont dissoutes et diluées dans l'eau) , 10 l de plasmide de modèle de 2 ng/L contenant le gène porteur synthétique et 1 polymérase de Phusion. Mélanger le mélange PCR par pipetting. Un mix maître pour les 10 modèles peut être préparé en même temps (préparer 11 réactions).
- Ajouter 90 OL du mix PCR à 10 oL de chaque amorce inversée de 10 M (PCR-N6-r1-r10, tableau 2). Mélanger par pipetting.
- PCR amplifie les modèles à l'aide du programme suivant : 98 oC pour 60 s, (98 oC pour 10 s, 50 oC pour 30 s, 72 oC pour 30 s) 35 cycles, 72 oC pour 10 min, tenir à 4 oC.
-
Purification de gel des modèles d'ADN amplifiés par PCR
- Préparer un gel d'agarose de 1 % (l'agarose à faible fusion est recommandée).
- Pour diminuer le volume, concentrez les mélanges de réaction PCR de 100 à 20 l volume total à l'aide du concentrateur à vide à basse température moyenne (30 à 40 oC).
- Ajouter 6 ll du colorant de chargement 6x, bien mélanger et charger sur le gel. Exécuter l'électrophoresis pendant 30 min dans un tampon TAE 1x à la tension appropriée pour le réservoir d'électrophoresis utilisé (5 à 10 V/cm). En parallèle, exécuter une échelle d'ADN de 100 bp ou 1 000 bp.
- À l'aide d'un scalpel propre, accisez les tranches de gel contenant le produit PCR cible. Évitez l'excès de gel agarose. Purifier les produits PCR à l'aide d'un kit d'extraction de gel (selon les instructions du fabricant).
REMARQUE: Les ratios A260/A230 d'ADN isolés dans les gels d'agarose sont généralement faibles (0,1 à 0,3). Les produits cibles et les produits latéraux attendus sont indiqués à la figure 2A. Les rendements attendus de 100 réactions PCR de l'Année sont de 1,2 à 3 g. Les réactions peuvent être redimensionnels pour obtenir un rendement plus élevé.
-
Transcription in vitro des molécules porteuses
- Transcrire l'ARN porteur in vitro en utilisant la polymérase d'ARN T7 selon les instructions du fabricant. Configurez 10 à 20 réactions L (le kit recommandé est dans le tableau des matériaux).
- Purifier l'ARN transcrit in vitro à l'aide d'un kit de purification de l'ARN. Configurez la digestion de l'ADN dans la solution en utilisant DNase I en suivant les instructions standard du fabricant, et élichez l'ARN dans 50 L d'eau. Pour augmenter le rendement de l'élution, laisser l'eau dans la colonne pendant 5 min avant la centrifugation.
REMARQUE: Veillez à ne pas dépasser la capacité de fixation maximale des colonnes (dans le kit mentionné dans le Tableau des Matériaux,la capacité est jusqu'à 100 g). Le rendement attendu des modèles PCR 1 à 10 (1 kbp à 200 pb de longueur) est de 25 à 50 g à partir de 10 réactions de transcription in vitro l'Une. Les réactions peuvent être à l'échelle pour obtenir un plus grand stock de molécules porteuses.
-
Plafonnement des molécules d'ARN porteuses transcrites in vitro
- Préparer le mélange de plafonnement en combinant 2 'L de 10x tampon de plafonnement, 1 'L de 10 mM GTP, 1 'L de 2 mM SAM (fraîchement dilué) , et 1 'L d'enzyme de plafonnement Vaccinia par ARN porteur.
- Mélanger jusqu'à 10 g de chaque molécule porteuse dans 15 l de volume total et dénaturer pendant 10 min à 65 oC. Placer immédiatement sur la glace pour empêcher la formation de la structure secondaire.
- Mélanger l'ARN porte-bébé dénaturé avec 5 ll du mélange de plafonnement et couver pendant 1 h à 37 oC.
- Purifez les molécules d'ARN plafonnées à l'aide d'un kit de purification de l'ARN — suivez le protocole de nettoyage du fabricant. L'ARN d'Elute dans 30 l'eau. Pour augmenter le rendement de l'élution, laisser l'eau dans la colonne pendant 5 min avant la centrifugation.
REMARQUE: Mesurer la concentration à l'aide du spectrophotomètre à microvolume. Le ratio A260/A280 prévu est de 2 et 20 000 euros. Notez que pour certains échantillons d'ARN A260/A230 peut être entre 1,3/2. Le rendement prévu lors de l'utilisation de 10 g d'ARN non plafonné est de 9 à 10 g d'ARN plafonné.
- Préparer le mélange du transporteur plafonné et non plafonné en combinant les montants décrits dans le tableau 3. Bien mélanger en faisant glisser le tube et mesurer la concentration à l'aide du spectrophotomètre à microvolume.
REMARQUE: Si une concentration plus élevée du support eurdoiteux est nécessaire pour s'adapter à la réaction de transcription inverse (voir ci-dessous), le mélange de porteur peut être concentré à l'aide du concentrateur de vide à basse température moyenne (30-35 oC) jusqu'à atteindre la concentration finale désirée. Les étapes 2 à 14 sont modifiées à partir du protocole standard nAnT-iCAGE signalé par Murata et coll.11
2. Transcription inversée
- Combiner 1 oL de l'amorce RT (2,5 mM TCT-N6 dissous dans l'eau, pour la séquence voir tableau supplémentaire 1), 10 ng d'ARN total d'intérêt et 4 990 ng de mélange de porte-bébés ( tableau3) en 10 oL de volume total dans une plaque PCR à faible liaison. Mélanger en faisant glisser le tube.
REMARQUE: Si l'ARN de l'échantillon est trop dilué pour la transcription inverse (voir ci-dessous), combinez-le avec la quantité appropriée du porteur, concentrez-vous à l'aide du concentrateur à vide à 9 l de volume total, et ajoutez 1 l de l'amorce RT. L'ajout du earl porteur, pour atteindre 5 g d'ARN au total empêche la perte d'échantillon. - Chauffer le mélange à partir de l'étape 2.1 à 65 oC pendant 5 min, et placer sur la glace immédiatement pour éviter la renaturation.
-
Préparer le mélange de transcription inversée (RT).
- Pour chaque échantillon, combiner 6,1 l d'eau (sans NNase et sans DNase), 7,6 l de tampon de 5 x premier brin, 1,9 L de 0,1 M TNT, 1 L de 10 mM dNTP, 7,6 l de mélange tréhalose/sorbitol (voir recette dans Murata et al.11) et 3,8 'L de la transcriptase inverse recommandée (voir 'c' 6-Table des Matériaux). Bien mélanger en faisant glisser le tube.
- Ajouter 28 L du mélange RT dans le tube PCR avec 10 L d'ARN, de support et d'amorce RT (volume total 38 l). Bien mélanger par pipetting.
REMARQUE: Le mélange est très visqueux en raison de trehalose/sorbitol. Mélanger jusqu'à consistance visiblement homogène. - Incuber dans un cycleur thermique en utilisant le programme suivant : 25 oC pour 30 s, 50 oC pendant 60 min, et tenir à 4 oC.
-
Purification de l'ADNc : hybrides à ARN à l'aide de perles magnétiques SPRI
- Ajouter 68,4 L des perles SPRI recommandées sans RNAse et Sans DNase (voir Tableau des matériaux) à 38 OL du mélange RT (perles au rapport échantillon 1,8:1). Bien mélanger par pipetting et incuber pendant 5 min à température ambiante (RT).
- Séparez les perles sur un support magnétique pendant 5 min. Jetez le supernatant et lavez les perles deux fois avec 200 ll d'éthanol de 70 % (fraîchement préparé).
REMARQUE: L'éthanol est ajouté aux perles sans mélanger et pendant que le tube est sur le support magnétique. L'éthanol ajouté est immédiatement retiré. Il faut prendre soin de ne pas perdre de perles pendant les lavages, car cela peut entraîner une perte d'échantillon. - Pendant que le tube est encore sur le support magnétique, enlever toutes les traces d'éthanol. Les gouttelettes d'éthanol peuvent être enlevées et poussées hors du tube à l'aide d'une pipette P10. Ne laissez pas sécher les perles.
- Ajouter 42 l'eau préchauffée à 37 oC aux perles et éliuter l'échantillon en faisant monter et descendre 60 x.
REMARQUE: Veillez à ne pas provoquer d'écume par tuyauterie, car elle peut causer la perte de perles (c.-à-d.échantillon lié) dans la mousse. - Incuber à 37 oC pendant 5 min sans couvercle pour permettre l'évaporation de traces d'éthanol.
- Séparer les perles sur un support magnétique pendant 5 min et transférer le supernatant dans une nouvelle plaque.
REMARQUE: Essayez de récupérer tous les supernatants pour éviter la perte d'échantillon tout en évitant le report de perles. Utilisez la pipette P10 pour obtenir les dernières gouttelettes d'échantillon.
3. L'oxydation
- Ajouter 2 'L de 1 M NaOAc (pH 4.5) dans la réaction RT purifiée. Mélanger par pipetting, ajouter 2 'L de 250 mM NaIO4 et mélanger à nouveau.
- Incuber sur la glace pendant 45 min. Couvrir la plaque de papier d'aluminium pour éviter la lumière.
- Ajouter 16 l de Tris-HCl (pH 8,5) dans le mélange d'oxydation pour neutraliser le pH.
- Purifez les hybrides oxydés d'ADNc : ARN à l'aide de perles magnétiques SPRI. Ajouter 108 l de perles SPRI à 60 oL du mélange d'oxydation (1,8 :1 perles par rapport à l'échantillon). Répétez la purification telle que décrite dans les étapes 2.6.1-2.6.6. Elute à l'aide de 42 l'eau préchauffée à 37 oC.
REMARQUE: Préparer fraîchement 250 mM NaIO4 en ajoutant 18,7 l d'eau par 1 mg de NaIO4. NaIO4 est sensible à la lumière; par conséquent, gardez la solution dans un tube recouvert de papier d'aluminium ou dans un tube résistant à la lumière.
4. Biotinylation
- Ajouter 4 OL de 1 M NaOAc (pH 6,0) dans le tube contenant l'échantillon oxydé purifié et mélanger par pipetting.
- Ajouter 4 'L de 10 mM de solution de biotine, mélanger par pipetting et incuber pendant 2 h à 23 oC dans un cycleur thermique pour éviter la lumière.
REMARQUE: Préparer la solution de biotine en mélangeant 50 mg de biotine avec 13,5 ml de DMSO. Faire des aliquots à usage unique et congeler à -80 oC. - Purifez les hybrides cDNA:RNA biotinylated utilisant des perles magnétiques SPRI. Ajouter 12 l de 2-propanol et mélanger par pipetting. Ajouter 108 l de perles SPRI (1,8:1 perles au rapport échantillon) et répéter la purification décrite dans les étapes 2.6.1-2.6.6. Elute à l'aide de 42 l'eau préchauffée à 37 oC.
REMARQUE: Le protocole peut être mis en pause ici, et les échantillons congelés à -80 oC.
5. RNase I Ddigestion
- Préparer le RNase je mélange en mélangeant 4,5 L de 10x RNase I tampon avec 0,5 L de RNase I (10 U/ L) par échantillon. Mélanger par pipetting.
- Ajouter 5 ll du mélange à chaque échantillon purifié (45 l au total). Mélanger par pipetting et incuber pendant 30 min à 37 oC.
6. Préparation des perles de Streptavidin
- Pour chaque échantillon, mélanger 30 ll de lisier de perles de streptavidin avec 0,38 l de 20 mg/mL de tRNA. Incuber sur la glace pendant 30 minutes et mélanger toutes les 5 minutes en faisant glisser le tube.
REMARQUE: Resuspendre la boue de perles de streptavidin bien avant pipetting en faisant glisser la bouteille. La solution tRNA doit être préparée selon Murata et al.11 - Séparez les perles sur le support magnétique pendant 2 à 3 min. Retirez le supernatant.
- Laver les perles en reconpendant dans 15 l de tampon A. Séparer les perles sur le support magnétique pendant 2 à 3 minutes et retirer le supernatant. Répétez le lavage et retirez le supernatant.
- Resuspendre les perles dans 105 l de tampon A et ajouter 0,19 l de 20 mg/mL de tRNA. Bien mélanger par pipetting.
REMARQUE: Les perles doivent être préparées fraîchement avant l'utilisation. Commencer la préparation des perles pendant la digestion RNase I. Pour plusieurs échantillons préparer les perles ensemble dans un seul tube.
7. Cap-piégeage
-
Liaison d'échantillon
- Ajouter 105 l de perles de streptavidin es préparées à 45 l'échantillon traité Par RNase I. Bien mélanger en pipetilage et incuber à 37 oC pendant 30 min. Mélanger en pipetilage toutes les 10 min.
- Séparez les perles sur le support magnétique pendant 2 à 3 min. Retirez le supernatant.
-
Léve zonale
- Ajouter 150 l de tampon de lavage A et resuspendre les perles par pipetting. Séparez les perles sur le support magnétique pendant 2 à 3 min et retirez le supernatant.
- Ajouter 150 l de la mémoire tampon de lavage B et resuspendre les perles par pipetting. Séparez les perles sur le support magnétique pendant 2 à 3 min et retirez le supernatant.
- Ajouter 150 l de la mémoire tampon de lavage C et resuspendre les perles par pipetting. Séparez les perles sur le support magnétique pendant 2 à 3 min et retirez le supernatant.
REMARQUE: Les tampons B et C doivent être préchauffés à 37 oC. Les recettes pour les tampons de lavage A, B et C sont décrites dans Murata et al.11
-
cDNA communiqué
- Préparer 1x RNase I tampon en mélangeant 58,5 L d'eau avec 6,5 L de 10x RNase I tampon.
- Resuspendre les perles en 35 oL de 1x tampon RNase I. Incuber à 95 oC pendant 5 min et transférer directement sur la glace pendant 2 min pour éviter la réassociation de l'ADNc. Tenez les couvercles pendant le transfert sur la glace, car ils peuvent sauter-off en raison de l'accumulation de pression.
- Séparer les perles pendant 2 à 3 min sur un support magnétique et transférer le supernatant dans une nouvelle plaque.
- Resuspendre les perles en 30 oL de 1x tampon RNase I. Séparez les perles sur le support magnétique pendant 2 à 3 min et transférez le supernatant au supernatant précédemment recueilli (le volume total de l'ADNc élué devrait être d'environ 65 l).
8. Enlèvement de l'ARN par RNase H et RNase I Digestion
- Par échantillon, combiner 2,4 l d'eau, 0,5 l de 10x rNase I tampon, 0,1 l de RNase H, et 2 'L de RNase I.
- Ajouter 5 ll du mélange aux 65 l l de l'échantillon d'ADNc libéré et mélanger par pipetting. Incuber à 37 oC pendant 15 min et tenir à 4 oC.
- Purifez l'ADNc du mélange de digestion RNase à l'aide de perles magnétiques SPRI. Ajouter 126 l de perles SPRI à 70 l de réaction de dégradation et mélanger par pipetting. Suivez les étapes de purification décrites pour la purification des perles SPRI en 2.6.1-2.6.6. Elute à l'aide de 42 l'eau préchauffée à 37 oC comme décrit.
- Préparer RNase je mix en combinant 4,5 L de 10x RNase I tampon et 0,5 L de RNase I.
- Ajouter 5 ll du mélange RNase aux 40 oL de l'échantillon purifié d'ADNc. Mélanger par pipetting et incuber à 37 oC pendant 30 min. Tenir à 4 oC.
- Purifie l'échantillon à l'aide de perles magnétiques SPRI. Ajouter 81 l de perles SPRI à 45 l de réaction de dégradation et mélanger par pipetting. Suivez les étapes de purification décrites pour la purification des perles SPRI en 2.6.1-2.6.6. Elute à l'aide de 42 L d'eau tel que décrit.
9. Ligation de 5' Linker
- Concentrez l'échantillon purifié d'adin à 4 'L à l'aide du concentrateur à vide. Maintenir la température à 30-35 oC. Testez le volume à l'aide d'une pipette. Si l'échantillon a séché à l'exhaustivité, dissoudre en ajoutant 4 L d'eau.
REMARQUE: Il est préférable d'éviter le séchage à l'exhaustivité pour éviter la perte de l'échantillon. - Incuber l'échantillon concentré à 95 oC pendant 5 min et le placer immédiatement sur la glace pendant 2 min pour éviter la renaturation. Maintenez les couvercles pendant le transfert des tubes car les couvercles peuvent éclater en raison de l'accumulation de pression.
- Incuber 4 'L du lien de 2,5 M 5' à 55 oC pendant 5 min et placer immédiatement sur la glace pendant 2 min pour éviter la renaturation.
- Mélanger 4 lL du lien de 2,5 M 5' avec 4 oL de l'échantillon.
REMARQUE: Le lien 5' doit être préparé selon le tableau supplémentaire 2, le tableau supplémentaire 3, le tableau supplémentaire 4, et le tableau supplémentaire 5. Diluer le lien de 10 M 5' à une concentration de 2,5 M m en utilisant 100 mM NaCl avant l'utilisation. - Ajouter 16 l de la ligature prémix (voir Tableau des matériaux) au lien mixte de 5' et à l'échantillon et bien mélanger par pipetting. Incuber à 16 oC pendant 16 h.
- Purifez le mélange de ligature à l'aide de perles magnétiques SPRI. Ajouter 43,2 L de perles SPRI et suivre les étapes 2.6.1-2.6.6. Elute tel que décrit à l'aide de 42 l d'eau préchauffée à 37 oC.
- Répétez la purification effectuée à l'étape 9.6 en ajoutant 72 l de perles SPRI au supernatant transféré (1,8:1 perles par rapport à l'échantillon).
REMARQUE : Les linkers de 5' contiennent des codes-barres qui permettent la mise en commun d'un jusqu'à huit échantillons avant le séquençage (huit codes-barres de trinuccléotide sont disponibles, comme décrit dans Murata et al.11 et tableau supplémentaire 1).
10. Ligation de 3' Linker
- Concentrez l'échantillon purifié à 4 L à l'aide du concentrateur à vide tel que décrit à l'étape 9.1.
- Incuber l'échantillon concentré à 95 oC pendant 5 min et le placer immédiatement sur la glace pendant 2 min pour éviter la renaturation. Maintenez les couvercles pendant le transfert des tubes, car les couvercles peuvent s'éteindre en raison de l'accumulation de pression.
- Incuber 4 'L du lien de 2,5 M 3' à 65 oC pendant 5 min et placer immédiatement sur la glace pendant 2 min pour éviter la renaturation.
- Ajouter 4 ll du lien de 2,5 M 3' au 4 oL de l'échantillon concentré.
- Ajouter 16 l de prémélange de ligature et bien mélanger par pipetting. Incuber à 16 oC pendant 16 h.
- Purifez le mélange de ligature à l'aide de perles magnétiques SPRI. Ajouter 43,2 L de perles SPRI et suivre les étapes 2.6.1-2.6.6. Elute tel que décrit à l'aide de 42 l d'eau préchauffée à 37 oC.
REMARQUE: Le lien 3' doit être préparé selon les tableaux complémentaires 6 et le tableau supplémentaire 7. Diluer le lien de 10 M 3' à une concentration de 2,5 M à l'aide de 100 mM NaCl.
11. Déphosphorylation
- Préparer le mix SAP en combinant 4 ll d'eau, 5 l de tampon SAP 10x et 1 an l d'enzyme SAP.
- Ajouter 10 l de mélange SAP à l'échantillon ligat purifié (volume total de 50 l) et incuber dans le thermocycleur en utilisant le programme suivant : 37 oC pendant 30 min, 65 oC pendant 15 min, et tenir à 4 oC.
12. Dégradation de 3' Linker Upper Strand Using Uracil Specific Excision Enzyme
- Ajouter 2 l'enzyme d'excision spécifique à l'uracil (voir Tableau des matériaux)à l'échantillon déphosphorylé, mélanger par pipetting et incuber dans le thermocycleur en utilisant le programme suivant : 37 oC pendant 30 min, 95 oC pendant 5 min, et placer immédiatement sur la glace pendant 2 min pour empêcher la réannealing du brin supérieur fragmenté.
- Purifiez le mélange de réaction en ajoutant 93,6 l de perles magnétiques SPRI au mélange de 52 l et mélangez bien par pipetting. Répéter les étapes de purification 2.6.1-2.6.6. Elute avec 42 l'eau préchauffée à 37 oC comme décrit.
13. Deuxième strand Synthèse
- Préparer le deuxième mélange de synthèse des brins (les volumes sont exprimés par échantillon) en combinant 5 L de tampon de réaction à la polymérase d'ADN 10x, 2 l d'eau, 1 L de 10 mL de dNTP, 1 L de 50 M nAnT-iCAGE deuxième brin (la séquence est dans le tableau supplémentaire 1) et 1 l de D Polymérase exonuccléenne-déficiente NA (voir polymérase recommandée dans tableau des matériaux).
- Ajouter 10 ll du mélange à l'échantillon purifié et bien mélanger par pipetting (volume total est de 50 l). Incuber dans le cycleur thermique en utilisant le programme suivant : 95 oC pendant 5 min, 55 oC pour 5 min, 72 oC pendant 30 min, et tenir à 4 oC.
14. Dégradation de l'amorce de synthèse de deuxième brin utilisant l'exonuclease I
- Ajouter 1 l d'Exonuclease I au mélange de synthèse du deuxième brin. Bien mélanger par pipetting et incuber à 37 oC pendant 30 min, puis tenir à 4 oC.
- Purifez l'ADN double brin en ajoutant 91,8 L de perles magnétiques SPRI à 51 l'échantillon traité Exonuclease I. Répéter les étapes de purification décrites en 2.6.1-2.6.6. et s'élinit avec 42 l'eau préchauffée à 37 oC comme décrit.
- Concentrez l'échantillon à l'aide du concentrateur à vide à 15 L, tel que décrit à l'étape 9.1.
15. Contrôle de la qualité et de la quantité
- Utilisez 1 L des échantillons concentrés et exécutez une puce d'ADN à haute sensibilité sur un analyseur de qualité d'ADN. Le profil/quantité attendu est présenté à la figure 3.
16. Première série de dégradation des transporteurs
- Préparer le mélange de dégradation en combinant 2 l d'eau, 2 l de tampon enzymatique de restriction 10x, 1 l d'I-SceI, et 1 l d'I-CeuI.
- Ajouter 6 ll du mélange de dégradation à 14 ll de l'échantillon concentré et mélanger par pipetting. Incuber à 37 oC pendant 3 h, puis 20 min de désactivation à 65 oC et tenir à 4 oC.
- Purifie le mélange de dégradation à l'aide de perles magnétiques SPRI. Ajouter 5 ll d'eau pour augmenter le volume du mélange de dégradation et ajouter 45 l de perles SPRI (1,8:1 perles par rapport à l'échantillon). Répéter la purification telle que décrite dans les étapes 2.6.1-2.6.6. et s'élindre avec 42 l'eau préchauffée à 37 oC.
- Concentrez l'échantillon éluté de 42 à 20 L du volume total tel que décrit à l'étape 9.1.
17. Contrôle du niveau de dégradation et détermination du nombre de cycles d'amplification PCR
- Préparer le mix qPCR pour amplifier des bibliothèques entières (mix adaptateur). Combiner 3,8 l d'eau, 5 'L de prémélange qPCR (2x), 0,1 'L'amorce adaptor'f1 (5'-AATGATACGGCGACCACCACCGA-3'), et 0.1 'L'de 10 'M adaptor'r1 amorce (5'-CAAGCAGAAGACGGGGCATACGA-3') pour chaque échantillon (voir tableau des matériaux pour le prémix qPCR recommandé).
- Mélanger 9 ll de mélange d'adaptateur qPCR avec 1 l d'échantillon à partir de l'étape 16.4 et bien mélanger par pipetting.
- Préparer le mélange qPCR pour amplifier l'ADN dérivé du transporteur (mélange de porteur). Combiner 3,8 L d'eau, 5 l de prémélange qPCR (2x), 0,1 l de 10 amorce de porte-eau (5'-GCGGCAGCGTTCGCTATATataAC-3'), et 0,1 'L'amorce adaptor'r1 pour chaque échantillon
- Mélanger 9 ll de mélange de porte-porteurs qPCR avec 1 l l de l'échantillon à partir de l'étape 16.4 et bien mélanger par pipetting.
- Définir le programme qPCR : 95 oC pour 3 min (95 oC pour 20 s, 60 oC pour 20 s, 72 oC pour 2 min) répété 40x, suivi d'une courbe de dénaturation spécifique aux instruments (65 à 95 oC), et se maintenir à 4 oC.
REMARQUE: Préparer un contrôle négatif en remplaçant l'échantillon par de l'eau.
18. Amplification pcR de la bibliothèque cible
- Préparer le mix d'amplification PCR en combinant 6 ll d'eau, 0,5 L d'amorce adaptateur f1 de 10 M, 0,5 L d'amorce adaptateur de 10 M et 25 l de prémix PCR (2x). Mélanger par pipetting (voir Tableau des matériaux pour le prémélange PCR recommandé).
- Ajouter 32 L du mix PCR à 18 oL de l'échantillon à partir de l'étape 16.4. Bien mélanger par pipetting.
- Définir l'amplification du PCR : 95 oC pour 3 min, (98 oC pour 20 s, 60 oC pour 15 s, 72 oC pour 2 min) 12-18 cycles, 72 oC pour 2 min et tenir à 4 oC.
REMARQUE: Le nombre exact de cycles PCR est déterminé par les résultats qPCR et correspond à la valeur Ct obtenue avec le mix d'apprêt adaptateur (le nombre de cycles PCR est égal à la valeur Ct). - Purifez l'échantillon amplifié en ajoutant 90 l de perles magnétiques SPRI à 50 oL de l'échantillon amplifié et mélangez soigneusement par pipetting. Répétez les étapes de purification décrites dans les étapes 2.6.1-2.6.6. et d'éliner l'échantillon à l'aide de 42 l'eau telle que décrite.
19. Deuxième ronde de dégradation des transporteurs
- Répétez les étapes 16.1-16.3.
- Purifie le mélange de dégradation à l'aide de perles magnétiques SPRI. Ajouter 10 l'eau à l'échantillon pour augmenter le volume et mélanger avec 30 l de perles SPRI (1:1 perles au rapport échantillon). Répéter la purification telle que décrite dans les étapes 2.6.1-2.6.6. et s'élinit avec 42 L d'eau préchauffée à 37 oC comme décrit.
- Concentrez l'échantillon éluté de 42 à 30 L de volume total.
20. Sélection de taille de bibliothèque
- Mélanger 24 l de perles magnétiques SPRI avec 30 oL de l'échantillon à partir de l'étape 19.3. (0,8:1 perles par rapport à l'échantillon). Répétez les étapes de purification décrites dans les étapes 2.6.1-2.6.6. et d'éliner l'échantillon dans 42 L d'eau tel que décrit.
- Concentrez l'échantillon à environ 14 L comme décrit à l'étape 9.1.
21. Contrôle de la qualité
-
Évaluation de la taille-distribution
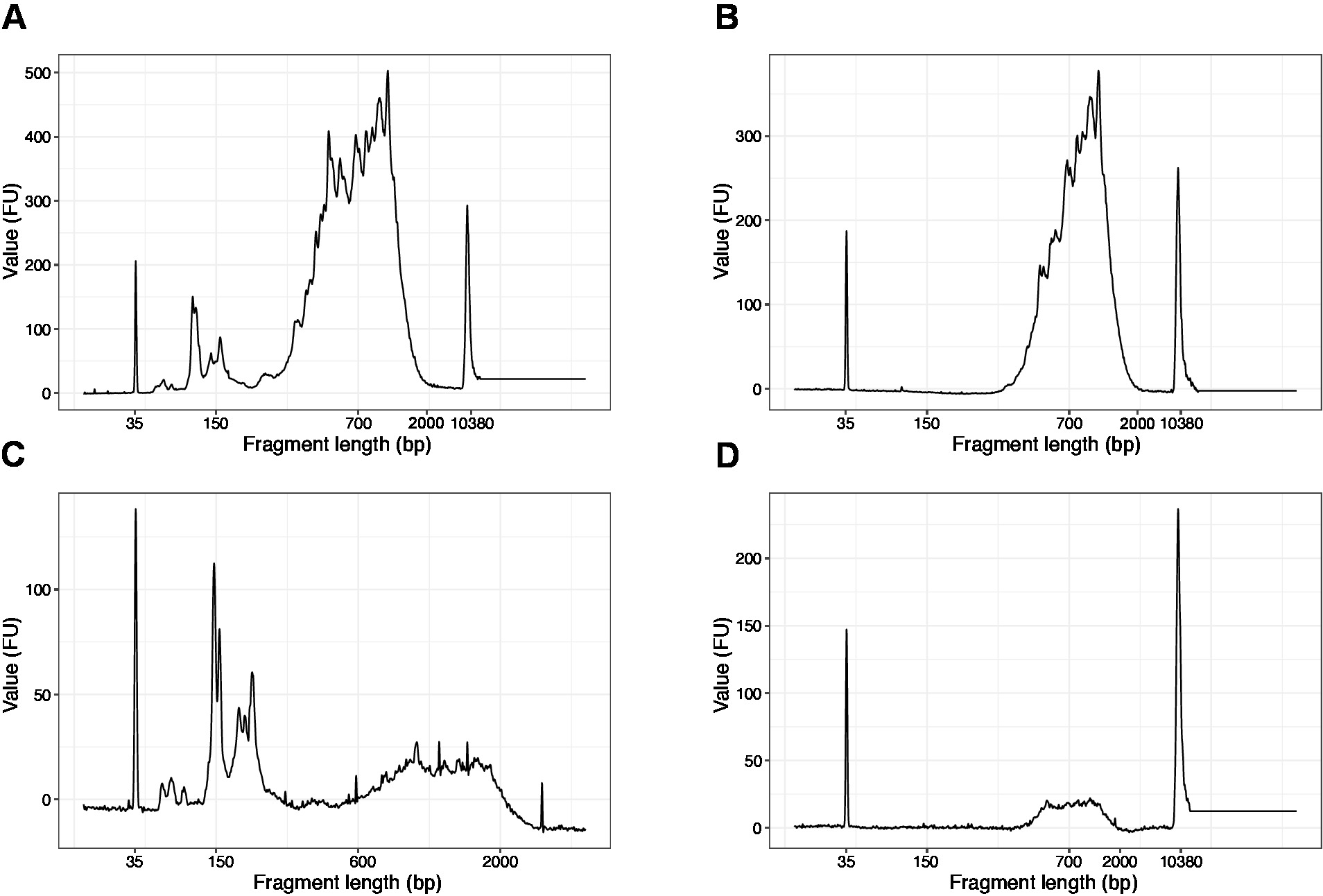
- Exécuter 1 L de l'échantillon sur la puce d'ADN à haute sensibilité. Les résultats attendus sont présentés à la figure 4.
REMARQUE: Si des fragments de moins de 200 pb sont visibles (voir l'exemple de la figure 4A,C),la sélection de la taille (étapes 20,1 à 20,2) doit être répétée jusqu'à ce que les fragments courts soient enlevés (Figure 4B,D). Habituellement, un tour supplémentaire de sélection de taille est suffisant. Si la quantité de fragments courts est grave (comme à la figure 4C),le rapport perles-échantillon devrait être ramené à 0,6:1.
- Exécuter 1 L de l'échantillon sur la puce d'ADN à haute sensibilité. Les résultats attendus sont présentés à la figure 4.
-
Contrôle de la qualité de la dégradation du transporteur
- Répétez les étapes 17.1-17.5.
REMARQUE: Selon la concentration des bibliothèques estimée dans la course de puces d'ADN HS (analyse de région), les échantillons doivent être dilués avant qPCR. Utilisez 0,5 L de l'échantillon pour éviter la perte de l'échantillon et diluer 100 à 500x dans l'eau (diluer à 1-20 pg/L concentration finale). La différence prévue entre les valeurs Ct obtenues avec l'adaptateur et le mix transporteur est de 5 à 10.
- Répétez les étapes 17.1-17.5.
-
Quantification des bibliothèques
- Préparer la dilution de travail de la norme d'ADN lambda en mélangeant 20 'L de 100 mg/mL lambda norme d'ADN avec 980 'L de 1x TE (préparer en diluant 20x TE fourni dans le kit de quantification de l'ADN). La dilution de l'ADN lambda peut être stockée à -20 oC.
- Préparer les dilutions sérielles standard de l'ADN lambda en mélangeant la norme lambda diluée et 1x TE selon le tableau supplémentaire 8.
REMARQUE: Pour une plus grande précision, il est recommandé d'ajouter 100 L de tampon 1x TE à tous les tubes et d'enlever le volume 1x TE au besoin par volume de l'lambda dilué à ajouter. N'utilisez pas plus d'un l de la bibliothèque; l'utilisation de 384 plaques de puits pour cette mesure est recommandée.
Résultats
Ce rapport décrit le protocole complet SLIC-CAGE pour l'obtention de bibliothèques prêtes à sequencing à partir de nanogrammes de matériel d'ARN total de départ (figure 1). Pour obtenir le mélange synthétique de porteur d'ARN, d'abord, les modèles de support de PCR doivent être préparés et gel-purifiés pour éliminer des produits latéraux de PCR (figure 2A). Chaque modèle PCR (dix au total) est produit à l'aide d'un avant commun, mais d'une amorce inverse différente (tableau 2), conduisant à des longueurs différentes du modèle PCR pour permettre la variabilité de la taille des porteurs d'ARN synthétiques. Une fois purifiés, les modèles PCR sont utilisés pour la transcription in vitro des molécules porteuses. On s'attend à ce qu'un seul produit porteur d'ARN soit prévu si les modèles sont purifiés en gel (voir l'analyse représentative du gel à la figure 2B). La préparation du transporteur peut être mise à niveau en fonction des besoins, et lorsqu'elle est préparée, mélangée et congelée à -80 oC pour une utilisation future.
En utilisant la quantité minimale recommandée d'ARN total d'échantillon (10 ng) combinée avec 16-18 cycles d'amplification de PCR, les bibliothèques sLIC-CAGE de haute complexité peuvent être réalisées. Le nombre de cycles PCR requis pour amplifier la bibliothèque finale dépend fortement de la quantité d'ARN d'entrée totale utilisée (le nombre prévu de cycles est présenté dans le tableau 4).
Après le premier cycle de dégradation, dans les résultats qPCR (étape 17), la différence attendue entre les valeurs Ct obtenues à l'aide de l'adaptateur f1 ou de l'amorce carrier-f1 est de 1-2, avec des valeurs Ct obtenues avec adaptor-f1 plus bas qu'avec le transporteur.f1.
La distribution des longueurs de fragment dans la bibliothèque finale est comprise entre 200 et 2 000 pb avec une taille moyenne de 700 à 900 pb (d'après l'analyse de la région à l'aide du logiciel Bioanalyzer, Figure 4B,D). Les fragments plus courts, tels que présentés à la figure 4A,C, doivent être enlevés par d'autres séries d'exclusion de taille (étapes 20-21). Ces courts fragments sont des artefacts d'amplification PCR et non la bibliothèque cible. Notez que les fragments plus courts se regroupent mieux sur les cellules de flux de séquençage et peuvent causer des problèmes de séquençage.
La quantité prévue de matériel de bibliothèque obtenu par échantillon se situe entre 5 et 50 ng. Des quantités significativement plus faibles sont indicatives de la perte d'échantillon pendant le protocole. Si la faible quantité obtenue est suffisante pour le séquençage (2-3 ng des bibliothèques mises en commun est nécessaire), les bibliothèques peuvent être de moindre complexité (voir ci-dessous).
Selon la machine de séquençage, la quantité de la bibliothèque chargée sur la cellule d'écoulement peut devoir être optimisée. À l'aide d'un Illumina HiSeq 2500, le chargement 8-12 pM SLIC-CAGE bibliothèques donne en moyenne 150-200 millions de lectures, avec 'gt;80% des lectures passant score de qualité Q30 comme seuil.
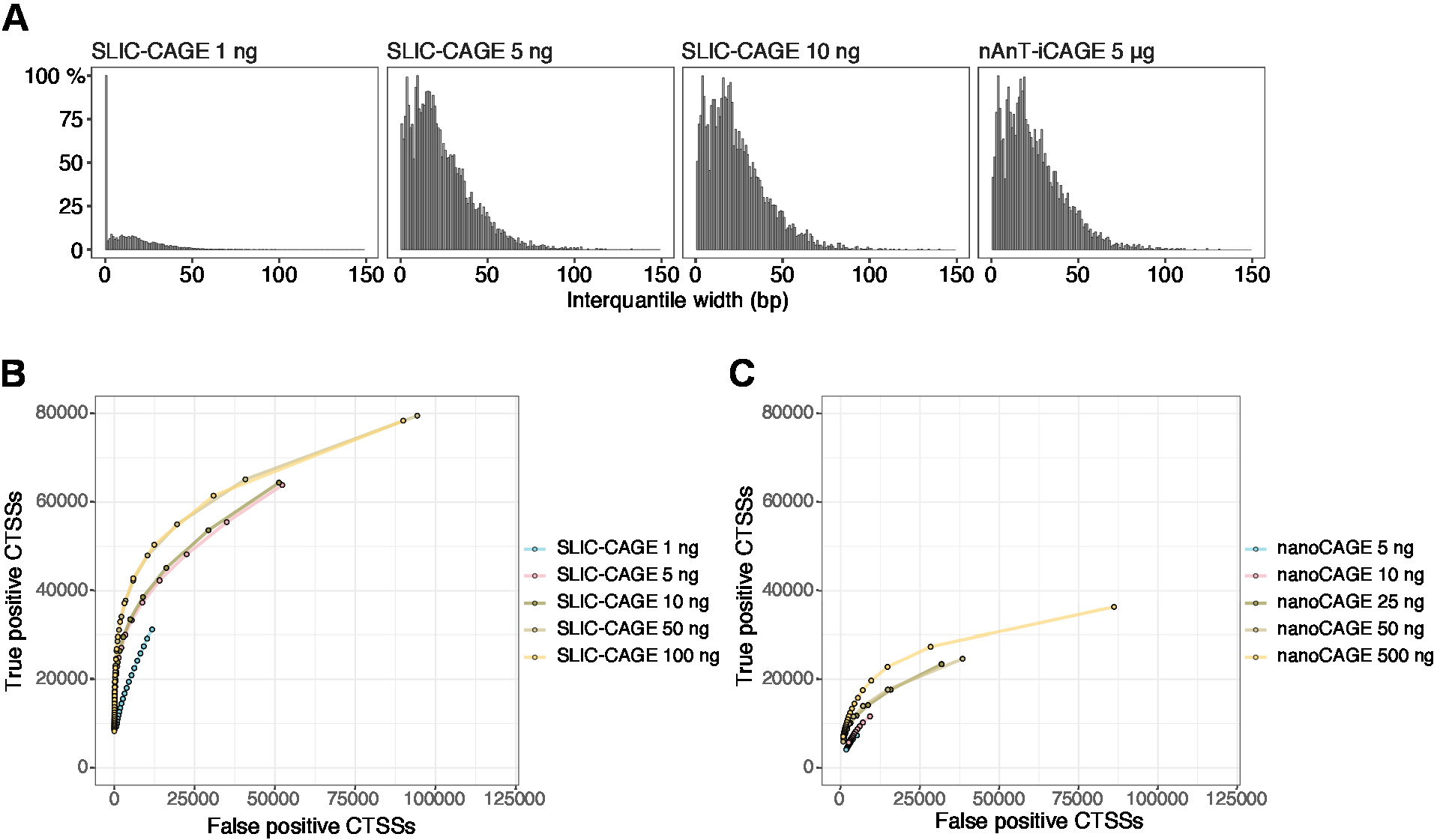
Les lectures obtenues sont ensuite cartographiées au génome de référence [pour 50 bp lit, Bowtie212 peut être utilisé avec des paramètres par défaut qui permettent zéro décalage par séquence de semences (22 bp)]. L'efficacité de la cartographie prévue dépend de la quantité totale d'entrée d'ARN et est présentée au tableau 5. Les lectures cartographiées uniques peuvent ensuite être chargées dans l'environnement informatique graphique et statistiqueR 13 et traitées à l'aide de CAGEr (paquet Bioconducteur14). La vignette de paquet est facile à suivre et explique le flux de travail et le traitement des données cartographiées en détail. Un contrôle visuel facile de la complexité de la bibliothèque est la distribution de la largeur des promoteurs, car les bibliothèques à faible complexité auront des promoteurs artificiellement étroits (Figure 5A, SLIC-CAGE bibliothèque dérivée de 1 ng de l'ARN total, pour plus de détails voir les précédents publication10). Cependant, même les bibliothèques SLIC-CAGE à faible complexité permettent d'identifier les véritables CTSS, avec une plus grande précision que les méthodes alternatives pour la cartographie TSS à faible/moyenne entrée (Figure 5B, C).

Figure 1 : Étapes du protocole SLIC-CAGE. L'ARN d'échantillon est mélangé avec le mélange de porteur d'ARN pour réaliser 5 'g de matériel total d'ARN. cDNA est synthétisé par transcription inversée et le bouchon est oxydé à l'aide de paroi de sodium. L'oxydation permet l'attachement de la biotine au bouchon à l'aide de biotin hydrazide. La biotine s'attache à l'extrémité de l'ARNm 3, car elle est également oxydée à l'aide de la diététité de sodium. Pour éliminer la biotine de l'ARNm : les hybrides d'ADNc avec l'ADNc incomplètement synthétisé et des extrémités de 3 degrés de l'ARNm, les échantillons sont traités avec RNase I. cDNA qui a atteint l'extrémité de 5 'de l'ARNm est alors choisi par purification d'affinité sur les perles magnétiques de streptavidin ( piégeage du bouchon). Après la libération de l'ADNc, 5 et 3-linkers sont ligated. Les molécules de bibliothèque qui proviennent du transporteur sont dégradées à l'aide d'endonucabals I-SceI et I-CeuI et les fragments sont enlevés à l'aide de perles magnétiques SPRI. La bibliothèque est ensuite amplifiée PCR. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
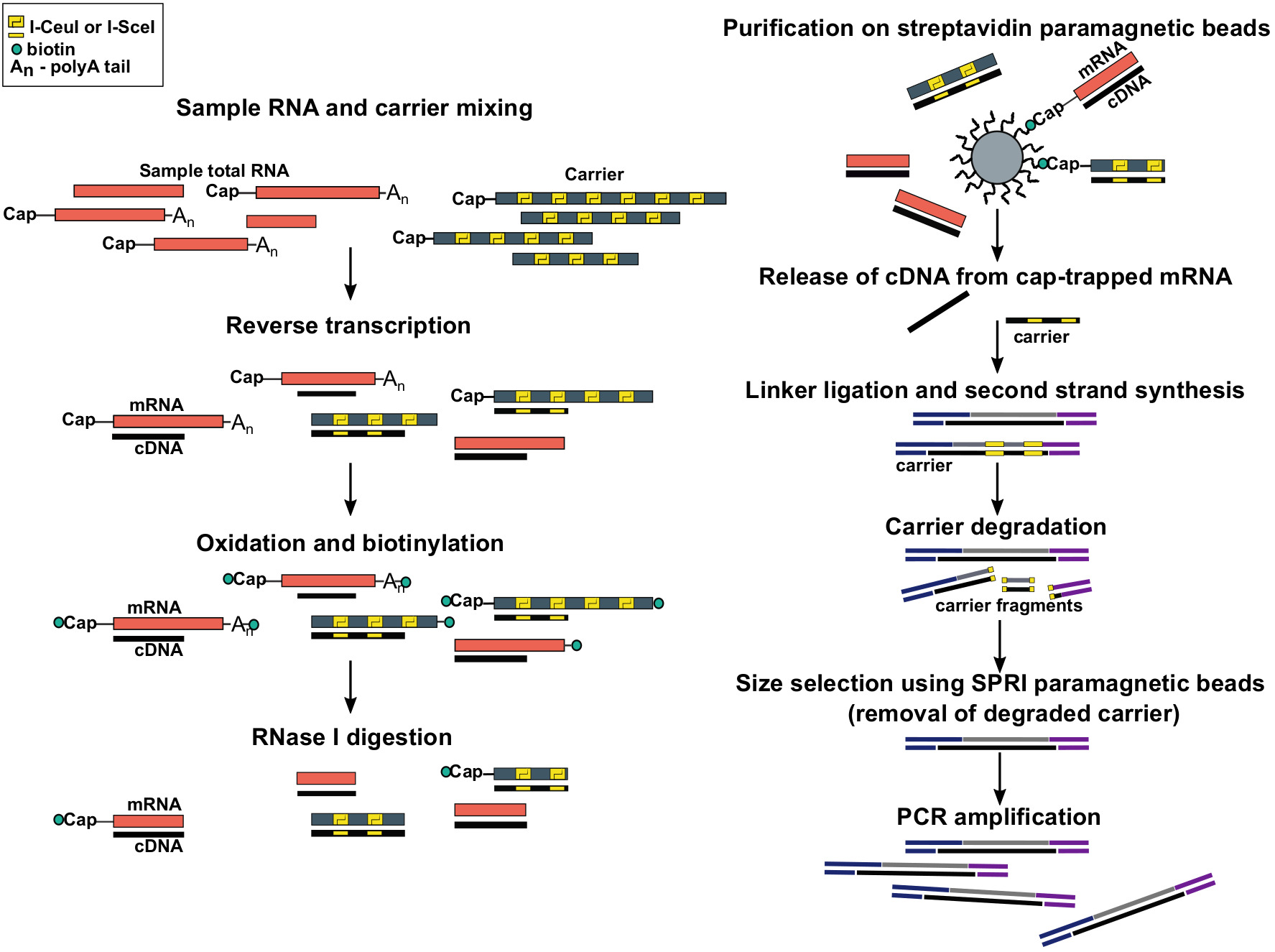{kind=link}

Figure 2 : Analyse représentative du gel des modèles DE PCR du transporteur et des transcriptions in vitro du transporteur. (A) Modèles PCR de porteur avant la purification de gel : le premier puits contient le marqueur de 1 kbp, suivi des modèles de PCR de porteur 1, 1-10. (B) Transcriptions in vitro du transporteur : le premier puits contient le marqueur de 1 kbp, suivi des transcriptions du transporteur 1-10. Les relevés de notes du transporteur ont été dénaturés par chauffage pendant 5 min à 95 oC avant le chargement. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Trace représentative de la qualité de l'ADN (puce d'ADN à haute sensibilité) de SLIC-CAGE avant le premier tour de dégradation du porteur. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 4 : La qualité de l'ADN représentatif (puce d'ADN à haute sensibilité) traces des bibliothèques SLIC-CAGE après l'amplification du PCR. (A) Bibliothèque SLIC-CAGE qui nécessite une sélection de taille supplémentaire pour l'enlèvement de fragments courts. (B) Bibliothèque SLIC-CAGE après sélection de taille en utilisant 0,6x perles SPRI par rapport à l'échantillon. (C) Bibliothèque SLIC-CAGE de montant de sortie inférieur qui nécessite la sélection de taille pour l'enlèvement du fragment court. (D) Bibliothèque SLIC-CAGE de plus faible quantité de sortie après sélection de taille en utilisant 0.6:1 perles SPRI au rapport d'échantillon. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 5 : Validation des bibliothèques SLIC-CAGE. (A) Distribution des largeurs interquantiles des amas d'étiquettes dans les bibliothèques SLIC-CAGE préparées à partir de 1, 5 ou 10 ng s'agitdent de l'ARN total de S. cerevisiae, et dans la bibliothèque nAnT-iCAGE préparée à partir de 5 g d'ARN total de S. cerevisiae. Une grande quantité de clusters d'étiquettes étroits dans la bibliothèque SLIC-CAGE de 1 ng indique sa faible complexité. (B) Courbes ROC pour l'identification CTSS dans les bibliothèques S. cerevisiae SLIC-CAGE. Tous les CTSS S. cerevisiae nAnT-iCAGE ont été utilisés comme véritable ensemble. (C) courbes ROC pour l'identification CTSS dans les bibliothèques S. cerevisiae nanoCAGE. Tous les CTSS S. cerevisiae nAnT-iCAGE ont été utilisés comme véritable ensemble. La comparaison des courbes de ROC montre que SLIC-CAGE surpasse fortement nanoCAGE dans l'identification CTSS. Les données d'ArrayExpress E-MTAB-6519 ont été utilisées. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}
Tableau 1 : Séquence du gène synthétique porteur. Les sites I-SceI sont audacieux et italiques en violet, et les sites de reconnaissance I-CeuI sont verts. Veuillez cliquer ici pour télécharger ce fichier.
| transporteur | amorce inverse 5'-3' | Longueur du produit PCR / bp | |
| 1 Fois | PCR-N6-r1: NNNNNNNNCTACGTGTCGCAGACGAATT | en 1034, états-unis | |
| 2 (en) | PCR-N6-r2: NNNNNNTATCCAGATCGTTGAGCTGC | 966 Annonces | |
| 3 (en) | PCR-N6-r3: NNNNNNNCACTGCGCGATCTCTTTACG | 889 Annonces | |
| 4 ( en plus) | PCR-N6-r4: NNNNNNGCCGTCGATAACTTGTTCGT | 821 Annonces | |
| 5 Annonces | PCR-N6-r5: NNNNNNAGTTGACCGCAGAAGTCTTC | 744 Annonces | |
| 6 Annonces | PCR-N6-r6: NNNNNNGTGAAGAATTTCTGTTCCCA | 676 Annonces | |
| 7 Annonces | PCR-N6-r7: NNNNNNNCTCGCCTCCAGTCATAAC | 599 Annonces | |
| 8 Annonces | PCR-N6-r8: NNNNNNTATACGCGATGTTGTCGTCGTAC | 531 Annonces | |
| 9 (en) | PCR-N6-r9: NNNNNNACCGCCGCGCCTTCCCGCAGG | 454 Annonces | |
| 10 Ans et plus | PCR-N6-r10: NNNNNNCAGGACGTTTTTGCCCAGCA | 386 Annonces | |
| L'amorce avant est la même pour tous les modèles de support. Souligné est la séquence de promoteur T7. PCR-GN5-f1: TAATACGACTCACTATAGNNNNNCAGCGTTCGCTA | |||
Tableau 2 : Amorces pour l'amplification du modèle de transporteur. L'apprêt avant est le même pour tous les modèles de support. Souligné est la séquence de promoteur T7. PCR-GN5-f1: TAATACGACTCACTATAGNNNNNNNCAGCGTTCGCTA. À l'aide d'amorces inverses différentes, des modèles PCR et donc des ARN porteuses de longueur différente sont produits.
| transporteur | longueur | non plafonné/g | plafonné/g |
| 1 Fois | en 1034, états-unis | 3,96 | 0,45 |
| 2 (en) | 966 Annonces | 8,36 | 0,95 |
| 3 (en) | 889 Annonces | 4,4 | 0,5 |
| 4 ( en plus) | 821 Annonces | 6,6 | 0,75 |
| 5 Annonces | 744 Annonces | 4,4 | 0,5 |
| 6 Annonces | 676 Annonces | 3,08 | 0,35 |
| 7 Annonces | 599 Annonces | 4,4 | 0,5 |
| 8 Annonces | 531 Annonces | 3,96 | 0,45 |
| 9 (en) | 454 Annonces | 2,64 | 0,3 |
| 10 Ans et plus | 386 Annonces | 2,2 | 0,25 |
Tableau 3 : Mélange de porteurs d'ARN. Au total, 49 g du mélange de porte-avions 0,3 à 1 kbp : non plafonnés à 44 g, plafonnés à 5 g.
| Entrée totale d'ARN /ng | Cycles PCR |
| 1 ng | 18 ans, états-unis qui |
| 2 ng | 17 Annonces |
| 5 ng | 16 Annonces |
| 10 ng | 15 à 16 ans |
| 25 ng | 14 à 15 ans |
| 50 ng | 13 à 15 ans |
| 100 ng | 12 à 14 ans |
Tableau 4 : Nombre prévu de cycles de PCR en dépendance de l'apport total d'ARN de l'échantillon. Le nombre approximatif de cycles est basé sur des expériences effectuées utilisant Saccharomyces cerevisiae, Drosophila melanogaster, et Mus musculus ARN total.
| Entrée totale d'ARN/ng | % de l'ensemble cartographié | % cartographié de façon unique | % transporteur |
| 1 ng | 30 Ans, états-unis ( | 20 à 30 ans | 30 Ans, états-unis ( |
| 2 ng | 60 Annonces | 20 à 50 ans | 10 Ans et plus |
| 5 ng | 60 à 70 ans | 40 à 60 ans | 5 à 10 ans |
| 10 ng | 60 à 70 ans | 40 à 60 ans | 5 à 10 ans |
| 25 ng | 65 à 80 ans | 40 à 70 ans | 0 à 5 |
| 50 ng | 65 à 80 ans | 40 à 70 ans | 0 à 3 |
| 100 ng | 70 à 85 ans | 40 à 70 ans | 0 à 2 |
Tableau 5 : Efficacité de cartographie prévue et dépendance de la quantité totale d'entrée d'ARN. Des nombres approximatifs sont présentés et basés sur des expériences effectuées à l'aide de Saccharomyces cerevisiae et Mus musculus ARN total.
Discussion
Pour réussir les préparations de la bibliothèque SLIC-CAGE, il est essentiel d'utiliser des pointes et des tubes à faible liaison pour prévenir la perte d'échantillons due à l'adsorption de l'échantillon. Dans toutes les étapes impliquant la récupération du supernatant, il est recommandé de récupérer l'ensemble du volume de l'échantillon. Comme le protocole comporte plusieurs étapes, la perte continue d'échantillons entraînera des échecs dans les bibliothèques.
Si CAGE (nAnT-iCAGE) n'a pas été effectué régulièrement, il est préférable de tester SLIC-CAGE avec différentes quantités d'intrants (10 ng, 20 ng, 50 ng, 100 ng, 200 ng) du même échantillon total d'ARN et de comparer avec les bibliothèques nAnT-iCAGE qui sont préparées à l'aide de 5 g d'ARN total. Si la bibliothèque nAnT-iCAGE échoue (moins de 0,5-1 ng de la bibliothèque d'ADN obtenue par échantillon), SLIC-CAGE est peu susceptible de fonctionner, et la perte d'échantillon doit être réduite au minimum.
Une étape cruciale pour s'assurer que les bibliothèques de haute qualité dépourvues d'ARN ou d'ARNr dégradé non plafonné sont le piégeage du bouchon décrit à la section 7. Il est très important que les perles de streptavidin soient complètement remises en suspension dans les tampons de lavage et que les tampons de lavage soient enlevés avant de continuer à l'étape suivante de lavage ou à l'élution de l'ADC.
Si les résultats du qPCR après le premier tour de dégradation du transporteur ne montrent aucune différence entre l'utilisation d'adaptor-f1 et d'amorces de transporteur-f1, la poursuite du protocole est toujours recommandée. Si après la deuxième série de dégradation du transporteur, la différence dans les valeurs Ct est inférieure à cinq, un troisième cycle de dégradation du transporteur est recommandé. Nous n'avons jamais trouvé un troisième cycle de dégradation nécessaire, et si elle se produit, il est recommandé de remplacer les stocks d'endonuclease homing.
Des séries supplémentaires d'amplification du PCR peuvent être ajoutées au protocole si le montant final de la bibliothèque obtenue n'est pas suffisant pour le séquençage. L'amplification de PCR peut alors être fixée avec le nombre minimal de cycles d'amplification nécessaires pour produire assez de matériel pour le séquençage, en tenant compte de la perte d'échantillon qui ne peut pas être évitée dans la sélection de taille. La purification ou la sélection de taille à l'aide de perles magnétiques SPRI doit alors être effectuée jusqu'à ce que tous les petits fragments (lt;200 bp) soient enlevés (si nécessaire, utiliser 0,6:1 perles au rapport d'échantillon), et la bibliothèque doit être quantifiée à l'aide de Picogreen.
Les bibliothèques peuvent être séquencées en mode unique ou à extrémité paire. À l'aide d'un séquençage à extrémité scariée, on peut obtenir des renseignements sur les isoformes de transcription. En outre, comme la transcription inversée est effectuée à l'aide d'une amorce aléatoire (TCT-N6, N6 étant un hexamer aléatoire), les informations de l'extrémité séquentée 3'-extrémité peuvent être utilisées comme identifiants moléculaires uniques (UMI) pour effondrer les doublons PCR. Comme un nombre modéré de cycles d'amplification de PCR est utilisé (jusqu'à 18), l'utilisation des IMI s'est avérée auparavant inutile.
Comme le cœur du protocole repose sur nAnT-iCAGE11, SLIC-CAGE utilise huit codes-barres. Par conséquent, le multiplexage de plus de huit échantillons n'est actuellement pas pris en charge. En outre, les deux SLIC-CAGE et nAnT-iCAGE ne sont pas adaptés pour capturer LES ARN plus court que 200 bp, que les protocoles sont conçus pour supprimer les linkers et les artefacts PCR par la taille-exclusion avec amPure XP perles.
SLIC-CAGE est la seule méthode impartiale de résolution à un seul nucléotide à faible entrée pour cartographier les sites de démarrage de transcription à l'aide de nanogrammes de matériel d'ARN total. Les méthodes alternatives reposent sur l'activité de commutation de modèle de la transcriptase inverse pour coder à barres l'ARN plafonné au lieu de cap-piégeage (par exemple, NanoCAGE15 et NanoPARE16). En raison de la commutation de modèle, ces méthodes présentent des biais séquence-spécifiques dans la détection de TSSs, menant à un nombre accru de TSS faux positifs et à une diminution du nombre de vrais TSS9,10.
Déclarations de divulgation
Un brevet pour l'ARN/ADN de porteur dégradable a été rempli.
Remerciements
Ces travaux ont été appuyés par la subvention du Wellcome Trust (106954) accordée à B. L. et au financement de base du Conseil de recherches médicales (CRM) (MC-A652-5QA10). N. C. a reçu le soutien de la bourse à long terme EMBO (EMBO ALTF 1279-2016); E. P. a reçu le soutien du Medical Research Council UK; B. L. a reçu le soutien du Medical Research Council UK (MC UP 1102/1).
matériels
| Name | Company | Catalog Number | Comments |
| 2-propanol, Bioultra, for molecular biology, ≥99.5% | Sigma-Aldrich | 59304-100ML-F | Used in RNAclean XP purification. |
| 3' linkers | Sequences are described in Murata et al. 2014 and Supplementary Table 1 of this manuscript. Annealing of strands to produce 3'linkers is described in the supplementary of this protocol. | ||
| 5' linkers | Sequences are described in Murata et al. 2014 and Supplementary Table 1 of this manuscript. Annealing of strands to produce 5'linkers is described in the supplementary of this protocol. | ||
| Agencourt AMPure XP, 60 mL | Beckman Coulter | A63881 | Purification of DNA |
| Agencourt RNAClean XP Kit | Beckman Coulter | A63987 | Purification of RNA and RNA:cDNA hybrids in CAGE steps. |
| Axygen 0.2 mL Polypropylene PCR Tube Strips and Domed Cap Strips | Axygen (available through Corning) | PCR-0208-CP-C | Or any 8-tube PCR strips (used only for water and mixes). |
| Axygen 1 x 8 strip domed PCR caps | Axygen (available through Corning) | PCR-02CP-C | Caps for PCR plates. |
| Axygen 1.5 mL Maxymum Recovery Snaplock Microcentrifuge Tube | Axygen (available through Corning) | MCT-150-L-C | Low-binding 1.5 mL tubes, used for enzyme mixes or sample concentration. |
| Axygen 96 well no skirt PCR microplate | Axygen (available through Corning) | PCR-96-C | Low-binding PCR plates - have to be used for all steps in the protocol. Note that plates should be cut to contain 2 x 8 wells for easier visibility of the samples |
| Bioanalyzer (or Tapestation): RNA nano and HS DNA kits | Agilent | To determine quality of RNA, efficient size selection and final quality of the library (Tapestation can also be used) | |
| Biotin (Long Arm) Hydrazide | Vector laboratories | SP-1100 | Biotinylation/tagging |
| Cutsmart buffer | NEB | Restriction enzyme buffer | |
| Deep Vent (exo-) DNA Polymerase | NEB | M0259S | Second strand synthesis |
| DNA Ligation Kit, Mighty Mix | Takara | 6023 | Used for 5' and 3'-linker ligation |
| dNTP mix (10 mM each) | ThermoFisher Scientific | 18427013 | dNTP mix for production of carrier templates (or any dNTPs suitable for PCR) |
| Dynabeads M-270 Streptavidin | Invitrogen | 65305 | Cap-trapping. Do not use other beads as these are optimised with the buffers used. |
| DynaMag-2 Magnet | ThermoFisher Scientific | 12321D | Magnetic stand for 1.5 mL tubes - used to prepare Streptavidin beads. |
| DynaMag-96 Side Skirted Magnet | ThermoFisher Scientific | 12027 | Magnetic stand for PCR plates (96 well-plates) - used with cut plates to contain 2 x 8 wells. |
| Ethanol, BioUltra, for molecular biology, ≥99.8% | Sigma-Aldrich | 51976-500ML-F | Used in AMPure washes. Any molecular biology suitable ethanol can be used. |
| Exonuclease I (E. coli) | NEB | M0293S | Leftover primer degradation |
| Gel Loading Dye, Purple (6x), no SDS | NEB | B7025S | agarose gel loading dye |
| HiScribe T7 High Yield RNA Synthesis Kit | New England Biolabs | E2040S | Kit for carrier in vitro transcription |
| Horizontal electrophoresis apparatus | purification of carrier DNA templates from agarose gels | ||
| I-Ceu | NEB | R0699S | Homing endonuclease used for carrier degradation. |
| I-SceI | NEB | R0694S | Homing endonuclease used for carrier degradation. |
| KAPA HiFi HS ReadyMix (2x) | Kapa Biosystems (Supplied by Roche) | KK2601 | PCR mix for target library amplification |
| KAPA SYBR FAST qPCR kit (Universal) 2x | Kapa Biosystems (Supplied by Roche) | KK4600 | qPCR mix to assess degradation efficiency and requiered number of PCR amplification cycles |
| Micropipettes and multichannel micropipettes (0.1-10 µL, 1-20 µL, 20-200 µL) | Gilson | Use of Gilson with the low-binding Sorenson tips is recommended. Other micropippetes might not be compatible. Different brand low-binding tips may not be of equal quality and may increase sample loss. | |
| Microplate reader | For Picogreen concentration measurement of the final library. Microplates are used to allow small volume measurement and reduce sample waste. | ||
| nuclease free water | ThermoFisher Scientific | AM9937 | Or any nuclease (DNase and RNase) free water |
| PCR thermal cycler | incubation steps and PCR amplficication | ||
| Phusion High-Fidelity DNA Polymerase | ThermoFisher Scientific | F530S | DNA polymerase for amplification of carrier templates (or any high fidelity polymerase) |
| QIAquick Gel Extraction Kit (50) | Qiagen | 28704 | Purification of carrier PCR templates from agarose gels. |
| qPCR machine | determining PCR amplification cyle number and degree of carrier degradation | ||
| Quant-iT PicoGreen dsDNA Reagent | ThermoFisher Scientific | P11495 | Used to measure final library concentration - recommended as, in our hands, it is more accurate and reproducible than Qubit. |
| Quick-Load Purple 100 bp DNA Ladder | NEB | N0551S | DNA ladder |
| Quick-Load Purple 1 kb Plus DNA Ladder | NEB | N0550S | DNA ladder |
| Ribonuclease H | Takara | 2150A | Digestion of RNA after cap-trapping. |
| RNase ONE Ribonuclease | Promega | M4261 | Degradation of single stranded RNA not protected by cDNA. |
| RNase-Free DNase Set | Qiagen | 79254 | Removal of carrier DNA templates after in vitro transcription. |
| RNeasy Mini Kit | Qiagen | 74104 | For cleanup of carrier RNA from in vitro transcription or capping |
| Sodium acetate, 1 M, aq.soln, pH 4.5 RNAse free | VWR | AAJ63669-AK | Or any nuclease (DNase and RNase) free solution |
| Sodium acetate, 1 M, aq.soln, pH 6.0 RNAse free | Or any nuclease (DNase and RNase) free solution | ||
| Sodium periodate | Sigma-Aldrich | 311448-100G | Oxidation of vicinal diols |
| Sorenson low binding aerosol barrier tips, MicroReach Guard, volume range 10 μL, Graduated | Sorenson (available through SIGMA-ALDRICH) | Z719390-960EA | Low-binding tips - recommended use throughout the protocol to minimise sample loss. |
| Sorenson low binding aerosol barrier tips, MultiGuard, volume range 1,000 μL , Graduated | Sorenson (available through SIGMA-ALDRICH) | Z719463-1000EA | Low-binding tips - recommended use throughout the protocol to minimise sample loss. |
| Sorenson low binding aerosol barrier tips, MultiGuard, volume range 20 μL , Graduated | Sorenson (available through SIGMA-ALDRICH) | Z719412-960EA | Low-binding tips - recommended use throughout the protocol to minimise sample loss. |
| Sorenson low binding aerosol barrier tips, MultiGuard, volume range 200 μL , Graduated | Sorenson (available through SIGMA-ALDRICH) | Z719447-960EA | Low-binding tips - recommended use throughout the protocol to minimise sample loss. |
| SpeedVac Vacuum Concentrator | concentrating samples in various steps to lower volume | ||
| SuperScript III Reverse Transcriptase | ThermoFisher Scientific | 18080044 | Used for reverse transcription (1st CAGE step) |
| Trehalose/sorbitol solution | Preparation is described in Murata et al. 2014. | ||
| Tris-HCl, 1 M aq.soln, pH 8.5 | 1 M solution, DNase and RNase free | ||
| tRNA (20 mg/mL) | tRNA solution. Preparation is described in Murata et al. 2014. | ||
| UltraPure Low Melting Point Agarose | ThermoFisher Scientific | 16520050 | Or any suitable pure low-melt agarose. |
| USB Shrimp Alkaline Phosphatase (SAP) | Applied Biosystems (Provided by ThermoFisher Scientific) | 78390500UN | |
| USER Enzyme | NEB | M5505S | Degradation of 3'linker's upper strand, Uracil Specific Excision Reagent/Enzyme |
| Vaccinia Capping System | NEB | M2080S | Enzymatic kit for in vitro capping of carrier molecules |
| Wash buffer A | Cap trapping washes. Preparation is described in Murata et al. 2014. | ||
| Wash buffer B | Cap trapping washes. Preparation is described in Murata et al. 2014. | ||
| Wash buffer C | Cap trapping washes. Preparation is described in Murata et al. 2014. |
Références
- Shiraki, T., et al. Cap analysis gene expression for high-throughput analysis of transcriptional starting point and identification of promoter usage. Proceedings of the National Academy of Sciences of the United States of America. 100 (26), 15776-15781 (2003).
- Haberle, V., Lenhard, B. Promoter architectures and developmental gene regulation. Seminars in Cell and Developmental Biology. 57, 11-23 (2016).
- Haberle, V., Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nature Reviews Molecular Cell Biology. 19 (10), 621-637 (2018).
- Andersson, R., et al. An atlas of active enhancers across human cell types and tissues. Nature. 507 (7493), 455-461 (2014).
- Consortium, E. P. An integrated encyclopedia of DNA elements in the human genome. Nature. 489 (7414), 57-74 (2012).
- Celniker, S. E., et al. Unlocking the secrets of the genome. Nature. 459 (7249), 927-930 (2009).
- Consortium, F., et al. A promoter-level mammalian expression atlas. Nature. 507 (7493), 462-470 (2014).
- Boyd, M., et al. Characterization of the enhancer and promoter landscape of inflammatory bowel disease from human colon biopsies. Nature Communications. 9 (1), 1661(2018).
- Adiconis, X., et al. Comprehensive comparative analysis of 5'-end RNA-sequencing methods. Nature Methods. , (2018).
- Cvetesic, N., et al. SLIC-CAGE: high-resolution transcription start site mapping using nanogram-levels of total RNA. Genome Research. 28 (12), 1943-1956 (2018).
- Murata, M., et al. Detecting expressed genes using CAGE. Methods in Molecular Biology. 1164, 67-85 (2014).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9, 357(2012).
- R Core Team. A language and environment for statistical computing. , Available from: https://www.R-project.org/ (2017).
- Haberle, V., Forrest, A. R., Hayashizaki, Y., Carninci, P., Lenhard, B. CAGEr: precise TSS data retrieval and high-resolution promoterome mining for integrative analyses. Nucleic Acids Research. 43 (8), e51(2015).
- Poulain, S., et al. NanoCAGE: A Method for the Analysis of Coding and Noncoding 5'-Capped Transcriptomes. Methods in Molecular Biology. 1543, 57-109 (2017).
- Schon, M. A., Kellner, M. J., Plotnikova, A., Hofmann, F., Nodine, M. D. NanoPARE: parallel analysis of RNA 5' ends from low-input RNA. Genome Research. 28 (12), 1931-1942 (2018).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.