
Method Article
Organoïdes épithéliales mucociliaires des cellules embryonnaires de Xenopus : génération, culture et imagerie vivante haute résolution
Dans cet article
Résumé
Nous décrivons un protocole simple pour développer des organoïdes épithéliales mucociliary des cellules profondes d’ectoderm isolées des embryons de Xenopus laevis. Les progéniteurs multipotents régénèrent les précurseurs épithéliales des cellules gobelet et permettent un suivi en direct de l’initiation et de la progression des transitions cellulaires à la surface des organoïdes.
Résumé
L’épithélium mucociliaire fournit la première ligne de défense en enlevant les particules étrangères par l’action de la production de mucus et du dégagement cils- médiatisé. Beaucoup de défauts médicalement pertinents dans l’épithélium mucociliary sont déduits car ils se produisent profondément dans le corps. Ici, nous introduisons un modèle 3D tractable pour l’épithélium mucociliary produit à partir d’ancêtres multipotents qui ont été microchirurgically isolés des embryons de Xenopus laevis. Les organoïdes épithéliales mucociliaires sont recouverts d’épithélium nouvellement généré à partir de cellules ectoderm profondes et décorés plus tard de cellules multiciliées à motifs distincts, de cellules sécrétrices et de cellules gobelet productrices de mucus qui sont indiscernables de l’épiderme indigène dans les 24 h. Les séquences complètes des transitions cellulaires dynamiques de la mésenchymie à l’épithéliale qui émergent sur la surface apical des organoïdes peuvent être suivies par imagerie vivante à haute résolution. Ces organoïdes épithéliales mucociliaires cultivés in vitro offrent des avantages distincts dans l’étude de la biologie de l’épithélium mucociliaire avec une efficacité élevée dans la génération, des conditions de culture définies, le contrôle du nombre et de la taille, et un accès direct à l’imagerie vivante pendant la régénération de l’épithélium différencié.
Introduction
Les dommages, l’infection, et la maladie de l’épithélium mucociliary sont associés à la production et au dégagement altérés du mucus qui se trouve souvent dans les désordres pulmonaires tels que la maladie pulmonaire obstructive chronique, l’asthme, la fibrose kystique, la bronchiectasie, et la dyskinésie ciliaireprimaire 1,2,3,4. Une avancée récente dans la technologie organoïde, par exemple, l’organoïde pulmonaire dérivé de cellules basales appelé trachéosphère qui récapitule la régénération de l’épithélium mucociliaire apparaissent comme un modèle prometteur avec un potentielthérapeutique 1,5,6. Cependant, son utilisation est actuellement limitée, en partie en raison de l’absence des conditions de culture définies et de la faible efficacité dans les productions organoïdes. L’épithélium mucociliaire dans les voies respiratoires humaines et l’épiderme de grenouille sont remarquablement semblables dans la morphologie de tissu, la composition cellulaire,et sa fonction 7,8,9,10,11,12. Dans les deux organismes, l’épithélium mucociliaire assure la défense de première ligne en sécrétant des substances mucus et antimicrobiennes et dégage des particules nocives et des agents pathogènes par l’action synchronisée des cils.
Ici, nous décrivons un protocole simple pour générer des organoïdes épithéliales mucociliaires utilisant les progéniteurs multipotents des embryons de Xenopus laevis 13,14. Précédemment, nous avonsrapporté 14 qu’en l’absence des facteurs exogènes de croissance et de la matrice extracellulaire, les cellules profondes microchirurgically isolées de l’ectoderm tôt de stade de gastrula s’assemblent spontanément en agrégats, régénèrent l’épithélium à sa surface, et mûrissent dans l’épithélium mucociliary en intercalant les cellules multiciliated et d’autres accessoires dans un délai de 24 h. En plus du développement rapide, ce protocole offre une occasion distincte d’accéder directement aux transitions des cellules ectoderm profondes multipotentes en progéniteurs épithéliales de cellules gobelet qui récapitulent les étapes de régénération d’un épithélium14 perturbé qui ne sont pas disponibles à partir d’embryons intacts et d’ectoderm (également connu sous le nom de capuchon animal)15,16,17. Le nombre et la taille des organoïdes produits sont évolutifs avec une grande efficacité en contrôlant les matériaux de départ des embryons Xenopus. Les organoïdes en culture flottante peuvent être facilement triés et transférés au stade désiré pour d’autres analyses, y compris l’imagerie à haute résolution, les essais mécaniques, le traitement médicamenteux et la caractérisationgénétique 14. Cette régénération spontanée et axée sur la mécanique tissulaire de l’épithélium à la surface des agrégats cellulaires embryonnaires entraîne des organoïdes épithéliales mucociliaires et fournit un nouveau modèle tridimensionnel (3D) pour étudier la biologie de l’épithélium mucociliaire.
Protocole
L’utilisation des animaux et les protocoles expérimentaux ont été approuvés par le comité institutionnel de soins et d’utilisation des animaux (IACUC) de l’Institute for Basic Science (IBS 18-01) et du Korea Advanced Institute of Science and Technology (KA2017-22).
1. Embryons
- Obtenir des embryons X. laevis à l’aide d’une procédure standard : recueillir manuellement les ovules des grenouilles femelles stimulées et effectuer la fécondation in vitro18,19.
- Dé-gelée les embryons fécondés avec agitation douce pendant environ 5 min dans 2% cystéine dans 1/3x modifié saline de Barth (MBS; voir la recette pour 1X MBS ci-dessous) au pH 819.
- Étape facultative pour l’imagerie en direct : pour étiqueter de façon fluorescente des protéines spécifiques et observer leur dynamique dans l’organoïde, passez à la section 5.
- (Facultatif) Pour surveiller la contamination des cellules superficielles d’ectoderm dans les organoïdes, étiquetez la surface apical des embryons avec NHS-rhodamine au stade 914. Incuber les embryons en 1 mg/mL NHS-Rhodamine en 1/3x MBS (pH 9.0) pendant 30 min avec nutation douce. Laver les embryons trois fois en les transférant dans une boîte de Pétri remplie de 1/3 x MBS pendant 15 min.
- Culture de l’embryon en 1/3x MBS à la température préférée (14-26 °C) jusqu’à ce que les premiers signes du stade 10 soient détectés (c.-à-d. l’apparition de cellules pigmentées foncées autour du blastopore à la vue végétale).
2. Préparation d’outils microchirurgaux, de solutions et de navires culturels
- Préparez les outils nécessaires, y compris une paire de forceps de qualité chirurgicale et des outils capillaire (boucle de cheveux et couteau à cheveux) pour la microchirurgie20.
- Préparer les supports culturels suivants pour les embryons : 1/3X MBS, où 1X MBS est fabriqué avec NaCl (88 mM), KCl (1 mM), NaHCO3 (2,4 mM), MgSO4 (0,82 mM), Ca (NO3)2 (0,33 mM), CaCl2 (0,41 mM) et HEPES (10 mM). Réglez le pH à 7,4 avec 10 M NaOH.
REMARQUE : Facultatif : ajouter des gouttes de phénol rouge pour indiquer le pH. - Préparez les médias culturels suivants pour les tissus embryonnaires et organoïdes : Danilchik’s for Amy (DFA)21 complété par une solution antibiotique et antimycotique fraîche de 1 %. Préparer le DFA avec nacl (53 mM), Na2CO3 (5 mM), gluconate de potassium (4,5 mM), gluconate de sodium (32 mM), CaCl2 (1 mM) et MgSO4 (1 mM). Ajuster le pH à 8,3 avec la bicine granulaire. FiltreR DFA (filtre bouteille-dessus de 0,2 μm), aliquot et le stocker à -20 °C.
- Préparez un DFA sans calcium et magnésium pour séparer les cellules profondes de l’ectoderme en utilisant la recette ci-dessus et en omettant CaCl2 et MgSO4. Aliquot et magasin à -20 °C.
- Préparer des tubes PCR non adhésifs pour l’agrégation cellulaire embryonnaire.
- Pour induire l’agrégation spontanée des cellules embryonnaires isolées, préparez des tubes PCR non adhésifs en enduissant les tubes PCR à fond rond de 200 μL de 1 % de BSA (1 g de BSA dans 100 mL d’eau distillée) pendant la nuit à 4 °C ou pendant 2 h à température ambiante. Chaque tube sera utilisé pour assembler un organoïde.
- Rincez trois fois les tubes PCR enduits de BSA pour éliminer les BSA résiduels.
- Remplissez les tubes PCR de 200 μL de DFA.
3. Isolement des cellules profondes d’ectoderm
- Sélectionnez et rassemblez des embryons lorsqu’ils atteignent le stade 10 précoce à l’aide d’outils capillaux sous stéréoscope.
- À l’aide d’une pipette de transfert jetable, transférer les embryons sélectionnés dans une boîte de Pétri remplie de DFA.
- Retirer la membrane vitelline des embryons à l’aide de forceps tranchants du côté végétal sans perturber le côté animal de l’embryon.
REMARQUE : Veillez à éviter d’exposer les embryons à l’air. L’introduction de bulles d’air dans la solution ou l’apport d’embryons à la surface provoqueront l’éclatement de l’embryon. - Pour isoler le capuchon animal, placez le côté animal de l’embryon vers le haut.
- Estimer visuellement l’étendue du bouchon animal à exciser et faire la première incision le long du bord du capuchon animal à l’aide d’un couteau à cheveux. Tirez le couteau à cheveux vers l’extérieur pour faire une coupe.
- Répétez l’étape 3.5 pour créer une chaîne de petites coupures pour exciser le bouchon animal.
- Coupez le bord épais du capuchon animal à l’aide d’un couteau à cheveux pour empêcher l’inclusion de précurseurs mésoderm.
REMARQUE : Pour éviter la guérison et l’agrégation de bouchons d’animaux isolés, passez à l’étape suivante dans les 10 minutes. En règle générale, nous isolons 5-10 bouchons d’animaux à la fois pour assembler plusieurs organoïdes. - Pour séparer les cellules profondes de l’ectoderm du bouchon animal, transférez les bouchons d’animaux excisés dans une boîte de Pétri remplie de DFA sans calcium et magnésium avec une pipette de transfert jetable. Veillez à ne pas introduire de bulles d’air pendant le transfert.
- Pour garder suffisamment d’espace pour les prochaines étapes, à l’aide des outils capillus, placez les bouchons d’animaux pour faire face au côté animal vers le haut et maintenez une distance généreuse par rapport aux autres explants.
- Attendez 5-10 min, puis surveillez les explants sous un stéréoscope. Une fois les cellules profondes desserrées ont été trouvées à partir du bord de la couche superficielle pigmentée par l’obscurité, commencer à soulever la couche superficielle loin des cellules ectoderm profondes de couleur claire à l’aide d’un couteau à cheveux sous le stéréoscope.
- Détachez soigneusement (décoller) la couche superficielle à l’aide d’un couteau à cheveux, à partir du bord.
- Recueillir les cellules ectoderm profondes avec aspiration minimale (10\u201215 μL) pour limiter la quantité de DFA sans calcium et magnésium qui est transférée dans le média d’agrégation dans l’étape suivante.
REMARQUE : Les cellules superficielles détachées peuvent être retirées du média pour éviter de contaminer les cellules ectoderm profondes restantes.
4. Génération d’organoïdes épithéliales mucocristalaires
- Transfert recueilli cellules ectoderm profondes à un tube PCR non adhésif contenant 200 μL de DFA. Pipette doucement le média (2\u20123 fois) pour disperser les cellules transférées dans le tube PCR.
REMARQUE : Le moment désigné comme heures après agrégation (hpa) commence à cette étape. La taille des organoïdes qui en résultent est contrôlée par le nombre de cellules ectoderm profondes ajoutées à un tube PCR. Des cellules ectoderm profondes d’une ou plusieurs capsules animales peuvent être utilisées, selon la taille désirée de l’organoïde. - Fermez le tube PCR. Maintenez les tubes PCR à la verticale pour induire une agrégation spontanée en bas.
- Surveillez le processus d’agrégation sous un stéréoscope. Les cellules se rassemblent généralement au bas du tube PCR en moins d’une heure et s’assemblent en agrégats sphériques à moins de 2-3 h, selon la taille.
- Pour effectuer des tests d’imagerie en direct ou de dépistage de drogues pendant le développement des organoïdes épithéliales mucociliaires, recueillir des agrégats à 2 hpa à l’aide d’une pipette de 200 μL munie de pointes agrandies (coupées avec du ciseaux stérilisé) afin d’éviter de causer des dommages aux agrégats pendant la collecte.
- Pour permettre aux agrégats de se développer dans l’organoïde épithélial mucociliaire en culture, recueillir des agrégats sphériques du tube PCR à 5 hpa et les transférer dans une boîte de Pétri remplie de DFA.
- Placez les agrégats loin des autres pour les empêcher de fusionner. Dans les 24 h de culture à température ambiante sans aucun facteur supplémentaire, on peut observer que les organoïdes épithéliales mucociliaires matures tournent avec l’action de battre les cils couvrant la surface de l’épithélium différencié
5. Imagerie en direct haute résolution (facultatif) des organoïdes en développement
- Préparer l’ARNm à la microinjection.
- Pour visualiser l’épithélialisation qui se produit au stade initial de la formation organoïde, préparez l’ARNm pour la protéine occludens zonula occludens épithéliale-1 (ZO-1) et pour la mise en valeur des membranes cellulaires en amplifiant les plasmides pCS2-ZO1-RFP et pCS2-mem-GFP (un cadeau de Lance Davidson).
- Extraire et linéariser l’ADN plasmide, puis transcrire l’ARNm plafonné à l’aide d’un kit de transcription in vitro SP6/T7.
- Aliquot l’ARNm transcrit et le stocker à -80 °C.
- Microinjecter l’ARNm dans un embryon fécondé
- Placer les embryons fécondés dans 3% Ficoll en 1x MBS.
- Chargez 3 à 4 μL d’ARNm à l’aide d’une pointe de micro-chargeuse dans une aiguille en verre tiré (une pointe d’aiguille conique longue et fine avec un diamètre intérieur de 10\u201230 μm).
- Attachez l’aiguille à un microinjecteur et ajustez le temps et la pression pour fournir un volume constant d’ARNm pour la microinjection.
- Injecter l’ARNm juste sous la surface apical du pôle animal. Une tache circulaire distincte de couleur pâle qui est causée par l’expansion du cortex est visible au moment de la microinjection.
- Transférer les embryons injectés à 1/3X MBS et les culture à l’étape 9.5.
- Recueillir les embryons étiquetés fluorescents sous un stéréoscope de fluorescence (paramètres d’excitation/émission pour GFP (488/510) et DP (532/588)).
- Procédez à l’étape 1.3.
- Assembler et culturer l’organoïde (sections 3 et 4) jusqu’au stade de développement souhaité.
- Effectuez l’imagerie en direct.
- Préparer une chambre d’imagerie à fond de verre en collant un verre de couverture dans une chambre acrylique moulue sur mesure à l’aide de graisse de silicium.
REMARQUE : Sceller hermétiquement la chambre pour éviter les fuites des médias culturels. - Remplissez la chambre d’imagerie de DFA.
- Choisissez une grille hexagonale de microscopie électronique de transmission (TEM) (75 mailles) dans un récipient à l’aide de forceps et appliquez une petite quantité de graisse sur le bord de la grille.
REMARQUE : La taille de la maille doit être plus petite que le diamètre de l’agrégat de sorte que l’agrégat se trouve sur la grille. - Appuyez légèrement pour fixer la grille TEM au fond de la chambre d’imagerie.
- Transférez les agrégats à la chambre d’imagerie et placez-les dans la grille.
REMARQUE : Évitez de positionner les agrégats à côté de la graisse. Tout au long de l’expérience, les agrégats doivent s’asseoir sur la grille TEM sans entrer en contact avec le fond de la chambre pour empêcher la compression physique. - Remplissez la chambre d’imagerie avec du DFA et scellez-la avec un verre de couverture et de la graisse.
REMARQUE : La chambre doit être étanche à l’air sans bulles d’air pour éviter toute turbulence ou mouvement pendant l’imagerie. - Pour suivre la progression de la formation organoïde épithéliale mucociliaire, recueillez des images z-stack time-lapse d’agrégats (à partir de 2 hpa) à l’aide d’un microscope confocal.
REMARQUE : Nous recueillons généralement des piles z d’environ 120 μm d’épaisseur toutes les 15 minutes à l’aide d’un objectif 20X pour suivre les comportements dynamiques des cellules, mais ces spécifications doivent être optimisées pour l’objectif des expériences.
- Préparer une chambre d’imagerie à fond de verre en collant un verre de couverture dans une chambre acrylique moulue sur mesure à l’aide de graisse de silicium.
6. Imagerie (optionnelle) mise au point d’organoïdes par fixation et immunostaining
- Fixez les organoïdes au stade désiré du développement en les transférant sur un flacon de verre rempli d’une solution fixative.
REMARQUE : Ajoutez un volume de solution fixative qui est >20 fois celui des échantillons pour assurer une fixation complète. Effectuez les processus suivants sur un nutator sauf indication contraire. En général, les organoïdes sont fixés avec 4% de paraformaldéhyde (PFA) dans PBS. Cependant, différents fixatifs peuvent être nécessaires pour détecter des protéines spécifiques. Par exemple, nous avons utilisé 4% de PFA avec 0,25% de glutaraldehyde dans PBS pour détecter la F-actine et la tubuline acétylée. Pour détecter l’intelectine (ITLN) et le ZO-1, les organoïdes sont fixés avec la solution glacée de Dent (4:1 méthanol: diméthyle sulfure) pendant la nuit à -20 °C. Les organoïdes fixes de Dent doivent être déshydratés en série avant le lavage (étape 6.3). La durée d’incubation et de lavage des anticorps peut être optimisée pour des besoins spécifiques. - Fixer les organoïdes pendant 15 min à température ambiante (RT) ou toute la nuit à 4 °C.
- Laver 3 fois avec PBST (PBS avec 0,1% Triton X-100) pendant 15 min à RT.
- Bloquer la reliure non spécifique avec 10% de sérum de chèvre dans PBST (PBSGT) pendant 1 h à RT.
- Incuber avec un anticorps primaire (1:200) en PBSGT pendant la nuit à 4 °C.
- Laver 3 fois avec PBST pendant 15 min à RT.
- Incuber avec un anticorps secondaire (1:200) en PBSGT pendant la nuit à 4 °C.
- Laver 3 fois avec PBST pendant 15 min à RT.
- Transférez les organoïdes fixes et immunotachés dans une chambre d’imagerie et procédez à l’imagerie confocale.
Résultats
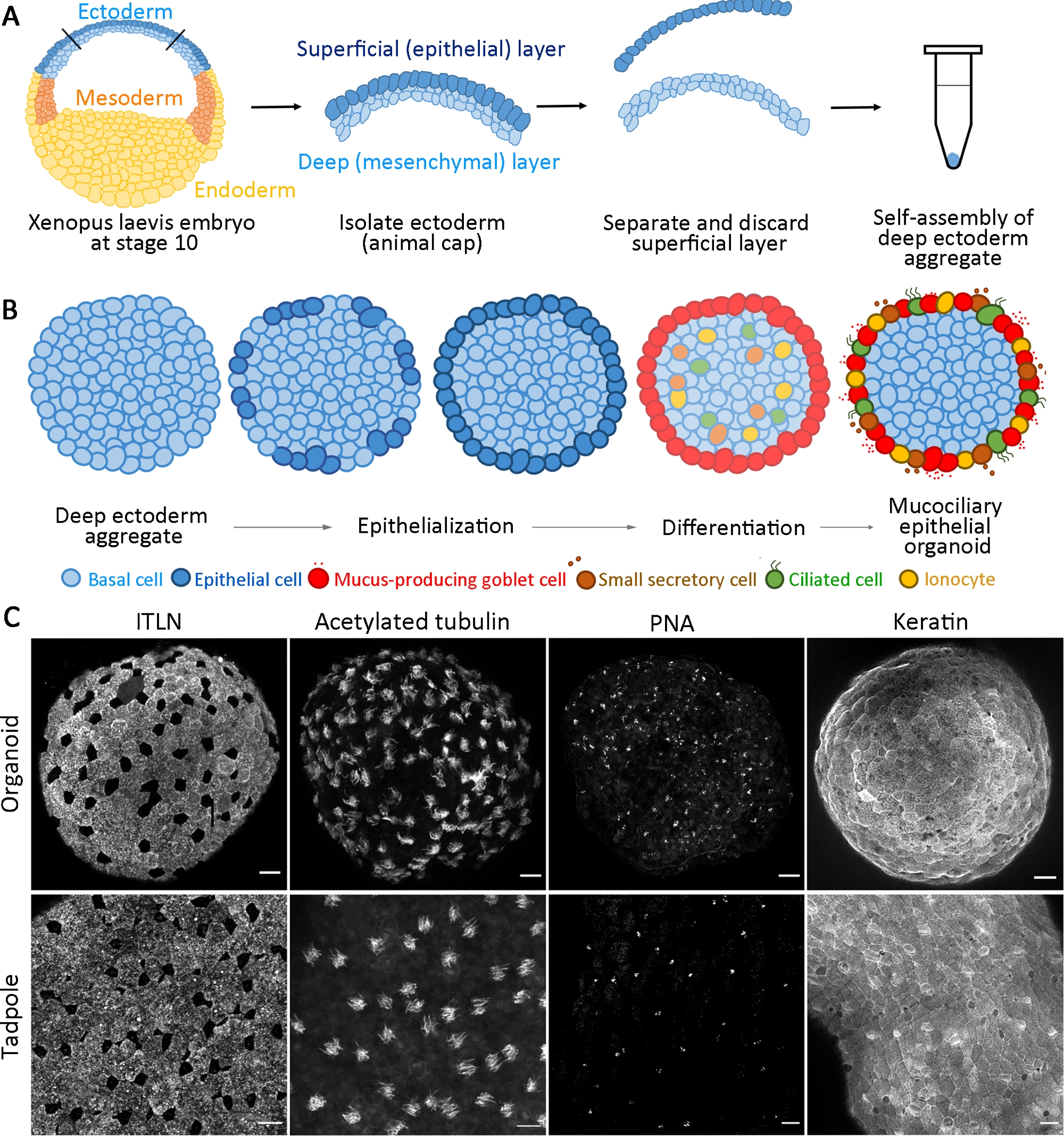
Ce protocole normalisé génère un organoïde épithélial mucociliaire à partir d’ancêtres multipotents isolés des embryons de gastrula X. laevis précoces dans les 24 h de culture14. Les cellules ectoderm profondes recueillies s’auto-assemblent pour former un agrégat dans un tube PCR non adhésif et subir une épithélialisation de surface et une différenciation des cellules gobelet. La surface nouvellement épithélialisée des agrégats fournit un substrat semblable à l’épithélium indigène que l’on trouve in vivo pour l’intercalation des cellules intérieures (p. ex., cellules multiciliées et autres cellules accessoires) et se développe pour former des organoïdes épithéliales mucociliaires(figure 1A,B). Dans les 24 h suivant l’agrégation, les organoïdes épithéliales mucociliaires auto-organisés régénèrent un épiderme mature indiscernable de l’épiderme d’un têtard. Les organoïdes comprennent l’épithélium entièrement différencié (kératine), les cellules de gobelet sécrétant le mucus (ITLN), les cellules multiciliées (tubuline acétylée) et les petites cellules sécréteurs (agglutinine d’arachide, PNA) (figure 1C).
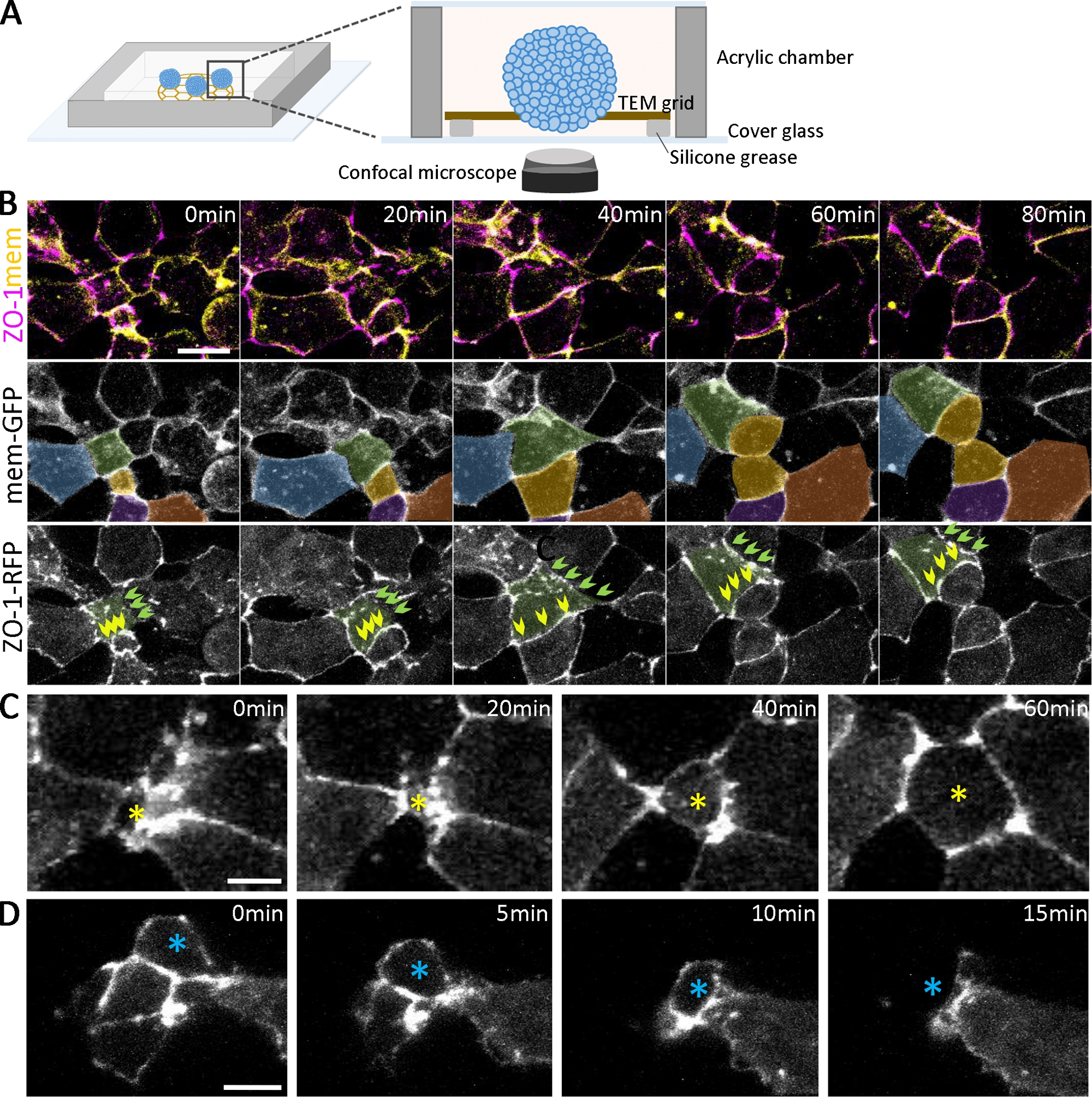
En plus de confirmer le développement de différents types de cellules avec immunostaining, la dynamique du développement organoïde peut être suivie par l’imagerie en direct (Figure 2A). Pour examiner l’épithélialisation qui émerge au stade précoce de la formation organoïde (figure 1B), nous avons étiqueté les embryons en exprimant des protéines de jonction serrées étiquetées fluorescentes (ZO-1-RFP) et des protéines de localisation des membranes (mem-GFP). Grâce à la double étiquetage, les étapes séquentielles de la formation de jonctions serrées ZO-1-positives peuvent être marquées et analysées quantitativement lors de l’épithélialisation (figure 2). Par exemple, pour les cellules(figure 2B,de couleur verte) à différents stades d’épithélialisation (à 0 min), certaines régions de l’adhérence cellule-cellule ont dispersé des ponctions de ZO-1(figure 2B, flèches vertes). En revanche, d’autres zones ont entièrement assemblé, contiguë expression ZO-1 (Figure 2B, flèches jaunes). Au fil du temps, la ponction se combine et se connecte pour former des jonctions étroites contiguës(figure 2B,flèches vertes), et les jonctions étroites contiguës maintiennent leur morphologie même pendant la division cellulaire(figure 2B,flèches jaunes). À mesure que les jonctions serrées mûrissent, les cellules se déplacent dynamiquement dans et hors de la surface le long des plans apical des organoïdes (figure 2C,D). De plus, en suivant les cellules spatiotemporally à la surface des organoïdes différeciants(figure 2B,cellules codées en couleur), une analyse à plusieurs échelles est possible, allant de la ponction individuelle aux jonctions étroites contiguës, aux limites des cellules cellulaires et aux sous-ensembles de populations cellulaires au sein des organoïdes.

Figure 1 : Génération d’organoïdes épithéliales mucocristalaires.
(A) Un schéma montrant le protocole d’assemblage d’agrégats ectoderm profonds à partir d’embryons X. laevis. (B) Un schéma pour un modèle de formation organoïde épithéliale mucociliaire provenant de cellules ectoderm profondes multipotentes (vue transversale). Les cellules placées en surface transitent vers les cellules épithéliales et se différencient en cellules gobelet. Les cellules ciliées différenciantes, les cellules sécrétaires et les ionocytes s’intercalent radialement dans la surface et régénèrent un épiderme mature. (C) Projection maximale z de mucociliary épithélium immunotaché pour ITLN (cellules gobelet productrices de mucus), tubuline acétylated (cellules ciliées), PNA (petites cellules sécrétrices), et kératine (cellules épithéliales) dans les organoïdes à 24 hpa (panneau supérieur) et épiderme têtard (panneau inférieur). Barre d’échelle = 30 μm. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : Imagerie vivante des organoïdes en développement.
(A) Un schéma de la chambre d’imagerie pour organoïdes vivants (à ne pas mettre à l’échelle). (B) Séquences time-lapse de piles confoccales recueillies à partir d’agrégats cellulaires ectoderm profonds exprimant ZO-1-RFP et mem-GFP à partir de 2,5 hpa. Barre d’échelle = 20 μm. Les cellules sont pseudo-colorées pour le suivi au fil du temps. Les cellules de couleur verte ont différents statuts d’adhérence cellulaire, y compris une adhérence positive ZO-1 en développement progressif (flèches vertes) et une adhérence positive contiguë ZO-1 (flèches jaunes) au fil du temps. (C, D) Les images confocales en accéléré des agrégats de cellules ectoderm profonds exprimant zo-1-RFP montrent les cellules intercalaires radialement se déplaçant vers la surface (C, étoile jaune) et se déplaçant à l’intérieur des agrégats (D, étoile bleue). Barres d’échelle = 10 μm. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
{kind=link}
Discussion
Les organoïdes épithéliales mucociliaires générés à partir de cellules ectoderm profondes de l’embryon X. laevis sont un outil puissant pour étudier l’épithélialisation et la différenciation des progéniteurs multipotents in vitro. Contrairement à l’essai de capuchon animal largement adopté16 utilisé pour l’organogenèse in vitro13 et le développement de l’épithélie mucociliaire15,17,22 qui utilisent l’ectoderm intact, les organoïdes dérivés de l’ectoderm profond présentés dans ce protocole offrent une occasion distincte de surveiller les phases de régénération mécanique tissulaire de l’épithélium de surface14. À environ 2 hpa, les cellules épithéliales positives ZO-1 nouvellement générées (figure 2) commencent à apparaître sur la surface apical des organoïdes et augmentent leur population pour couvrir l’organoïde entier pendant que le tissu solidifie ou réduit la conformité14. La régénération de l’épithélium et les spécifications de lignée subséquentes pour les cellules de gobelet productrices de mucus procèdent spontanément dans un média de culture chimiquement défini en un jour. Ces organoïdes épithéliales mucocristalaires qui se développent rapidement fournissent une plate-forme pour examiner les comportements cellulaires dynamiques en temps réel, en haute résolution, lors des étapes progressives de la régénération épithéliale. Ils permettent également d’investigation des questions fondamentales qui se posent pendant le développement mucociliary d’épithélium, l’homéostasie, et les maladiesassociées 2,9,23. En particulier, la sensibilité mécanique des cellules progénitrices profondes pendant la transition vers les précurseurs épithéliales de cellules de gobelet identifiés dans les organoïdes14 peut servir à relier les maladies respiratoires associées à la différenciation basale anormale où les cellules de gobelet sécrétant le mucus sont surproduiteou sous-produites 23.
Bien que ce protocole offre une approche simple pour générer ces organoïdes, il ya plusieurs étapes critiques pour le succès dans les expériences. Pour éviter la contamination des cellules épithéliales superficielles pendant l’isolement des cellules profondes d’ectoderm de la calotte animale, il faut surveiller le capuchon animal placé dans le DFA sans calcium et magnésium sous un stéréoscope et détecter le bon moment pour initier la séparation de la couche superficielle pigmentée foncée de la calotte animale. Si le tissu est maintenu dans le DFA sans calcium et magnésium pendant trop longtemps, le tissu entier se dissociera et la distinction entre les cellules profondes et superficielles serait alors impossible pour les agrégats profonds d’ectoderm. Pour confirmer l’absence de cellules superficielles dans les agrégats profonds d’ectoderm, nous recommandons d’étiqueter fluorescentement la surface apical de l’embryon avec nhs-rhodamine (étape1.4 14) avant la microchirurgie ; cela permettrait une identification facile des cellules de surface si elles existent dans les organoïdes qui en résultent. Puisque la régénération épithéliale est réglée par la mécaniquetissulaire 14,il est essentiel d’éviter la génération involontaire de force pour les organoïdes auto-organisateurs. En particulier, nous suggérons d’éviter tout contact avec le fond de verre de la chambre d’imagerie pendant l’imagerie en direct en plaçant des agrégats sur les bords des grilles TEM, car cela permet un contact libre avec la fenêtre d’imagerie des agrégats vivants (étape 5.1.2.). Ce modèle 3D in vitro-cultivé et auto-organisé pour l’épithélium mucociliaire servira d’outil tractable pour répondre aux questions fondamentales qui se posent lors de la régénération de l’épithélium et de la spécification de lignée des cellules gobelet.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous remercions les membres de Kim Lab et Lance Davidson pour leurs commentaires et leur soutien. Ces travaux ont été soutenus par la Bourse des jeunes scientifiques à HYK de l’Institute for Basic Science (IBS-R0250Y1).
matériels
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Dual-stage Glass Micropipette Puller | Narishige | PC-100 | |
| Picoliter microinjector | Warner Instruments | PLI-100A | |
| Confocal Laser Microscope | |||
| Stereoscope | |||
| Tools | |||
| Forcep | Dumont | Dumont #5 | |
| Hair knife | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| Hair loop | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| hCG injection | |||
| human chorionic gonadotropin | Sigma | cg10-10vl | |
| MBS solution | |||
| 10M Sodium hydroxide | Sigma | 72068 | |
| Calcium chloride | Sigma | C3881 | |
| Calcium nitrate | Sigma | C1396 | |
| HEPES | Sigma | H4034 | |
| Magnesium sulfate | Sigma | 230391 | |
| Phenol-red | Sigma | P0290 | |
| Potassium chloride | Sigma | 7447-40-7 | |
| Sodium bicarbonate | Sigma | S6014 | |
| Sodium chloride | Sigma | S9625 | |
| Sodium hydroxide reagent grade, 97%, powder-25g | Sigma | 655104 | |
| dejellying solution | |||
| L-Cysteine hydrochloride monohydrate | Sigma | C7880 | |
| Sodium hydroxide 10M | Sigma | 72068 | |
| Ficoll solution | |||
| Ficoll | Sigma | F4375 | |
| DFA solution | |||
| Sodium chloride | Sigma | S9625 | |
| 0.22mm Filter | Millipore | S2GPT05RE | |
| Antibiotic Antimycotic Solution | Sigma | A5955 | |
| Bicine | Sigma | B3876 | |
| Calcium chloride | Sigma | C3881 | |
| Magnesium sulfate | Sigma | 230391 | |
| Potassium gluconate | Sigma | G4500 | |
| Sodium carbonate | Sigma | 222321 | |
| Sodium gluconate | Sigma | G9005 | |
| mRNA in vitro transcription | |||
| SP6/T7 in vitro transcription kit | Invitrogen | AM1340 | |
| mRNA microinjection | |||
| Borosilicate glass capillary tubes | Harvard Apparatus | GC100-10 | |
| Eppendorf microloader pipette tips | ThermoFisher | A25547 | |
| Mineral oil | Sigma | M5904 | |
| PCR tube coating | |||
| BSA | Thermofisher | 26140079 | |
| PCR tubes | SSI | SSI-3245-00 | |
| Imaging | |||
| Custom-milled acrylic chamber | |||
| Coverglass 24mmX50mm | Duran | B01_001650 | |
| SPI Hexagonal TEM Grids, Gilded Nickel (50mesh) | SPI | 275HGN-XA | |
| SPI Hexagonal TEM Grids, Gilded Nickel (75mesh) | SPI | 2775GN-XA | |
| Silicone grease | Shinetsu | HIVAC-G | |
| Fixation | |||
| 20ml screw top-cap vial | Wheaton | WH.986580 | |
| 2ml screw top-cap vial | |||
| Benzyl alcohol | Sigma | 305197 | |
| Benzyl benzoate | Sigma | B6630 | |
| Dimethyl sulfoxide (DMSO) | Sgima | D4540 | |
| Glutaraldehyde 10% EM GRADE | Electron Microscopy Sciences | 16120 | |
| Goat serum | Jackson | 005-000-121 | |
| Methanol | Sigma | 322415 | |
| Paraforlamdehyde | Sigma | P6148 | |
| Phosphate-buffered saline (PBS) | LPS Solution | CBP007B | |
| Triton X-100 | Sigma | T8787 | |
| Primary antibody (1:200) | |||
| acetylated tubulin | Sigma | clone 6-11B-1 | |
| Itln1 | Proteintech | 11770-1-AP | |
| Keratin | Developmental Studies Hybridoma Bank | 1h5 | |
| ZO1 | Invitrogen | 402200 | |
| Vectors | |||
| pCS2-mem-GFP | Gift from Dr. Lance Davidson | ||
| pCS2-ZO1-RFP | Gift from Dr. Lance Davidson |
Références
- Barkauskas, C. E., et al. Lung organoids: current uses and future promise. Development. 144 (6), 986-997 (2017).
- Puchelle, E., Zahm, J. M., Tournier, J. M., Coraux, C. Airway Epithelial Repair, Regeneration, and Remodeling after Injury in Chronic Obstructive Pulmonary Disease. Proceedings of the American Thoracic Society. 3 (8), 726-733 (2006).
- Tilley, A. E., Walters, M. S., Shaykhiev, R., Crystal, R. G. Cilia dysfunction in lung disease. Annual Review of Physiology. 77, 379-406 (2015).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clinical Microbiology Reviews. 24 (1), 210-229 (2011).
- Rock, J. R., et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12771-12775 (2009).
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300(2019).
- Dubaissi, E., et al. A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development. 141 (7), 1514-1525 (2014).
- Hayes, J. M., et al. Identification of novel ciliogenesis factors using a new in vivo model for mucociliary epithelial development. Developmental Biology. 312 (1), 115-130 (2007).
- Walentek, P., Quigley, I. K. What we can learn from a tadpole about ciliopathies and airway diseases: Using systems biology in Xenopus to study cilia and mucociliary epithelia. Genesis. 55 (1-2), (2017).
- Werner, M. E., Mitchell, B. J. Understanding ciliated epithelia: The power of Xenopus. Genesis. 50 (3), 176-185 (2012).
- Dubaissi, E., Papalopulu, N. Embryonic frog epidermis: a model for the study of cell-cell interactions in the development of mucociliary disease. Disease Models & Mechanisms. 4 (2), 179-192 (2011).
- Walentek, P., et al. A novel serotonin-secreting cell type regulates ciliary motility in the mucociliary epidermis of Xenopus tadpoles. Development. 141 (7), 1526-1533 (2014).
- Asashima, M., et al. In vitro organogenesis from undifferentiated cells in Xenopus. Developmental Dynamics. 238 (6), 1309-1320 (2009).
- Kim, H. Y., Jackson, T. R., Stuckenholz, C., Davidson, L. A. Tissue mechanics drives regeneration of a mucociliated epidermis on the surface of Xenopus embryonic aggregates. Nature Communications. 11 (1), 665(2020).
- Haas, M., et al. DeltaN-Tp63 Mediates Wnt/beta-Catenin-Induced Inhibition of Differentiation in Basal Stem Cells of Mucociliary Epithelia. Cell Reports. 28 (13), 3338-3352 (2019).
- Green, J. Molecular Methods in Developmental Biology: Xenopus and Zebrafish. Guille, M. , Humana Press. 1-13 (1999).
- Stubbs, J. L., Davidson, L., Keller, R., Kintner, C. Radial intercalation of ciliated cells during Xenopus skin development. Development. 133 (13), 2507-2515 (2006).
- Nieuwkoop, P. D., Faber, J. Normal table of Xenopus laevis (Daudin) : a systematical and chronological survey of the development from the fertilized egg till the end of metamorphosis. , Garland Pub. (1994).
- Sive, H. L., Grainger, R. M., Harland, R. M. Early development of Xenopus laevis : a laboratory manual. , Cold Spring Harbor Laboratory Press. (2000).
- Joshi, S. D., Kim, H. Y., Davidson, L. A. Microscopy tools for quantifying developmental dynamics in Xenopus embryos. Methods in Molecular Biology. 917, 477-493 (2012).
- Sater, A. K., Steinhardt, R. A., Keller, R. Induction of neuronal differentiation by planar signals in Xenopus embryos. Developmental Dynamics. 197 (4), 268-280 (1993).
- Sedzinski, J., Hannezo, E., Tu, F., Biro, M., Wallingford, J. B. Emergence of an Apical Epithelial Cell Surface In Vivo. Developmental Cell. 36 (1), 24-35 (2016).
- Rock, J. R., Randell, S. H., Hogan, B. L. M. Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Disease Models & Mechanisms. 3 (9-10), 545-556 (2010).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.