
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
Mesure du potentiel d’action optique unicellulaire dans les cardiomyocytes dérivés de cellules souches pluripotentes induites par l’homme
Dans cet article
Résumé
Nous décrivons ici l’acquisition optique et la caractérisation des potentiels d’action à partir de cardiomyocytes dérivés de cellules souches pluripotentes induites à l’aide d’un système de photométrie modulaire à grande vitesse.
Résumé
Les techniques conventionnelles de microélectrode intracellulaire pour quantifier l’électrophysiologie des cardiomyocytes sont extrêmement complexes, nécessitent beaucoup de main-d’œuvre et sont généralement réalisées à faible débit. L’expansion rapide et continue de la technologie des cellules souches pluripotentes induites (CSI) constitue une nouvelle norme dans la recherche cardiovasculaire et d’autres méthodes sont maintenant nécessaires pour augmenter le débit de données électrophysiologiques au niveau d’une seule cellule. VF2.1Cl est un colorant sensible à la tension récemment dérivé qui fournit une réponse rapide à canal unique et de grande magnitude aux fluctuations du potentiel membranaire. Il possède une cinétique supérieure à celle des autres indicateurs de tension existants et met à disposition des données fonctionnelles équivalentes à celles des techniques traditionnelles de microélectrode. Ici, nous démontrons une caractérisation simplifiée et non invasive du potentiel d’action dans des cardiomyocytes humains dérivés de l’iPSC à rythme externe à l’aide d’un système de photométrie modulaire et très abordable.
Introduction
La modélisation électrophysiologique des cardiomyocytes et la construction de plateformes efficaces pour le dépistage des médicaments cardiaques sont essentielles pour le développement de stratégies thérapeutiques pour une variété de troubles arythmiques. L’expansion rapide de la technologie des cellules souches pluripotentes induites (IPSC) a produit des percées prometteuses dans la modélisation des maladies humaines et l’investigation pharmacologique à l’aide de cardiomyocytes dérivés de patients isolés (iPSC-CM). Les techniques de « référence » pour la caractérisation électrophysiologique de ces cellules par patch-clamp (current-clamp) peuvent quantifier la morphologie et la durée du potentiel d’action (AP), cependant, cette méthode est incroyablement complexe et lente, et mal adaptée à l’acquisition de données à haut débit1. On rapporte régulièrement que les iPSC-CM ont un potentiel de membrane diastolique accru et un courant de fuite accru par rapport aux cardiomyocytes natifs adultes2. Il est suggéré que la plus petite taille des cellules et la capacité membranaire réduite observée dans les iPSC-CM peuvent produire une erreur systématique lors de l’utilisation de la technique de serrage de courant, expliquant peut-être ces écarts3. Afin de maximiser l’utilité d’une plate-forme iPSC-CM, une méthode supplémentaire est précieuse pour augmenter le débit et assurer la précision des données lors de la caractérisation des changements de tension transmembranaire au niveau d’une seule cellule dans les iPSC-CM.
Les colorants sensibles à la tension (VSD) sont depuis longtemps une méthode proposée pour fournir une analyse plus rapide, non invasive et équivalente de la cinétique des AP cardiaques par rapport à celles des techniques traditionnelles4. Une étude récente a démontré l’adéquation de la photométrie de sonde sensible à la tension ratiométrique pour quantifier avec précision l’APcardiaque 5. De plus, la capacité d’étendre facilement les approches de photométrie optique prête cette technique aux écrans de cardiotoxicité à grande échelle essentiels au développement de médicaments thérapeutiques (par exemple, CiPA). Le développement de protocoles de cardiotoxicité standardisés dans le cas d’une étude multi-sites en aveugle utilisant des techniques optiques de réseau de microélectrodes et de détection de tension a démontré la valeur clé de cette approche6.
De nombreux colorants potentiométriques sont disponibles dans le commerce, et le développement synthétique en cours de nouvelles sondes montre un potentiel passionnant pour rationaliser leur efficacité dans une variété de constructions cardiaques et neuronales. Le VSD idéal aura une cinétique et une sensibilité accrues, tout en affichant une diminution de la charge capacitive, du photobleaching et de la cytotoxicité. Le VF2.1Cl (FluoVolt) récemment synthétisé exprime bon nombre de ces propriétés bénéfiques en grande partie en raison de sa nouvelle structure moléculaire filaire, partagée par d’autres membres de la nouvelle famille VoltageFluor (VF)7. Contrairement aux VSD électrochromes courants dans lesquels de simples sondes se conjuguent moléculairement et électriquement à la membrane plasmique, ce colorant consiste en un fil synthétique inséré passivement et couvrant la membrane qui associe un donneur riche en électrons à un fluorophore de fluorescéine modifié (FITC). Les détails mécanistes sont fournis à la figure 1. Ce colorant démontre une excellente sensibilité aux fluctuations de tension de la membrane, affichant un changement de 27% de l’intensité d’émission par 100 mV contre environ 10% observé dans d’autres sondes courantes à des vitesses comparables7. En outre, les systèmes PeT à base de fil n’interagissent pas directement avec le champ électrique cellulaire, ce qui produit des interférences électriques minimales et des changements négligeables de la charge capacitive cellulaire.

Figure 1 : Paramètres chimiques, spectraux et mécanistes du colorant VF2.1Cl. (A) Structure chimique de VF2.1Cl. Les caractéristiques moléculaires à noter comprennent de multiples groupes alkyles dans le fil moléculaire phénylène vinylène qui facilitent l’insertion dans la membrane plasmique. Un groupe d’acide sulfonique chargé négativement conjugué à la sonde FITC assure la stabilisation du fluorophore sur la surface extracellulaire et facilite l’insertion près perpendiculaire par rapport au champ électrique de la bicouche lipidique. (B) Schéma simplifié de VF2.1Cl perpendiculaire s’intégrant dans la membrane plasmique d’une cellule cible. (C) Spectres d’absorption et d’émission du colorant VF2.1Cl. Spectra est identique à celui des sondes FITC et GFP standard. (D) Représentation du mode d’action mécaniste de VF2.1Cl. Dans des conditions de repos (hyperpolarisées), des tensions intracellulaires négatives entraînent les électrons libres vers le fluorophore rostral. L’abondance d’électrons garantit que le transfert d’électrons photo-induit (PeT) est privilégié comme une voie hors de l’état excité après l’excitation optique, éteignant efficacement la fluorescence. En revanche, un potentiel membranaire dépolarisé influence le mouvement descendant des électrons favorisant la fluorescence lors de l’excitation optique. La réponse fluorescente qui en résulte est linéairement liée à la tension de la membrane et peut être utilisée avec précision pour recueillir des informations temporelles détaillées sur la cinétique électrophysiologique cellulaire. (E) Images représentatives en champ lumineux (supérieur) et fluorescence à 470 nm (inférieur) de cardiomyocytes de léporine chargés de VF2.1Cl. (F) Z pile d’un seul cardiomyocyte chargé. Les flèches indiquent les zones de localisation claire de VF2.1Cl à la membrane cellulaire. Les images ont été acquises avec un système confocal à disque rotatif composé d’une tête confocale à disque rotatif X-lightv3 avec un motif sténopé de 50 μm; Illuminateur LDI-7; Caméra Prime95B et objectif PlanApo Lambda 100x. Barre d’échelle : 20 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
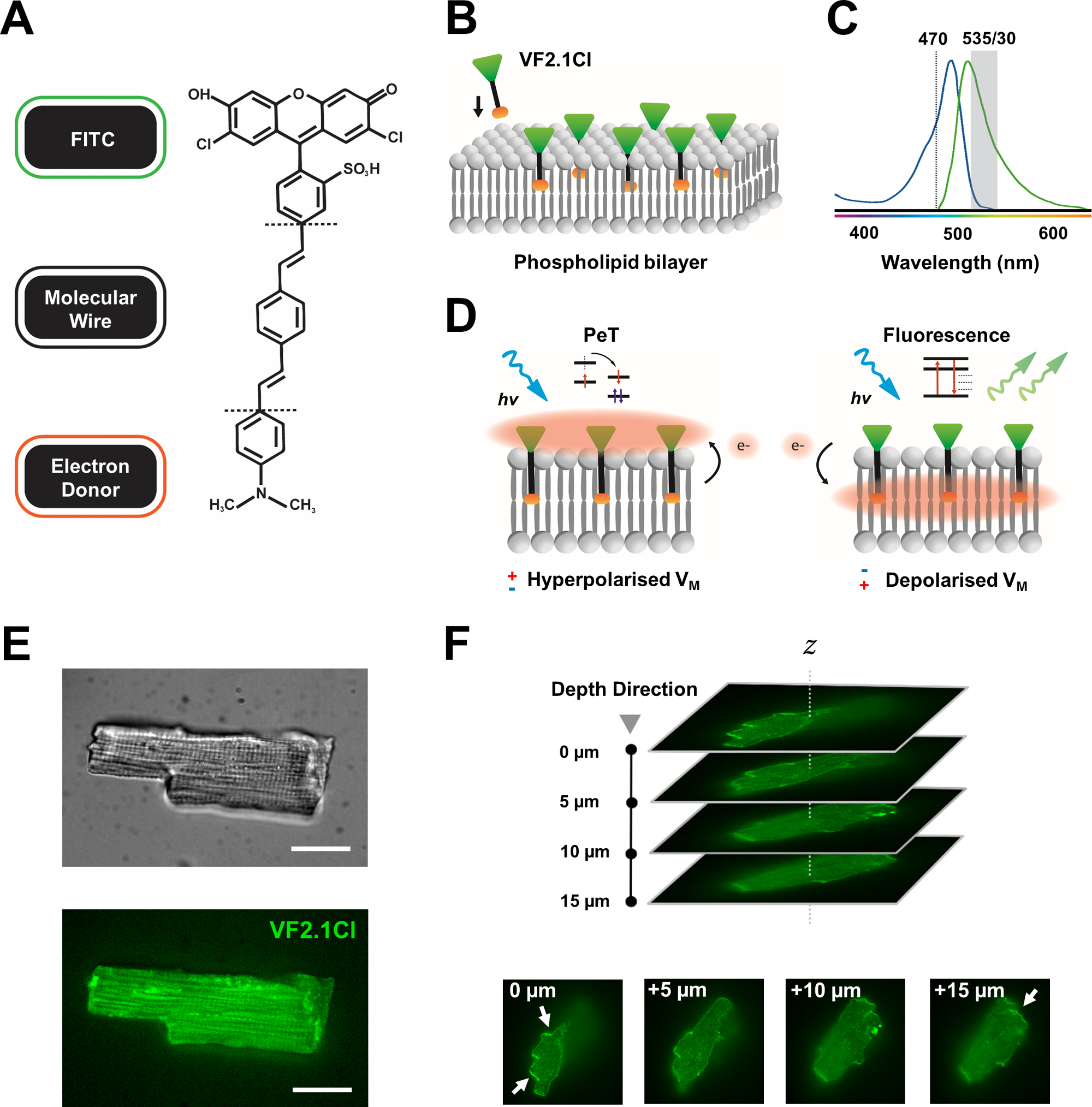{kind=link}
La sonde FITC conjuguée à VF2.1Cl garantit qu’elle peut être utilisée efficacement dans des configurations de filtre standard et GFP, et qu’elle ne nécessite qu’un système d’acquisition à canal unique, qui sont toutes deux des caractéristiques communes des plates-formes d’imagerie fluorescente. L’analyse de monocouches iPSC-CM humaines denses avec ce colorant a été récemment rapportée8,9,10,11. Notre protocole diffère de ces études en raison de notre étude des iPSC-CC uniques et isolés, non perturbés par les influences électriques et paracrines des monocouches syncytaires denses, et de notre utilisation d’un système de photométrie abordable et personnalisable par opposition à des arrangements complexes d’imagerie confocale ou à grand champ.
Nous décrivons ici notre protocole pour l’acquisition et l’analyse rapides de points d’accès optiques robustes à partir de cardiomyocytes humains isolés dérivés de l’iPSC et de cardiomyocytes natifs (voir Dossier supplémentaire). Nous utilisons VF2.1Cl couplé à une plate-forme de pointe personnalisable pour les mesures de photométrie à cellule unique. Ces protocoles expérimentaux ont été approuvés par le comité d’éthique du Centre médical universitaire de Göttingen (n° 10/9/15).
Protocole
1. Préparations cellulaires
REMARQUE: Les CS IPS humains utilisés dans ce protocole ont été dérivés de donneurs sains et différenciés en monocouches en utilisant une modulation de petites molécules entièrement définie des techniques de signalisation WNT et de purification du lactate comme décrit précédemment12,13,14. Les iPSC-CM ont été maintenus tous les 2-3 jours avec un milieu de culture décrit ci-dessous.
- Préparer un milieu de culture de milieu basal (RPMI 1640) et un supplément à 2% (B27). Conserver à 4 °C. Utilisation à température ambiante (RT).
- Préparer un milieu de placage de milieu basal (RPMI 1640), de supplément à 2% (B27) et d’inhibiteur de ROCK 1:2000. Conserver à 4 °C. Utilisation à RT.
- Recouvrir les couvercles en verre rond #0 de 10 mm stérilisés avec 150 μL de matrice de membrane basale sans facteur 1:60 et incuber à 4 °C pendant 4 h.
REMARQUE: L’optimisation du volume de glissement des couvercles est nécessaire pour s’assurer que tout le verre est recouvert tout en maintenant une tension superficielle adéquate pour éviter les déversements. 150 μL est recommandé pour les couvercles ronds de 10 mm. La taille du lot, le type de couvercle, le volume du couvercle et le type de plaque de culture peuvent être adaptés aux besoins des expérimentateurs. - Commencez la dissociation iPSC-CM avec un réactif de dissociation cellulaire à base d’EDTA. Assurez-vous que la monocouche est complètement détachée en rinçant doucement avec une pipette de 1 000 μL.
- Transférer la suspension cellulaire dans un tube de 15 mL et ajouter un milieu de placage à double volume. Centrifuger pendant 10 min à 100 x g.
- Resuspendez la pastille avec le volume souhaité (volume de remise en service) du milieu de placage. Compter les cellules manuellement ou électroniquement.
- Sélectionnez la densité optimale par couvercle (15 000) qui permettra l’analyse cellulaire isolée plus tard.
- Calculez le volume de suspension cellulaire « active » (A) nécessaire pour plaquer tous les couvercles à cette densité souhaitée. Appliquez la formule suivante et retirez dans un tube séparé:

- Calculez le volume de milieu de placage supplémentaire (B) nécessaire pour accueillir chaque couvercle à un volume souhaité. Appliquez la formule suivante et ajoutez le volume obtenu au tube de suspension active:

- Retirez la matrice des couvercles et appliquez le « volume du couvercle » de la suspension cellulaire (A + B) sur chaque couvercle. Ressuspendez régulièrement dans le tube pour assurer une distribution cellulaire uniforme.
- Incuber à 37 °C pendant 1 h. Remplissez doucement le puits avec du milieu de placage.
- Après 24 h, échangez les médias avec le milieu de culture normal et maintenez tous les 2-3 jours.
2. Configuration expérimentale
- Équipez un microscope à épifluorescence inversée d’une lentille à haute ouverture numérique à grossissement de 40x (N.A: > 0,75) pour mener des expériences.
- Couplez une LED blanche chaude à commutation rapide sur le port d’éclairage transmis du microscope. Insérez un simple filtre rouge de 660 nm dans le trajet de la lumière transmise.
REMARQUE: Cette lumière peut être activée tout au long des expériences de photométrie pour observer l’échantillon sans contaminer le signal fluorescent vert. - Montez une tête LED à commutation rapide de 470 nm pour l’enregistrement photométique. Insérez un filtre d’excitation 470/40 à l’angle épifluorescent du microscope pour nettoyer la lumière générée par la LED.
REMARQUE: Pour une quantification optimale du signal, un système d’éclairage avec contrôle de rétroaction à haute vitesse de la sortie optique est recommandé. - Insérez un cube de microscope contenant un séparateur de faisceau de passage de 495 nm de long dans le carrousel de l’unité miroir du microscope.
- Ajustez un bras de détection contenant un diaphragme de champ réglable sur le port de monture C du microscope pour permettre la sélection de la région d’intérêt.
- Couplez séparément un détecteur photomultiplicateur (PMT) et une caméra USB au microscope. Cela constituera la base du système de détection des émissions.
- Insérez un cube filtrant contenant un séparateur de faisceau passe-long de 565 nm et un filtre d’émission 535/50 dans le port PMT. Cela divise la lumière d’émission entre les deux détecteurs.
REMARQUE: Une caméra fixée au port transmis du système de détection des émissions peut détecter la lumière transmise sous fond clair tout au long de toutes les expériences. - Couplez le PMT à une alimentation et à un amplificateur PMT. Connectez la sortie de l’amplificateur PMT à une broche d’entrée analogique d’un système d’acquisition de données.
- Filtrez les données analogiques du PMT à 1 kHz ou plus.
- Numérisez les données à une fréquence au moins le double de celle de la fréquence la plus élevée présente dans le signal analogique (2 kHz ou plus) pour répondre aux critères de Nyquist et empêcher l’aliasing.
3. Chargement cellulaire avec VF2.1Cl
REMARQUE: Toutes les étapes impliquant ce colorant doivent être effectuées dans des conditions de faible luminosité.
- Préparer une solution de bain de Tyrode de (en mM): 140 NaCl, 10 HEPES, 10 Glucose, 4 KCl, 1 MgCl2,2 CaCl2,pH = 7,35 et chaud à 37 °C.
- Préparer une aliquote de solution de chargement dans un tube de microcentrifugation en mélangeant 5 μL de 1 000 x VF2.1Cl et 50 μL de solution de poloxamère solubilisante à 20 %.
- Appliquer 5 μL de la solution de chargement sur 5 mL de solution de Tyrode chauffée (concentration totale de colorant de 0,1x) dans une boîte de Petri de 20 mm.
REMARQUE: La concentration finale de colorant est de 0,1x. C’est1/10ème de ce qui a été suggéré par le fabricant. Cela permet de préserver les ressources, d’assurer une cytotoxicité négligeable et, surtout, de conserver des signaux optiques clairs provenant de cellules chargées avec des rapports signal/bruit élevés. - Ajouter un seul couvercle iPSC-CM au plat et incuber à 37 °C pendant 20 min.
- Assemblez une chambre d’imagerie à cellules vivantes chauffée. Montez sur l’étage du microscope et remplissez avec 500 μL de solution fraîche de Tyrode.
- Laver le couvercle avec la solution fraîche de Tyrode à 37 °C.
- Appliquez soigneusement le couvercle iPSC-CM sur la chambre de bain préchauffée à l’aide de pinces à pointe fine.
REMARQUE: Assurez-vous que la chambre et son contenu sont toujours chauffés à des températures physiologiques. Si vous le souhaitez, commencez par une perfusion continue de la solution de Tyrode réchauffée.
4. Stimulation du champ électrique
REMARQUE: Le déclenchement externe d’iPSC-CM est facultatif mais utile pour la normalisation de la dynamique cellulaire et des paramètres expérimentaux. Il augmente la facilité d’analyse et permet d’enquêter sur les effets dépendant de la fréquence.
- Fixez un insert de stimulation avec deux électrodes en platine espacées de 5 mm dans la chambre d’enregistrement.
- Connectez un stimulateur externe à l’insert de stimulation. Réglez sur des impulsions de champ bipolaire de 5 ms à 0,5 Hz.
- Déterminez la tension de stimulation optimale en augmentant le stimulus de 1 V vers le haut. Le stimulus de seuil est défini comme la tension la plus basse à laquelle les cellules commencent à se contracter. Appliquez des tensions d’environ 25 % au-dessus de ce seuil. La plage normale est comprise entre 1 V et 30 V.
- Fixez la fréquence de stimulation avec le stimulateur externe ou déclenchez-la avec un logiciel d’acquisition.
5. Acquisition de potentiel d’action optique
REMARQUE: Ce protocole utilise un logiciel commercial pour l’acquisition et l’analyse.
- Visualisez les myocytes en mode champ clair à l’aide du chemin de lumière transmis et de la caméra USB.
- Sélectionnez une cellule isolée et recadrez étroitement son chemin optique avec le diaphragme de champ en veillant à ce que seule la lumière de la cellule d’intérêt soit surveillée.
- Activez l’amplificateur PMT et réglez l’alimentation PMT sur 750 V.
- Exécutez le protocole de stimulation (voir étape 4) avec le logiciel d’acquisition et activez simultanément la lumière d’excitation de 470 nm. Ce dernier peut se faire via un panneau distant ou automatisé à une intensité fixe (signal TTL).
- Ajustez le gain et le décalage de l’amplificateur PMT pour vous assurer que le signal ne sature pas et est optimisé pour la plage de détection du système d’enregistrement.
- Enregistrez 10 balayages en veillant à ce que des potentiels d’action stables soient détectés.
- Continuez l’enregistrement et déplacez immédiatement l’étage du microscope pour acquérir brièvement le signal de fond d’une région dépourvue de cellules. Éteignez le voyant d’excitation.
REMARQUE: Cette valeur de fond(décalage F) sera utilisée pour tenir compte de toute fluorescence de fond. - Si vous le souhaitez, perfuser localement des médicaments de référence tels que la nifédipine de 1 μM pour identifier les réponses cellulaires à la manipulation pharmacologique.
- De manière séquentielle, répétez les étapes 5.2 à 5.7, en sélectionnant chaque fois une nouvelle cellule. Remplacez les couvercles si vous le souhaitez pour assurer un roulement expérimental élevé en une seule séance.
REMARQUE : Les protocoles de chargement et d’acquisition d’images sont décrits à la figure 2.
6. Analyse des données
- Ouvrez un enregistrement enregistré avec le logiciel d’analyse et en moyenne 10 balayages contenant des potentiels d’action stimulés à partir d’une seule cellule.
- Prenez une moyenne du signal de base représentant ledécalage F et soustrayez-la de la trace moyenne.
- Calculer ∆F/F0 avec la formule suivante (où F est mesuré fluorescence et F0 est fluorescence diastolique) :

- Identifier la trace de référence (diastolique) et la zone d’intérêt (PA) et mesurer les paramètres de potentiel d’action cardiaque souhaités. Cela inclut, mais sans s’y limiter, le temps de désintégration pour la repolarisation à50%(APD 50) età 90%(APD 90).
- Exportez les données de cette cellule unique vers un tableur.
- Répétez les étapes 6.1 à 6.5 pour tous les enregistrements. Évaluer les résultats à l’auprès de tests non appaire appropriés ou d’une analyse de la variance.

Figure 2 : Protocoles de chargement et d’acquisition d’images. (A) Organigramme du protocole complet de chargement VF2.1Cl pour les iPSC-CM et les cardiomyocytes natifs. (B) Schéma simplifié des configurations de séparateur de faisceau (BS) et de filtre utilisées dans ce protocole pour l’excitation et la détection de l’émission de VF2.1Cl en réponse aux changements de tension transmembranaire. Veuillez cliquer ici pour voir une version agrandie de cette figure.
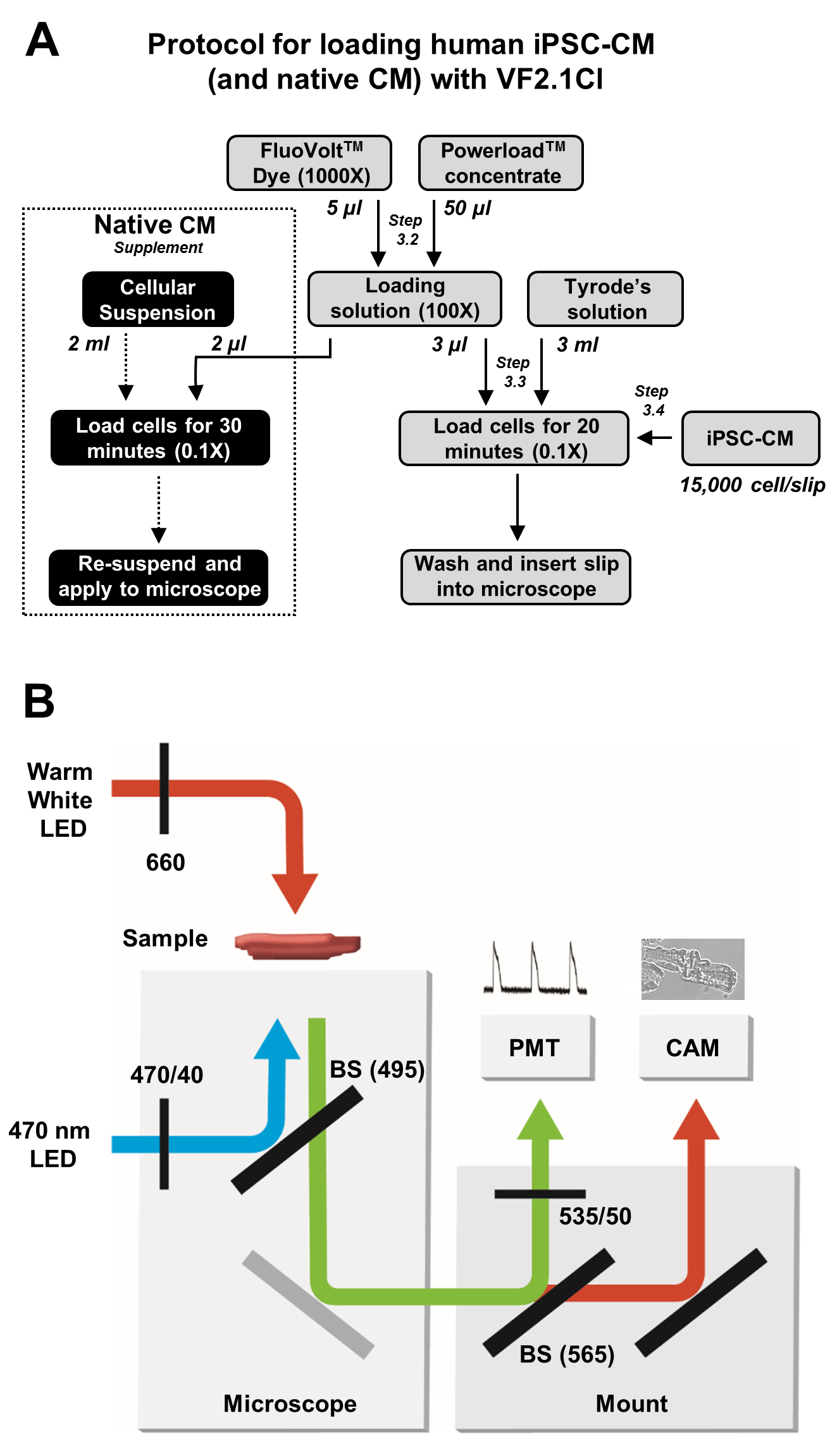{kind=link}
Résultats

Figure 3 : Profils de potentiel d’action optique (PA) de cardiomyocytes natifs isolés et de cardiomyocytes dérivés de cellules souches pluripotentes induites par l’homme (iPSC-CM). (A) AP optique représentatif d’un seul cardiomyocyte murin (au centre) avec un SEM moyen ± d’APD50 et aPD90 (n = 7, à droite). (B) AP optiqu...
Discussion
Nous décrivons ici un protocole de base permettant d’acquérir facilement des profils AP détaillés à partir d’iPSC-CM isolés adaptés à la modélisation électrophysiologique et au dépistage de médicaments cardiaques. Nous détectons des points d’accès réguliers et robustes à partir de nos iPSC-MC peu ensemencés, ce qui suggère à la fois la fonctionnalité des indicateurs et la fidélité méthodologique.
En raison du large éventail de méthodologies commerciales pour la r...
Déclarations de divulgation
Cairn Research Ltd a soutenu cette publication en couvrant les coûts de production du fichier vidéo.
Remerciements
Les auteurs tiennent à remercier Cairn Research Ltd. pour leur aimable contribution financière qui a couvert les coûts de production de cette publication. De plus, nous remercions Mme Ines Mueller et Mme Stefanie Kestel pour leur excellent soutien technique.
Les recherches des auteurs sont soutenues par le Centre allemand de recherche cardiovasculaire (DZHK), la Deutsche Forschungsgemeinschaft (DFG, Fondation allemande pour la recherche, VO 1568/3-1, IRTG1816 RP12, SFB1002 TPA13 et dans le cadre de la stratégie d’excellence de l’Allemagne - EXC 2067/1- 390729940) et la Else-Kröner-Fresenius Stiftung (EKFS 2016_A20).
matériels
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| 0.25 Trypsin EDTA | Gibco | 25200056 | |
| B27 Supplement | Gibco | 17504044 | |
| CaCl2 | Carl Roth | HN04.2 | |
| D(+)-Glucose anhydrous BioChemica | ITW Reagents | A1422 | |
| Fetal Bovine Serum | Gibco | 10270-106 | |
| FluoVolt Membrane Potential Kit | Invitrogen | F10488 | |
| HEPES | Carl Roth | HN77.4 | |
| KCl | Sigma-Aldrich | 6781.1 | |
| Lamanin | Sigma-Aldrich | 114956-81-9 | |
| Matrigel | BD | 354230 | |
| NaCl | Sigma-Aldrich | 9265.2 | |
| Nifedipine | Sigma-Aldrich | 21829-25-4 | |
| Penicillin/Streptomycin | Invitrogen | 15140 | |
| ROCK Inhibitor Y27632 | Stemolecule | 04-0012-10 | |
| RPMI 1640 Medium | Gibco | 61870010 | |
| Versene EDTA | Gibco | 15040033 | |
| Equipment | |||
| 495LP Dichroic Beamsplitter | Chroma Technology | ||
| Axopatch 200B Amplifier | Molecular Devices | ||
| Circle Coverslips, Thickness 0 | Thermo Scientific | CB00100RA020MNT0 | |
| Digidata 1550B | Molecular Devices | ||
| Dual OptoLED Power Supply | Cairn Research | ||
| ET470/40x Excitation Filter | Chroma Technology | ||
| ET535/50m | Chroma Technology | ||
| Etched Neubauer Hemacytometer | Hausser Scientific | ||
| Filter Cubes | Cairn Research | ||
| IX73 Inverted Microscope | Olympus | ||
| MonoLED | Cairn Research | ||
| Multiport Adaptors | Cairn Research | ||
| Myopacer Cell Stimulator | IonOptix | ||
| Optomask Shutter | Cairn Research | ||
| Optoscan System Controller | Cairn Research | ||
| PH-1 Temperature Controlled Platform | Warner Instruments | ||
| Photomultiplier Detector | Cairn Research | ||
| PMT Amplifier Insert | Cairn Research | ||
| PMT Supply Insert | Cairn Research | ||
| RC-26G Open Bath Chamber | Warner Instruments | ||
| SA-OLY/2AL Stage Adaptor | Olympus | ||
| T565lpxr Dichroic Beamsplitter | Chroma Technology | ||
| T660lpxr Dichroic Beamsplitter | Chroma Technology | ||
| TC-20 Dual Channel Temperature Controller | npi Electronic | ||
| UPLFLN 40X Objective | Olympus | ||
| USB 3.0 Colour Camera | Imaging Source | ||
| Software | |||
| Clampex 11.1 | Molecular Devices | ||
| Clampfit 11.1 | Molecular Devices | ||
| IC Capture 2.4 | Imaging Source | ||
| Prism 8 | Graphpad |
Références
- Miller, E. W. Small molecule fluorescent voltage indicators for studying membrane potential. Current Opinion in Chemical Biology. 33, 74-80 (2016).
- Liang, P., et al. Drug screening using a library of human induced pluripotent stem cell-derived cardiomyocytes reveals disease-specific patterns of cardiotoxicity. Circulation. 127 (16), 1677-1691 (2013).
- Horváth, A., et al. Low resting membrane potential and low inward rectifier potassium currents are not inherent features of hiPSC-derived cardiomyocytes. Stem Cell Reports. 10 (3), 822-833 (2018).
- Salama, G., Morad, M. Merocyanine 540 as an optical probe of transmembrane electrical activity in the heart. Science. 191 (4226), 485-487 (1976).
- Hortigon-Vinagre, M., et al. The use of ratiometric fluorescence measurements of the voltage sensitive dye Di-4-ANEPPS to examine action potential characteristics and drug effects on human induced pluripotent stem cell-derived cardiomyocytes. Toxicological Sciences. 154 (2), 320-331 (2016).
- Blinova, K., et al. International multisite study of human-induced pluripotent stem cell-derived cardiomyocytes for drug proarrhythmic potential assessment. Cell Reports. 24 (13), 3582-3592 (2018).
- Miller, E. W., et al. Optically monitoring voltage in neurons by photo-induced electron transfer through molecular wires. Proceedings of the National Academy of Sciences. 109 (6), 2114-2119 (2012).
- Bedut, S., et al. High-throughput drug profiling with voltage- and calcium-sensitive fluorescent probes in human iPSC-derived cardiomyocytes. American Journal of Physiology-Heart and Circulatory Physiology. 311 (1), 44-53 (2016).
- McKeithan, W. L., et al. An automated platform for assessment of congenital and drug-induced arrhythmia with hiPSC-derived cardiomyocytes. Frontiers in Physiology. 8, 766 (2017).
- Duncan, G., et al. Drug-mediated shortening of action potentials in LQTS2 human induced pluripotent stem cell-derived cardiomyocytes. Stem Cells and Development. 26 (23), 1695-1705 (2017).
- Asakura, K., Hayashi, S., Ojima, A., Taniguchi, T., Miyamoto, N. Improvement of acquisition and analysis methods in multi-electrode array experiments with iPS cell-derived cardiomyocytes. Journal of Pharmacological and Toxicological Methods. 75, 17-26 (2015).
- Lian, X., et al. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nature Protocols. 8 (1), 162-175 (2013).
- Burridge, P. W., et al. Chemically defined generation of human cardiomyocytes. Nature methods. 11 (8), 855-860 (2014).
- Kleinsorge, M., Cyganek, L. Subtype-directed differentiation of human iPSCs into atrial and ventricular cardiomyocytes. STAR Protocols. , 100026 (2020).
- Knollmann, B. C., Katchman, A. N., Franz, M. R. Monophasic action potential recordings from intact mouse heart: validation, regional heterogeneity, and relation to refractoriness. Journal of Cardiovascular Electrophysiology. 12 (11), 1286-1294 (2001).
- Leopold, J. A., Loscalzo, J. Emerging role of precision medicine in cardiovascular disease. Circulation Research. 122 (9), 1302-1315 (2018).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. Journal of Visualized Experiments. (77), e50235 (2013).
- Voigt, N., et al. Enhanced sarcoplasmic reticulum Ca2+ Leak and increased Na+-Ca2+ exchanger function underlie delayed afterdepolarizations in patients with chronic atrial fibrillation. Circulation. 125 (17), 2059-2070 (2012).
- Voigt, N., et al. Cellular and molecular mechanisms of atrial arrhythmogenesis in patients with paroxysmal atrial fibrillation. Circulation. 129 (2), 145-156 (2014).
- Fakuade, F. E., et al. Altered atrial cytosolic calcium handling contributes to the development of postoperative atrial fibrillation. Cardiovascular Research. , 162 (2020).
- Gross, E., Bedlack, R. S., Loew, L. M. Dual-wavelength ratiometric fluorescence measurement of the membrane dipole potential. Biophysical Journal. 67 (1), 208-216 (1994).
- Matiukas, A., et al. Near-infrared voltage-sensitive fluorescent dyes optimized for optical mapping in blood-perfused myocardium. Heart Rhythm. 4 (11), 1441-1451 (2007).
- Mutoh, H., et al. Spectrally-resolved response properties of the three most advanced fret based fluorescent protein voltage probes. PLoS One. 4 (2), 4555 (2009).
- Hochbaum, D. R., et al. All-optical electrophysiology in mammalian neurons using engineered microbial rhodopsins. Nature Methods. 11 (8), 825-833 (2014).
- Huang, Y. L., Walker, A. S., Miller, E. W. A photostable silicon rhodamine platform for optical voltage sensing. Journal of the American Chemical Society. 137 (33), 10767-10776 (2015).
- Deal, P. E., Kulkarni, R. U., Al-Abdullatif, S. H., Miller, E. W. Isomerically pure tetramethylrhodamine voltage reporters. Journal of the American Chemical Society. 138 (29), 9085-9088 (2016).
- Fluhler, E., Burnham, V. G., Loew, L. M. Spectra, membrane binding, and potentiometric responses of new charge shift probes. Biochemistry. 24 (21), 5749-5755 (1985).
- Fromherz, P., Muller, C. O. Voltage-sensitive fluorescence of amphiphilic hemicyanine dyes in neuron membrane. Biochimica et Biophysica Acta. 1150 (2), 111-122 (1993).
- Salama, G., et al. Properties of new, long-wavelength, voltage-sensitive dyes in the heart. Journal of Membrane Biology. 208 (2), 125-140 (2005).
- Jin, L., et al. Single action potentials and subthreshold electrical events imaged in neurons with a fluorescent protein voltage probe. Neuron. 75 (5), 779-785 (2012).
- Kralj, J. M., Douglass, A. D., Hochbaum, D. R., MacLaurin, D., Cohen, A. E. Optical recording of action potentials in mammalian neurons using a microbial rhodopsin. Nature Methods. 9 (1), 90-95 (2012).
- Tsutsui, H., Karasawa, S., Okamura, Y., Miyawaki, A. Improving membrane voltage measurements using FRET with new fluorescent proteins. Nature Methods. 5 (8), 683-685 (2008).
- Lundby, A., Mutoh, H., Dimitrov, D., Akemann, W., Knöpfel, T. Engineering of a genetically encodable fluorescent voltage sensor exploiting fast Ci-VSP voltage-sensing movements. PLoS One. 3 (6), 2514 (2008).
- Bradley, J., Luo, R., Otis, T. S., DiGregorio, D. A. Submillisecond optical reporting of membrane potential in situ using a neuronal tracer dye. The Journal of neuroscience. 29 (29), 9197-9209 (2009).
- Herron, T. J., Lee, P., Jalife, J. Optical imaging of voltage and calcium in cardiac cells & tissues. Circulation Research. 110 (4), 609-623 (2012).
- Kappadan, V., et al. High-resolution optical measurement of cardiac restitution, contraction, and fibrillation dynamics in beating vs. blebbistatin-uncoupled isolated rabbit hearts. Frontiers in Physiology. 11, 464 (2020).
- Kettlewell, S., Walker, N. L., Cobbe, S. M., Burton, F. L., Smith, G. L. The electrophysiological and mechanical effects of 2,3-butane-dione monoxime and cytochalasin-D in the Langendorff perfused rabbit heart. Experimental Physiology. 89 (2), 163-172 (2004).
- Képiró, M., et al. para-Nitroblebbistatin, the non-cytotoxic and photostable Myosin inhibitor. Angewandte Chemie International Edition. 53 (31), 8211-8215 (2014).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.