
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
במבחנה גידול דבורים בודד: כלי להערכת גורמי סיכון זחל
In This Article
Summary
תרסיסים פטריות על צמחים פורחים עשויה לחשוף הדבורים בודד כדי ריכוזים גבוהים של שאריות אבקה בעקיצות פטריות. באמצעות מבוסס מעבדה בניסויים המערבים במבחנה-דבורה גדלו הזחלים, מחקר זה בוחן את ההשפעות אינטראקטיבי של צריכת פטריות שטופלו אבקה נגזר צמחים המחשב המארח ופונדקאי.
Abstract
למרות דבורים בודד לספק שירותי האבקה מכריע עבור גידולי פרא ו מנוהלים, קבוצה זו מינים עתירי התעלמו במידה רבה במחקרים תקנה חומרי הדברה. הסיכון של חשיפה שאריות פטריות הוא עשוי להיות גבוה במיוחד אם הספריי מתרחשת על, או ליד צמחים פונדקאים בזמן הדבורים הן איסוף אבקה להקצות את קיניהם. עבור מינים של Osmia צורכים אבקה מתוך קבוצה נבחרת של צמחים (oligolecty), חוסר היכולת להשתמש זני צמחים פונדקאים שאינו יכול להגביר שלהם גורם הסיכון הקשורות פטריות רעילות. כתב יד זה מתאר פרוטוקולים המשמשים בהצלחה נמצאת מאחור oligolectic מייסון דבורים, Osmia ribifloris . שאתה לאטו, מביצה לשלב prepupal בתוך התא צלחות תרבות בתנאי מעבדה סטנדרטית. ה במבחנה-דבורים גדלו משמשות לאחר מכן לחקור את ההשפעות של מקור החשיפה ואבקה פטריות על כושר דבורה. בהתבסס על עיצוב העצרת 2 × 2 מלא חצה, הניסוי בוחן את ההשפעות העיקריות ואינטראקטיבי של מקור החשיפה ואבקה פטריות על כושר זחל, לכמת ביומסה prepupal, זחל התפתחותית, וזמן שאירים. היתרון העיקרי של שיטה זו הוא כי באמצעות במבחנה-דבורים גדלו מפחית את השתנות רקע טבעית ומאפשרת מניפולציה בו זמנית של מספר פרמטרים ניסיוני. הפרוטוקול המתואר מציג כלי רב-תכליתי עבור השערות בדיקות המערבים את הסוויטה של הגורמים המשפיעים על בריאות דבורה. עבור מאמצי השימור ייפגש עם הצלחה משמעותית, מתמשך, כאלה תובנות הגומלין המורכבים של גורמים פיזיולוגיים וסביבתיים נהיגה דבורה ירידות תוכיח כדי להיות קריטי.
Introduction
בהינתן תפקידם כמו הקבוצה השלטת של חרקים ומאביקים1, אובדן הכללית בקרב אוכלוסיות דבורה מהווה איום על ביטחון תזונתי ועל המערכת האקולוגית יציבות2,3,4,5,6 ,7. מגמות הפוחתת שתי האוכלוסיות דבורה מנוהל ופרוע יוחסו מספר גורמי סיכון משותף כולל פיצול בית גידול, המתעוררים טפילים פתוגנים, אובדן של מגוון גנטי, ואת המבוא של מין פולש3 ,4,7,8,9,10,11,12. בפרט, העלייה החדה בשימוש של חומרי הדברה, (למשל, neonicotinoids) ישירות נקשר השפעות מזיקות בין דבורים13,14,15. מספר מחקרים הראו כי הסינרגטיות בין neonicotinoids וקוטלי (עבי) ergosterol ביוסינטזה-עיכוב יכול להוביל תמותה גבוהה על פני מספר דבורה מינים16,17,18 , 19 , 20 , 21 , 22. בכל זאת, קוטלי פטריות, זמן נחשבת "דבורה-בטוח", להמשיך להיות ריססו על גידולים חקלאיים, בלום בלי הרבה לביקורת23. דבורים הרעיה תועדו באופן שגרתי יחזיר המון אבקה מזוהם עם פטריות שאריות24,25,26. צריכת פטריות-ladenpollen כזה יכול לגרום תמותה גבוהה בין דבורים זחל27,28,29,30וחבילת אפקטים תת קטלני בין דבורים למבוגרים16 , 31 , 32 , 33 , 34. מחקר שנערך לאחרונה עולה כי קוטלי פטריות עלולה לגרום הפסדים דבורה על ידי שינוי בקהילה חיידקים בתוך המאוחסנות בכוורת אבקה, ובכך לשבש את symbioses קריטי בין דבורים חיידקים בעקיצות אבקה35.
למרות דבורים בודד הם חיוניים עבור ההאבקה של מספר צמחים פרועה, החקלאית36,37,38, זו קבוצה מגוונת של המאביקים קיבלה הרבה פחות תשומת לב בהדברה ניטור מחקרים. הקן של אישה מבוגרת בודד מכיל 5-10 אטומים בתאי הרבייה, אחד מצויד עם מסה סופית של אסף maternally אבקה צוף, ואת ביצה בודדת39. לאחר הבקיעה, הזחלים לסמוך על הפרשה שהוקצה אבקה, microbiota בעקיצות אבקה המשויך כדי לקבל תזונה מספקת40,41. מכיוון שהם חסרים את היתרונות של אורח חיים חברתי, דבורים בודד ייתכן לפגיע יותר חשיפה לחומרי הדברה42. למשל, בעת לגירעונות דבורים חברתית בעקבות תרסיס צבע עשויים להיות מתוגמלים על קצת להרחיב על-ידי עובדי ומסתיים לאחרונה מתעוררים מיוחמת, מותו של נקבה יחידה בודד למבוגרים פעילות הרבייה כל43. כאלה הבדלי רגישות מדגישים את הצורך לשלב דבורה מגוונות taxa במחקרים ecotoxicological כדי להבטיח הגנה ראויה על דבורים מנוהל ופרוע כאחד. עם זאת, מלבד קומץ של מחקרים, חקירות ההשפעות של חשיפה פטריות בעיקר התמקדה דבורים חברתיות18,23,32,44,45 ,46,47,48,49.
דבורים הבודדים השייכים סוג Osmia (איור 1) היו בשימוש ברחבי העולם כמאביקים יעיל של מספר חשוב פירות, אגוזים יבולים39,50,51,53, 53. כפי עם אחרים המאביק מנוהל קבוצות24,54,55,56,57,58, דבורים Osmia למבוגרים הם באופן שגרתי חשוף קוטלי פטריות ריססו על היבולים בבלום44. הנקבה הבוגרת שיחור מזון על יבולים לאחרונה מותז עשויים לאסוף, במניה לחדריהם מיוחמת עם פטריות עמוסי אבקה, המהווה את הדיאטה בלעדית עבור הזחלים המתפתח מאוחר יותר. צריכת בהוראות אבקה מזוהמים לאחר מכן לחשוף את הזחלים להוסיף פטריות שאריות42. הסיכון של חשיפה עשוי להיות גבוה יותר בקרב מינים oligolectic מספוא רק על60,59,צמחים הקשורים קשר הדוק מארח מספר61. דבורים megachilid מסוימים, לדוגמה, נראה מעדיפים לחפש אבקה מורכבים באיכות נמוכה, כאמצעי להפחתת הטפילות62. עם זאת, במידה שבה קוטלי פטריות להשפיע על כושר זחל בין דבורים בודד oligolectic יש לא הייתה מדעית לכמת. מטרתו של מחקר זה היא לפתח פרוטוקול לבחון הראשי וגדלתי השפעות אינטראקטיבי של מקור החשיפה ואבקה פטריות על הכושר של במבחנה דבורים בודד. לחקור, ביצים של ribifloris או . שאתה לאטו (ס) ניתן להשיג מסחרית (טבלה של חומרים). אוכלוסייה זו אידיאלית בגלל חשיבותו של המאביק מקורית, וכן שלה ברורה עבור צוף-עשיר Mahonia aquifolium (אורגון ענבים) נמצא במרחק אזור53,63,64 (איור 2).

איור 1. תמונה ברזולוציה גבוהה של מבוגרים Osmia ribifloris. צילום האשראי ד ר ג'ים מקל, אנטומולוג, משרד החקלאות-ARS אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.

באיור 2. Phragmite קינון הקנים של Osmia ribifloris (ס) עם נקבה הקינון בחזית. מחיצות קאמרית התקעים והשקעים מסוף עבור הקנים בנויים כעיסה עלים. צילום האשראי מר קימבל קלארק, NativeBees.com אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
המטרה הראשונה של מחקר זה היא להעריך את ההשפעה של צריכת פטריות שטופלו אבקה על כושר זחל (נמדד במונחים של זמן פיתוח וביומסה prepupal). בזמן חשיפה propiconazole פטריות יישומית בדרך כלל מקושר תמותה מוגברת בין דבורים למבוגרים על פני מספר מינים 23,24,32,44,45, 54,55,56,57,58,65,66,67, השפעתה על דבורים הזחל הוא פחות ידוע. המטרה השנייה של מחקר זה היא להעריך את ההשפעות של רב-פונדקאי אבקה על כושר זחל. מחקרים קודמים מצביעים על כי הזחלים דבורים oligolectic מצליחים לפתח כאשר נאלץ לצרוך אבקה-פונדקאי68. תוצאות כאלה לייחס בווריאציות דבורה פיזיולוגיה69, אבקה ביוכימיה70ו את microbiome מועיל המשויך אבקה טבעית הוראות71. המטרה השלישית של מחקר זה היא להעריך את ההשפעות אינטראקטיבי של פטריות וטיפול תזונתיים אבקה על כושר זחל.
תכונות ביולוגיות רבות כולל גודל הגוף האמהי, הקצאת משאבים קצב, אסטרטגיה הרעיה, אבקה כמות72,73,74,75 ידועים להשפיע על כושר זחל בין דבורים בודד. גורמים אלה יכולים להציג את השתנות משמעותי בין קני הסוף, אשר מציב אתגר בפיתוח עיצובים מוטוריים בדירת בעת הערכת בריאות זחל. יתר על כן, בהתחשב בכך התפתחות הזחל מתרחשת בתוך קנים הקינון אטום, ההשפעות של השתנות כזה על הוא צאצא הינם קשה לדמיין כימות ללא שימוש בטכניקות קטלניים (איור 3). כדי להתגבר על האתגר הזה, כל השערות במסגרת מחקר זה נבחנים באמצעות רימות גדלו מחוץ הקנים הקינון שלהם. הנבחנים מייצג באופן מלא מצטלבים 2 × 2 העצרת set-ups, עם כל גורם בהיקף של 2 רמות; גורם 1: פטריות חשיפה (פטריות; אין פטריות); פקטור 2: אבקה מקור (אבקה המארח, Non-פונדקאי אבקה). דבורים מגודלים מהביצה ועד השלב prepupal בתוך התא multiwell סטרילי צלחות תרבות בתנאי מעבדה מבוקרת. אחד זה אינדיבידואלי מצויד היטב עם כמות מתוקננת של אבקה הוראה, ביצה אחת. לאחר הבקיעה, הזחל ניזון אבקה שהוקצה בתוך הבאר, השלמת פיתוח זחל, יוזם להתגלמות. מחקרים שנעשו בעבר הראו כי התמותה לא מוסברת הוא נמוך בין דבורים העלה בתוך סביבה זו rearing מלאכותי יותר נתקל פראי49,76. השימוש במבחנה-דבורים גדלו יש כמה יתרונות על פני השדה מבוססי מחקרים: 1) מצמצם את ההשפעות מבלבלים של השתנות טבעי וגורמים בלתי מבוקרת, הקשורות בדרך כלל לימודי המבוסס על שדה; 2) היא מאפשרת רמות מרובות של מניפולציה עבור factor(s) כל עניין להיבדק בו זמנית על פני קבוצות הטיפול; 3) מספר משכפל יכול להיות מוגדר מראש, גורמים ניסיוני עבור כל שכפול ניתן לטפל בנפרד; 4) תגובה זחל משתנים ניתן בקלות דמיינו להקליט ולצפות באופן עצמאי ללא מפריע הזחלים סמוכים; 5) הפרוטוקול יכול להיות שונה כדי להתאים עיצובים מוטוריים מורכבים יותר מעורבים גורמים רבים ומשתנים התגובה.

איור 3. תוכן בתוך פיה הקינון הטבעי של Osmia ribifloris (ס). (א) פיה גזור מציג צ'יימברס בודדים, הוראות אבקה, ואת מחיצות ואמצעי (B) טריים שנקטפו הוראות אבקה, והביצים המשויך (מסומן בעיגול שחור) מבט מקרוב. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Protocol
1. מכינים את Propiconazole פתרונות לניסויים חשיפה פטריות
- להכין 0.1 x פטריות פתרון על ידי המסת כרכים המתאים של propiconazole מסחרית שנרכשו 14.3% במים סטריליים היום של הניסוי. ודא כי הפיתרון היחיד של פטריות טריות משמש עבור כל הטיפולים.
- הוסף µL 23 של 0.1 x פתרון פטריות לגרם של אבקה הוראה כדי להשיג את הריכוז המרבי של propiconazole דיווח בעבר מאסף-דבורה אבקה24 (0.361 PPM או µg של המרכיב הפעיל g-1 של אבקה).
2. הקציר ביצים והוראות אבקה מארח בין קני הסוף Osmia
- באמצעות אזמל מעוקר, לנתח טרי מחובר קינון הקנים של Osmia, שמחלקים את זה לשני חלקים לאורך הקנה לחשוף את התאים הבודדים.
הערה: כל קן עשוי להכיל בין 8 עד 14 תאי ביצה אחת בתוך תא. - לבדוק את הקנים באופן חזותי כדי לזהות את התאים המכילים ביצים זכר בהתבסס על הנחיות שפורסמו בעבר77. השימוש מחט כפוף סטיריליים כדי להסיר את כל הוראה אבקה יחד עם הביצה המשויך הקנה הקינון, מקום בסביבה נקייה לשקול את הסירה.
- בעדינות להפריד את הביצה הפרשה בעזרת מברשת צבע נקי יפה רושמות את משקל טריים הפרשה אבקה, ביצה באמצעות איזון מעבדה סטנדרטיים. לחשב את המשקל הממוצע של הוראות אבקה זכר.
- בצע את השלבים הבאים עם השהיה המינימלי כדי להקטין את הסיכוי לנזק לביצה מחשיפה עודף חום והתייבשות.
3. מכינים מארח המפעל אבקה הוראות
- מבחינה ויזואלית לבדוק את האבקה מצמח פונדקאי maternally שנאספו נחפר מן התאים הקינון כדי להבטיח כי נוכח78אין טפילים. על מנת להפחית כל הטיה אימהי פוטנציאליים, לשלב בהוראות אבקה לתוך מסה יחידה בצלחת פטרי סטריליות ומערבבים היטב באמצעות מחט מעוקר.
- לחלק המסה המשולב החדש הוראות אבקה, המבטיח כי המשקל של כל הוראה משוקם שווה בקירוב להמשקל הממוצע של הוראה זכר שהוקצה באופן טבעי (כלומר ± SE, 0.35 ± 0.01 g, N = 42).
הערה: מכיוון Osmia sp. מקצה הוראות אבקה קטנים הצאצאים הזכריים, התוצאה משקולות הגוף התחתון של הזחלים הגברי בהשוואה לזו של נקבות77. כדי למנוע כל כזה הטיה הנובעת הבדלי מין ספציפי, השתמש רק ביצים הזכר בניסויים.
4. מכינים מצמח פונדקאי ללא אבקה הוראה
- Pulverize דבש מסחרית שנרכשו שנאספו דבורה אבקה אבקה באמצעות טחנת הכדור מעבדה סטנדרטיים.
- בהתבסס על הלחות של המארח שנאספו maternally אבקה הוראות (~ 20%), מימה את האבקה האבקה שימוש באמצעי אחסון המתאים של 40% לעקר סוכר פתרון79 , לערבב ביסודיות כדי ליצור עקביות דמוי בצק.
- לחלק אבקה בודדים ההמונים, כל אחת שוקלת כ זהה המשקל הממוצע של הוראה זכר שהוקצה באופן טבעי.
הערה: תכולת הלחות של אבקה מארח שנאספו maternally הוראות יכול להיות מוגדר ב מראש על-ידי השוואת המשקל טרי ויבש של אבקה הוראות 30 צ'יימברס זכר שנבחרו באקראי80. כדי להשיג את המשקל היבש, אבקה הוראות צריך להיות קפוא ומיובש, איזה שהוא לופילייזר (1.5 הרשות במשך 72 h).
5. מכינים צלחות תרבות תא Multiwell
- קו בארות בודדים של התרבות 48-ובכן סטרילי צלחת עם כוסות פח בלוק (5 × 9 ס מ). באמצעות מלקחיים סטרילי, פלייר בעדינות את השפה העליונה של הקפסולה כך זה עשוי להכיל הפרשה אבקה.
- למקם מסה יחידה של הוראה אבקה מארח או שאינו מארח בתוך בגביע פח בעזרת כלי סטרילי מבוסס על קבוצת הטיפול.
הערה: כדי למנוע זיהום צולב, השתמש צלחות נפרדות עבור טיפול וקבוצות שליטה.
6. מוסיפים קוטלי פטריות
- ליצור גומה שמוקם מרכזי בתוך המסה אבקה באמצעות מקל עץ סטרילי. השתמש מקל חדש עבור כל הוראה אבקה.
- להוסיף אמצעי אחסון המתאים של פתרון פטריות (לטיפול), או במים סטריליים (עבור פקדים) לתוך השפל. לצבוט את הפתיחה של הדיכאון באמצעות מלקחיים סטרילי כדי למזער את משטח המגע בין ונעביר / סטרילי הביצה והמים.
- ודא הסידור העצרת של הנבחנים מיושר עם זה מתואר בצורה הסמלית (איור 4).

באיור 4. ייצוג סכמטי של ההתקנה ניסיוני. הניסוי מייצגת חצה מלא 2 × 2 העצרת setups. גורם 1 מייצג פטריות חשיפה, מורכב מ- 2 רמות: (i) לא פטריות (N = 10), ו (ii) פטריות (N = 10). פקטור 2 מייצג מקור אבקה והיא מורכבת 2 רמות: אבקה (i) מארח (N = 8), ו (ii) Non-פונדקאי אבקה (N = 8). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
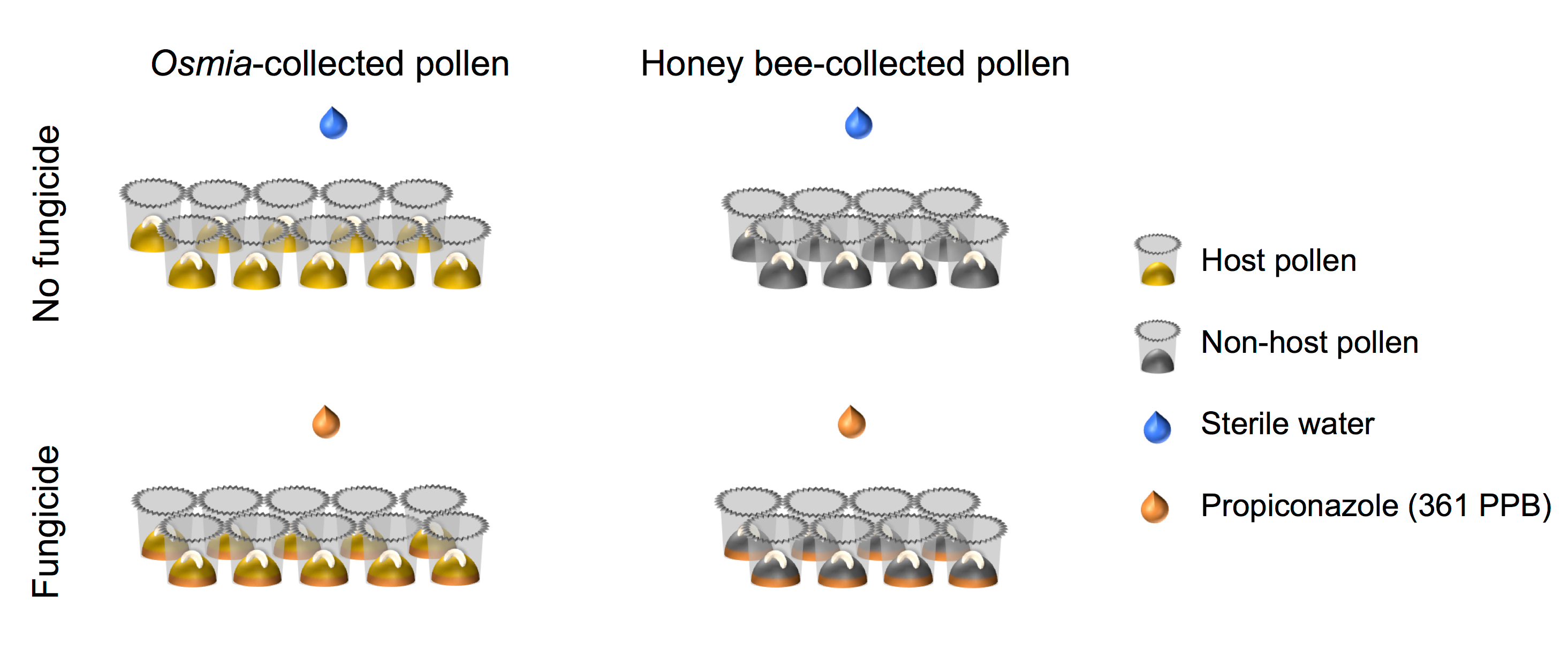{kind=link}
7. אחורי ולבחון את הזחלים
- במקום ביצה זכר שנבחרו באקראי על המשטח העליון של הוראה אבקה בעזרת מברשת צבע יפה נקי. פעם ביצים הונחו על כל הוראות, החלף את המכסה של לוח תרבות התא, כדי לאבטח אותו עם תיוג הקלטת על הפינות.
- למקם את הצלחות היטב על מגש נקי, לכסות את זה עם מטלית כהה כדי למנוע מגע עם אור ישיר. מקום טוב 6 לוח המכיל 30 מ ל מים סטריליים בתוך המגש כדי למנוע לייבוש. יוצאים הדגירה מגשים ללא הפרעה בתוך אינקובטור בטמפרטורת החדר.
- להתבונן צלחות טוב מדי יום תחת מיקרוסקופ ויבתר מבלי להסיר את המכסה של הלוחות היטב. להבטיח כי הזחלים בחיים על-ידי בדיקת לתנועה. אם אין תנועה מזוהה, למחוק בגביע פח המכילים את הזחלים מת ואת הפרשה אבקה הנותרים. אפשר כל הזחלים ששרדו לפתח הפרעה בתוך הלוחות היטב עד שהם מגיעים לשלב prepupal.
- הסר הזחל בגביע בדיל ברגע שהוא מגיע שלב prepupal41. שימוש במברשת כדי לנקות את כל צרכיהם המעטפת משי. בזהירות לחתוך דרך המעטפת משי באמצעות מיקרוסקופ ויבתר ולחלץ את prepupa עם מלקחיים גומי.
- להתמודד עם prepupa בעדינות כדי להבטיח כי הכלים לא לנקב את גוף רך. להקליט את המשקל טריים של prepupa (ביומסה prepupal) ואת פרק הזמן התפתחותית מביצה ועד השלב prepupal (זחל פעם התפתחותית).
הערה: כל זחל מתים צריכים להיות מושלך מיד כדי למנוע צמיחת חיידקים רצויה על גופה ועל מתן שאריות אבקה. זה מפחית את הסיכון של זיהום לזחלים בריא הנותרים.
Access restricted. Please log in or start a trial to view this content.
תוצאות
כושר זחל היה לכמת באמצעות שלושה מדדים (i) זחל התפתחותית זמן, ביומסה (ii) prepupal, שאירים (iii) אחוז. אנובה דו-כיווני נערך באמצעות חשיפה פטריות (שתי רמות: אין פטריות, פטריות) ומקור אבקה (שתי רמות: אבקה המארח, Non-פונדקאי אבקה) ' המשתנים תלויים, וכן זחל התפתחותית זמן המשתנה התלוי. ההשפעה ה...
Access restricted. Please log in or start a trial to view this content.
Discussion
גידול דבורים מחוץ הקנים הקינון הטבעי שלהם, בתנאי מעבדה, מאפשר בדיקה של השערות הנוגעים כושר זחל. כך גורמים לא מזוהה להמשיך לגרום לתמותה דבורה, לימודי הערכת סיכונים באמצעות במבחנה ניסויים יכול לעזור לזהות איומים פוטנציאליים ולהודיע לשיטות ניהול עבור קבוצת מינים עתירי למאביקים פראי
Access restricted. Please log in or start a trial to view this content.
Disclosures
המחברים אין לחשוף.
Acknowledgements
המחברים תודה קלארק קימבל, טים Krogh למתן הקנים קינון Osmia , מרדית נסביט, מולי בידוול לסיוע המעבדה, ד"ר קמרון קורי, קריסטל Guédot, טרי גריסוולד, מייקל Branstetter, שלושה אנונימיים בודקים עבור את הערותיהם שימושי זה לשפר את כתב היד. עבודה זו נתמכה על ידי שירות המחקר החקלאי-USDA הופקעו קרנות (מחקר מידע המערכת הנוכחית 3655-21220-001), מחלקת החקלאות של ויסקונסין, במסחר הגנת הצרכן (#197199), הקרן הלאומית למדע (תחת מענק מס DEB-1442148), DOE הגדול אגמים Bioenergy המרכז לחקר (DOE משרד המדע בער דה-FC02-07ER64494).
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| eggs of O. ribifloris sensu lato (s.l.) | Kaysville, Davis County, Utah, USA | ||
| Osmia reeds | Nativebees.com | NA | Freshly plugged reeds |
| Dissection set | VWR | 89259-964 | Sterilize before use |
| Long Nose Pliers | Husky | 1006 | |
| 6 well culture plates | VWR | 10062-892 | Sterile sealed |
| 48 well culture plates | VWR | 10062-898 | Sterile sealed |
| Petri dishes | VWR | 25373100 | Sterile sealed |
| Square Weighing Boats | VWR | 10770-448 | |
| Camel Hair Brush | Bioquip | 1153A | |
| Tin capsules | EA Consumables | D1021 | Sterilize before use |
| Sucrose | VWR | 470302-808 | |
| Propiconazole 14.3 | Quali-Ppro | 60207-90-1 | Propiconazole 14.3% |
| Honey bee pollen | Bee energised | 897098001244 | Untreated, natural, raw pollen |
| Microbalance | VWR | 10204-990 | |
| Pulverisette | LAB SYNERGY INC. | 30334913 | |
| Wooden sticks | VWR | 470146908 | Sterilize before use |
| Sealing tape | VWR | 89097-912 | |
| Microscope | VWR | 89403-384 | |
| Planting tray | VWR | 470150-632 | |
| Ethanol | VWR | BDH1158-4LP | |
| Centrifuge tube | VWR | 21008936 | |
| Microsyringe | Cole-Palmer | UX-07940-07 | |
| Rubber tweezer | Amazon | B0135HWPN4 | |
| Syringe needles | VWR | 89219-334 | |
| Freeze drier | Labcono | LFZ-1L | |
| Statistical software | SPSS | Version 21.0 |
References
- Klein, A. -M., et al. Importance of pollinators in changing landscapes for world crops. P Roy Soc Lond B Bio. 274 (1608), 303-313 (2007).
- Biesmeijer, J. C. J., et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science. 313 (5785), 351-354 (2006).
- Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O., Kunin, W. E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol Evol. 25 (6), 345-353 (2010).
- Cameron, S. A., et al. Patterns of widespread decline in North American bumble bees. P Natl Acad Sci USA. 108 (2), 662-667 (2011).
- Gallai, N., Salles, J. M., Settele, J., Vaissière, B. E. Economic valuation of the vunerability of world agriculture confronted with pollinator decline. Ecol Econ. 68 (3), 810-821 (2009).
- Fontaine, C., Dajoz, I., Meriguet, J., Loreau, M. Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. Plos Biol. 4 (1), 0129-0135 (2006).
- Kluser, S., Peduzzi, P. Global pollinator decline: a literature review. , UNEP/GRID Europe. (2007).
- Brown, M. J. F., Paxton, R. J. The conservation of bees: a global perspective. Apidologie. 40 (3), (2009).
- Lebuhn, G., et al. Detecting insect pollinator declines on regional and global scales. Conserv Biol. 27 (1), (2013).
- Vanengelsdorp, D., Meixner, M. D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J Invertebr Pathol. , Suppl 103. S80-S95 (2010).
- Pettis, J. S., Delaplane, K. S. Coordinated responses to honey bee decline in the USA. Apidologie. 41 (3), 256-263 (2010).
- Sandrock, C., Tanadini, L. G., Pettis, J. S., Biesmeijer, J. C., Potts, S. G., Neumann, P. Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agr Forest Entomol. 16 (2), (2014).
- Van der Sluijs, J. P., Simon-Delso, N., Goulson, D., Maxim, L., Bonmatin, J. M., Belzunces, L. P. Neonicotinoids, bee disorders and the sustainability of pollinator services. Curr Opin Env Sust. 5 (3), (2013).
- Goulson, D., Nicholls, E., Botías, C., Rotheray, E. L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 347 (6229), (2015).
- Johnson, R. M., Ellis, M. D., Mullin, C. A., Frazier, M. Pesticides and honey bee toxicity - USA. Apidologie. 41 (3), (2010).
- Iwasa, T., Motoyama, N., Ambrose, J. T., Roe, R. M. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Protection. 23 (5), 371-378 (2004).
- Glavan, G., Bozic, J. The synergy of xenobiotics in honey bee Apis mellifera: mechanisms and effects. Acta Biol. Slov. 56, 11-27 (2013).
- Biddinger, D. J., et al. Comparative toxicities and synergism of apple orchard pesticides to Apis mellifera (L.) and Osmia cornifrons (Radoszkowski). PLoS ONE. 8 (9), e72587(2013).
- Thompson, H. M., Fryday, S. L., Harkin, S., Milner, S. Potential impacts of synergism in honeybees (Apis mellifera) of exposure to neonicotinoids and sprayed fungicides in crops. Apidologie. 45 (5), 545-553 (2014).
- Jansen, J. -P., Lauvaux, S., Gruntowy, J., Denayer, J. Possible synergistic effects of fungicide-insecticide mixtures on beneficial arthropods. IOBC-WPRS Bulletin. 125, 28-35 (2017).
- Robinson, A., Hesketh, H., et al. Comparing bee species responses to chemical mixtures: Common response patterns? PLoS ONE. 12 (6), (2017).
- Sgolastra, F., Medrzycki, P., et al. Synergistic mortality between a neonicotinoid insecticide and an ergosterol-biosynthesis-inhibiting fungicide in three bee species. Pest Management Science. 73 (6), 1236-1243 (2017).
- Ladurner, E., Bosch, J., Kemp, W. P., Maini, S. Assessing delayed and acute toxicity of five formulated fungicides to Osmia lignaria and Apis mellifera. Apidologie. 36 (3), 449-460 (2005).
- Mullin, C. A., et al. High levels of miticides and agrochemicals in North American apiaries: implications for honey bee health. PloS one. 5 (3), e9754(2010).
- Pettis, J. S., Lichtenberg, E. M., Andree, M., Stitzinger, J., Rose, R., Vanengelsdorp, D. Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PloS one. 8 (7), e70182(2013).
- David, A., et al. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environ Int. 88, 169-178 (2016).
- Zhu, W., Schmehl, D. R., Mullin, C. A., Frazier, J. L. Four common pesticides, their mixtures and a formulation solvent in the hive environment have high oral toxicity to honey bee larvae. PloS one. 9 (1), e77547(2014).
- Simon-Delso, N., Martin, G. S., Bruneau, E., Minsart, L. A., Mouret, C., Hautier, L. Honeybee colony disorder in crop areas: The role of pesticides and viruses. PLoS ONE. 9 (7), (2014).
- Park, M. G., Blitzer, E. J., Gibbs, J., Losey, J. E., Danforth, B. N. Negative effects of pesticides on wild bee communities can be buffered by landscape context. P Roy Soc B-Biol Sci. 282 (1809), 20150299-20150299 (2015).
- Bernauer, O. M., Gaines-Day, H. R., Steffan, S. A. Colonies of bumble bees (Bombus impatiens) produce fewer workers, less bee biomass, and have smaller mother queens following fungicide exposure. Insects. 6 (2), 478-488 (2015).
- Williamson, S. M., Wright, G. A. Exposure to multiple cholinergic pesticides impairs olfactory learning and memory in honeybees. J Exp Biol. 216 (10), 1799-1807 (2013).
- Artz, D. R., Pitts-Singer, T. L. Effects of fungicide and adjuvant sprays on nesting behavior in two managed solitary bees, Osmia lignaria and Megachile rotundata. PLoS ONE. 10 (8), e0135688(2015).
- Pilling, E. D., Bromleychallenor, K. A. C., Walker, C. H., Jepson, P. C. Mechanism of synergism between the pyrethroid insecticide lambda-cyhalothrin and the imidazole fungicide prochloraz, in the honeybee (Apis mellifera L). Pestic Biochem Phys. 51 (1), 1-11 (1995).
- Johnson, R. M., Wen, Z., Schuler, M. A., Berenbaum, M. R. Mediation of pyrethroid insecticide toxicity to honey bees (Hymenoptera: Apidae) by cytochrome P450 monooxygenases. J. Econ. Entomol. 99 (4), 1046-1050 (2006).
- Steffan, S. A., Dharampal, P. S., Diaz-Garcia, L. A., Currie, C. R., Zalapa, J. E., Hittinger, C. T. Empirical, metagenomic, and computational techniques illuminate the mechanisms by which fungicides compromise bee health. JoVE. (128), e54631(2017).
- Batra, S. W. T. Solitary bees. Sci Am. 250 (2), 120-127 (1984).
- Linsley, E. G. The ecology of solitary bees. Hilgardia. 27 (19), 543-599 (1958).
- Garibaldi, L. A., et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science. 339 (6127), 1608-1611 (2013).
- Bosch, J., Kemp, W. P. How to manage the blue orchard bee. , Sustainable Agriculture Network. Washington, DC. (2001).
- Keller, A., Grimmer, G., Steffan-Dewenter, I. Diverse microbiota identified in whole intact nest chambers of the red mason bee Osmia bicornis (Linnaeus 1758). PLoS ONE. 8 (10), e78296(2013).
- Bosch, J., Kemp, W. P. Development and Emergence of the Orchard Pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environmental Entomology. 29 (1), 8-13 (2000).
- Brittain, C., Potts, S. G. The potential impacts of insecticides on the life-history traits of bees and the consequences for pollination. Basic and Applied Ecology. 12 (4), 321-331 (2011).
- Arena, M., Sgolastra, F. A meta-analysis comparing the sensitivity of bees to pesticides. Ecotoxicology. 23 (3), 324-334 (2014).
- Ladurner, E., Bosch, J., Kemp, W. P., Maini, S. Foraging and nesting behavior of Osmia lignaria (Hymenoptera: Megachilidae) in the presence of fungicides: cage studies. J Econ Entomol. 101 (3), 647-653 (2008).
- Huntzinger, A. C. I., James, R. R., Bosch, J., Kemp, W. P. Fungicide tests on adult alfalfa leafcutting bees (Hymenoptera: Megachilidae). J Econ Entomol. 101 (4), 1088-1094 (2008).
- Tsvetkov, N., et al. Chronic exposure to neonicotinoids reduces honey bee health near corn crops. Science. 356 (6345), 1395-1397 (2017).
- Mao, W., Schuler, M. A., Berenbaum, M. R. Disruption of quercetin metabolism by fungicide affects energy production in honey bees (Apis mellifera). P Natl Acad Sci. 114 (10), 2538-2543 (2017).
- Blacquière, T., Smagghe, G., Van Gestel, C. A. M., Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology. 21 (4), 973-992 (2012).
- Sgolastra, F., Tosi, S., Medrzycki, P., Porrini, C., Burgio, G. Toxicity of spirotetramat on solitary bee larvae, Osmia cornuta (Hymenoptera: Megachilidae), in laboratory conditions. Journal of Apicultural Science. 59 (2), 73-83 (2015).
- Mader, E., Spivak, M., Evans, E. Managing Alternative Pollinators. , Sustainable Agriculture Research and Education (SARE), US Dept. of Agriculture. (2010).
- Bosch, J., Kemp, W. P. Developing and establishing bee species as crop pollinators: the example of Osmia spp.(Hymenoptera: Megachilidae) and fruit trees. B Entomol Res. 92 (1), 3-16 (2002).
- Sampson, B. J., Rinehart, T. A., Kirker, G. T., Stringer, S. J., Werle, C. T. Phenotypic variation in fitness traits of a managed solitary bee, Osmia ribifloris (Hymenoptera: Megachilidae). J Econ Entomol. 108 (6), 2589-2598 (2015).
- Sampson, B. J., Cane, J. H., Kirker, G. T., Stringer, S. J., Spiers, J. M. Biology and management potential for three orchard bee species (Hymenoptera: Megachilidae): Osmia ribifloris Cockerell, O. lignaria (Say) and O.chalybea Smith with emphasis on the former. Acta Hort. 810, 549-555 (2009).
- Hladik, M. L., Vandever, M., Smalling, K. L. Exposure of native bees foraging in an agricultural landscape to current-use pesticides. Sci Total Environ. 542, 469-477 (2016).
- Long, E. Y., Krupke, C. H. Non-cultivated plants present a season-long route of pesticide exposure for honey bees. Nat Commun. 7, (2016).
- Krupke, C. H., Hunt, G. J., Eitzer, B. D., Andino, G., Given, K. Multiple routes of pesticide exposure for honey bees living near agricultural fields. PLoS ONE. 7 (1), e29268(2012).
- Stoner, K. A., Eitzer, B. D. Using a hazard quotient to evaluate pesticide residues detected in pollen trapped from honey bees (Apis mellifera) in Connecticut. PLoS ONE. 8 (10), e77550(2013).
- Sánchez-Bayo, F., Goulson, D., Pennacchio, F., Nazzi, F., Goka, K., Desneux, N. Are bee diseases linked to pesticides? - A brief review. Environ Int. 89, 7-11 (2016).
- Steffan-Dewenter, I., Klein, A. -M., Gaebele, V., Alfert, T., Tscharntke, T. Bee diversity and plant-pollinator interactions in fragmented landscapes. Specialization and generalization in plant-pollinator interactions. , 387-410 (2006).
- Kremen, C., Ricketts, T. Global perspectives on pollination disruptions. Conserv Biol. 14 (5), 1226-1228 (2000).
- Memmott, J., Waser, N. M., Price, M. V. Tolerance of pollination networks to species extinctions. P Roy Soc B-Biol Sci. 271 (1557), 2605-2611 (2004).
- Spear, D. M., Silverman, S., Forrest, J. R. K. Asteraceae pollen provisions protect Osmia mason bees (Hymenoptera: Megachilidae) from brood parasitism. The American Naturalist. 187 (6), 797-803 (2016).
- Rust, R. W. Biology of Osmia (Osmia) ribifloris Cockerell (Hymenoptera: Megachilidae). J Kansas Entomol Soc. 59, 89-94 (1986).
- Torchio, P. F. Osmia ribifloris, a native bee species developed as a commercially managed pollinator of highbush blueberry (Hymenoptera: Megachilidae). J Kansas Entomol Soc. 63 (633), 427-436 (1990).
- Sanchez-Bayo, F., Goka, K. Pesticide residues and bees - A risk assessment. PLoS ONE. 9 (4), e94482(2014).
- Kasiotis, K. M., Anagnostopoulos, C., Anastasiadou, P., Machera, K. Pesticide residues in honeybees, honey and bee pollen by LC-MS/MS screening: Reported death incidents in honeybees. Sci Total Environ. 485 (1), 633-642 (2014).
- Stanley, J., Sah, K., Jain, S. K., Bhatt, J. C., Sushil, S. N. Evaluation of pesticide toxicity at their field recommended doses to honeybees, Apis cerana and A. mellifera through laboratory, semi-field and field studies. Chemosphere. 119, 668-674 (2015).
- Praz, C. J., Müller, A., Dorn, S. Specialized bees fail to develop on non-host pollen: Do plants chemically protect their pollen? Ecology. 89 (3), 795-804 (2008).
- Sedivy, C., Müller, A., Dorn, S. Closely related pollen generalist bees differ in their ability to develop on the same pollen diet: Evidence for physiological adaptations to digest pollen. Funct Ecol. 25 (3), 718-725 (2011).
- Williams, N. M. Use of novel pollen species by specialist and generalist solitary bees (Hymenoptera: Megachilidae). Oecologia. 134, (2003).
- Graystock, P., Rehan, S. M., McFrederick, Q. S. Hunting for healthy microbiomes: determining the core microbiomes of Ceratina, Megalopta, and Apis bees and how they associate with microbes in bee collected pollen. Conserv Genet. 18 (3), 1-11 (2017).
- Bosch, J., Vicens, N. Relationship between body size, provisioning rate, longevity and reproductive success in females of the solitary bee Osmia cornuta. Behav Ecol Sociobiol. 60 (1), 26-33 (2006).
- Bosch, J., Vicens, N. Body size as an estimator of production costs in a solitary bee. Ecol Entomol. 27 (2), 129-137 (2002).
- Radmacher, S., Strohm, E. Factors affecting offspring body size in the solitary bee Osmia bicornis (Hymenoptera, Megachilidae). Apidologie. 41 (2), 169-177 (2010).
- Seidelmann, K. Open-cell parasitism shapes maternal investment patterns in the Red Mason bee Osmia rufa. Behav Ecol. 17 (5), (2006).
- Becker, M. C., Keller, A. Laboratory rearing of solitary bees and wasps. Insect Science. 23 (6), 918-923 (2016).
- Bosch, J. The nesting behaviour of the mason bee Osmia cornuta (Latr) with special reference to its pollinating potential (Hymenoptera, Megachilidae). Apidologie. 25, 84-93 (1994).
- Krunić, M., Stanisavljević, L., Pinzauti, M., Felicioli, A. The accompanying fauna of Osmia cornuta and Osmia rufa and effective measures of protection. B Insectol. 58 (2), 141-152 (2005).
- Elliott, S. E., Irwin, R. E., Adler, L. S., Williams, N. M. The nectar alkaloid, gelsemine, does not affect offspring performance of a native solitary bee, Osmia lignaria (Megachilidae). Ecol Entomol. 33 (2), 298-304 (2008).
- Toby Mordkoff, J. The Assumption(s) of Normality. , Available from: http://www2.psychology.uiowa.edu/faculty/mordkoff/GradStats/part 1/I.07 normal.pdf (2016).
- Hendriksma, H. P., Härtel, S., Steffan-Dewenter, I. Honey bee risk assessment: New approaches for in vitro larvae rearing and data analyses. Methods Ecol and Evol. 2 (5), 509-517 (2011).
- Aupinel, P., et al. Improvement of artificial feeding in a standard in vitro method for rearing Apis mellifera larvae. B Insectol. 58 (2), 107-111 (2005).
- Beekman, M., Ratnieks, F. L. W. Long-range foraging by the honey-bee, Apis mellifera L. Funct Ecol. 14 (4), 490-496 (2000).
- Gathmann, A., Tscharntke, T. Foraging ranges of solitary bees. J Anim Ecol. 71 (5), 757-764 (2002).
- Greenleaf, S. S., Williams, N. M., Winfree, R., Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia. 153 (3), 589-596 (2007).
- Bee Pollen Supplement - Bee Rescued. , Available from: https://beerescued.com/product/bee-rescued-bee-pollen-supplement/ (2018).
- Holly-Leaf Oregon-Grape (Mahonia aquifolium) Species Details and Allergy Info, Teton county, Wyoming. , Available from: http://www.pollenlibrary.com/Local/Specie/Mahonia+aquifolium/in/Teton County/WY/ (2018).
- Cane, J. H., Griswold, T., Parker, F. D. Substrates and Materials Used for Nesting by North American Osmia Bees (Hymenoptera: Apiformes: Megachilidae). Annals of the Entomological Society of America. 100 (3), 350-358 (2007).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved